JOURNAL OF VIROLOGY, May1990,p. 2110-2116 0022-538X/90/052110-07$02.00/0
Copyright © 1990,AmericanSocietyforMicrobiology
Structure and Function of the Epstein-Barr
Virus BZLF1
Protein
GRAHAMPACKHAM, ANDROULLA ECONOMOU, CLIONAM. ROONEY,DAVID T. ROWE,
AND PAUL J. FARRELL*
Ludwig Institute for Cancer Research, St. Mary's HospitalMedicalSchool, Norfolk Place, London W2IPG, England Received 20 November 1989/Accepted 14 January 1990
FiveDNA-binding sites for the Epstein-Barr virus BZLF1 protein have been identified withinthreeof the early viral promoters, and four of thesebindingsites containaconsensusAP-1 site. Thepartof theBZLF1 protein required for sequence-specific DNA bindingtooneof these AP-1-like siteswas identifiedby deletion mapping. Site-directedmutagenesis of this DNA target suggests that BZLF1mayworkpartlybyovercoming
acellularrepressorof viral transcription.
Epstein-Barr virus (EBV) isahumanherpesvirusthatcan
follow either a latent (i.e., nonproductive) or aproductive life cycle. When EBV infectsan appropriate B lymphocyte invitro,thelymphocyte becomesimmortalizedandalatent infection is established (reviewed in reference 13). The switch tothe virus-productive infection can be induced in
vitro by treating the latently infected lymphocytes with a variety of chemical or biological inducers (e.g.,
12-O-tet-radecanoylphorbol-13-acetate, transforming growth factor
,B,
orantiimmunoglobulin)
orby superinfection
with certainrearranged, defective EBV genomes (e.g., het virus). In
common with other herpesviruses, the productive-cycle
geneexpression of EBV is organizedas acascade ofgroups
ofgenes (reviewed in reference 6). Present evidence indi-catesthat expression of the BZLF1geneof EBV(also called
EB1 and ZEBRA) is the first step in the productive-cycle cascade (2, 5, 20, 21). Transfection of a transcriptionally active BZLF1constructissufficienttoinduce theproductive cycle (4, 5, 18). The BZLF1 and BRLF1 proteinsaregene regulatory molecules, both of whichcanactivate the BSLF2 +BMLF1 gene (also called EB2), and then various
combi-nations of these three gene products appear to activate further stages of the cascade (3, 8, 10-12, 15-18, 22). We showedpreviously that BZLF1 isasequence-specific
DNA-binding protein thatcanbindtoaDNAsequencewithinthe promoter region of the BSLF2+BMLF1 gene (7). This binding site contains a consensus recognition sequence for
the AP-1 transcription factor family (TGAGTCA; EBV position 84429). BZLF1 has localized protein sequence homologytoc-fos and c-jun, which are components of the AP-1factor, particularly inaregion called the "basic motif," which is thoughtto be thepartof the protein that interacts with DNA. We have shown elsewhere (7, 18) that approxi-mately theC-terminal half of BZLF1 (from the SmaI siteto the Cterminus) is sufficienttobind specificallytothetarget but will not activate transcription. N-terminal sequences within the BZLF1 protein aretherefore required forthefull activity of the protein butnotfor DNA bindingper se.Inthis
paper we have mapped five DNA-binding sites for the BZLF1 protein within the EBV genome; not all of these contain a consensus AP-1 site. We also delineate more precisely those parts ofBZLF1 thatare required for DNA
binding at one of the sites and for transactivation ofthat EBV promoter. Site-directed mutagenesis of this target
*Correspondingauthor.
indicates that BZLF1 may workpartly by counteracting a
cellular factor thatrepressesviraltranscription. MATERIALS ANDMETHODS
Construction of plasmids. The starting point for all the BZLF1 deletionmutantswastheplasmid SP64-BZLF1(18),
which isan870-base-pair BamHI-EcoRI cDNA for BZLF1
cloned between the BamHI and EcoRI sites of SP64.
SmaI-C.The N-terminalSmaIfragmentwasdeletedfrom
SP64-BZLF1 and replaced with adouble-stranded (ds) oli-gonucleotideofsequence CATCGATG, which suppliesthe
initiator methionine codon.
N-Hincd. SP64-BZLF1 was cut with HincIl, and a ds
oligonucleotide of sequence TAGAATTCTA was added. Afterdigestion with EcoRI, the DNA wascircularized and cloned.
N-PstI. SP64-BZLF1 was cutwithPstI, and the PstI ends wereligatedto adsadaptor ofsequence
GCGAATTCTTACTGCA CGCTTAAGAATG
After cleavage with EcoRI and BamHI, the resulting BamHI-EcoRI fragment containing the appropriate part of BZLF1 was cloned between theBamHI and EcoRI sitesof
SP64.
N-StyI. SP64-BZLF1wascutwithStyI, and the endswere
made blunt withmung bean nuclease. A dsoligonucleotide
ofsequence TAGAATTCTAwasligatedtothe ends. After digestion withEcoRI,theplasmid wascircularized.
HindIll-delta. SP64-BZLF1 was cut with HindIII, giving three pieces. The N-terminal HindIII fragmentwas cloned into the large vector/C-terminal piece, omitting the small centralHindIIIfragment.
BsmI-C. SP64-Sma-Cwasdigested with EcoRI and BsmI. An adaptorconsisting of the ds oligonucleotide
GATCCAAGATGTGCG GTTCTACAC
wasusedtojoin theBsmIendtothe BamHI site in thelarge BamHI-EcoRI fragment of SP64.
BsmI-HincII. SP64-BsmI-C was digested with PstI. The
smallerfragmentwascloned into thelarge PstI fragment of SP64-N-HincII.
The aboveconstructsin SP64werefor in vitro transcrip-tion and translation. To assay the activity of the BZLF1
deletion mutants by transfection, the sequences derived from BZLF1 in the SP64 constructs were subcloned as 2110
Vol.64,No. 5
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE AND FUNCTION OF EBV BZLF1 PROTEIN 2111
XbaI-EcoRI fragments into the 3.1W plasmid, as for 3.1W-BZLF1 in (18). This results in the EBV BamHI W latent
cycle promoterdriving expression ofthe BZLF1 genes. The S-CAT plasmid, the BglII derivative of pSVOCAT, and CMV-BZLF1 have been described elsewhere (18).
CMVLTR-BRLF1 contained EBV sequences 105415 to 103081 (BglII toHindIII) cloned between a simian virus 40
polyA site andastrongpromotercomprisingahuman T-cell leukemia/lymphoma virus 1 long terminal repeat and the
cytomegalovirus immediate early enhancer.
In vitro transcription and translation of BZLF1 mutants. TheSP64-BZLF1 constructs werelinearizedwithEcoRI and
transcribed with SP6 polymerase at 40°C for 45 min.
Tran-scriptionreactionscontained2.5 ,ugofDNA;0.5 mM eachof
ATP, GTP, CTP, and UTP; 100 ILM 7MeGpppG; 10 mM
dithiothreitol; 40 mM Tris hydrochloride (pH 7.5); 6 mM
MgCl2; 2 mM spermidine hydrochloride; 62.5 U ofRNasin (Promega);and 25 Uof SP6polymerase inavolumeof50
RI.
Afterthe transcription, the templateDNA wasremovedby digestionwith DNase I(Pharmacia; fastprotein liquid
chro-matographygrade, 2.5 U)at37°C for15 min. The RNA was
phenol-chloroform extracted, ethanol precipitated, and
dis-solved in 20
RI
of water. Translation was in themRNA-dependent reticulocyte lysate (Amersham) with 1.5 ,ul of
RNA in a totalvolume of15 ,ulat 30°C for 35 minorin the wheat germ system(Amersham)with 1 ,ulofRNA inatotal
volume of13
pul
at25°Cfor60min. Asamplewasdiluted in sodiumdodecyl sulfategelsample bufferandanalyzedon a20%polyacrylamide gel. The remaining translation product
wasfrozenat -70°C forDNA-bindingassays.
Site-directed mutagenesis of the promoter for BSLF2+ BMLF1. The 257-CAT, AE4-CAT, and AE4K-CAT plas-mids were prepared, starting with an M13 clone (257.RIF) from the DNA-sequencing program. This contained EBV sequences 84325 to 84767 (1) in M13mp8 at the SmaI
site,
oriented with the 84325 end close totheBamHI siteof the M13
polylinker.
Position84325 is thetranscription
initiation nucleotide ofthe spliced RNA encoding BSLF2+BMLF1,and 257.RIFincludesthe promoter
region
forthisRNAfrom +1 to -442 relative to the transcription start. Asynthetic
oligonucleotide of sequence GCGAAGCACTCTCGAGTG AAGGTGAC wasusedfor site-directed
mutagenesis
(14) of257.RIF, resulting in the replacement of the GACTCA at EBV 84430 with CTCGAG (an XhoI site). This gave M13AE4. ds DNA from 257.RIF and M13AE4 was
pre-pared, and theEBV sequencescouldbeexcised
by using
theflanking BamHI and EcoRI sites in the
polylinker.
The EcoRI site was filled in with Klenow DNApolymerase;
a BamHIlinkerwasadded,and theresulting BamHIfragment
wasclonedinto theBglIIsite ofpSVOCAT,giving257-CAT and AE4-CAT, resulting in either the
wild-type
or mutant promoterdrivingthe CATgene. AE4-CATwasthen cutat theunique XhoIsite,filled in with Klenowpolymerase,and recircularized with DNA ligase, giving AE4K-CAT. The sequencesof257.RIF, AE4, and AE4K around thepoint
ofmutagenesis werecheckedby DNA
sequencing.
Footprinting. DNase I footprinting used the same prepa-ration ofBZLF1 fusion protein
(pREX-SmaA),
which is a fusion ofproteinAtotheSmaI-CBZLF1 sequences,as was described elsewhere (7, 18). Reaction conditions forfoot-printingwere asdescribedpreviously (7).
Gel retardation.
32P-labeled
DNA substrates were either the 261-base-pair 374RIF fragment from the promoter for BSLF2+BMLF1 (7) ordsoligonucleotides
overlapping
its consensus AP-1site. The wild-typeoligonucleotide
292/293 had ds sequenceGAAGCACTGACTCATGAAG,
and theAE4
oligonucleotide
was ds GCGAAGCACTCTCGAGTG AAGGTGAC.Binding
wasperformed
at roomtemperature as describedpreviously (7).
The fusionprotein
orin vitro translationproduct
(usually
1to3pI)
wasdiluted in 18pul
of100 mMKCl-20mM HEPES
(N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid)
(pH 7.9)-10%
glycerol-0.2
mMEDTA-4 mM
dithiothreitol,
and tothis,
4 ,ulofpremixed
DNA
[i.e.,
poly(dIC),
32P-labeled
targetDNA,
and anycompetitor
oligonucleotide]
was added. This resulted in afinal concentration of 100
pug
ofpoly(dIC)
per ml. Concen-trations of32P-labeled
DNAsandcompetitors
aregiven
in thefigure
legends.
Afterincubationatroomtemperaturefor20to30min,3
pu1
ofdye
(xylene
cyanol
FFandbromophenol
blue in thesame
buffer)
wasadded and the wholesample
wasloadedon toeithera1%agarose
gel
or a4%polyacrylamide
gel.
Gelswere runeither in 0.5x TBEorbuffer G(380
mMglycine,
50 mMTris,
5 mM EDTA[pH
8.5]).
The shiftedcomplexes
were detectedby
autoradiography
of the driedagarose
gel
orthewetpolyacrylamide gel.
Transfection
experiments. (i) Assay
of BZLF1 deletion mutants. 3.1W-BZLF1 orthe3.1W-BZLF1 deletion mutant DNAs wereelectroporated
with S-CATin BL41/CL16 cells aspreviously
described(18).
Cells were harvested after 1day,
andCATactivity
wasdetermined(18).
(ii) Assay
of site-directedmutantsin AP-1 siteupstreamof BSLF2+BMLF1.S-CAT, 257-CAT,
AE4-CAT,
andAE4K-CATwere
electroporated
withorwithoutCMV-BZLF1,
orCMVLTR-BRLF1,
into BL41cellsaspreviously
described(18).
Cells were harvested after 2days,
and CATactivity
wasassayed (18).
RESULTS
Deletion
mapping
the part of BZLF1required
for DNAbinding.
A cDNAencoding
the BZLF1protein
was clonedpreviously
between theBamHIand EcoRIsitesofSP64,
and this clone could be transcribed and translated in vitro toproduce
authentic BZLF1protein
(18).
Various deletion mutants wereprepared by
restrictiondigestion,
using
syn-thetic
oligonucleotides
torepair
the resected ends(Fig.
1).
By
this means, aninitiatormethionineresiduewassupplied
totheN-terminal deletionsandatranslationterminatorwasrestored tothe C-terminal deletions. All thedeletionswere
precise
truncations ofthe BZLF1 sequence, and no extra-neous aminoacids were introduced(except
forthe initiator methionine residues in the N-terminaldeletions).
All ofthedeletions ofBZLF1 weretranscribedwith SP6
polymerase,
and the
resulting
RNAwastranslatedineitherthereticulo-cyte
lysate
or wheat germ translationsystem,
using
[35S]cysteine
astheradioactive label.Analysis
of thetrans-lation
products
on sodiumdodecyl
sulfategel
electrophore-sisshowed that allofthemutantsgave
proteins
ofappropri-ate relativesizes
(Fig.
2; BsmI-HincII
isnotshown).
Itwasnecessary to use the wheat germ system to demonstrate translation of the smaller
proteins
because thehigh
concen-tration of
globin
in thereticulocyte
lysate
distorted the sodiumdodecyl
sulfategel
in that size range. The invitro-translated
proteins
were thenused ingel
retardationassays to test theirability
to bindspecifically
to a261-base-pair
32P-labeled
fragment
of EBV DNAspanning
thepromoter
region
of BSLF2+BMLF1 andcontaining
the consensusAP-1 site. This is the same DNA
fragment
to which wepreviously
showedbinding
of BZLF1 fusionproteins
and invitro-translatedBZLF1
(7).
Thefull-length
BZLF1,
SmaI-C,
BsmI-C,
and N-HincIImutantsallshifted thetarget,
butthe N-PstI mutant did not(Fig.
3).
Because ofits smallsize,
VOL.64,1990on November 10, 2019 by guest
http://jvi.asm.org/
2112 PACKHAM ET AL.
0 200 400 600 800bp
I I I
BamHI Hindlil Hindill Smal Styl Bsmi
BZLF1
BZLF1 N-Hincil
BZLF1 Hindlil defta
BZLF1 N-Psti
BZLF1 Smal-C
BZLF1 N-Styl
BZLF1 BsmI-C
BZLF1 Bsml-Hincil
I I
PstI Hincil EcoRt
boundaries
MMD... LNF
- MMD ... LDV
..FVQAYA..
MMD ...LLQ
MGA ...LNF
MMD ... GAN
MCD... LNF
- MCD... LDV
RegionofhomologywithfosrjunDNAbindingdomain
Non-codingsequences
Codingsequences
Bsml Pstl Hincll
v
+
~~~+
v+
BZLF1 SLEECDSELEIKRYKNRVASRKCRAKFKQLLQHYREVAAAKSSENDRLRLLLKQMCPSLDVD
v <--baSiC Otif --> L v L L L L
C-FOS LSPEEEEKRRIRRERNKMAAAKCRNRRRELTDTLQAETDQLEDEKSALQTEIANLLKEKEKL
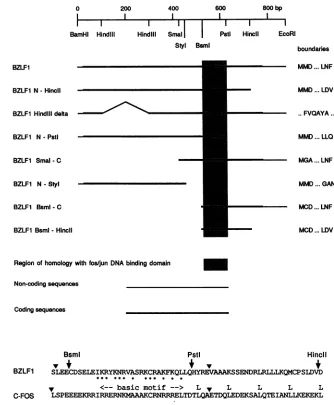
FIG. 1. BZLF1 deletion mutants. The structures of the BZLF1 deletion mutants are shown beneath a restriction map of the EBV sequences inthe BZLF1 cDNA(SP64-BZLF1). The amino acidsatthe relevant boundaries of the deletionsareindicated,andthe amino acid sequenceof theBsmI-HincIIportion that isrequired forDNAbinding is shown below. This sequence iscomparedwith part ofc-fosto illustrate the basic motif, the leucine zipper region (L) of c-fos, and the boundaries(V)ofexon2ofBZLF1andexon3ofc-fos.Conserved residues within the basic motif are marked with dots (R-K substitutions are consideredconserved).
BsmI-HincII was tested by using an oligonucleotide as the target(seebelow).
The shifts represent site-specific binding because assays were performed in the presence of excess poly(dIC) and
because the binding was abolished by an excess ofa ds
oligonucleotide containing the AP-1 site but not by an unrelated dsoligonucleotide. The results indicate that pro-tein sequences essential for binding lie between the BsmI and HincII sites. This region of 58 amino acids includes the
basic motifand, in c-jun and c-fos, the leucine zipper (the
leucinezipper is not apparent in BZLF1). Deletion from the
C terminustothe PstIsite removes the region equivalent to the leucine zipper and abolishes binding. Although the
BsmI-C mutant retains DNA binding, the protein-DNA
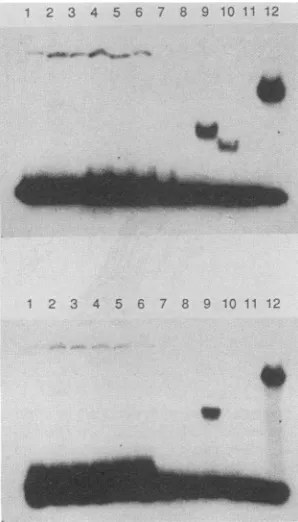
complex is less stable to salt than, for example, SmaI-C. This isillustrated in Fig.4, wheregelretardation assays of a
ds oligonucleotide covering the AP-1 site in the BSLF2+ BMLF1 promoter were performed in either 0.5x TBE or
bufferG(highersalt). Although the complex is seen in both
buffer systems with SmaI-C (lane 9), the BsmI-C complex
(lane 10) is only stable in the lower-salt buffer. A similar
phenomenonwasobservedby comparing full-lengthBZLF1 with N-HincII (data not shown). The BsmI-HincII mutant (lane 11) did notgive a significant retardation complex in either buffer system(an extremely weaksignalwas seenon theoriginal autoradiograph).
BZLF1 mutants that retain transactivation retain DNA binding. The BZLF1 cDNA and the deletion mutants HindIIIdelta, N-HincII, N-StyI, SmaI-C, andN-PstI were recloned downstreamofastrongEBV promoter(thelatent promoter in BamHI W) so as topermit expression of the BZLF1 protein. These constructs were
electroporated
intoBL41/CL16cells (whichcontain EBV) withaplasmid
con-tainingthe promoter for BSLF2+BMLF1 fusedtothe CAT gene. This promoter is dependent on transactivation from BZLF1 or BRLF1. The BZLF1 deletion mutant N-HincII was aseffective asfull-lengthBZLF1intransactivatingthis promoter, but N-PstI, N-StyI, and SmaI-C were inactive J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.136.470.71.476.2]STRUCTURE AND FUNCTION OF EBV BZLF1 PROTEIN 2113
(.3
= _
C.)
-O
=(cr
C)-C =Cf> < >2Y
z
I C)
Ns 9Is 0 E o
m z = Z C } co z
- 46
-30
-21
C ,;
... _ ._ __. _ ,._ " ,J::LC.sr^v9C0s
... ..
.; j':''
...:
'.
*' ':
* "e
>,.ji;t
{.i'.
=
_w w*
. _
_tj #4Sl _wP
-14
[image:4.612.111.252.78.331.2]-7
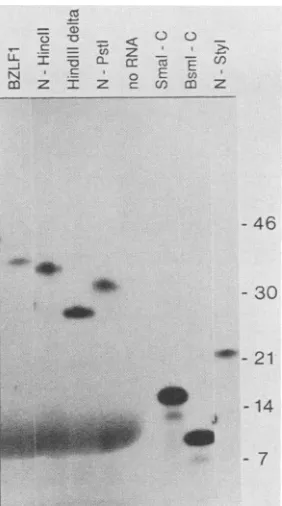
FIG. 2. In vitro translation of BZLF1 deletionmutants.DNAfor the various BZLF1 deletion clones in SP64 was linearized with EcoRIandtranscribed with the SP6 polymerase, and the resulting RNA was translated in the mRNA-dependent reticulocyte lysate (lanes BZLF1, N-HincII, HindIII delta, and N-PstI) orthe wheat
germ system (no RNA, SmaI-C, BsmI-C, and N-StyI), using
[35S]cysteineasthe radioactivelabel. The translation productswere
detectedbysodium dodecyl sulfate gel electrophoresis and fluorog-raphy. The positions to which radioactive protein size markers migratedareshownonthe right in kilodaltons.
(Fig. 5). These results are consistent with a model of the BZLF1protein in which sequence-specific DNA binding is mediatedby amino acids between the BsmI andHincII sites butsequences N terminal of the BsmI sitearealso required
fortransactivation. The deletionof amino acids between the HindIIIsitesresults only inapartial loss of the transactiva-tionfunctionofthe N-terminaldomain (Fig. 5, lane 2).
BZLF1binds specificallytoDNAsequencesupstreamof the promotersforBZLF1 andBRLF1.Since BZLF1 is thoughtto stimulate expression of BZLF1, BRLF1, and BSLF2+ BMLF1,wetested theability ofa BZLF1fusion proteinto footprintsequencesaround theBZLF1and BRLF1
promot-ers. Inthese experiments, the SmaI-C fusionto protein A
wasused, sincewehadpreviously shown this footprintedto the consensus AP-1 site in the promoter for BSLF2+ BMLF1 (7). The fusion protein was titrated into the foot-printing reactions, and this gave some measure of the
relativeaffinity ofbindingatdifferent sites. Two clear sites werereproducibly found upstream of the BRLF1promoter (probes RBR.511 and 7.FAL, Fig. 6). Some other sites of lowaffinity werealso observed;thesegave variable results
in footprinting. Upstream of the BZLF1 promoter (probe 48.BAZ), one region was found to footprint but this was unusuallywidecompared withthe other footprints, and the titration (lanes 2 through 4) indicates two binding sites of different affinitiesadjacent toeach other. The sequences of
allthe sitesaresummarizedinFig.7andarecomparedwith the site (M) previously mapped upstream of the BSLF2+ BSLF1promoter. All the sitesexceptfor thehigher-affinity
FIG. 3. Gel retardationassayforBZLF1 deletion mutants bind-ingto the promoter for BSLF2+BMLF1. The 32P-labeled probe from374.RIF (1 ngper assay,final concentration3 nM) wasmixed with variousBZLF1deletion mutants made by in vitro translation in the reticulocytelysate or wheat germ system (see Materials and Methods). Allreactions contained100
jig
ofpoly(dIC)per ml. Lanes 0, No oligonucleotide competitor; lanes 1, cognate 292/293 ds oligonucleotide GAAGCACTGACTCATGAAG (0.5 ,uM); lanes 2, anunrelated ds oligonucleotide sequence GGGTACCC (0.5JIM).
Electrophoresiswason a1%agarosegelin0.5x TBE.
site upstreamofBZLF1 arecentered on a close match to the consensusAP-1site. In thepossible alignment shown in Fig. 7, it can beseen that someother nucleotides also match in theflankingsequences thatarecoveredby the footprint.The
high-affinity site upstream of BZLF1 does not match the AP-1 consensus. A possible alternative consensus which matches all of the sites is indicated.
Site-directed mutagenesis of theconsensus AP-1 site in the promoterfor BSLF2+BMLF1. To assess thesignificanceof theconsensus AP-1 siteat 84429 in the promoterregionof
BSLF2+BMLF1, six nucleotides of the AP-1 site were
replaced by unrelated nucleotides without changing the
surrounding sequence. The site was changed from TGAC TCA to TCTCGAG. Analogous CAT fusions ofthe wild-type promoterand themutant(called AE4)wereprepared.A further mutant (AE4K) was made by cutting the XhoI site
(CTCGAG) introduced to AE4, repairing the ends with Klenow polymerase, and religating. This resulted in the inclusion of fourextranucleotides; therefore,in this mutant, the sequencewaschanged from TGAGTCAtoTCTCGATC GAG. These constructs were transfected into BL41 cells with or without CMV-BZLF1 to assay their sensitivity to
transactivation. Our principal objective here was to study
the effectofthesemutationson BZLF1transactivation,but
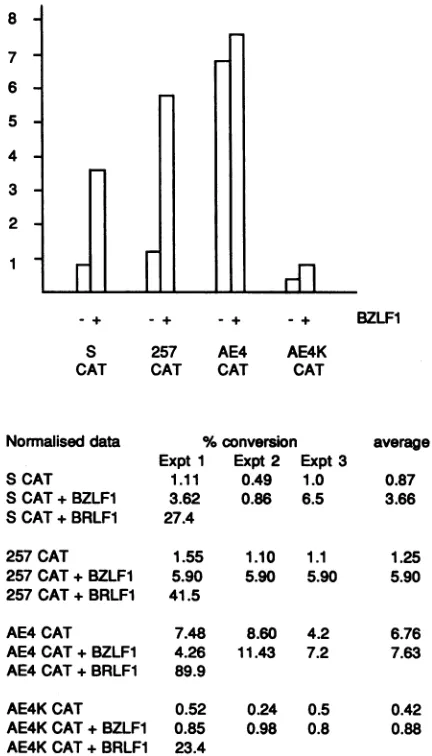
for illustrationandcomparison, the greatereffect of BRLF1 onthis promoter is shown inoneexperiment(Fig. 8).BZLF1 activated CATexpressionfrom thewild-typepromoter,but no effectwas seen on the AE4 construct because the AE4 promoterwas alreadyactive evenwithout BZLF1
(Fig.
8).The AE4K promoter was inactive with or without BZLF1
(but could still be activated by BRLF1). These results suggest that BZLF1isactingonthis target in BL41cells
by
counteractingacellular repressorthatrecognizessequences
involving the consensus AP-1 site. When the consensus AP-1site isremoved, theBZLF1 nolongerhasaneffect in
VOL.64, 1990
40
-, 4"..'W j
40
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.354.522.81.274.2]2114 PACKHAM ET AL.
1 2 3 4 5 6 7 8 9 10 11 12
S
A 7FAL 48BAZ 1234512345
tiEIf lA
{B..U ..
-rn
.-*abI a
aW
1 2 3 4 5 6 7 8 9 10 11 12
S V.
B RBR511 Ml 2345M
be4 .-529
_-406 m -406
1111
*
~~~~~~~-
311 |**-1.g- 240 .11:1 w-240
-219
i-182 * -0
*-162 . -182
*-i49 e *-162
R t , !r-~149
-125
*-112 -1251
92~ 7.. £ -92 * > 9
78 4-92
-, - 69
.. -6
- -_ _ . -69 1*Zi*ft
FIG. 4. Salt sensitivity ofgeJretardation complexes. Gel retar-dation assays were analyzed on polyacrylamide gels in0.5x TBE (upper panel) or buffer G (lowerpanel). Lanes 1 through 6, AE4 ds oligonucleotide (0.4 ,ug/ml)as acontrol; lanes 7 through 12,292/293 dsoligonucleotide (0.4 ,ug/ml). Lanes 1 and 7, No added protein; lanes 2 through 5 and 8 through 12, 2 ,u1 ofreticulocyte lysate translations of: lanes 2 and 8, no RNA; lanes 3 and 9,SmaI-C;lanes 4and 10,BsmI-C; lanes 5 and 11,BsmI-HincII.Lanes 6and 12, 1pI (0.5 pug)ofpREX-SmaA&fusion protein. Allreactions contained 100
pig
of poly(dIC) per ml.this assay system. An important promoter element must, however, surround or overlap this region because
introduc-ing anextrafour nucleotides in AE4K destroys this
basal-level promoter activity. The pREX-SmaA BZLF1 fusion
9
4lb
...
4-36
I.,afff *
MD .i ifs
_^ ^N
4b:JI.
I&a
&.
-36
FIG. 6. Footprinting of BZLF1 SmaI-C (pREX-SmaA) fusion proteintopromoterregionsofBZLF1and BRLF1. 32P-end-labeled DNAswereprepared forfootprinting as describedpreviously (7) by usinga32P-end-labeledprimertocopyasecond strandonM13 ss DNA. Theprimerwasthe 17-base sequencing primer; the clones usedwere (A) 48BAZ(BZLF1 promoter) and (B)7 FAL(BRLF1 promoter)andRBR511(BRLF1 promoter), and allwere cutwith EcoRI.TheEBV contentof these clones is 48BAZ, 103188toabout 103650;RBR511, 106107 toabout106750; 7FAL, 106192toabout 106670.ProteinDNAcomplexeswereformedasforgelretardation, and then 1 pu1of25 mM MgCl2and1 pulofDNase (4 U/,u) were added. Afterdigestion for60 s at roomtemperature,phenol extrac-tion, and ethanol precipitaextrac-tion, samplesweredissolved in formam-ide andelectrophoresedon6% polyacrylamide gels in 8M urea.Size markers areanMspI digest of pBR322, endrepaired with Klenow DNA polymerase, and
[a-32P]dCTP.
Lanes 1 and 5, No added protein;lane2,1pul(0.5pug)of protein;lane3,2pA of protein; lane 4,4,ulof protein.protein bound an oligonucleotide spanning the consensus AP-1siteofthe standard promoter in agelretardation assay butdidnotbindto an oligonucleotide correspondingtothe AE4 mutant sequence (Fig. 4, lanes 6 and 12).
DISCUSSION
* * * *v Deletionanalysisof the BZLF1proteinhas shown thatthe
part ofthe protein between the BsmI and Hincll sites is required for sequence-specific DNA binding by BZLF1 at
C
i 2 3 4 5 6 the siteupstream
of thepromoter
forBSLF2+BMLF1. Thisisarefinement ofourearlier
mapping
of thisfunction. TheFIG. 5. CATassay of induction of S-CAT construct by BZLF1
deletion
mutant that lostDNA binding(N-PstI) also lostthe deletion mutants. p3.1W-BZLF1 fusions of the BZLF1 deletionability
to activate the promoter forBSLF1+BMLF1,
con-mutants(20 ,ug) were electroporated with the S-CATplasmid (20p.g)
sistent with the idea thatsequence-specific DNA binding is intoBL41/CL16cells. CAT activity was assayed 1 daylater. Lane part of the mechanismbywhich BZLF1 inducesproductive-C, No BZLF1; lane 1, BZLF1; lane 2, HindIII delta; lane 3, cycle transcription.
N-HincII; lane 4,N-StyI;lane 5,SmaI-C; lane 6,N-PstI. Although the BZLF1 sequences involved in the protein-J. VIROL.
6 .,
.--.'.- M'41-. 4b
M.-Ai..
ft. .
-ft &.
4ft
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.347.523.74.368.2] [image:5.612.105.254.77.338.2] [image:5.612.67.298.515.672.2]STRUCTURE AND FUNCTION OF EBV BZLF1 PROTEIN 2115
Promoter for BZLF1, probe 48.BAZ
170 180 190
103310 103320 103330
TACATTAGCAATGCCTGTGGCTCATGCATAG
<---ACCGAGT-->
ATGTAATCGTTACGGACACCGAGTACGTATC
Promoter for BRIE1, probe RBR.511
150 160 170
106210 106220 106230 CTGGTCTTTTATGAGCCATTGGCATGGGCG
<---TGAGCCA--->
8 -7 -6 -5
-4
-3 -2
-310 320 330
106370 106380 106390 AACTAAGCTTATGAGCGATTTTATCACAGG
<---TGAGCGA--->
TTGATTCGAATACTCGCTAAATAGTGTCC
Promoter for BRLF1, probe 7.FAL
60 70 80
106210 106220 106230 CTGGTCTTTTATGAGCCATTGGCATGGGCG
<----TGAGCCA--- -> GACCAGAAAATACTCGGTAACCGTACCCGC
220 230 240
106370 106380 106390 AACTAAGCTTATGAGCGATTTTATCACAGG
<---TGAGCGA--->
TTGATTCGAATACTCGCTAAAATAGTGTCC
1
F
r-
,n
- + - + - + - + BZLF1
S 257 AE4 AE4K
CAT CAT CAT CAT
Normalised data S CAT
S CAT+BZLF1 S CAT+BRLF1
%conversion Expt 1 Expt2 Expt 3
1.11 0.49 1.0 3.62 0.86 6.5 27.4
average 0.87 3.66
Alignment
AE4mutant ttcATctcgaggt
R R R
z
z
?consensus? AP1
ttcATGAGTCAgt cttATGAGCGAtt tttATGAGCCAtt tgcATGAGCCAca tacATtAGCaAtg Y-YAT-AGY-A- -TGAGTCA
FIG. 7. Relationship of footprints seen in Fig. 6 to DNA se-quence ofEBV. Theextent of each high-affinity footprint is indi-cated by the arrows and dotted lines. The numbers above each segmentrelate the EBVgenomenumbers(1) to thegel size markers inFig.6.Below this is shownanalignment of thefootprintswiththe previously established footprint(M;seereference7)in the promoter region of BSLF2+BMLF1. Thesequenceofthe AE4 mutantof the M sequence, which does not bind BZLF1, is also shown. The alignment of four ofthefootprint targets to a consensus AP-1 site is indicated together with a possible alternative consensus that matchesallfive bindingsites.
DNA interaction apparently lie between the BsmI and HincII sites, the SmaI-BsmI and HincII-C parts seem to
contribute significantly to the stability of this protein
do-main. We presumethatthe virtual absence ofaretardation complexwith BsmI-HincII isaconsequenceofthe instabil-ity oftheproperly folded structure (the DNA-protein
inter-action is
evidently
not very salt sensitive, since SmaI-Cbinds well in both high and low salt). The salt sensitivity
impliesthationic interactions areimportantinstabilizingthe
protein structure of the DNA-binding domain. There are several acidic residues in the SmaI-BsmI and HincII-C sections which may be involved in interactions with basic
amino acids in the basic-motif region stabilizingthe struc-ture. Inourassays, theSmaI-C segmentseemstobindjust
aswellasthe wholeproteinand,therefore, we conclude that the wholeDNA-binding domain liesC terminal of theSmaI site.
257CAT 257CAT+BZLFI 257 CAT+BRLF1 AE4CAT AE4CAT+BZLF1 AE4 CAT+BRLF1 AE4KCAT AE4KCAT + BZLF1 AE4KCAT+BRLF1
1.55 5.90 41.5
7.48 4.26 89.9
0.52 0.85 23.4
1.10 1.1 1.25 5.90 5.90 5.90
8.60 4.2 6.76 11.43 7.2 7.63
0.24 0.5 0.42 0.98 0.8 0.88
FIG. 8. Site-directed mutagenesis of the AP-1 site in the pro-moterforBSLF2+BMLF1. ReporterconstructsS-CAT, 257-CAT, AE4-CAT, and AE4K-CAT were electroporated alone or with CMV-BZLF1orCMVLTR-BRLF1 intoBL41 cells. After 2 days, extractsof the cellswereassayed for CATactivity.A totalof three experiments were performedwithBZLF1, butonly one was per-formed withBRLF1. The percentconversion ofchloramphenicolto acetyl chloramphenicol was determined, and to make the three experimentscomparableandgiveequal weighttoeachexperiment, the resultswerenormalizedonthe 257CATplusBZLF1figure. The true percentconversion figures for 257CAT plus BZLF1 were as follows: experiment 1, 2.02; experiment 2, 4.8; and experiment 3, 5.90. The normalizedfigureswereaveraged,and these averagesare plotted inthehistogramatthe top.
We identified binding sites for BZLF1 upstream ofthe promotersfor BRLF1, BZLF1, and BSLF2+BMLF1. Four of the five sitescontain close matches tothe AP-1 consen-sus, but the fifth site(inthe BZLF1promoter) doesnot. This site has also beenmappedaccurately by E.Flemingtonand S. H. Speck (submitted for publication). The variation in sites suggestsoneof threepossibilities:
(i)
twoDNA-bindingdomains are present in BZLF1, (ii) the same part of the BZLF1proteincanrecognizetwoapparentlyquitedifferent DNA sequences, or (iii) the truebinding site for BZLF1 is common to all the targets and is overlapping a consensus AP-1site infour of the five targetsmapped. Alignmentofall five sites shows a possible consensus that matches all the VOL.64, 1990
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.325.541.77.455.2]2116 PACKHAM ET AL.
targets, but ourfootprintingisnotyetsufficiently precise to determine the important nucleotides for binding. This will
require mutagenesis of individual nucleotides in the target
regions. We cannot yetexclude the possibilitythat BZLF1
contains two differentDNA-binding domains, but this can nowbe tested byusing our deletionmutantsand synthetic
DNAscontainingan AP-1-likeBZLF1 target orthe BZLF1 promoter target thatlacks the consensus AP-1 site. So far
our experiments on mapping the part of the protein that
binds DNA have all used one ofthe AP-1-like targets (the
oneupstreamof BSLF2+BMLF1).
In the EBV-negative Burkitt's lymphomacell line BL41, bothBZLF1andBRLF1areabletotransactivate the BSLF2
+BMLF1 promoter but it is clear that the activation by
BRLF1 isquantitively muchgreater. In Blymphocytelines
carrying EBV, the major routeby which BZLF1 activates
BSLF2+BMLF1 is, therefore, presumed to be through
BZLF1first activating BRLF1,whichthenactivates BSLF2
+BMLF1. Nevertheless, thereisamodest effect ofBZLF1 on thepromoterfor BSLF2+BMLF1 andwe have
investi-gatedthis. Oneinteresting explanation forourresults is that
atleastpartof the effect ofBZLF1onthispromoterinvolves counteracting acellularrepressorof thepromoterfunction.
That repressor seems to require sequences overlappingthe consensus AP-1 site in the promoter
region. Although
we have not studied this repression function directly, obvious candidatestomediate suchrepression wouldbe membersofthe cellular AP-1 family of transcription factors. There are well-established precedents for these factors acting as re-pressors, aswell as in their more usualactivating function, for example, in the promoters ofc-fos (19) and c-myc (9).
The effectiveness of such repression would presumably be
influencedby endogenous levels ofthe appropriatecellular AP-1factors and the mode of presentation andquantity of
DNA in transfection assays. These variables may account
for differences between cell lines and another group not
observing
thiseffect ofBZLF1onthe promoterforBMLF2 +MLF1 (10).At present, we do nothave the methods to testwhether
this apparent ability of BZLF1 to counteract a cellular
repressoris importantin the EBV genomeduring anormal infection. The EBVproductivecycleispresumablyinduced
by cellular changeswhicharereflectedin alteredpatternsof cellulartranscription factors.Theasyetunidentified factors
that switch on BZLF1 transcription are presumably the
primary trigger, butautoactivation by BZLF1 andaloss of repressionby cellular factors that recognize sequences
con-tainingtheconsensusAP-1sitesmayassist theswitchof the
virus from being latent to fully productive. Perhaps the
high-affinitynon-AP-1-likesite upstreamof BZLF1, which is
involved in autoactivation of BZLF1 in B lymphocytes
(Fleming and Speck, submitted) permits accumulation of
sufficient BZLF1 to overcome repressing cellular factors fromother AP-1-like targets and thenswitchonthe produc-tivecycle.
LITERATURE CITED
1. Baer, R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P. J. Farretl,T.J. Gibson, G. Hatfull, G. S. Hudson, S. C.Satchwell, C. Seguin, P. S. Tuffnell, and B. G. Barrell. 1984. DNA sequence and expression of the B95-8 Epstein-Barr virus
ge-nome.Nature(London)310:207-211.
2. Biggin,M., M. Bodescot, M. Perricaudet, and P.Farrell. 1987. Epstein-Barr virus gene expression in P3HR1-superinfected Raji cells.J.Virol.61:3120-3132.
3. Chevallier-Greco, A., E. Manet, P. Chavrier, C. Mosnier, J.
Daillie,and A.Sergeant.1986. BothEpstein-Barrvirus
(EBV)-encoded transacting factors, EB1 and EB2, are required to activatetranscriptionfromanEBVearlypromoter. EMBO J. 5:3243-3250.
4. Countryman, J., H.Jenson, R. Seibi,H.Wolf,and G. Miller. 1987.Polymorphicproteinsencoded within BZLF1 of defective and standard Epstein-Barr viruses disrupt latency. J. Virol. 61:3672-3679.
5. Countryman,J., and G. Miller. 1985. Activation ofexpression
of latent Epstein-Barr herpes virus aftergene transfer with a smallclones subfragment ofheterogeneous viral DNA. Proc. Natl.Acad. Sci. USA 82:4085-4089.
6. Farrell,P.J. 1989. TheEpstein-Barrvirusgenome.Adv. Viral Oncol. 8:103-132.
7. FarreUl,P. J., D. T.Rowe, C.M. Rooney, and T.Kouzarides. 1989. Epstein-Barr virus BZLF1 trans-activator specifically
bindsto a consensusAP-1site and is relatedtoc-fos. EMBO J. 8:127-133.
8. Hardwick, J. M.,P. M.Lieberman,and S. D.Hayward.1988. A newEpstein-Barrvirustransactivator, R,inducesexpressionof acytoplasmic early antigen. J. Virol. 62:2274-2284.
9. Hay, N.,M.Takimoto,andJ.M.Bishop.1989.AFOSprotein
is present inacomplexthat bindsanegative regulatorof MYC. GenesDev. 3:293-303.
10. Kenney,S.,E.Holley-Guthrie,E.-C.Mar,and M. Smith.1989. TheEpstein-BarrvirusBMLF1promotercontainsanenhancer elementthat isresponsivetothe BZLF1 and BRLF1
transacti-vators.J. Virol. 63:3878-3883.
11. Kenney, S., J. Kamine, E. Holley-Gutherie, J.-G. Lin, E.-C. Mar, and J. Pagano. 1989. The Epstein-Barr virus (EBV)
BZLF1 immediate-earlygene product differentially affects
la-tent versusproductiveEBVpromotors.J.Virol. 63:1729-1736.
12. Kenney,S.,J.Kamine,E.HoUley-Guthrie,E. C.Mar,J.C.Lin, D. Markovitz, and J. Pagano. 1989. The Epstein-Barr virus immediate-early gene product, BMLF1, acts in trans by a
posttranscriptional mechanism which is reporter gene
depen-dent.J.Virol.63:3870-3877.
13. Knutson, J. C., and B. Sugden. 1989. Immortalization of B lymphocytes by Epstein-Barr virus: what does the virus con-tributetothecell? Adv. Viral Oncol. 8:151-172.
14. Kunkel,T.A.1985.Rapidand efficientsite-specificmutagenesis
without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
15. Lieberman, P.M., J.M.Hardwick,andS. D.Hayward.1989. ResponsivenessoftheEpstein-BarrvirusNotIrepeatpromoter totheZtransactivator is mediatedinacell-type-specificmanner
bytwoindependent signal regions.J. Virol.63:3040-3050. 16. Lieberman, P. M., P. O'Hare, G. S. Hayward, and S. D.
Hayward. 1986. Promiscuous transactivation ofgene expres-sionbyanEpstein-Barrvirus-encodedearly protein. J. Virol. 60:140-148.
17. Oguro,M.O.,N.Shimizu,Y.Ono,and K. Takada.1987.Both therightwardand theleftwardopenreadingframeswithin the BamHI M DNAfragmentofEpstein-Barr virus act as trans-activators ofgeneexpression. J. Virol.61:3310-3313.
18. Rooney,C.M.,D. T.Rowe,T. Ragot,and P.J. Farrell.1989. ThesplicedBZLF1 geneofEpstein-Barrvirus(EBV) transac-tivatesanearlyEBVpromoterand inducesthe virusproductive cycle.J.Virol.63:3109-3116.
19. Schontal, A.,M.Buscher,P.Angel,H.J.Rahmsdorf,H.Ponta, K.Hattori,R.Chiu,M.Karin,and P. Herrlich. 1989. The Fos andJun/AP-1proteinsareinvolved in thedownregulationof Fos transcription. Oncogene4:629-636.
20. Takada, K., and Y. Ono. 1989. Synchronous and sequential
activation oflatentlyinfected Epstein-Barrvirus genomes. J. Virol. 63:445-449.
21. Takada, K.,N. Shimizu,S. Sakuma,andY. Ono. 1986. trans Activation of the latentEpstein-Barrvirus(EBV)genomeafter transfection of theEBV DNAfragment.J.Virol.57:1016-1022. 22. Urier, G.,M.Buisson,P.Chambard,and A.Sergeant.1989.The Epstein-Barr virus early protein EB1 activates transcription from differentresponsiveelementsincludingAP-1bindingsites. EMBO J. 8:1447-1454.
J. VIROL.