0022-538X/91/084078-13$02.00/0
Copyright C) 1991,American Society for Microbiology
A
Cellular
Function
Can Enhance
Gene
Expression
and
Plating
Efficiency of
a Mutant
Defective
in the Gene for ICPO, a
Transactivating Protein of
Herpes
Simplex Virus Type 1
WEIZHONG CAI ANDPRISCILLA A. SCHAFFER*
Laboratory of Tumor Virus Genetics, Dana-FarberCancer Institute, and Department of Microbiology and Molecular Genetics, Harvard MedicalSchool, Boston, Massachusetts 02115
Received 25 February 1991/Accepted7 May 1991
ICPO transactivates herpessimplex virus type 1 genesofall classes as well as a number ofheterologousviral and cellulargenes, yetit is not essentialforvirusreplicationinvitro or in vivo. Stocks of ICPO deletion mutants, however, exhibit significantly lower plating efficiencies on standard24-h-old Vero cell monolayers than do stocksof wild-typevirus. In an attempt todeterminewhether thegrowthstatusof cellsinthemonolayeraffects the abilityofICPO mutants toinitiateplaque formation, theplating efficiencies and abilities of an ICPO null mutant (7134) andofwild-type virus (KOS) to express selected viral proteinswere determined onVero cell monolayers whose growth had been arrested either by contact inhibition-trypsinization or by isoleucine deprivationand had then been released fromgrowtharrest.Theproportionofcellscyclingsynchronouslyafter releasefromgrowtharrest wasassessed by flow cytometry.The results of these studies indicate thattheplating efficiencyof 7134wasgreateston Verocellmonolayers8hafterreleasefrom growtharrestinducedbyeither treatment. Monolayers of both types released from growth arrest at other times supported 7134 plaque formation lessefficiently.Incontrast, theplating efficiency of KOSwasnearly equalonmonolayers at alltimes after release from growth arrest. Notably, both KOS and 7134 were equally efficient in enteringcells and inducing expression of the immediate-early protein ICP4in either 8- or 24-h monolayers. Relativeto KOS, however, 7134was significantly impaired in the expression of selected earlyand late genesin cells at 24 h postrelease. When theplating efficiencies of 7134 and KOS wereexaminedin0-28 cells (Vero cells that are stably transformedwith theICPO gene)whosegrowthhad beenarrested and thenreleased, nodifferences in theplatingefficiencies ofthe twoviruses as afunction of growthstatus werenoted. Thesefindings suggestthat acellularfunctionexpressedmaximally in cells8hafterreleasefrom growtharrest cansubstitute operationally forICPOtoenhanceplaqueformation and viral geneexpression by7134. They further suggest that onerole ofICPOin viral infection is tofacilitatevirus replicationin cells that do not express thisfunction.
The regulation ofviral geneexpression involvescomplex interactions among multiple viral and cellular factors. Be-causeoftherelativeeasewithwhich viruses canbe manip-ulatedgenetically,the functions ofviral proteinsinvolvedin
theseregulatoryinteractions have been better characterized than those ofcellular proteins. Specifically, the regulatory
proteins encoded by herpes simplex virus type 1 (HSV-1) have beenextensivelycharacterizedintermsof their roles in viral replication and their effects on specific classes of viral promoters. Without adetailed understanding of the roles of cellular factors in this process, however, the mechanisms
underlying regulation of HSV-1 gene expression cannot be
completely understood. Inthisreport weshow thataviable but replication-deficient mutant with a mutation in the HSV-1 regulatory gene specifying ICPO has facilitated the identification ofa cellularfunction that activates viral gene
expression.
The HSV-1 genomecontainsatleast72genes(49-51, 59). These genes areclassified into three major groups basedon the kinetics of their expression following viral infection: immediate-early(IE), early (E), andlate (L)(32). Viralgene
expression is coordinately regulated at the transcriptional and posttranscriptional levels (10, 25, 34, 75). The
expres-sion of the IE genes is activated by a component of the incoming virus particle,VP16, immediately following
infec-*Correspondingauthor.
tion (5, 44). The IEproteinsinturn induceandregulate the
expression ofEand L genes(33) whoseprotein productsare involved in viral DNAsynthesisandconstitutevirion struc-tural components, respectively.
Five IEproteins have been identified and characterized:
infected-cell proteins(ICPs) 0, 4, 22, 27, and 47 (32).Twoof
these, ICP4 andICP27,areessentialfor virusreplication,as demonstratedby thepropertiesoftemperature-sensitiveand other lethal mutations in these two genes (14, 16, 47, 67).
ICP4 represses the expression of all five IE genes and activates theexpression ofEandLgenes(13, 15-17,23,24,
46, 55, 56, 62). ICP27 modulates the expression of both E and L genes inacomplexandpoorly understoodmanner(17,
47, 52, 63, 64, 67, 71). The other three IE proteins
(ICPO,
ICP22, and ICP47) are not essential for virus replicationin cell culture, since mutant viruses lacking genes for these
proteins are replication competent, although replication is impairedto somedegree (43, 61, 68, 70, 72).
In transient expressionassays
ICPO
activates the expres-sion of all classes of HSV-1 genes (IE, E, and L) andgenes of otherviruses, aswellasanumberof cellular genes(3, 18,19, 23, 24, 46, 53-56, 62, 71). Despite its promiscuous
transactivating activity, ICPOis notrequired forvirus repli-cation; however,deletion of bothcopiesof thediploidICPO
generesults inanoverall reduction in theexpressionof viral proteins,especiallythose of theLkinetic class (68, 72). ICPO mutants replicate in cultured cells in a
multiplicity-depen-dentfashion, i.e., they grow poorly following
low-multiplic-4078
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 ICPO-LIKE CELLULAR FUNCTION 4079 ity infection but well following high-multiplicity infection
(68, 72). Consistent with the requirement for ICPOfor viral growth at low multiplicities, deletion of the ICPOgenes also results in substantially reduced de novo synthesis of infec-tious virus following transfection of cells with infecinfec-tious mutant viral DNA (3). Given their impaired ability to repli-catein cell culture at low multiplicities of infection, it is not surprising that ICPO mutants are compromised in their ability to produce infectious virus after reactivation from latent infections (4, 9, 39). Notably, the relative ability of ICPO deletion and nonsense mutant viruses to produce infectious virus following reactivation from latency in a mouse ocular model correlates well with the ability of the corresponding mutant proteins to transactivate HSV pro-moters in transient assays (3, 4). This observation indicates that the transactivating activity ofICPO is critical for virus replication associated with acute infection and for virus reactivation from latency.
The mechanism by whichICPO transactivates so broad a spectrum of eukaryotic promoters is not known. ICPO is a nuclear phosphoprotein that may bind to DNA nonspecifi-cally (1, 30, 58). It contains two potential zinc fingerdomains that are probably involved in DNA binding and transactiva-tion (19, 20, 60). Available evidence indicates that ICPO activates any promoter that exhibits a basal level ofactivity, although it does not appear to require any particular cis-acting element, including the TATA box, for this activity (54). It is thought that ICPO mediates its activity through cellular transcriptional machinery; however, it is notknown which cellular factor(s)interacts with or is induced by ICPO or under what circumstances this interaction and induction occurs. Moreover, it is possible that ICPO mediates its effects at the posttranscriptional level.
In an attempt to further characterize the transactivating function ofICPOand to identify putative cellularfunctions involved in the activation process, we have examined the replication efficiency of anICPOnull mutant relative to that of the wild-type virus in partially and highly synchronized cell populations. Our results indicate that a cellularfunction expressed at maximal levels shortly after release from growth arrest can substitute forICPO to enhance viral gene expression and plaque formation by the null mutant. They further suggest that one role ofICPO in viral infection is to facilitate virus replication in cells that do not express this function.
MATERIALS AND METHODS
Cells and viruses. Vero cells and ICP0-transformed Vero cells (0-28 cells) (68) were grown andmaintained in Dulbec-co'smodified Eagle's medium supplemented with 10% fetal bovine serum (growthmedium) as previously described (67). Cells were passed twiceweekly and reached numbersof 2 x
107 to3 x 107 cells per 150-cm2 flask.
Wild-type HSV-1 KOS and anICPO null mutant derived from KOS, 7134 (3), were propagated and assayed as de-scribed previously (69).
Determination of particle numbers in virus stocks.Cell-free stocks of KOS and 7134 werediluted to contain an estimated 108 particles per ml. Quantitation of virus particles was done by the droplet-pseudoreplication method described previ-ously (48). The averagenumber of particles in five fields was determined for each stock.
Determination of HSV DNA content of viral stocks. Ali-quots of mutant andwild-type virus stocks were diluted in 300 ,ul ofTris-buffered saline (8.0 g of NaCl, 0.38 g ofKCI,
and 3.0 g of Tris base per liter, pH 7.4). Viral DNA associated with cell debris wasremoved by low-speed cen-trifugation. Supernatant fluidswere subjectedto
high-speed
centrifugation inamicrocentrifugefor 30 mintopellet
virus particles. Supernatantfluids and associated viralDNAwere discarded, and virusparticleswereresuspended
in TEbuffer (10 mMTrisHCl,1 mMEDTA, pH8.0)containing
0.2 mgof proteinaseKpermland0.6% sodiumdodecyl
sulfate(SDS)
for4 h. Knownconcentrations of
purified
KOS DNA were also incubated with theproteinase K-SDSsolution to serve as standards. Solutions were then extracted with anequal
volume ofphenol-chloroform-isoamyl alcohol(25:24:1),
andviral DNA was pelleted after ethanol
precipitation.
After resuspension and denaturationbyheating
at100°C
for 5min,viral DNA wastransferredto anitrocellulose filter in aslot blot apparatus (Minifold II; Schleicher &
Schuell,
Inc.,
Keene, N.H.). The blot was
probed
with the EcoRI-DfragmentofHSV-1KOS DNA. After exposureto
X-ray
film and development oftheautoradiogram,
thesignal
in each slotwasmeasuredbyscanning
densitometry
and theamount of DNAwascalculatedby
using
astandardcurvegenerated
from the signals of known HSV-1 DNA standards.Synchronization of Vero cells by trypsinization and
by
isoleucine deprivation.
Partially growth-arrested
Vero and 0-28 cells were obtainedby
trypsinization
andseeding
of confluent cells in normalgrowth
medium.Highly
growth-arrested Vero and 0-28 cells were obtained
by
isoleucinedeprivation
essentially
as describedby
Hamlin and Pardee (28) andLeyandTobey
(41). Cells tobe blocked ingrowth
arrest werefirsttrypsinizedand seeded in
growth
mediumat37°Cfor24h. Mediumwasthen
changed
togrowth
mediumlackingisoleucine andwas
supplemented
with 10%dialyzed
fetal bovineserum. Medium
lacking
isoleucinewasprepared
on the basis of thecomposition
of Dulbecco's modified Eagle's mediumasdescribedby
FlowLaboratories(Product
no. 10-331), except that no isoleucine was added. After growth in medium
lacking
isoleucine for 42h,
cells were grown inisoleucine-containing growth
medium to reverse the block induced by isoleucinedeprivation,
harvested foranalysis by flow cytometry, or infected at various times
postrelease fromgrowth arrest and harvested at later times for further analysis.
Isoleucine-deprived
Vero cells aregrowth arrested in the
GdG1
stage of the cellcycle
(see
below)and donotdivide until 20to24hafter the addition of isoleucine tothe culture medium.
Flowcytometry. The
proportions
of cells in theG0/G1,
S,andG2/M phases ofthe cell
cycle
were determinedby
flow cytometryusing
chromomycin A3
as theDNA-binding
fluo-rescent dye (22). For flowcytometric
analysis,
monolayers
in 35- or 60-mm-diameter
petri
dishes were harvestedby
trypsinization, and cells were
pelletted
by
centrifugation
at 1,500 rpm for 10 min at4°C
in a Sorvall RC-3centrifuge.
Afterbeingwashed with
phosphate-buffered
saline(PBS) (8
g ofNaCl, 0.2 g of
KCl,
1.44 g ofNa2HPO4,
and 0.24 g ofKH2PO4 per liter, pH 7.4) and
being
repelleted,
cells wereresuspended in 1 ml ofcold PBS and added
dropwise
to anequalvolume of cold
(4°C)
80% ethanol withgentle
mixing.
Ethanol-fixed cells were stored at
4°C
beforebeing
stained. For chromomycinstaining
and flow cytometry, cells were pelleted, washed withPBS,
repelleted,
andresuspended
in 0.5 mlofasolutioncontaining
0.02 mgofchromomycin
A3 and 3 mgofMgCl26H20
per ml of PBS. After incubation onicefor 30 min indarkness,
cellswerecounted inaCoulter EPICS 753 flow cytometer, and data files were transferred into flow cytometry standard formatby
using
Coulter Veclink software.Histograms
weregenerated,
and areasVOL. 65,1991
on November 10, 2019 by guest
http://jvi.asm.org/
undereachpeakwereintegrated by usingBecton Dickinson
Consort/VAXsoftwaretodetermine thepercentages of cells in
Go/G1,
S,andG2/Mof the cellcycle. The flowcytometric method used inthese studiescannotdistinguish betweenGo
and G1orbetweenG2andM; hence,we usetheterms
Go/Gl
andG2/M tosignify all cellsinthese collectivestages of the cellcycle.
Immunofluorescence assays. Growth of Vero cell cultures
wasarrested byincubation in mediumlackingisoleucine for 42 h. Petri dishes (diameter, 35 mm) containing sterile coverslipswere thenseeded with 2.7 x 105 growth-arrested
cells, and resulting monolayers were grown in standard mediumto release themfrom growth arrest. At 8 and 24 h after release and seeding, monolayers were infected with either KOS or 7134. Monolayers infected 8 h postrelease fromgrowtharrestwereinfected with 0.03 PFUpercell and thesamenumberof PFUwasusedtoinfectmonolayersat24 hpostrelease. Since the cell numberin 24-hmonolayerswas
twice that in8-h monolayers, the actualmultiplicity in 24-h monolayers was 0.015 PFU per cell. At 8-h postinfection, coverslipswere removed, fixed, and stained with monoclo-nal antibodies directed against individual viral proteins. Cellsoncoverslips were stained bytheindirect
immunoflu-orescence procedure describedby Quinlan et al. (62). The
antibodies used inthesetestswere asfollows: 58S, directed against the IE protein ICP4; 39S, directed against the E protein ICP8; and56S, directedagainsttheLprotein ICP5. Monoclonalantibodies werepreparedasascites fluidsfrom hybridoma cell lines obtained from the American Type CultureCollection (39Sand 58S) orMartinZweig(56S).
Proteinlabeling and immunoprecipitation. Vero cells (2.7 x 105) whose growth had been arrested by isoleucine
dep-rivation were trypsinized, seeded in 35-mm-diameter petri dishes, incubated at 37°C in medium containing isoleucine, and infected 8 or 24 h later with either KOS or 7134. The multiplicity of infection used to infect 8-h cultures inthese experimentswas0.03 PFUpercell,andthesamenumber of PFUwasused toinfect24-hcultures.Because the number of
cellseffectively doubledinthe16-hinterval,themultiplicity ofinfection in 24-hcultureswas0.015 PFUpercell. Infected cultures were pulse-labeled at6 and 8 h postinfectionwith 200 RCi of[35S]methionine (DuPont NEN Research Prod-ucts, Boston, Mass.)in 100 ,ulof Tris-buffered saline for 20 min. After the pulse, cells were washedwith Tris-buffered saline and lysed with lysis buffer containing 1 mM Na-p-tosyl-L-lysine chloromethyl ketone and 5% dry milk to inhibitproteaseactivitypresent in Vero cellextracts(2). Cell extracts were then subjected to immunoprecipitation as
described by Holland et al. (31) by using the following pooled monoclonal antibodies directedagainstthe indicated HSV-1 IE, E,and Lproteins: 58S (ICP4; IE), 56S (ICP5; L), 51S (ICP6; IE),39S (ICP8; E), 3S (gB; L),and 19S (gC; L). Monoclonal antibodies 39S and 58S were prepared as de-scribed above for immunofluorescence tests; 56S, 51S, 3S, and 19Swerekindly provided byMartin Zweig (Frederick,
Md.). Precipitated proteins were separated by
SDS-poly-acrylamide gel electrophoresis (36, 45) and visualized by autoradiography.
RESULTS
Propertiesof KOS and7134 virusstocks. TheICPOgeneis present in two copies in the inverted repeat regions
sur-rounding the unique long component of the viral genome
(60). An ICPO null mutant, 7134,wasgenerated by deletion
[image:3.612.325.564.88.280.2]of the two ICPO coding regions and insertion of the coding
TABLE 1. Propertiesof KOS and 7134 stocks
Parameter KOS 7134
Physical particles(1011 PP/ml)a 1.4 2.3
HSVgenome content(1010 8.3 7.9
genomes/ml)b
Titer(PFU/ml)c
Verocells 2.5 x 109 1.7 x 108
0-28 cells 1.3 x 109 1.8 x 109
Physical particlenumber-to- 1.7 2.9
genomenumber ratio Physical particle-to-PFU ratio
Vero cells 56 1,353
0-28 cells 108 128
Genome number-to-PFU ratio
Vero cells 33 465
0-28 cells 63 44
" Physical particle (PP) numbers were determined by electronmicroscopic quantitationofnegatively stained virions in cell-free stocks byusing standard procedures(48).
bTheconcentration of HSV-1 DNA in virus stockswasdeterminedbyslot
blothybridization and scanningdensitometry. Genome numberswere calcu-latedonthe basis of themolecularweight of the HSV-1 genome(108)byusing knownamountsof HSV-1DNA asstandards.
' Virus titersweredetermined bystandardplaque assaysonmonolayers24 hpostseeding.
region of the lacZ gene (3). By definition, 7134 also lacks sequences that specify portions of the latency-associated transcripts (LATs) whichareencoded by the strand opposite to that which encodes ICPO. Like ICP0, the LATs are not essential for virus replication, as LAT mutants are replica-tion competent (38). 7134 expresses
P-galactosidase
but does not expressICPO. Because theplating efficiency of 7134 was significantly lower than that of the wild-type virus on Vero cellmonolayers,it isprobablethatstocks of this mutant, like stocks of other ICP0 mutants,exhibithigher particle-to-PFU ratios than stocks of wild-type virus (72).Inordertofurther characterize 7134, it was first necessary to normalize the particle number, the genome-containing particle number, andthetiter of the 7134 stock preparation with respect to the same parametersof the KOS stock (Table 1). Physical particle counts were determined by electron
microscopy of negatively stained preparations (48). The
physical particlecount was1.4 x
1011/ml
for the KOS stock and 2.3 x1011/ml
for the 7134stock. Because the number of genomes in a virus preparation may be a more sensitive measure of the number ofpotentially infectious virions it contains thanparticle numberis(i.e., notall virusparticles contain viral DNA), we also measured the numbers ofgenome-containing particles in the two stocks. For this purpose, the DNAcontent of each stock wasquantified by slotblot hybridization and scanning densitometry by using knownamountsof viralDNA asstandards. Theprobe used to detect viral genomes by slot blot analysis was derived from the EcoRI-D fragment which contains no replication
originsorpackagingsequences(26). Hence, the genomes of the classical noninfectious defective interfering particles
which lack sequences in EcoRI-D but are present in all virus stocks were not detected (42, 74). The genome numbers obtained in this way shouldreflect the number of
genome-containingvirions in each stock. The genome contentofthe
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 ICPO-LIKE CELLULAR FUNCTION 4081 TABLE 2. NumbersofVerocellsexpressingICP4,
ICP8,andICP5'
Ageof Avg no. of fluorescence-positive cells/avg Virus monolayer no. of cells per field
(h)b ICP4 ICP8 ICP5
KOS 8 8.0/100 4.2/105 3.5/99
24 8.1/226 4.1/219 3.3/222
7134 8 9.0/108 0.79/108 0.21/106
24 8.6/225 0.04/220 <0.04/220
aEight-and 24-hVero cell monolayerswereinfectedwiththesamenumber of PFUwhichwasdeterminedon0-28 cells(andthereforeapproximately the
same numbers ofparticles [Table 1]) of either KOS or7134. For 8-h-old
cultures, the multiplicity of infectionwas 0.03 PFU percell,and for 24-h
cultures, themultiplicity of infectionwasapproximately0.015 PFU percell,
ascell numbershad nearly doubledbetween 8 and24h.At8-hpostinfection, cellsoncoverslipswerefixed andstained withantibodytoICP4,ICP8, or
ICP5.Atotal of24fieldswascountedpersample. Theaveragenumber of cells
perfieldand the averagenumber offluorescence-positive cellsperfieldwere thendetermined.
bAttime ofinfection.
KOS stock was 8.3 x
1010
genomes perml and thatof the 7134 stock was 7.9 x1010
genomes perml. These numbersindicate that the stock of KOS contained 1.7-fold more physicalparticlesthangenome-containing particles and that thestock of 7134contained 2.9-foldmorephysical particles thangenome-containing particles. Stocks were then assayed
simultaneously on standard 24-h-old Vero and 0-28 cell monolayers. 0-28 cells are Verocells that are stably
trans-formed with the ICPO gene (68). These cells complement ICPO-deficient mutants for virus growth and enhance the platingefficiency ofICPOmutants15- to20-fold (68). In these assaysthetiter oftheKOS stock wasconsistently approxi-mately 20-fold higher than the titer of 7134 on Vero cell
monolayers,whereastitersofboth viruseswerenearly equal
on 0-28 cell monolayers. On the basis of these data, the
physical particle-to-PFU ratioonVero cellswas56for KOS
and 1,353 for7134, and on 0-28cells,itwas108forKOS and 128 for 7134. Genome number-to-PFU ratios were also calculatedforthetwovirusstocksonVero and 0-28cells. As wasthe casefor the physicalparticle-to-PFU ratio,theratio of genome number to PFU of7134 relative to KOS was
significantlyhigheronVero cells(14-fold)thanon0-28cells
(<1-fold).Because theparticle-to-PFUand genome number-to-PFUratios ofthe 7134 stockon0-28 cells approximated
thoseoftheKOS stock,the 7134 stockcontainednearlyas manyinfectiousparticles permilliliter asthe KOS stock.
To exclude thepossibilitythatthelowerplatingefficiency
of 7134 virus on 24-h-old Vero cell monolayers was a consequence of7134 virions entering these cells less effi-ciently than KOS did, monolayers ofVero cells were
in-fected with KOS or7134at a multiplicity of infection of1 PFU per cellonthebasis of titers on 0-28 cells. Extracellular virus(including virusthat had adsorbedtocells but had not
entered)wasremovedby low pH treatmentfor 1minat2h
postinfection (2). Totalcellular DNAwasthenisolated,and viral genome numbersweredetermined by slot blot hybrid-ization.Approximately 10%of theinputviral genomeswere
detected in DNA from cells infected with either KOS or
7134, indicating that genome-containing particles of both KOS and 7134 entered the cells with equal
efficiency.
Moreover, wehaverecently shown that 7134andanumber ofICPOnonsense mutantsarenotdefective in entryorin the
expressionof ICP4 in Vero cells (4) (confirmatorydataare
presentedinTable2). On the basis of thesefindingsand the
Virus titer
(PFU/plate)
2000 \
7134
100-
KOS
0
04 812 24
Time
post-seeding
(hr)
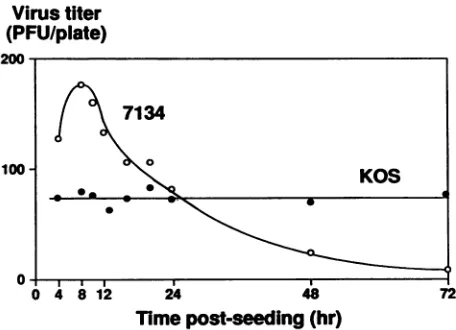
FIG. 1. Platingefficiencyof 7134and KOS on Veromonolayers at various times after trypsinization and seeding. Petri dishes (diameter, 35mm)were seeded with 2.7 x 105cells. Theresulting monolayers were infected at various times postseeding with 100
PFU of either 7134or KOS(input multiplicities of infection were
calculatedby usingtiters determinedon 24-h-old Vero cell mono-layers [Table 1]). Infectedmonolayerswereoverlaidwith methyl-cellulose, and plaques were visualized and counted on day 5 postinfection. Plaque numbers shown in this and allfigures repre-sent theaveragenumber ofplaquesobtained fromduplicatedishes
unless otherwise indicated. Variations inplaquenumbersin dupli-catedisheswere less than 10%.
datapresentedin Table 1,we conclude that thenumbers of
genome-containing particlesin the stocksof KOSand 7134 were approximately equal andthat thetwo viruses entered
thecells withequalefficiency.
Dependence of 7134plating efficiencyonthe ageofthe Vero cellmonolayer. The data
presented
inTable 1 demonstrate thatalthoughthestock of 7134contained approximatelythe same numberofpotentially
infectiousparticles
asthestockofKOS,theabilityof 7134
particles
toproduceplaques
wasmarkedlyreducedcomparedwith thatofKOSon24-h Vero cellmonolayers. One
explanation
for thisobservation is that asubset of cells in 24-hmonolayers mightnotsupportvirusreplicationby7134virus and thatpermissivity mightvaryas a function ofthe growth status ofindividual cells
and, by
extension,
as afunction ofthe age ofthemonolayer
post-seeding. Totestthishypothesis,Vero cellswere
trypsinized
ahd seeded in petri
dishes,
and theresulting monolayers
wereinfectedatvarious timespostseeding with 100 PFUof
KOSor7134. Titersof virus stocks usedtoinfect
monolay-ers werethosedetermined
by
plaqueassaysonstandard 24-h Verocell monolayers (Table 1).As shown in
Fig.
1, theplating
efficiency
of KOS wasrelatively independent ofthe ageofthe Verocell
monolayer
atthe timeof
plating:
KOS scored withapproximately
equal
efficiency
on 4- to 72-h-oldmonolayers.
In contrast, theplating
efficiency
of 7134washighly dependentuponthe age ofthemonolayer
postseeding;
titerswerehighest
whenvirus was addedto monolayers thatwere 8 h old anddeclined on oldermonolayers.
Theplating efficiency
of 7134 on 8-h-oldmonolayers was always two- to fourfold
higher
than thatobservedon 24-h-old
monolayers
in repeatexperiments.
As shown inFig.
1,7134produced nearly
200plaques
on8-h-oldmonolayers, a number twofold
higher
than the number of KOSplaques. Recall that this is because 7134stockscontainlarge numbers of
nonscoring
particles
that arepotentially
VOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.47.293.96.175.2]Virus
titer
(PFU/plate)
KOS KOS
5.6x1
031.7x10
100 - particles particles 300
60
- 1 73 - 3000l |
~~~168
-200
100
0
0
20-0 7134 5 19.6 7134
4_20
200
184 1.4xl
1.4xpr0i-20
1
00
1
I0
0
1w
Im
I0
8
24
8
24
[image:5.612.74.303.71.325.2]Time
post-seeding (hr)
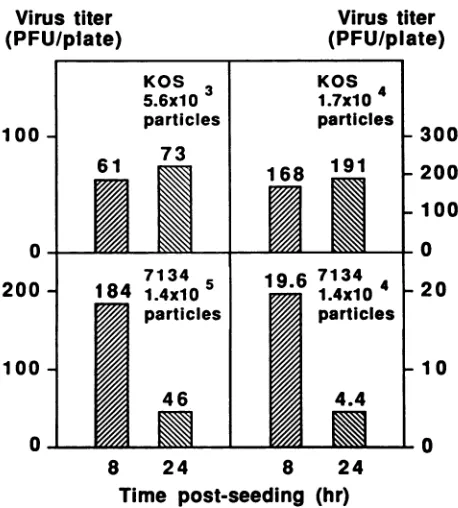
FIG. 2. Effect ofmultiplicity of infection of virus particlesonthe
plating efficiency of 7134. Petri dishes (diameter, 35 mm) were
seeded with2.7 x 105Vero cellsasdescribed in the legendtoFig. 1, and the resulting monolayerswereinfectedat8or24 h postseed-ing with the number of virus particlesindicated within eachpanel. Infected monolayers were overlaid and plaques were counted on
day5postinfection. Numbers shown representtheaverage plaque
countfrom two dishes, exceptfor numbers shown in the bottom
rightpanel, which represent the average number ofplaques in 10 dishes.
infectious (i.e., particles that do not produce plaques on
24-h-old Vero cell monolayersbut that do produce plaques
on 0-28 cells). Thus, although the number of7134 plaques that scoredon8-h-monolayerswasgreater thanthatofKOS, theplating efficiencyof 7134on8-hmonolayerswasactually
lower thanthat of KOS.
Thedifferences in theplatingefficiencies of KOS and 7134
on Vero cell monolayers can be explained either by the
presence ofa cellular function that facilitates 7134 plaque formation or by differences in the total multiplicities of infection of KOS and 7134 viruses used in these studies. In ordertoaddress the latterpossibility, 8- and 24-h-old Vero cellmonolayerswereinfected either withanequalnumberof KOSor7134 PFUorwith approximately equalnumbers of
virusparticles. The data shown in the left panels ofFig. 2 represent a repeat of the experiment shown in Fig. 1, in whichapproximately 100PFU ofKOS or7134was usedto infect 8-and24-h-oldmonolayers. Inthistestasinprevious tests, the plating efficiency of KOS on 8-h-old monolayers was nearly the same as that on 24-h-old monolayers,
whereastheplatingefficiency of 7134on8-h-oldmonolayers was fourfold higher than thaton 24-h-old monolayers. The
numberof7134particleswas24-fold higher thanthat of KOS
in this experiment. We next repeated the experiment, this time controllingfor the numberofgenome-containing parti-cles of the two viruses (Fig. 2, right panels). For this
purpose, the number of genome-containing particles was
increasedbythreefold forKOS, resultinginacorresponding
threefold increase in KOS PFU, and the number of 7134
genome-containing particles wasreducedby 10-fold, result-ing in a 10-fold decrease in 7134 PFU. Although the input multiplicities of infection of KOS and 7134 particles were
nearlyequal with these adjustments, the plating efficiency of KOS remained constant on 8- and 24-h-old monolayers, whereas that of 7134 on 8-h monolayers was fourfold higher than on 24-h monolayers. Therefore, the variation in 7134 plating efficiency was not a consequence of the total number of virus particles with which monolayers were infected but of the age of the monolayer at the time of infection. We further conclude that a Vero cell function expressed in 8-h-old monolayers can substitute for the viral function missing in 7134 to enhance plaque formation.
This conclusion is further supported by the following simple calculations. When 2.7 x 105 cells are infected with 100 PFU of 7134 (determined on the basis of the titer on 24-h-old Vero cell monolayers), each cell receives on aver-age 0.5 physical particles, 0.17genome-containing particles, or 0.005 potentially infectious particles (particles that score on 0-28 cells) (Table 1). At this very low multiplicity of infection, few if any cells are infected with more than one infectious particle. As shown in Fig. 2, a 10-fold dilution further reduces the probability that a single cell is infected by multiple particles.
In order to test whether the differences in the plating efficiencies of KOS and 7134 on Vero cell monolayers of various ages were a consequence of the absence of ICPO (and not the LATs) in 7134-infected cells, plating efficiency tests were repeated on 0-28cell monolayers. In these tests, theplatingefficiencies of the two viruses were nearly equal and exhibited no dependence on the age of the monolayer (data not shown; similar results are presented in Fig. 6 below). Therefore, the differences in plating efficiency of 7134 on Vero cell monolayers of various ages is a conse-quence of the absence of the ICPO gene in 7134 and the presence of a plaque-enhancing cellular function able to substitute forICPO.
The activity ofthe plaque-enhancing cellular function cor-responds with the growth status of cells in the monolayer. Because the processoftrypsinization andreseeding induces
avariety of changes in cells which might affect theabilityof 7134 toproduce plaques on Vero cell monolayers, we next attempted toexamine other properties that vary with the age of the monolayerfor their roles in the enhancement of 7134 plating efficiency. One such property is cell density, which increases as afunction oftime postseeding. The number of Vero cells in monolayers in these studies nearly doubles between 8 and 24 h postseeding, such that the cell density
increases significantly during this 16-h period. In order to test the effect of cell density on 7134 plating efficiency we seeded twice the numberofcells (i.e., 5.4 x
105
perdish),and monolayers were infected with 100 PFU of either KOS or7134 at 8 h postseeding. The platingefficiencies on 7134 were approximately the same on 8-h-old monolayers of the twodensities and were, again, twice those ofKOS, demon-strating that cell density is not responsible for the depen-denceof 7134 plating efficiency on the age of the Vero cell monolayer.
Other factors which vary with the age of the monolayer
include the growth status of cells (whether they areactively
cycling or growth is arrested) and changes in the concentra-tions of nutrients or growth factors in the medium. These possibilities are addressed below.
If the cellular function that can substitute for ICPO is associated with the growth statusof cells, then cells such as
Virus titer
(PFU/plate)
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 ICP0-LIKE CELLULAR FUNCTION 4083 those in the monolayers used in Fig. 1 should be at least
partially synchronized with regardtotheir growthstatus. In order to examine this possibility, Vero cells were trypsin-ized, seeded, harvested at various times postseeding, and subjected to flow cytometry to determine the proportionof cells thatwasactivelycyclingandtheproportion that was in growth arrest. The results of these tests indicated that the cellsinmonolayerssimilarto those used to generate thedata showninFig. 1 were indeed partiallysynchronized ingrowth arrest at thetime of seeding and for 7 h thereafter (data not shown). Unfortunately, the extent of synchrony was not sufficient to permit us to identify definitively the relative proportions of growth-arrested and cycling cells at the time of peak plating efficiency (8 h postseeding). These cells were probably synchronized by contactinhibition-trypsinization, since it has been demonstrated by others thatdetachment of cellsby trypsinization delays some cells in G1 from entering S, resulting in partial growth arrest (6). Thus, the depen-dence of 7134 plating efficiency on the ageofthe monolayer postseeding (Fig. 1 and 2) correlated to some extent withthe degreeof synchrony of the cell population caused by growth arrestand release.
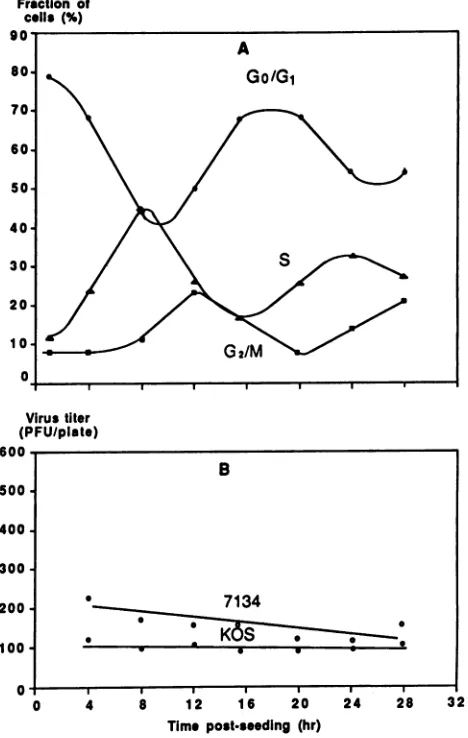
In order to assess the role of growth status in 7134 plating efficiency more definitively, it was necessary touse a more highly synchronized cell population. A higher level of syn-chrony was achieved byisoleucinedeprivation ofVerocells (28, 41). Vero cells were culturedfor 42 hinmediumlacking isoleucine, and then the block was released by cultivating cells in mediumcontaining isoleucine. Cells cultured inthis manner were harvested at varioustimes postrelease, andthe growth status of thepopulation wasassessed byflow cytom-etry (Fig. 3). Immediately beforerelease oftheblock (t = 0), the growth of approximately 85% of cells was arrested and had accumulated in the GJ/G1 phase of the cycle (Fig. 3A). The remainder of the cells occupied the SorG2/Mphasesin approximately equal numbers. Cells in S or G2/M phase could notbe induced to enterGJG1 byincubation forlonger periods of time in isoleucine-deficient medium. Cells in GJG1 moved into the S phase between 8 and 12 h postre-lease and entered G2/M between 16 and 20 h postrelease. When cells were counted at various times postrelease, the majority were shown to have completed cell division by approximately 24 h (Fig. 3B). These cells again entered S phasebetween 24and 28hpostrelease. On the basis of these data, we estimate that growth ofapproximately 85% ofthe cells was arrested by isoleucine deprivation. Moreover, these cells continued to cycle synchronously after being released fromgrowth arrest. Cells synchronized by contact inhibition-trypsinization and cells deprived of isoleucine appeared to be blocked at the same stage in the cell cycle (GJG1); however, isoleucine-deprived cells achieved a far greater degree ofsynchrony.
To determine whether isoleucine-deprived-released cells could provide theplaque-enhancingfunction more efficiently thantrypsinized cells, Vero cellssynchronized by isoleucine deprivation wereinfected with KOS or7134 at various times postrelease fromgrowth arrest (Fig. 3C). Only slight fluctu-ations in theplatingefficiency of KOS were evident inhighly synchronized monolayers through one complete cycle. By contrast, the platingefficiency of 7134reached a peak at 8 h postrelease from growth arrest.
To further examine the effect of trypsinization on the induction of the cellular function, cells synchronized by isoleucine deprivation were trypsinized, reseeded in isoleu-cine-containing medium, and released from the block. A similardegree ofsynchrony was achieved in these tests (data
Fractlonof
cells(%)
Numberof cells(x108) 1.!
1.c
0.I
0.1 0.7
0.1 0.1
Virus tier (PFU/plate)
600I
500
400
300 200
100 0
12 16 20
Timepost-riess (hr)
FIG. 3. Plating efficiencies of 7134 and KOS on Vero cell mono-layers synchronized by isoleucine deprivation. Twenty-four-hour-old Vero cell monolayers were harvested and seeded in 60-mm-diameter petri dishes (5 x 105 cells per dish). Monolayers were maintained in isoleucine-free medium from 24 to 66 h postseeding. Cells were then released from the block by growth in medium containing isoleucine. At various times postrelease, cells were (A) harvested and subjected to analysis by flow cytometry, (B) har-vested to determine the cell number per dish, or(C)infectedwith 40 PFU of7134 or 100 PFU of KOS (input multiplicities of infection were based on titers determined on 24-h-old Vero cells [Table 1]). Forty PFU of 7134 was used to infect each 60-mm dish rather than 100 PFU per 35-mm dish in order to ensure better plaque separation and thus facilitate more accurate plaque counting. These changes were necessary because many more plaques were produced inthis experiment than in the experiment shown in Fig. 1. Plaques were visualized on day 5 postinfection. Plaque numberswere multiplied by 2.5 for comparison with the data shown in Fig. 1, whicharebased on an inoculum of 100 PFU per monolayer.
5
B a
D
-I
-I19 0
r 0 a
0
5---l
5 if I I I I I I I
VOL. 65,1991
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.317.545.99.534.2]Virustiter Fraction of
cells (%)
12 16 20
Time post-seeding (hr)
FIG. 4. Platingefficiencies of 7134 and KOSonVerocell
mono-layers whose growth had been arrested by isoleucine deprivation and trypsinization and then released. Twenty-four-hour-old Vero cellmonolayers were subjectedtoisoleucine deprivation for 42 h. Cellswere then harvested by trypsinization, seeded at 6.3 x 105
cells per 60-mm-diameter dish, and released from the block by
growth in medium containing isoleucine. At various times post-seeding, monolayers were infected and plaques were counted as
described in thelegendtoFig. 3.
notshown), and the plating efficiency of 7134 again peakedat 8 hpostseeding(Fig. 4). The efficiency of 7134 plating under these conditions was sixfold (between four- and sixfold in repeat experiments)greaterthan that observed on
monolay-ers trypsinized and cultured in standard medium (compare
Fig. 1A with Fig. 4). The combination of isoleucine depriva-tion andtrypsinization appeared to havea synergistic effect
on 7134 plating efficiency in this experiment, suggesting the possibilitythattrypsinization andisoleucine deprivationmay
enhance7134 plating efficiency bytwodifferentmechanisms. Trypsinization temporarily arrests cells within G01G1 but notinotherstagesof the cellcycle(6). Inordertodetermine whether the cellular functionwas induced by trypsinization
per se or whether trypsinization must occur at a specific
stage ofthe cell cycle toinducethe cellularfunction, cells deprived of isoleucinewerereleasedfromgrowtharrestand then trypsinized and seeded at 8 rather than at 0 h post-release. The resulting monolayers were then infected at varioustimes postseeding. No obvious enhancement of 7134 plating efficiency was observed on these monolayers, and
theplating efficiency of the mutant was generally low (Fig.
5B), although thecellsexhibitedahigh degree of synchrony
(Fig. 5A). Although asynergistic effectwas observed when cells synchronized by isoleucine deprivation were
trypsin-ized (Fig. 4), this effect was observed only when trypsin-ization occurred at the time of release from growth arrest andnot at8 hpostrelease(late GgG1orpre-S). Therefore,as
notedabove, trypsinizationper sedidnothaveastimulatory
effect on 7134 plating efficiency. In addition, because no
obvious peakof7134plating efficiencywasobserved at 8h postseedingwiththe addition ofnewmedium,we conclude
that changes in the concentrations of medium components
were not responsible for the pattern of expression ofthe cellularplaque-enhancing function.
In all tests,the platingefficiency of7134 wasgreatest on
Verocellmonolayers 8 h after trypsinization and seedingor
on highly synchronized monolayers at 8 hpostreleasefrom
80-10
Virus titer (PFU/pIate) 600
B 500
400 300
-200- * _ _ 7134
100 0
0 , . . . .
0 4 8 1 2 1 6 20 24 28 32
Time post-seeding (hr)
FIG. 5. Plating efficiencies of7134and KOSonVero cell mono-layers releasedfrom the block induced by isoleucinedeprivationat
t = 0 and trypsinized 8 h later. Twenty-four-hour-old Vero cell monolayersweresubjectedtoisoleucinedeprivation for 42 h. Cells were then releasedfrom the block by growth for 8 h in medium containing isoleucine. They were then trypsinized and seeded in isoleucine-containing medium. At various times postseeding, mono-layers were analyzed by flow cytometry (A) or infected (B) as describedin thelegendtoFig. 3.
growtharrestinducedbyisoleucinedeprivation (Fig. 1, 3C, and4). This timepointcorresponds most closelyto late G1 phase(Fig. 3A).Whenhighly synchronizedcellscycledback andagainreachedG1 (atapproximately24 hpostrelease),as judged by flow cytometry, a second major peak of 7134 plaque formationwas notobserved(acompletesecond cycle is shown in Fig. 5). This observation indicates that the cellular plaque-enhancing function exhibits at least one featurewhich is characteristic of factors expressed in cells after release from growth arrest, namely, it is expressed during entryinto the cellcycle butnotwithinthe cell cycle perse. Thus,the marked increase in 7134plating efficiency correspondswith release of thecellpopulationfromgrowth arrest, suggestinganassociation betweengrowth statusand expression of the 7134 plaque-enhancing cellular function. Despite the close correspondence ofthese two properties,
however,acausalrelationshipbetweenrelease from growth A
Go/Gi
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.78.310.75.247.2] [image:7.612.334.568.79.448.2]HSV-1 ICPO-LIKE CELLULAR FUNCTION 4085
Virustiter
(PFU/pbate) 500
400 300 200 100 0
7134
0 4 8 12 16 20
Timepost-release (hr)
FIG. 6. Plating efficiencies of 7134 and KOSon0-28 cell
mono-layers generated by trypsinization of isoleucine-deprived cells and seeding in isoleucine-containing medium. 0-28 cellsgrownfor 42 h in mediumlacking isoleucine were trypsinized and seeded (6.3 x 105
cells per 60-mm-diameter petri dish) in medium containing
isoleu-cine. At various times postseeding, monolayers were infected and
analyzedas described in thelegendtoFig. 3.
arrestand expression of the cellular function remains tobe
proven, asother factors inducedbythesetreatmentsbutnot associated with the growthstatusof cellsmayberesponsible
for theenhancement of7134plating efficiency.
The absence of ICPO is responsible for the differences in plating efficiency of 7134 in isoleucine-deprived cells at dif-ferent times postaddition of isoleucine. In order to test whetherthe differences in the behavior of 7134 and KOS in isoleucine deprived-released cellsare a consequenceof the absence of ICPO in 7134-infected cells, as is the case for trypsinized cells, we tested the plating efficiencies of 7134
and KOS on 0-28 cell monolayers synchronized by isoleu-cine deprivation. In these tests, the plating efficiencies of 7134 and KOS were nearly equal on 0-28 monolayers to which isoleucine-containing medium had been added 4 to28 h previously (Fig. 6). This finding (i) demonstrates that the absence of ICP0 in 7134-infected Vero cells isresponsiblefor the marked difference in the plating efficiency of 7134 on
isoleucine-deprived Vero cellmonolayers at different times postaddition of isoleucine and (ii) confirms the previous observation that the cellularplaque-enhancingfunction sub-stitutes specificallyfor ICP0.
KOS and 7134 initiate infection of isoleucine-deprived-released Vero cells withequal efficiency.Two possible expla-nations for the reducedplating efficiencyof 7134 relative to KOS on 24-h monolayers are that 7134 virions enter these cells less efficiently thanKOS virions or if the two viruses enterwithequal efficiency, that7134virionsareless efficient ininitiatinginfectionleading toplaqueformation. Although
we had shown previously that 7134 was unimpaired in its abilityto enterpartially synchronized Vero cell monolayers 8 and 24 h after trypsinization and seeding, we wished to confirm this observation by using isoleucine deprived-re-leased cells. Therefore, we infected Vero cell monolayers whose growth had been arrested by isoleucine deprivation and then released fromgrowth arrest8 and 24 hpreviously withequal numbers of KOS or7134 PFU. At 8 h postinfec-tion, monolayerswerefixed and thenumberofcells
express-ing ICP4 (an IE protein) was assessed by immunofluores-cence.Themultiplicityofinfectionwas0.03 PFUpercellon
monolayersinfectedat8 hpostreleaseofthe block. Because thesamenumber of PFUwasusedtoinfectmonolayersat8 and 24 h postrelease and because the number of cells had doubled duringthe 16-h interval, the actual multiplicity of infectionon24-hmonolayerswas approximately0.015 PFU percell.
As seenin Table
2,
the numberofcellsexpressing
ICP4 wasnearly
the same as that inmonolayers
infected with either KOS or7134 at8 or24 hpostaddition
ofisoleucine. Recall that anequal
numberof KOS and 7134 PFU(deter-mined on thebasis of assays on 0-28
cells)
reflects thefact that stocks of the two viruses contain a similar number ofgenome-containing particles. Therefore,
KOS and 7134 areequally
efficient in theirability
to enter cells from cultures thatdiffer in the timeof isoleucineadditionby
16h,
and the lowerplating
efficiency
of7134onmonolayers
24 hpostad-dition is not a consequence of cellular factors that exclude
attachmentor
penetration
of 7134particles.
These observa-tions confirm ourprevious
finding
that both viruses wereequally
capable
ofentering
8- and 24-h-old isoleucinede-prived-released
Verocellmonolayers,
whichwas measuredby
thetotal number of genomes detected in infected cells. Viralproteinsynthesis
inisoleucine-deprived-released
Vero cells.Asnotedpreviously,
ICP0is abletoactivateall kineticclasses of HSV-1 promoters tested to date in transient
expression
assays. One would thereforepredict
ageneral
reduction in viral gene
expression
in cells infected with mutant viruseslacking
bothcopies
of the gene for ICPO.Similarly,
if a cellular function can substitute for ICPO to stimulate viralgrowth,
this function should also enhance geneexpression
in cells infected with ICP0 mutants. Ageneral
reduction in viral geneexpression
has indeed been documented for ICP0 mutants.Although
it is difficult todetermine how many and
specifically
which genes areaf-fected in this way
(21,
68, 72),
we haverecently
demon-strated that 7134 and other ICPO mutants exhibit reducedexpression
of E and Lproteins
at lowmultiplicities
ofinfection
(4).
In order to examine viral geneexpression
inisoleucine-deprived-released cells,
we infectedisoleucine-deprived
Verocellmonolayers
at8and24h afteraddition of isoleucine with lowmultiplicities
of infection of KOS and 7134. Inlow-multiplicity-of-infection
experiments,
however,
levels ofviral
protein
synthesis
areminimal,
and individual bands are difficulttoidentify
andquantify against
theback-ground
of cellularprotein
bands. Tocircumvent thisproblem,
weexamined the
synthesis
of viralproteins
whichrepresentspecific
kinetic classesby
immunoprecipitation
of infected cellextractswithamixtureof monoclonal antibodies. Theseproteins
includedICP4(an
IEprotein),
ICP6and ICP8(both
E
proteins),
andICP5,
gB,
andgC (all
Lproteins).
Isoleucine-deprived
Verocellmonolayers
8 and24hafteradditionofisoleucinewereinfectedataverylow
multiplicity
ofinfection
(0.03
PFU percell)
andpulse-labeled
at6or8 hpostinfection,
which was followedby
immunoprecipitation
and
SDS-gel
electrophoresis
of theprecipitated
protein (Fig.
7).
KOS inducedhigh
levelsofEand Lprotein
synthesis
inmonolayers
at both 8 and 24 hpostaddition
of isoleucine(compare
lanes 1 and2with lanes 5 and6).
Incontrast, 7134 induced lower levels of theseproteins
inmonolayers
in-fected at both timespostaddition
of isoleucine.Notably,
protein
levels inmonolayers
to which isoleucine had been added 8 hpreviously
weresignificantly
higher
than those inmonolayers
towhich isoleucine had been added 24 hprevi-ously (compare
lanes 3 and 4 with lanes 7 and8). Thus,
the enhancedexpression
of viralproteins
in 8-hmonolayers
relativeto
expression
in 24-hmonolayers
infected with 7134corresponded
with theresults ofplating efficiency
tests. On the basis of the demonstrated enhancement of viral geneexpression
in Vero cells in 8-hmonolayers,
the cellular function shares with ICP0 the property ofstimulating
theexpression
of viral genes of all kinetic classes.The results of studies of viral
protein
synthesis
as mea-KOSVOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.92.256.74.186.2]infect
at
vi
rus:pulse at
ICP4
-ICP5
-ICP6
-ICP8
-gB
-gC
-8
24
KOS 7134 KOS 7134
6 8 6 8 6 8 6 8
40' m"o4
...
686
686
Is3
ES 41
1 2 3 4 5 6 7 8
FIG. 7. Viral polypeptide synthesis in isoleucine-deprived Vero
cellsas afunction ofthe time of addition of isoleucine. Petri dishes
(diameter, 35 mm)were seededwith 2.7 x 105Vero cellsdeprived
ofisoleucine for 42 h; cellswereseeded in normal growth mediumto
reversetheblock. Monolayerswereinfectedat8or24 hpostseeding witheither KOSor7134.Eight-hourmonolayerswereinfectedata
multiplicity of infection of 0.03 PFU percell (determined on the
basis of virus titers on 0-28 cell monolayers). Twenty-four-hour monolayers were infected with the same number ofPFU of each
virus. Because cell numbers had doubled by 24 h the multiplicity of infection inthesecultureswas0.015 PFUpercell. When double the
number of 8-hcells wasusedand themultiplicity of infectionwas
0.015 PFU per cell, no differences in the levels of viral protein
synthesiswerenoted fromthose of 8-h cellsinfectedat0.03PFUper
cell(data not shown). Infected cultures were pulse-labeled for 20
minat6or8 hpostinfection, and then infected cell extracts were
immunoprecipitated and subjectedtoSDS-gelelectrophoresis.
sured by immunoprecipitationwere confirmed by
immuno-fluorescencetests(Table 2). Thus, incontrast tothe results ofteststhatshowed thatICP4 expressionwas thesamefor
7134 and KOS in monolayers infected 8 and 24 h after addition ofisoleucine, results of this test showed dramatic differences intheability of 7134toinducetheexpression of ICP8 and ICP5 relative to that of KOS in monolayers infectedat both 8and24 hpostadditionand in the ability to induce the expression ofthese proteins in monolayers in-fectedat8versus24hpostaddition. Althoughthenumber of cellsexpressing ICP8wasabouthalfthe number expressing
ICP4whenmonolayerswere infected with KOSat8and24 hpostaddition ofisoleucine, this numberwasnearlyequalin
monolayers of bothtypes(4.2 in 8-hmonolayers and 4.1in 24 hmonolayers). Thesame canbe said for expression ofthe
lateprotein, ICP5,in KOS-infected cells, exceptthatfewer cells in both 8 and24 h monolayers expressed this protein (3.5in8 h monolayers and 3.3 in24h monolayers). Insharp contrast toKOS-infected cells, the number of 7134-infected cells expressing ICP8 and ICP5 was reduced severalfold
when8-hmonolayerswereinfected(ICP8, 0.79; ICP5, 0.21).
In monolayers infected with 7134 24 h after addition of isoleucine, the numberwas even smaller(ICP8, 0.04;ICP5, <0.04). Cells expressing low levels ofICP8 and ICP5 may
wellhaveescapeddetection because of the low sensitivity of thefluorescence assay. Nevertheless, these results
demon-h ps
stratethatisoleucine-deprived
cells to which isoleucine had been added 8 h previously express a function which, likeICPO,is able toenchance expression of E and L
proteins.
The dataon plating efficiency presented in previous sec-hP1| tions suggest that anICPO-like cellular function isexpressed
atmaximallevels inmonolayers 8 h aftertrypsinization and seeding and 8 h after addition of isoleucine to isoleucine-deprived cells. Since plaque formation involves several rounds ofvirusreplication in many cells, itwasimportantto demonstrate that the increase in 7134 platingefficiencywas the result of a cellular function expressed during the first round ofreplication and notthe result of events thatoccur during subsequent rounds of replication in neighboring cells that may differ in growth status or other properties when infected. Theprotein synthesis assays fulfill thisrequirement
because protein synthesis was measured during the first round ofreplication.
DISCUSSION
Virus replication involves complex interactions between viral and cellular proteins. Because susceptible cells may vary with respect to their physiological conditions at the time of infection and because variations in these conditions arecharacterized by differences in the kinds and concentra-tions of proteins that the cells contain, viruses must be capable ofinitiating infection despite these differences in the intracellular environment. The fact that smaller,genetically less complex viruses are moredependent for their replica-tion on the funcreplica-tions of cellular proteins than are larger, more complex viruses has been recognized for some time. This is reflected in the fact that the genomes oflarger viruses have acquired homologs of critical cellular genes, which permits them toreplicate in cells not expressing these genes. Forexample, cellular proteins involved in DNA precursor metabolism are expressed maximally immediately before and during the S phase of the cell cycle. Levels of these proteins are low in other stages of the cell cycle and are especially low in growth-arrested cells (57). HSV-1 encodes avariety of proteins involved in DNA precursor metabolism such as thymidine kinase(TK) and ribonucleotide reductase which permit the virus to replicate in cells that do not express adequate levels of the cellular counterparts of these proteins.Thus,thefact that HSV-1carriesits own genes for TK andribonucleotide reductase permits the virus to repli-catein cells irrespective of their growth status.
In the studies described herein we have presented evi-dence for the existence of a cellular function which is expressedathighlevels inisoleucine-deprived cells 8 h after addition ofisoleucineorincells 8 haftertrypsinizationand which can substitute for ICPO, a major HSV-1 transactivator of viral gene expression. The effectsof this function onthe replication competence of 7134 raise the possibility that cellular factors thatperform this functionor asubset of these factors may be involved in productive infection and in the reactivation of HSV from latency.
Properties of the ICPO-like cellular function. Our results demonstrate(i)thatamutantviruslackingbothcopies of the ICPO gene is impaired in its abilityto form plaques and to induce expression of E and L viral proteins in Vero cell
monolayers24haftertrypsinizationandreseedingor
isoleu-cinedeprivation and release and (ii) that thisdefect canbe
compensated for in monolayers 8 hafter thesetreatments.
Thus,acellularfunctionexpressed maximally in 8-h
mono-layers cansubstitute forICPO. Wedesignatethisan "ICPO-like" function. Mechanistically, the cellular function may
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.75.296.71.282.2]HSV-1 ICP0-LIKE CELLULAR FUNCTION 4087 represent (i) an ICPO homolog, (ii) a component(s) of the
cascade through which ICPO operates, e.g., an
ICPO-acti-vated factor which in turn activates expression of other genes, or (iii) a transactivator whose activity is inhibited at most stages of the cell cycle (e.g., 24 h) but not shortly after release from growth arrest (8 h). The last two possibilities raise the question of whether the cellularfunction is specific. We have shown that the cellular function enhanced the plating efficiency and viral protein expression of 7134 but not of KOS and that its effect on 7134 was abolished whenICPO
was provided in trans in 0-28 cells. These findings argue that the cellular function can substitute specifically for ICPO
regardless of its mechanism of action.
The conclusion that the cellular function is able to substi-tute forICPOis based on two criteria: enhancement of 7134 plaque formation (complementation) and protein synthesis. Complementation is an especially informative criterion be-cause it assesses the cellular function in the context of the entire family of HSV-1 genes. Because the cellularfunction complemented 7134 plaque formation efficiently, it can by definition transactivate the same viral genes as ICPO, a conclusion which is supported by the ability of the function to enhance the expression ofrepresentative HSV-1 proteins of E and L kinetic classes. The ability of the cellularfunction to stimulate expression of all selected E and L viral genes whose expression isstimulated byICPOfurthersupports the notion that it can substitute specifically forICPO.
The observation that peak expression of the ICPO-like cellular function occurred at 8 hpostseeding in monolayers prepared from trypsinized cells and in monolayers 8 h postrelease from growth arrest induced by isoleucine depri-vation indicates that bothprocedures block cells in the same or a similar stage of the cell cycle. The finding that the trypsinization at 0 h (earlyGJG1) but not at 8 h (lateGJG1) induces the ICPO-like function strongly suggests (but by no means proves) its association with the cell cycle. The results of flow cytometric analysis and quantitation of cells in the monolayer demonstrate that the growth of Vero cells syn-chronized by isoleucine deprivation is arrested in theG01G1
stage of the cell cycle, which is consistent with findings in other cell types (28, 41). Although these reports suggest that isoleucine deprivation blocks cells inG1 phase, the evidence presented is alsoconsistent with the possibility that the cells are blocked at a stageexhibiting some characteristics of
Go.
Unfortunately, it is not clear from our studieswhether Vero cells are arrested in
Go
orG1 or what stage of the cell cycle precisely corresponds to 8 h postrelease of the block induced by isoleucine deprivation. The failure of growth arrested-released cells to provide the enhancing function as cells traverse G1 of the second cycle would argue that the block is in or nearGo
and that the ICPO-like cellular function is expressed maximally during entry into the cell cycle. Con-siderableprecedent exists for such a pattern of cellular gene expression. In mouse cells, for example, many IE cDNA clones have been isolated that represent cellular genes expressed only transiently following stimulation by serum or growth factors but not within the cell cycle itself (7, 8, 11, 27, 35, 37, 40, 66, 73). Some of these transiently expressed proteins include cellular transcription factors, such as c-Fos, c-Jun, c-Myc, and Krox-20. These proteins are purported to initiate expression of subsequent kinetic classes of cellular genes which themselves might be transiently expressed. If indeed the ICPO-like cellular function is expressed tran-siently as a function of the cell cycle, it would not be expected to be detected at high levels in highlysynchronizedcells that have not experienced growth arrest. We are currently investigating this possibility.
Roles of ICPO and the ICPO-like cellular function in pro-ductiveinfection and latency. (i) Productive infection.ICPO is known to play animportant butnonessential role in
produc-tiveinfection because growth ofICPO mutantsisimpairedin standard monolayers at low multiplicities of infection. Be-cause of the rather vaguephenotype of ICPOmutants (i.e.,
they are replication competent yet exhibit reduced expres-sionof E and L genes and reduced synthesis of infectious virus), the precise role of ICPO invirusreplication has been difficult to assess. Theresultspresented in this reportclarify
much of the confusion concerning the function ofICPO in productive infection bydefining therelationshipbetweenan ICPO mutant and the host cells itinfects, in this case, Vero cells. Cells canbeclassified as eitherpermissive or nonper-missive in terms of their ability to support the growth of ICPO mutants. Permissive cells support mutantgrowth be-cause they express afunction that can substitute for ICPO. These cellsexpress the cellularfunctionatmaximal levels 8 h aftertrypsinization andreseeding and 8 h after addition of isoleucine to deprived cells. Cells not subjected to these treatments are less permissive for growth of ICPO null mutants unless they are infected at high multiplicities of infection (21, 68, 72). A high multiplicity of infection may confer permissivity by inducing expression of the cellular function. In contrast toICPO mutants, thewild-typevirus is able to express ICPO and can therefore replicate in cells regardlessof whetherthey express the cellular function
(i.e.,
irrespectiveof their growthstatus). In this way, the
ability
of HSV-1 to express ICPOconfers a significant growth advan-tage to the virus.(ii) Latency. The significance of the fact that the HSV-1 genomecarries two genesfor the nonessential ICPO function may indicate that many cells that the virus encounters in nature do not express this function. This
hypothesis
is especially relevant to the problem oflatency
and reactiva-tion. HSV-1 establishes latent infections in neuronal cells which are in a growth arrested, quiescent state. Since the pattern of expression of the ICPO-like function resembles that of IE cellular transcription factors after serum stimula-tion and since these proteins are not expressed inquiescent
cells, the ICPO-like function would not be
expected
to be expressed in such neuronalcells. If ICPOplays
animportant
roleinreactivation fromlatencyashas been
postulated
(39),
suppression of ICPO expression, as well as of the cellular ICPO-like cellular function and other IE
transactivating
proteins (ICP4 andICP27), would assist the virus in
entering
the latent state and thus prevent productive infection asso-ciated withreactivation. A role for ICPOinreactivation was initially postulated on the basis of two observations:
(i)
deletion of the ICPO gene results ina
significant
decrease in the ability of latently infected mouseganglia
toproduce
progenyvirusfollowingexplantation(i.e.,
toreactivate from latency) (4, 9,39)and(ii) ICPO-deficient andnullmutantsfail toreactivate inanin vitro system (65).Notably,
ICPO alonein the background ofan adenovirus vector was sufficientto inducereactivation in this in vitro model
(29, 76).
Assuming
thatreactivationconstitutesa
resumption
ofinfectionin that IE gene expression precedes and isrequired
for E and L gene expression, it is unclear whether the role of ICPO in reactivation is to enhance E and L geneexpression
or to enhance IE, E, and L geneexpression.
During
productive
infection, ICPO is thought to be
required
for E and L gene expression but not for IE geneexpression (4)
(Fig.
7 and Table 2) because IE genesare activatedby
thevirionprotein
VOL.65, 1991