Inducing Scale Insect
Apiomorpha
Riibsaamen
(Hemiptera: Sternorrhyncha: Coccoidea).
Lynette Gai Cook
Division of Botany and Zoology
The Australian National University
Canberra, ACT, Australia
A thesis submitted for the degree of Doctor of Philosophy of The Australian National University
FRONTISPIECE
.. 11
Declaration:
The research and text presented in this thesis is the original work of the author, except where acknowledged.
ACKNOWLEDGMENTS
Firstly, I would like to thank my supervisors Penny Gullan and Dave Rowell for support and guidance throughout my postgraduate studies. I am indebted to Penny for stimulating and sustaining my interest in scale insects. She was generous with her time and willingly shared her considerable expertise while assisting with most aspects of this study. I would like to thank Dave for tutelage in allozyme electrophoresis and his enthusiasm and capability in all things chromosomal.
Thanks to Mike Crisp for assistance with eucalypt identification and I apologise for some of the poor material he was given to work with. Ian Scott was an excellent instructor in molecular techniques - it was not his fault that it took six months to get the first DNA sequences from Apiomorpha. Assistance from other staff and students in the lab has been much appreciated. Jeremy Brownlie (BOZO and CSIRO Entomology) searched Apiomorpha DNA for transposable elements.
I am grateful for the critical reviews of my 1nanuscripts provided by my supervisors and others, especially Peter Cranston. I have enjoyed the intellectual stimulation gained from the Coopers and Cladistics discussion group, the Calypso coffee group - past and present (David, Hanna, Holly, Jim, John, Mike, Scot), behavioural ecology discussion group and EES postgraduate discussion groups.
The Division of Botany and Zoology provided most of the facilities used and many members of staff have provided advice and technical support. I would especially like to thank Keith Herbert (photographies), Aimorn Stewart (SEM and slide preparation) and Alan Muir (workshop). I'd also like to thank the staff of the Plant Culture Facility and the SEM Unit of the ANU, especially Roger Heady. The CSIRO Division of Entomology provided a depository for galls of Apiomorpha.
lV geographic localities of collections used in this project was made possible by the many colleagues who recognised and collected Apiomorpha during the course of their work or when on holidays. Penny and Peter provided many collections of Apiomorpha.
This project was partially funded by an Australian Postgraduate Research Award with addition monies from Penny Gullan's research pool. Some of the molecular work was partially funded by a small ARC grant to Penny Gullan and Dave Rowell. I appreciate the opportunity to have attended several overseas conferences through funding by the Michael White Travel Award (ANU) and national conferences with financial assistance from the Division of Botany and Zoology.
ABSTRACT
Apiomorpha is an unusual genus of gall-inducing scale insects. Both males and females
induce galls on Eucalyptus - the galls of females of some species are spectacular and amongst the largest induced by gall-inducing insects. Adults of Apiomorpha are highly sexually dimorphic and this is reflected in the relative size and shape of their galls. Galls of males are typically small and tubular whereas those of females are larger and usually have species-specific morphologies.
The highly derived morphology of adult females, probably a result of their gall-inducing habit, has meant that the phylogenetic affinities of Apiomorpha have been unclear. In this study, DNA sequence data were used to generate a phylogeny of scale insects including Apiomorpha and eriococcid taxa to which it may be related. The molecular data do not provide support for a monophy letic Eriococcidae and there is strong evidence that Eriococcus sensu lato is polyphyletic. Apiomorpha appeared to be most closely related to a clade of non-gall-inducing eriococcids that includes some eucalypt-feeding Australian eriococcids, such as Phacelococcus. Several morphological features of immature instars were found to support this relationship. The DNA sequence data suggest that gall-induction has probably evolved at least three times amongst eriococcid taxa and that other Australian gall-inducers on Eucalyptus are not closely related to
Apiomorpha.
Vl mate predominantly with females on their natal plant. A female may also mate with her
male descendants because of an overlap in generations between a mother and her
offspring. High levels of inbreeding are likely.
Reproduction in Apiomorpha was found to be primarily sexual because males are known
for most species and sperm was found in the reproductive tracts of most gravid females
examined. The one apparent exception is an unusual undescribed taxon from
Queensland for which no males or sperm were found, despite the presence of embryos. Males of all species of Apiomorpha examined had a Comstockiella form of chromosome
loss prior to meiosis. Heterochromatisation of the paternally inherited chromosomes in
hybrid male embryos of one female supports the hypothesis that it is the paternal
genome that is eliminated in scale insects. Females of Apiomorpha appeared to have
typical chiasmate meiosis, with one chiasma per bivalent. Reports that chiasmata are
terminalised prior to metaphase I in scale insects were not supported in Apiomorpha. In
addition, the observed orientation of bivalents perpendicularly to the metaphase plate
suggests that the first division of meiosis may be reductional and therefore that meiosis
in at least some scale insects may not be inverted relative to that in most organisms.
The range of chromosome numbers found in Apiomorpha (2n=4 to 2n = about 192)
exceeds that reported for most other animal genera. There is extensive chromosomal
variation within and among most morphologically defined species of Apiomorpha,
suggesting that cryptic species complexes are present. Allozyme electrophoresis was
used to delineate biological species boundaries for several taxa in which chromosomal
variation was detected in conjunction with some morphological differences. In each
case, taxa that exhibited morphological differentiation were found to be distinct
biological species. In contrast, different chromosomal forms within some taxa had
identical allozyme profiles. These data indicate that Apiomorpha may be more speciose
than previously reported.
It is argued that high levels of inbreeding combined with a chromosome system in which
males pass on only their maternally inherited chromosomes have enhanced the rate of
n1utations in Apiomorpha may be high relative to that in most other animals. It is suggested that the activity of transposable elements (reported in one species) and fragmentation at fragile sites such as rRNA gene clusters may have contributed to increased rates of chromosomal rearrangement in Apiomorpha. The nuclear small subunit ribosomal rRNA gene of some Apiomorpha taxa was found to be longer than that reported for any other organism.
In this study, an examination of host associations of Apiomorpha taxa revealed an hierarchical pattern of host specificity between the insect and its host. Apiomorpha is restricted to Eucalyptus sensu stricto and each species-group of Apiomorpha is restricted to a single informal eucalypt subgenus. Lower-level taxa of Apiomorpha are restricted to eucalypt sections, series or species. The pattern of associations between Apiomorpha taxa and their Eucalyptus hosts suggests that the two may be cospeciating. The relationships among the eucalypt taxa on which Apiomorpha taxa occur have not been fully elucidated and therefore hypotheses of cospeciation were not tested.
At some locations, different chromosomal forms of the one morphologically defined taxon of Apiomorpha were specific to different host taxa. The association of chromosomal differentiation with different host use suggests that chromosomal rearrangements may be playing an active role in taxon divergence in Apiomorpha. A
TABLE OF CONTENTS
Declaration ... 11 ..
Acknowledgments ... iii
Abstract ... V Table of Contents ... viii
List of Tables ... xiv
List of Figures ... xv
Chapter 1. General Introduction ... 1
1.1. Scale insects in general ... 2
1.2. Apiomorpha ... 5
1.2.1. Galls ... 5
1.2.2. Adult morphology ... 6
1.2.3. Systematics ... 7
1.2.4. Geographic distribution ... 8
1.3. Scope of the present study ... 9
Chapter 2. The Systematic Placement of Apiomorpha ... 12
2.1. Introduction ... 13
2.2. Methods ......... 16
2.2.1. Specimens and DNA extraction ... 16
2.2.2. PCR and sequencing ... 16
2.2.3. Analysis ... 18
2.2.3 .1. Editing and alignment. ... 18
2.2.4. Phylogenetic analysis ... 19
2.2.4.1. Maximum parsimony ... 19
2.2.4.2. Neighbor-Joining ... 20
2.2.4.3. Maximum Likelihood ... 21
2.2.4.4. Statistical tests ... 21
2.2.5. Length of ssu rRNA ... 21
2.3. Results ... 22
2.3.1. ssu rDNA ... 22
2.3.1.1. Sequence variation ... 22
2.3 .1.2. Length of ssu rDNA sequence using primers 2880 and B- ... 22
2.3.1.3. Inferred phylogeny ... 23
2.3.2. corr ... 24
2.3.2.1. Sequence variation ... 24
2.3.2.2. Inferred phylogeny ... 25
2.3.3. Combined data ... 25
2.4. Discussion ... 26
2.4.1. Systematics of the Cocco idea ... 26
2.4.2. Systematics of the Eriococcidae ... 26
2.4.3. Systematic placement of Apiomorpha ..................... 29
2.4.4. Origins of gall induction among eriococcid taxa ... 31
2.4.5. Length of ssu rRNA ... 32
2.5. Summary ... 32
Chapter 3. Life History and Morphology ... 42
3 .1. Introduction ... 43
3 .2. Methods ... 46
3 .2.1. Collection ... 46
3.2.2. Rearing ... 47
3.2.3. Life history and biology ... 47
3.2.4. Morphology of crawlers ... 48
3.2.5. Sex determination ... 49
3 .3. Results ... 50
3 .3 .1. Life history and biology ... 50
3.3.3.1. Secondary sex ratios ... 52
3 .3 .3 .2. Parasitism, predation and ants ... 53
3.3.2. Morphology ... 53
3.3.2.1. Crawler morphology ... 54
3 .3 .2.2. Sexual dimorphism among crawlers ... 56
3. 3.2.2.1. A. pharetrata species-group ...... 57
3.3.2.2.2. A. munita species-group ... 57
3.4. Discussion ... 59
3.4.1. Life history and biology ... 59
X
3 .4.2.1. Anal lobes ... 60
3.4.2.2. Crawlers ... 61
3.4.2.3. Sexual dimorphism among crawlers ... 64
3.5. Summary ... 67
Chapter 4. Chromosome Number Variation in Apiomorpha ... 76
4.1. Introduction ... 77
4.2. Methods ... 79
4.2.1. Specimens and tissue ... 79
4.2.2. Chromosome preparations ... 80
4.2.2.1. Photographs ... 80
4.2.2.2. Nucleolar organiser regions ... 81
4.2.2.3. C-banding ... 81
4.2.3. Chromosome mapping and evolution ... 81
4.2.4. Crosses between different chromosomal forms of A. pharetrata ................ 82
4.3. Results ... 82
4.3 .1. Chromosome numbers in Apiomorpha ... 82
4.3.2. Intraspecific chromosomal variation (variable species only) ... 83
4.3 .3. Nucleolar Organiser Regions (NO Rs) ... 89
4.3.4. Hybrid matings ... 89
4.3.5. Chromosome numbers in Eriococcidae and so1ne other scale insects ... 90
4.3.6. Chromosome size and evolution ... 90
4.4. Discussion ... 91
4.4.1. Chromosome number and karyotype evolution ... 91
4.4.2. Factors possibly contributing to chromosomal variation ... 95
4.4.3. Chromosome mediated speciation? ... 98
4.4.4. Intraspecific chromosomal variation - taxonomic in1plications ... 100
4.5. Summary ... 102
Chapter 5. Sex, Meiosis and Chromosome Systems in Apiomorpha ... 119
5.1. Introduction ... , ... 120
5.2. Methods ... 124
5 .2.1. Chromosome preparations ... 124
5.2.3. Evolution of chromosome systems ... 124
5.2.4. Sex ratio among embryos ... 124
5.3. Results ... 125
5.3.1. Chromosome systems ... 125
5.3.1.1. Heterochromatic bodies ... 125
5.3.1.2. Male meiosis ... 125
5.3.1.3. Female meiosis ... 126
5.3.1.4. Presence of sperm ... 127
5.3.2. Evolution of chromosome systems ... 128
5.3.3. Sex ratio ... 128
5.4. Discussion ... 129
5.4.1. Reproduction ... 129
5.4.1.1. Modes of reproduction ... 129
5.4.1.2. Sex ratio ... 131
5.4.2. Female meiosis ... 132
5.4.3. Male meiosis ... 134
5.4.4. Evolution of chromosome systems ... 135
5.4.5. Consequences of chromosome system ... 136
5.4.5.1. Fixation or maintenance ofkaryotypic forms ... 136
5.4.5.2. Recombination rates ... 137
5.5. Summary ... 138
Chapter 6. Allozyme electrophoresis and species delineation in Apiomorpha .... 156
6.1. Introduction ... 157
6.2. Methods ... 158
6.3. Results ... 159
6.3.l. Apiomorpha minor .................................................... 160
6.3.2. Apiomorpha munita ... 160
6.3.3. Apiomorpha pharetrata ... 161
6.3.4. Apiomorpha pileata ... 162
6.3.5. Apiomorpha strombylosa ... 162
6.4. Discussion ... 162
6.4. l. Apiomorpha minor .............................................................. 162
XU
6.4.3. Apiomorpha pharetrata ... 164
6.4.4. Apiomorpha pileata ... 164
6.4.5. Apiomorpha strombylosa ... 164
6.4.6. General ... 165
6.5. Summary ... 166
Chapter 7. Host Associations of Apiomorpha ... 17 4 7 .1. Introduction ... 17 5 7 .2. Methods ... 177
7 .2.1 Collection and host identification ... 177
7.2.2. Glasshouse hosts ... 178
7.3. Results ... 179
7 .3 .1. Host associations ... 179
7 .3 .1.1. Apiomorpha malleeacola .......................... 179
7.3.1.2 Apiomorpha minor species-group ... 179
7.3.1.3. Apiomorpha munita ... 180
7 .3 .1 .4. Apiomorpha pharetrata ... 181
7 .3 .1.5. Apiomorpha strombylosa species-group ... 181
7 .3 .1.6. Other patterns of host use ... 181
7.3.2. Glasshouse hosts ... 182
7.4. Discussion ... 183
7.4.1. Host specificity ... 183
7.4.1.1. Apiomorpha minor species-group ... 183
7.4.1.2. Apiomorpha munita ...... 184
7.4.1.3. Apiomorpha pharetrata ... 185
7.4.1.4. Apiomorpha strombylosa species-group ... 186
7.4 .1. 5. Other patterns of host use ... 187
7.4.2. Coevolution, cospeciation, host-tracking or host-switching? ... 187
7.5. Summary ... 191
Chapter 8. Modes of Speciation in Apiomorpha ... 197
8.1. Modes of speciation in Apiomorpha ................................................. 198
8.1.1. Allopatric distributions ... 198
8.1.1.2. Model: cospeciation in allopatry ... 200
8.1.2. Sympatric distributions ... 202
8.1.2.1. Different chromosomal forms of Apiomorpha in sympatry ... 202
8.1.2.2. Cospeciation in allopatry with secondary sympatry ... 203
8.1.3. Alternative models of speciation in Apiomorpha ... 205
8.1.3.1. Alternative models ... 205
8.1.3.2. Comparing and testing the models ... 206
8.1.4. Applicability of the extended model to other taxa ... 207
8.2. Conclusions and summary ... 208
.
XlV
LIST OF TABLES
Chapter 2. The Systematic Placement of Apiomorpha Based on DNA Sequence Data
Table 2.1 Scale insects used for DNA sequencing ... 34
Table 2.2. Pairwise genetic distances ... 35
Table 2.3. Base composition ... 36
Chapter 3. Life History and Morphology Table 3 .1. Species reared in the glasshouse ... 69
Table 3 .2. Sexual dimorphism ... 70
Chapter 4. Chromosome Number Variation in Apiomorpha Table 4.1. Diploid chromosome counts in Apiomorpha ... 104
Table 4.2. Chromosome numbers in some other scale insects ... 107
Chapter 5. Sex, Meiosis and Chromosome Systems Table 5.1. Heterochromatic chromosomes ... 139
Table 5.2. Presence of sperm ... 140
Table 5.3. Sex ratios ....... 141
Chapter 6. Allozyme electrophoresis and species delineation in Apiomorpha Table 6.1. Allozyme systems trialed ... 167
Table 6.2 Apiomorpha minor ........... 168
Table 6.3. Apiomorpha munita ...... 169
Table 6.4. Apiomorpha pharetrata ........... 170
Table 6.5. Apiomorpha pileata ... 171
Table 6.6. Apiomorpha strombylosa ... ... 172
Chapter 7. Host Associations of Apiomorpha Table 7 .1. Host associations of Apiomorpha ....... 193
LIST OF FIGURES
Chapter 1. General Introduction
Figure 1.1. Apiomorpha ... ............................. 11
Chapter 2. The Systematic Placement of Apiomorpha Based on DNA Sequence Data Figure 2.1. Length of ssu rDNA ... 37
Figure 2.2. Strict consensus of MP tree derived from ssu rDNA ... 38
Figure 2.3. Transitions versus transversions ... 39
Figure 2.4. NJ phylogram derived fr01n COII sequences ... 40
Figure 2.5. Single MP tree derived from combined data ... 41
Chapter 3. Life History and Morphology Figure 3 .1. Life history ... 71
Figure 3 .2. Anal lobes ... 72
Figure 3.3. Crawler morphology ... 73
Figure 3.4. Crawlers ... 74
Figure 3.5. Antenna! pegs ... 75
Chapter 4. Chromosome Number Variation in Apiomorpha Figure 4.1. Mitotic metaphases ... 108
Figure 4.2. Mitotic metaphases ... 109
Figure 4.3. Diploid complements in A. minor ... 110
Figure 4.4. Diploid complements in A. munita tereticornuta ........... 111
Figure 4.5. Diploid complements in A. munita malleensis ...... 112
Figure 4.6. Diploid complements in A. munita munita ................. 113
Figure 4. 7. Nucleolar organising regions ... 114
Figure 4.8. Geographic ranges of A. munita subspecies ... 115
Figure 4.9. Geographic distribution of karyotypes of A. munita ... 116
Figure 4.10. Frequency distributions of chromosome complements ... 117
XVI
Chapter 5. Sex, Meiosis and Chromosome Systems
Figure 5 .1. Inverse meiosis ... 14 2
Figure 5.2. Heterochr01natic chromosomes ... 143
Figure 5.3. Chromosome loss ... 144
Figure 5 .4. Spermatogenesis ... 145
Figure 5. 5. Chromosome elimination in A. pileata ... ... 146
Figure 5.6. Diplotene/diakinesis in A. variabilis ... 147
Figure 5.7. Meiosis in a female of A. pharetrata ... 148
Figure 5.8. Meiosis in a female of A. strombylosa .................................................. 149
Figure 5.9. Chiasma position ... 150
Figure 5 .10. Meiosis in females of Apiomorpha .. ... 151
Figure 5 .11. Meiosis in females of A. munita tereticornuta ........................................... 152
Figure 5.12. Sperm in reproductive tract of A. regularis ... 153
Figure 5.13. Elongating "sperm" in reproductive tract of A. regularis .... 154
Figure 5.14. Evolution of chromosome systems in scale insects ... 155
Chapter 6. Allozyme electrophoresis and species delineation in Apiomorpha Figure 6.1. UPGMA dendrogram of genetic distance in A. munita ... 173
Chapter 7. Host associations of Apiomorpha Figure 7.1. Phylogenies of A. munita and Symphyomyrtus ... 196
Chapter 8. Modes of Speciation in Apiomorpha Figure 8.1. Model of cospeciation in allopatry with secondary sympatry ... 211
CHAPTER 1.
"A day before, the scale insects had two tarsal claws and their mates were flying
creatures; thus they were not yet scale insects. The mysterious events of that day are completely veiled by our ignorance; we even cannot date the day. However, the scale
insects of the day after can be the subject of scientific research ... "
Jan Koteja ( 1996, p. 65) referring to the sudden appearance in the fossil record of a disparate
range of scale insect taxa, some nearly identical to extant species.
1.1. Scale insects in general
2
Scale insects (Hemiptera: Sternorrhyncha: Coccoidea) are sap-sucking obligatory plant
parasites and are one of the most diverse and unusual groups of animals ( e.g., Miller and
Kosztarab, 1979; Gullan and Kosztarab, 1997). Scale insects exhibit the greatest range
of reproductive modes known for any one animal group (Nur, 1980). Development
differs among the sexes, and males and females are highly sexually dimorphic (Danzig,
1980). Coccoidea are known from all major terrestrial plant groups and are found in all
terrestrial zoogeographical regions except Antarctica (Miller and Kosztarab, 1979;
Williams, 1984).
Despite the potential for scale insects to be used as model organisms for exploring some
possibly unique evolutionary, biogeographical and speciation-related questions, the
ecology, biology and host relationships of most scale insects have been little studied.
The majority of scale insect studies have focussed on taxonomy, or on the economic
impact and control of pest species. The unusual chromosome systems, sex determination
and reproductive modes of some coccoids also have been reported. While a sound
systematic framework is essential as a basis for other biological studies, there has been little extension of earlier work.
There are estimated to be more than 7500 species of coccoids (Ben-Dov et al., 2000)
assigned variously to twenty or more families (Koteja, 1974a; Gullan and Kosztarab
(=Orthezioidea) and the neococcoids (=Coccoidea s. str.) (Borchsenius 1950; Koteja,
1974a, 1996; Danzig, 1980; Miller, 1984a; Kosztarab and Kozar, 1988; Kosztarab,
1996). The archaeococcoids comprise the Margarodidae ( as in Morrison, 1928), the
Ortheziidae, sometimes also the monotypic Phenacoleachiidae (Danzig, 1980; Koteja,
1996) and, if it is recognised as a separate family, also the Carayonemidae
(Richard, 1986; Kosztarab, 1996). Most of the morphological characters that define the
archaeococcoids (abdominal spiracles, compound eyes in males, and an XX-XO
chromosome system) are plesiomorphies that occur more widely in the Hemiptera and
thus paraphyly of archaeococcoids is likely (e.g., Foldi, 1997; Cook, Gullan and
Trueman, unpubl.).
The neococcoids, which comprise all of the other families and most of the species of
scale insects, are generally treated as a monophyletic group. It has been defined by
synapomorphies such as a chromosome system involving paternal genome elimination
(Nur, 1980; Danzig, 1980), needle-like apical setae on the labium (Koteja, 1974a,b,
1996), shared structural and developmental features of the ovaries (Szklarzewicz, 1998),
and the absence of abdominal spiracles (Morrison, 1928).
Scale insects are a geologically old group. Although the coccoid fossil record extends
only to the Lower Cretaceous (Koteja, 1990, 2000), the scale insect clade must be the
same age as its sister group. It is generally agreed, on the basis of morphology (Carver et
al., 1991) and DNA sequence data (Campbell et al., 1994, 1995a,b; Dohlen and Moran,
1995), that the aphids are sister to the scale insects. Aphids appear first in the fossil
record in the Middle-Upper Triassic at about 235-225 Ma (Shcherbakov and Wegierek,
1991 ). However, the diversity of wing features found in the two Triassic fossils suggests
that aphids evolved much earlier (Shcherbakov and Wegierek, 1991 ), perhaps as early as
the Permian as proposed by Heie (1999).
The taxonomy of scale insects has traditionally been based on morphological features of
adult females, probably as a result of their greater longevity and relative
conspicuousness compared with adult males or immature instars. Additionally, because
4
rare or are not produced at all (Boratynski and Davies, 1971; Danzig, 1980).
Nevertheless, males and immature instars of both sexes are important for systematic
studies and pest identification ( e.g., Giliomee, 1995; Hodgson and Henderson, 1996;
Williams and Miller, 1999; Gullan, 2000).
Adult scale insects display marked sexual dimorphism. In general, adult males are
typically small and winged (mesothoracic pair only) and are capable of weak flight
(Ghauri, 1962; Boratynski and Davies, 1971 ). They have eyes, compound in
archaeococcoids but simple in most neococcoids (Giliomee, 1995), and well-developed
antennae. The abdomen is typically slender but its length varies considerably among
taxa dependent on the configuration of mating ( e.g., Gullan, 1978, 1998) or its other uses
(e.g., phoretic transport of female crawlers (Gullan and Cockburn, 1986)). Males do
not
feed after their second instar and adult males have no mouthparts.
Female coccoids are wingless and those of most taxa have much reduced legs, antennae
and eyes (e.g., Miller and Kosztarab, 1979; Beardsley, 1984). Adult females of some
species have lost all appendages, having no legs and with the antennae reduced to
patches of sensory setae ( e.g., Gullan and Kosztarab, 1997). In contrast to the general
reduction of features, the cuticle of adult females typically has a variety of
well-developed ducts, pores and other glandular organs, and commonly has specialised setae
(Foldi, 1997; Gullan and Kosztarab, 1997).
The reduced and much modified morphology of adult female coccoids is presumably a
consequence of their highly specialised parasitic lifestyles (Danzig, 1984). In general,
adult females are much less mobile than first-instar nymphs (crawlers) or adult males,
and those of many species are sessile. Females of some taxa feed only from roots or are
found exclusively in soil and leaf-litter samples ( e.g., Morrison, 1928; Gullan, 1998;
Kozar and Miller, 2000). Females of many families produce specialised coverings of
waxes, resins and/or discarded cuticles of immature stages (reviewed in Gullan and
Kosztarab, 1997) whereas others induce galls ( e.g., Gullan, 1984c; Beardsley, 1984;
Australia has a diverse scale insect fauna and a disproportionately high number of gall
inducing taxa, primarily belonging to the traditionally recognised family Eriococcidae
(Gullan, 1984c; Beardsley, 1984). Because the relationships among Australian
inducing coccoid taxa are not clear, it is not known whether the prevalence of
gall-induction is due to a radiation in a single lineage or separate origins in numerous
different lineages. The variation in the complexity and morphology of galls induced by
different taxa suggests that there may have been more than one origin of gall formation
among the Australian eriococcids. The most simple gall form is a pit, such as that
induced by some species of Lachnodius and Opisthoscelis (Froggatt, 1921 a,b;
Beardsley, 1984). The modified bract-galls induced on Allocasuarina by males and
females of Cylindrococcus are also relatively simple (Gullan, 1984b ). The most complex
eriococcid-induced galls are those of females of Cystococcus (Beardsley, 1984; Gullan
and Cockburn, 1986) and Apiomorpha (Kuster, 1937; Beardsley, 1984; Gullan, 1984a).
Apiomorpha is the most speciose genus of strictly gall-inducing coccoids and is unusual
in respect of the morphology of galls induced by females, the size of females of some
taxa, and host specificity. Other than morphological and systematic studies (Froggatt,
1893, 1894a,b,c; Gullan, 1979; 1983; 1984a), Apiomorpha has been the subject of few
biological papers ( e.g., Short, 194 7; Gullan et al., 1997).
1.2. Apiomorpha
1.2.1. Galls
Both males and females of Apiomorpha induce galls on Eucalyptus. The galls induced
by females of some species of Apiomorpha, such as A. duplex (Fig. 1. lA), are
spectacular and amongst the largest (up to 20 cm) induced by any insects. They are also
among the most structurally complex of all plant galls (Kuster, 1937; Beardsley, 1984;
Meyer, 1987). Galls of males of Apiomorpha are typically small and tubular (Fig. 1. lB).
6
some species, such as A. spinifer, induce galls that coalesce to form a compound gall
structure (Fig. 1.1 C) on the gall of a conspecific female.
Adult females of Apiomorpha are each confined within the gall that she initiated when
she first began feeding as a crawler. The galls of females of Apiomorpha each have a
chamber in which the insect has limited mobility and is orientated with its head towards
the base of the gall and its anal lobes towards the apex (Fig. 1. 1D). As the gall of a
female matures an apical opening is formed as plant tissue in the region becomes
necrotic. The opening allows elimination of honeydew, mating and the escape of
crawlers. The high sugar content of the honeydew produced by second-instar and adult
females suggests that they feed from phloem (Gullan, 1981 ), the common food source of
aphids and scale insects (Carver et al., 1991 ). Most scale insects and aphids harbour
endosymbionts (Tremblay, 1989, 1997), presumably in a mutualistic relationship that
provides the insect with nutrients not available directly from the plant sap (Moran and
Baumann, 2000). Unusually, endosymbionts have not been found in Apiomorpha
(Tremblay, 1989).
1.2.2. Adult morphology
The morphology of adult females of Apiomorpha is highly modified, presumably as a
result of their endophytic (gallicose) lifestyle. The body shape of females varies among
taxa but, in general, is pyriform (Fig. 1. lE) with the head large and rounded and a pair of
large, sclerotised anal lobes posteriorly. The legs are short and robust and not lost
completely as they have been in many other gall-inducing scale insects. Adult females of
Apiomorpha have very small simple eyes that may function only to differentiate night from day.
The integument of females of Apiomorpha is mostly membranous, with some regions of
sclerotisation (Gullan, 1984a). The membranous nature of the cuticle of coccoids allows
females to continue to grow considerably after moulting to the adult instar (Kosztarab,
1987). Females of some species of Apiomorpha are among the largest of all known
females of A. pharetrata may be only 2 mm in length whereas those of A. duplex can
reach lengths of over 40 mm (Gullan, 1984a; Cook and Gullan, unpubl.). The cuticle has
numerous multilocular wax-pores (Gullan, 1979) but there are neither other pore types
nor macro- or micro-tubular ducts. Hair-like setae occur on most body segments. Many
species have robust sclerotised spine-like setae on the posterior abdominal segments
and, less commonly, on the dorsal surface of other segments.
Adult males of Apiomorpha are not highly modified relative to other eriococcids. They
are small (less than 5 mm in body length) and possess well-developed antennae, eyes,
legs and wings (Theron, 1968; Gullan, 1978, 1981). At maturity, two long wax filaments
develop at the end of the abdomen and protrude from the gall prior to the emergence of
the adult male. Males leave their galls as adults and search for mates.
1.2.3. Systematics
Gullan (1984a) extensively revised the taxonomy of Apiomorpha and since then only
one new species has been described (Gullan and Jones, 1989). Currently there are 39
species of Apiomorpha described. To date, the systematics of Apiomorpha have been
based primarily on the morphology of adult females. Cuticular features of the
integument such as the number and distribution of setae, wax pores and spine-like setae, as well as general body shape and characteristics of the sclerotised anal lobes, spiracles
and legs, provide the majority of taxonomic characters used for distinguishing species.
The galls of adult females of Apiomorpha are an extended phenotype because the shape
is generally species-specific (Gullan, 1984a). Galls of males of some species are also
diagnostic and therefore the morphology of galls induced by both males and females of
Apiomorpha is used to help distinguish among species. Adult males have been little
studied and those of only two species have been described (Theron, 1968). As yet, adult
male morphology is not used in the taxonomy of the genus.
The currently circumscribed species of Apiomorpha may be termed morphospecies because their taxonomy, with its inferred species boundaries, is based solely on
8
biological species. Biological species are based on an assumption of reproductive isolation and may be defined as "groups of interbreeding natural populations that are reproductively isolated from other such groups" (Mayr, 1970). An inference of this definition is that individuals of a species share a common gene pool that is not shared by individuals of another species (Ayala, 1982).
Each Apiomorpha morphospecies has been assigned to one of 14 species-groups (Gullan, 1984a) based on morphological similarities of adult females and their galls. The species groupings are phenetic - although suggesting that taxa within groups are more closely related to each other than to members of other species-groups, this provides no other insights about the evolutionary relationships among constituent morphospecies or among species-groups. To date, there is no published phylogenetic analysis of relationships among morphospecies of Apiomorpha.
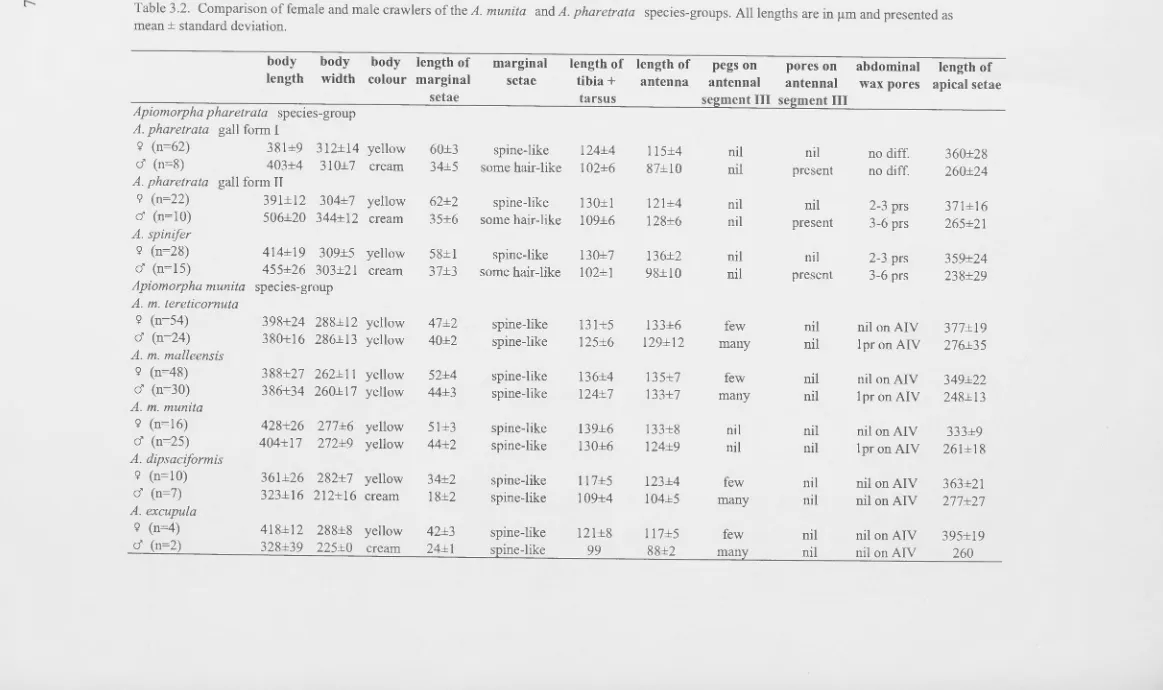
Intraspecific variation in gall and/or adult female morphology has been reported in nine morphospecies of Apiomorpha (Gullan, 1984a): A. attenuata, A. malleeacola, A. minor, A. munita, A. pharetrata, A. pileata, A. strombylosa, A. subconica and A. urnalis. Gullan found that in most cases the intraspecific variation was host-plant-specific and in one case (A. pileata) it appeared to be geographic. However, chromosome number variation concomitant with the morphological variation has been reported in each of A. minor, A. pharetrata and A. strombylosa (Cook, 1992), suggesting that each may represent
species-complexes. In addition, extensive chromosomal variation has also been detected within morphospecies exhibiting no obvious morphological variation (Cook, 1992), thus implying that cryptic species-complexes may be relatively common in Apiomorpha.
1.2.4. Geographic distribution
Apiomorpha has been recorded in all Australian states and territories as well as from
factors such as colonisation events and local extinctions probably playing a large role in their observed distribution (Gullan, 1984a; Cook, 1992; Gullan et al., 1997). The majority of collections of Apiomorpha have been made in south-eastern Australia
(Gullan, 1984a; Cook, 1992; LeBreton and Vaarwerk, 1992, 1993). Although this may
represent the focus of collecting effort, the pattern may reflect host availability because
eucalypts are abundant and species-rich in this region (Wardell-Johnson et al., 1997). Gullan (1984a) reported that the majority of morphospecies of Apiomorpha are relatively host specific. Therefore, it may be expected that species richness of
Apiomorpha in a region may reflect the number of available host species. This appears
to be the case in south-eastern Australia, with the number of gall-inducing insect species
covarying with the number of myrtaceous host species (Blanche and Westaby, 1995).
1.3. Scope of the present study
Apiomorpha may be a valuable study organism for understanding speciation and
evolution in phytophagous insects. It is fully dependent on its host, relatively
host-specific and, because it produces galls, the hosts can be recorded with certainty.
Speciation within a group of related organisms may be determined to a considerable
extent by their genetic systems and population structure (Bush, 1975). In addition, the
process of evolution may be better understood by comparing patterns in one taxon with
those of its close relatives (Sanderson and Donoghue, 1994 ), including sister-group
comparisons (Wiegmann et al., 1993; Barraclough et al., 1998; Farrell, 1998). There is already a strong taxonomic framework for Apiomorpha based on morphology, although chromosomal variation among populations of morphospecies has raised doubt as to
whether adult female morphology reflects biological species boundaries in this taxon.
However, there are few empirical data on genetic and population structuring in
Apiomorpha, and its systematic relationships are not known with certainty. Additionally,
many of the host records for Apiomorpha are derived from museum labels and are not
able to be confirmed.
This project" aims to:
10 • Explore the extent of chromosomal variation in Apiomorpha and determine the extent to which it differs from its closest relatives.
• Determine whether chromosomal variation within some morphospecies of Apiomorpha is attributable to the presence of cryptic species.
• Describe the major aspects of life history of Apiomorpha.
• Describe meiosis and determine the chromosome system employed by Apiomorpha. • Determine the likely breeding system in Apiomorpha.
• Determine the specificity of host associations in Apiomorpha.
• Evaluate the possible modes of evolution and speciation in Apiomorpha.
0
r
m
11
Figure 1.1. Apiomorpha. (A) Galls of adult females of A. duplex. (B) Individual galls of males of A. minor on a leaf. (C) Compound gall (black arrow) of males of A. spinifer on
the gall of their mother (white arrow). (D) Gall of A. calycina with the side cut away to reveal the adult female. Her head is towards the base of the gall. (E) Adult female of A.
ovicoloides ( courtesy of P.J. Gullan). She has recently given birth to crawlers (small and
yellow). (F) Stand of Eucalyptus nortonii. There is a population of A. conica on the small sapling (arrow) but not on other saplings in the stand. Inset (G). Enlargement of the sapling. The population of A. conica is very dense, with males galls covering the
[image:30.804.47.792.36.1139.2]CHAPTER 2.
THE SYSTEMATIC PLACEMENT OF APIOMORPHA BASED ON DNA SEQUENCE DATA
Parts of this chapter are adapted from the manuscript "A molecular phylogeny of the scale insects (Hemiptera: Sternorrhyncha: Coccoidea) based on nuclear small-subunit ribosomal DNA" by LG Cook, PJ Gullan and HE Trueman. The manuscript is near completion and is intended for submission to the journal "Molecular Phylogenetics and Evolution". Thus, there have been contributions by the other authors, particularly Penny Gullan. Penny provided background research and discussed ideas but the text used in this chapter is my own. Penny also collected, or obtained, and slide mounted most of the specimens used for DNA sequencing other than those of Cylindrococcus, Frenchia and Pseudococcus, and she confirmed
identifications of most specimens. All sequences were derived and analysed by me, except that Holly
Trueman provided ssu rDNA sequences for Jcerya purchasi and Orthezia urticae and that two sequences
(Aonidiella aurantii and Dysmicoccus neobrevipes) were obtained from GenBank
(http://www.ncbi.nlm.nih.gov) (Benson et al., 2000). Apiomorpha was not included in the above study and
"The systematic positions of Apiomorpha and other Australian gall-forming coccoid
genera are problematical."
Penny Gullan (1984a, p. 132)
"I'm having a lot of trouble with my character(s)."
Chris Hodgson (ISSIS, 1998) referring to his morphological data set.
2.1. Introduction
13
Apiomorpha belongs within the neococcoids because it possesses the putative
synapomorphies of that group: a chromosome system involving paternal genome elimination (Nur, 1980; Danzig, 1980), needle-like apical setae on the labium (Koteja, 1974a; 1996), and no abdominal spiracles (Morrison, 1928). However, the systematic
placement of Apiomorpha among the neococcoids is less certain.
It is generally agreed that Apiomorpha is an eriococcid (Froggatt, 1921 a; Ferris, 1957;
Hoy, 1963; Gullan, 1984a; Ben-Dov et al., 2000), although some authors have treated the taxon as a separate family within the Acanthococcidae family group (Koteja, 1974b; 197 6), or a distinct family with uncertain affinities (Borchsenius, 19 5 8; Cox and Williams, 1987; Koteja, 1996). Theron's (1968) study of adult male characters supported a relationship with the Eriococcidae. He further postulated that Apiomorpha was closely related to Opisthoscelis on the basis that both were eucalypt-feeding gall-inducing Australian endemics. Several other authors (Signoret, 1869; Balachowsky, 1942) have also thought Apiomorpha and Opisthoscelis to be closely related, including Froggatt (1921a) who also included Ascelis, Cystococcus and Frenchia (actually an. asterolecaniid) in the same subfamily. A close relationship between Apiomorpha and
The recognition of the Eriococcidae per se is probleTI?-atic. Cox and Williams (1987) argue that the Eriococcidae are primarily separated from other coccoid families by the absence of the characters that help define other families, rather than by the possession of unique characters (autapomorphies) that define the Eriococcidae. For example, all characters used by Ferris (1957) and Hoy (1962) to define the Eriococcidae (anal lobes, well developed antennae, well developed legs, tubular ducts, sessile pores and anal ring) are found also in other scale insect families. It appears likely that the anal lobes of eriococcids are homologous with those of coccids (Cox and Williams, 1987; Hodgson, 1995). The macrotubular ducts with cupped inner ends found in eriococcids are also common in the Aclerdidae, Cerococcidae, Lecanodiaspididae, Coccidae (Williams, 1985b; Cox and Williams, 1987), Kermesidae and Putoidae. Microtubular ducts are found in many coccoid taxa, including Pseudococcidae, and their morphology often differs even among eriococcid taxa. The anal ring structure is common to many of the neococcoid families and quinquelocular and multilocular pores are shared with most scale insect families. If these structures are homologous among the Eriococcidae and the other scale insect families in which they occur, then either the structures are plesiomorphic for the eriococcids (i.e., they occurred in a common ancestor of eriococcids and the other taxa in which they are found), or the Eriococcidae are not monophy letic.
Most inferences about scale insect relationships have been based on features of adult females and, to a lesser extent, crawlers and adult males. Consequently, the affinities of
Apiomorpha are difficult to assess because adult females of Apiomorpha are highly derived morphologically, presumably due to their specialised life style of inhabiting galls. The enlarged spines, multilocular disc pores and anal ring structure of crawlers of
Apiomorpha support an affinity with the Eriococcidae but they and other features do not
clearly indicate an affinity with any particular eriococcid genera. Although morphology has been useful in elucidating the systematics of some other scale insect groups ( e.g., Qin and Gullan, 1995), it may be of limited use for inferring the relationships of
15
i.e. homoplasy. Therefore, the relationships inferred from the morphologies of adult females of gall-inducing species may be misleading and need to be treated with caution. The morphologies of adult males of some other gall-inducing taxa also exhibit adaptations to the galling habit of females ( e.g. Gullan, 1978) and do not appear to clarify the placement of Apiomorpha (Gullan, 1984a). Immature instars have not been
extensively studied previously in Apiomorpha, or in most other scale insects.
It is necessary to know the phylogenetic affinities of Apiomorpha in order to better make
inferences about the evolution of traits such as the origins of gall induction, chromosome system evolution, chromosome number evolution, and host specificity. In this chapter, partial sequences from two genes, one nuclear and the other mitochondrial, are used to determine the systematic placement of Apiomorpha. In addition, the monophyly of the
Eriococcidae and Eriococcus is tested, and the origins of gall induction examined.
Molecular data were used in view of the apparent difficulties with inferring the relationships of Apiomorpha using only morphological data.
Nuclear small subunit ribosomal DNA (ssu rDNA) sequence data were obtained as part
of a project undertaken to estimate a molecular phylogeny for the whole of the
Coccoidea. This gene is appropriate for high-level scale insect studies because it is slowly evolving (conservative) (Hillis and Dixon, 1991; Caterino et al., 2000) and therefore may be informative for ancient divergence events. The scale insects, as a group, probably arose near the Pennian (i.e., prior to 245 Ma) (Gullan and Kosztarab, 1997). Partial DNA sequences of ssu rDNA were obtained for representatives of 10 families of scale insects, including a diverse range of eriococcids, to test whether
Apiomorpha is more closely related to eriococcids than to other neococcoid taxa.
Partial sequences from the mitochondrial gene Cytochrome Oxidase II (COII) were derived as part of a separate study to resolve internal relationships within Apiomorpha.
COII is more informative than ssu rDNA for lower-level systematics because it evolves faster (Simon et al., 1994). Only a small region (about 300 bp) of COII, and no other
as putative outgroups, probably because of primer mismatch. Due to limited funding, no custom primers were designed for other taxa or gene regions.
Because the SSU rDNA and
con
sequences were gathered for different projects, not alltaxa are represented in both the data sets. However, there is sufficient overlap of taxa to use these data to address the main objectives of this chapter. Both data sets included Opisthoscelis and several other Australian gall-inducing taxa and therefore the suggestion of a close relationship between Apiomorpha and Opisthoscelis was tested.
2.2. Methods
2.2.1. Specimens and DNA extraction
Margarodids and ortheziids were used as the outgroups in phylogenetic analyses of ssu rDNA data. A pseudococcid, identified in the ssu rDNA sampling as a suitable outgroup, was used to polarise
con
trees. The taxa used, their current taxonomic classification,and collection localities are shown in Table 2.1. The cuticles of adult females from which material was dissected were slide mounted to allow identification and to provide voucher specimens. If the specimen was destroyed during DNA extraction, other females from the same collection were used as vouchers. Embryos and/or ovarian tissues were dissected from larger females. Small females and eggs were homogenised whole. DNA was extracted from fresh or ethanol-preserved specimens using the salting-out method of Sunnucks and Hales (1996). A single chloroform wash was performed prior to precipitation with ethanol if solids or excessive pigments were present.
DNA was extracted separately from three individuals of Eriococcus buxi and two individuals of E. aceris, to provide independent confirmation of the SSU rDNA sequence for these two tax a.
2.2.2. PCR and sequencing
I
17
Dohlen and Moran, 1995). Six additional primers from von Dohlen and Moran (1995, and unpubl.); B [for] (5'-ctggtgccagcagccgcgg), 2882 [for] (5'-agaggtgaaattcttgga), 2883 [rev] (5'-tccaagaatttcacctct), E [for] (5'-ggtggtgcatggccg), E- [rev] (5'-cggccatgcaccacc), 2881 [rev] (5'-gatccttccgcaggttcaccta), and six novel primers designed during the present study were used in combination to amplify the entire ssu rRNA gene of Apiomorpha
munita tereticornuta. The new primers are:
El 0for (5'-ggaaggaacgctcttattag)
E 1 0rev ( 5 '-c ggttttgatctaataagagc)
E23 for ( 5 '-ttagagtgctcaaagcaggc)
E23rev (5'-gttccattattccatgcgacc)
E4 3 for ( 5 '-ttcttagagggacaggcggcg)
E43rev (5'-catcacagacctgttatcgc).
An internal region of the COII gene was amplified using the primers C2-J-3400 (Liu and Beckenbach, 1992) and C2-N-3661 (Simon et al., 1994).
PCR reactions (25µ1) contained 5pmol of each primer, l0mM Tris-HCI (pH 8.3), 3mM MgCI, 50mM KCI, 0.2mM of each dNTP, 1 unit of Taq-polymerase (Fisher Taq Fl,
Biotech) and 2 µI of template. Amplification was carried out in a Corbett Research FTS-960 thermal sequencer using a 45 sec denaturation at 94 °C, 30 sec extension at 72°C and using a 5°C stepdown program for annealing, with the first cycle at 65°C and annealing
in later cycles reduced by 5°C after every second cycle until reaching 45°C. An
A 1 0µl sequencing reaction was carried out using ABI (Perkin-Elmer) BigDye terminator chemistry following the manufacturer's instructions. Sequencing was performed using an ABI Prism 3 77 automated DNA sequencer. All DNA fragments were sequenced in both the forward and reverse directions.
2.2.3. Analysis
2. 2. 3.1. Editing and alignment
Sequences were edited using Sequencher 3 .0 (Gibbs and Cockerill, 1995) and a similarity search was performed on all sequences using the "basic blast" option (Altschul etal., 1997) at the NCBI website (http://www.ncbi.nlm.nih.gov/BLAST/). Three scale insect ssu rDNA sequences (Table 2.1) were obtained from Genbank (Benson et al., 2000).
A preliminary alignment was made using CLUSTAL _ X (Thompson et al., 1997) using default weightings. Adjustments were made by eye to the ssu rDNA alignment with reference to the predicted secondary structure model for eukaryote nuclear ssu rRNA (Van de Peer et al., 1998). Regions of conflict between the model and each of the scale insect sequences were compared using minimum energy RNA folds obtained for each helix using Mfold (Mathews et al., 1999; Zuker et al., 1999). Some highly variable regions, particularly E 10, did not have the same predicted secondary structures and were not able to be unambiguously aligned across all taxa. These regions were excluded from further analyses. The ssu rDNA alignment was partitioned into 3 categories on the basis of secondary structure: stems, loops and single stranded regions.
There were no indels in the COII region and no further editing was required. Because of the possibility of transposed copies of COII (Sunnucks and Hales, 1996), each nucleotide sequence was translated to amino acid sequence to check for the occurrence of stop codons. Stop codons would indicate that the copy is nonfunctional, and probably of nonmitochondrial origin (i.e., a nuclear pseudo gene). In addition, the distribution of substitutions between 1st, 2nd and 3rd codon positions was compared among taxa. In
19
occurring in 3rd codon positions, with fewer occurring in 1st positions and fewest in 2nd postions (Simon et al., 1994 ). If this pattern is not followed in pair-wise comparisons among closely related taxa, the possibility that at least one of the sequences is that of a pseudogene should be considered.
2.2.4. Phylogenetic analysis
A variety of different approaches to phylogenetic reconstruction was used, primarily to guard against undetected violations of underlying assumptions in any one model. It was assumed that, if different models and weighting schemes resulted in the same topology,
then the resultant phylogeny was robust in respect of inference method. Robustness does not imply that the phylogeny is accurate.
2.2.4.1. Maximumparsimony
Heuristic searches comprising 100 random addition sequence starting trees, TBR branch swapping and no maxtrees restrictions were performed using PAUP* 4.0b4a (Swofford,
1999). Three weighting schemes were used for the ssu rDNA data:
1. equal weights,
2. transversions weighted twice that of transitions because the estimated transition:transversion ratio was 2,
3. stem sites weighted half that of loops and single-stranded regions on the nai've basis that double-stranded regions are involved in non-independent mutations because compensatory change occurs on the opposite strand in order to maintain pairing. That is,
a change in each strand (2 sites) is one event (see Soltis et al., 1999).
were weighted 5, and 3rd positions 1. The latter is arbitrary weighting to determine whether there is high sensitivity to the weighting scheme.
A combined analysis was performed using only those taxa that were common to both data sets. A partition homogeneity test, as implemented in PAUP* indicated that the two data matrices were combinable (p=0.615). A branch and bound search was conducted using a weighting scheme where all positions were weighted 3 except that 3rd codon positions in the COII data set were weighted 1. This weighting scheme was chosen because it is simple but still allows for the down-weighting of the problematic, but informative, third codon positions in the COII data set.
2. 2. 4. 2. Neighbor-Joining
Models of varying complexity (base frequencies, substitutional classes, percentage of
invariant sites (I), site-specific rates (1)) were compared using log likelihood ratio tests
(Goldman, 1993; Yang et al., 1998; Sullivan and Swofford, 1997). The most simple
model which did not differ significantly from more complex models was chosen ( see Maddison et al., 1999 for justification). For the ssu rDNA data, this model approximated
a Kimura two-parameter model (K2P+r) (Kimura, 1980) assuming equal base
frequencies and site specific rates (1 shape parameter = 0 .16). The LogDet model
(Lockhart et al., 1994) was chosen for the COII data because significant base-compositional bias was present among taxa. Two analyses were run, one with the full
21
2. 2. 4. 3. Maximum Likelihood
ML analysis of the ssu rDNA data was run under three evolutionary models: Jukes Cantor (Jukes and Cantor, 1969), K2P assuming equal rates for all sites and with site-specific rates (gamma) and transition/transversion ratio estimated from the data.
2. 2. 4. 4. Statistical tests
Most methods of phylogeny reconstruction assume that the base composition is at equilibrium (stationary) (Page and Holmes, 1998). That is, the base composition of sequences is the same for all taxa under study. Differences in base composition among taxa (non-stationarity) were tested using a Chi-square test as implemented in PAUP*. Non-stationarity was further tested by running the analysis using only several taxa which differed markedly from each other in base composition.
Bootstrap tests (Felsenstein, 1985) were conducted using 1000 replicates and the same settings as the original analyses but including only one representative of taxa with identical aligned sequences. Internal nodes were also assessed using Bremer support ( decay index) (Bremer, 1988). Groupings assumed to be monophyletic under previous hypotheses (Eriococcidae and Eriococcus) but not appearing in the unweighted MP trees were tested using topology-dependent permutation tail probability (T-PTP) tests (Faith, 1991).
2.2.5. Length of ssu rRNA
2.3. Results
2.3.1. ssu rDNA
2.3.1.1. Sequence variation
Identical sequences were obtained from different individuals of each of E. buxi and E. aceris. No taxa of scale insects shared identical complete nucleotide sequences although, in the aligned regions used in phylogenetic analyses, E. aceris was identical to E.
spurius, and Opisthoscelis mammularis was identical to Lachnodius sp .. The four bases occurred in equal proportions and there was no compositional bias among taxa. In the aligned regions (508 bp), there is up to 10% divergence (K2P, gamma shape parameter=0.14) among scale insects and 0.6-2. 7% divergence among the species of Apiomorpha sampled (Table 2.2).
2. 3.1. 2. Length of ssu r DNA sequence using primers 2 880 and
B-The length of the fragment amplified using primers 2880 and B- in non-Apiomorpha scale insects ranged from 545 in Austrolecanium sassafras to 643 in Puto yuccae. In each species of Apiomorpha, the fragment was considerably longer than that known for
other scale insects. PCR amplification of the whole of ssu rDNA using primers 2880 and 2881, which prime near the 5' and 3' ends respectively, resulted in a product around 4500 nucleotides long in Apiomorpha sp. (Fig. 2. lA). The sum of the length of fragments amplified using internal primers (Fig. 2. lB) also is about 4500 nucleotides in A. munita tereticornuta. Digestion of genomic DNA extracts of A. m. tereticornuta (Fig.
~ 23
for the amplified 5' region of ssu rDNA of A. m. tereticornuta (1159bp) is 240
nucleotides longer than that for the same region in X vesparum (919bp ).
A PCR-based survey, using primers 2880 and B-, of other species of Apiomorpha
indicates that the fragment is longer in all species of Apiomorpha surveyed than it is in
all other scale insects. The length of the fragment varies considerably among surveyed
Apiomorpha species, from 760 bp in A. pomaphora to 1159 bp in A. m. tereticornuta
(Fig. 2.lE). The length of the ssu rDNA fragment is shorter in A. munita munita than it is in A. munita tereticornuta (Fig. 2. lE).
2. 3.1. 3. Inferred phylogeny
The same two MP trees were obtained under the three different weighting schemes (strict consensus Fig. 2.2). The neococcoids, excluding Puto yuccae, comprise a monophyletic group but support is weak. Puto (Putoidae) appears outside the
neococcoids as sister to the ortheziids in all trees but there is no bootstrap support for this placement and Bremer support of only 1. There is strong support for the monophy ly of each of Pseudococcidae, Coccidae, Diaspididae and Asterolecaniidae. A monophy letic Eriococcidae was not recovered under any model or method because the diaspidid and asterolecaniid taxa fell among the eriococcid taxa in all analyses. The relative positions of the Diaspididae and Asterolecaniidae differed between MP and NJ methods. In unweighted MP analyses, there was no bootstrap support, and Bremer support of only 1, for the positioning of each of the Diaspididae and Asterolecaniidae among the taxa currently recognised as belonging in the Eriococcidae. An a priori
assumption of the monophyly of the Eriococcidae could not be rejected by the ssu rDNA data (T-PTP, p=l.0).
A monophy letic Eriococcus sensu lato was not recovered under any method or model
and was rejected by T-PTP (p=0.001). Taxa currently assigned to Eriococcus fall into three clades which each include taxa currently assigned to other genera. There is strong support for a sister relationship between E. eucalypti and Madarococcus (T-PTP, p=0.001), with both appearing as sister group to the gall-inducing taxa Opisthoscelis,
Cylindrococcus form the best supported clade in the analysis (bootstrap= 100, T-PTP p=0.001) with 12 character state changes also supporting the node, six with a CI of 1 and the others unique to the node. There is weak support for a clade comprising E. aceris, E. coccineus, E. spurius, Dactylopius, Phacelococcus, Ourococcus and Apiomorpha. Each of the three eriococcid clades includes gall-inducing taxa.
2.3.2. COIi
2. 3. 2.1. Sequence variation
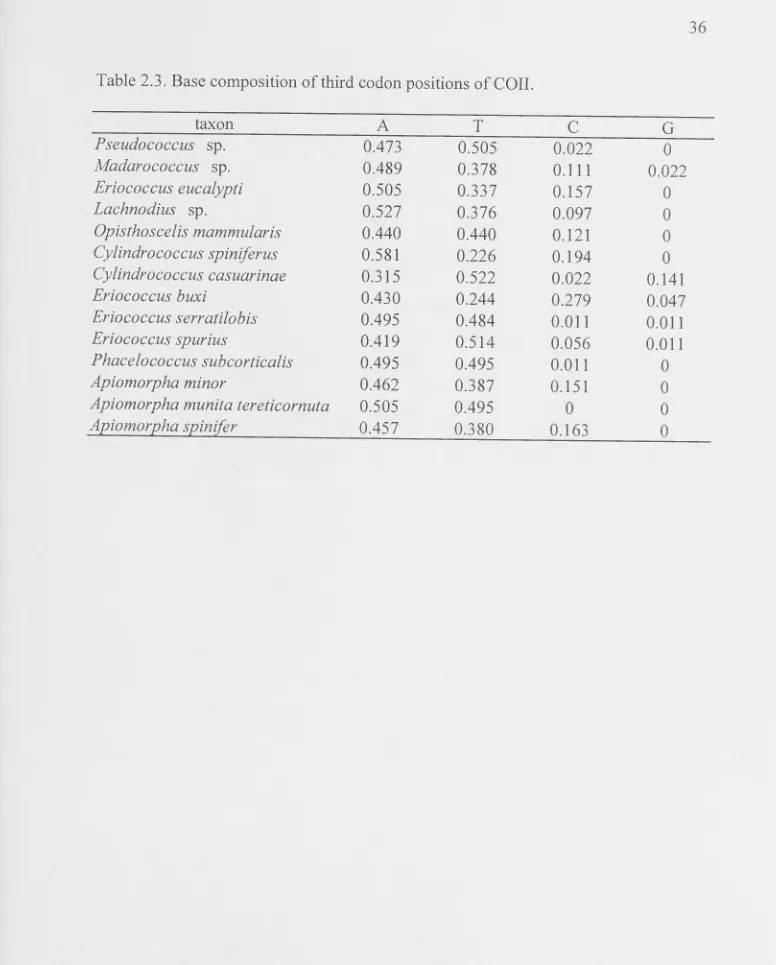
There was no length variation (indels) among taxa and all sequences appeared to code for functional products, with no stop codons and the expected distribution of substitutions among codon positions. There is significant base composition bias towards very high AT but very low G content, especially in 3rd codon positions (Table 2.3). There is also significant compositional bias among taxa which is strongest in 3rd codon positions, with AT content ranging from 67% in E. buxi to 100% in A. munita tereticornuta (Table 2.3), but also present in 1 st+2nd codon positions. The non-stationarity in 3rd codon positions is partially due to high A in some taxa but high T in others, rather than solely a difference in A+ T. Additionally, the proportion of C varies
among taxa and C. casuarinae has high G (14%) relative to other taxa. The inferred
amino acid sequences were not used for phylogenetic analyses because the nucleotide compositional bias, and/or codon bias, may affect the inferred amino acid sequence (Foster et al., 1997; Singer and Hickey, 2000) and therefore exacerbate, rather than correct for, compositional bias.
There is very high divergence among taxa, even after LogDet transformation to take account of different base compositional biases. There is up to 20% divergence within
Apiomorpha, and 15 to 30% between Apiomorpha and other eriococcid taxa. Plots of transitions against transversions for 3rd codon positions alone (Fig. 2.3A), and for
25
2.3.2.2. Inferred phylogeny
The eriococcid taxa fall into three clades (Fig. 2.4) which do not conflict with those recovered from the ssu rDNA data. Madarococcus sp. and E. eucalypti are sisters, as are Lachnodius and Opisthoscelis. These four taxa form a clade (hereafter referred to as the "yellow" clade) under all methods and models. The two Cylindrococcus species have a sister relationship despite their different base compositional biases. Cylindrococcus and
E. buxi are sisters (hereafter referred to as the "blue" clade ), as they are in the ssu
rDNA-derived trees. The blue clade is labelled separately from the yellow clade because Cylindrococcus and E. buxi do not appear as sister to the yellow clade under an MP weighting scheme of 5: 5: 1. The yellow and blue clades are sisters under all other models. Apiomorpha clusters with Phacelococcus and E. spurius, as it did in the ssu rDNA derived trees, with E. serratilobis (not sequenced for ssu rDNA) also in the clade (hereafter referred to as the "red" clade). Apiomorpha appears as monophyletic.
The two NJ models (LogDet and HKY85) gave the same topology (Fig. 2.4 ), which is the same as the ML tree and very similar to that derived using MP with a 2:3: 1 weighting scheme. The only difference between this MP model and the NJ and ML models concerns the placement of E. spurius. In the NJ and ML phylograms, E. spurius is sister to E. serratilobis whereas it is placed as sister to the rest of the red clade in MP trees.
2.3.3. Combined data
I
2.4. Discussion
2.4.1. Systematics of the Coccoidea
The ssu rDNA data support the recognition of the neococcoids with the exclusion of Putoidae. Although too few taxa were sampled for each of Pseudococcidae, Coccidae, Diaspididae and Asterolecaniidae to rigorously test the monophy ly of each, there was strong support for sister relationships among the disparate taxa sampled for each. Relationships among these families are not well resolved by these ssu rDNA data. Although Pseudococcidae consistently appeared as sister to the rest of neococcoidea, there was little support for this relationship. There is weak support for a sister relationship between Coccidae and Kerriidae, consistent with Koteja's (1976) observations that the salivary pumps of each are similar and the morphology of crawlers and adult females (Miller, 1991).
2.4.2. Systematics of the Eriococcidae
The ssu rDNA data do not support the monophyly of the Eriococcidae. These results are congruent with those observed for morphological data. Eriococcidae is primarily defined by the lack of characters that are synapomorphic for other families, leading Cox and Williams (1987) to suggest that Eriococcidae may be a paraphyletic grouping. In the present study, Eriococcidae is rendered paraphyletic by the inclusion of representatives of the Diaspididae and the Asterolecaniidae. However, the ssu rDNA data do not present strong evidence for the inclusion of either of these two families among taxa currently recognised as belonging to Eriococcidae. Additionally, the monophyly of the