0022-538X/81/050621-11$02.00/0 Vol.38, No. 2
Restriction Alleviation by Bacteriophages Lambda and
Lambda Reverse
PENELOPETOOTHMANt
Department of Chemistry and Institute ofMolecular Biology, University of Oregon, Eugene, Oregon 97403 Received24September1980/Accepted 29 January 1981
Deletion analysisindicated that the phage A restriction alleviation gene(s) ral resides between thecIIIand N genes. TheRal+ phenotype was expressed only whenXral+carried amodificationsuch that it wasresistant to restriction by the host specificity system. Under theseconditions, Ralfunction protected superin-fecting unmodified phages from restriction by EcoK or EcoB but not from restriction by EcoPl.Ral-protectedphage DNA was notconcomitantlyK and B modified, but rather received onlythe modification specified by the system of the restricting host. Possible mechanisms for Ral action are discussed. Of the other lambdoidphages tested, thehybrid phageArevhadRal activity, whereas
08Ovir
andone X-P22 hybrid did not. The restriction alleviation activity ofXrevcalled Lar, may be the same as theactivity expressed in sbcA- strains of Escherichia coli,but it wasfunctionallyseparable from exonuclease VIII activity (the product ofthe recE gene), which is also expressed in sbcA- strains.Host modification restriction systems were firstdescribedby Arber and Dussoix (2, 11)and havebeen identified inanumber of speciesand strains of bacteria (41). Presumably, modifica-tion andrestriction have evolvedtoprotect bac-teriafrominvading foreign DNA, although ad-ditionalroles havenotbeen ruledout.The mod-ification restriction loci of Escherichia coliK-12 and E.coli B and the SB system ofSalmonella typhimuriumareallelic and include three genes
(hsdS, hsdM,
andhsdR)
(5, 15, 24, 53). Thespecificity
ofthe system is determinedby
the hsdS gene(5).
Modification and restriction in these bacterialspecies
have beendesignated
typeIandarecharacterized
by
anenzyme com-plex which requires the cofactorsS-adenosyl-methionine,ATP, and
Mg2e
torestrict unmodi-fied DNA but only theS-adenosylmethionine
cofactorto
modify
DNA(28,
32,
37,55).
Meth-ylation occurswithin therecognition
sequence (45, 52), whereasendonucleolytic cleavage
oc-cursoutside thissequence andshowsno appar-ent sequence
preference
for the actual DNA cleavage reaction (1, 23,38).Bacteriophageshave evolved several waysto disruptthe
complex
process ofmodificationand restrictionand thus escape lethalcleavage.
The temperatephage Mu is resistant to restriction by avariety of restriction enzymes because ofmethylation
byarathernonspecific
modification function (mom) after induction from thepro-tPresent address:DepartmentofBotany,AJ10,University
ofWashington,Seattle,WA98195.
phage state (51). The T-even phages escape restriction becauseDNA isglucosylated (12). T3 and T7expressocrfunctions
immediately
upon infection, which preventnotonly restriction but also modificationby
EcoB orEcoK (27, 47,48). Although T3ocr-, T7ocr-, and T-evenglucosyl-ation-defective phages are sensitive to restric-tion, the rare phages that doescaperestriction aremodified. Another virulentphage,
T5,
must have some mechanism(s) to escape restriction byEcoRI, asT5 DNAhassix sites sensitiveto EcoRIinvitro (54),yetnoEcoRI restrictionor modificationoccursin vivo(8).TheXrestrictionalleviationfunction,
Ral,
was firstcharacterizedby
Zabeau(M.Zabeau,
Ph.D. thesis,Rijkuniversiteit Gent, Ghent,
Belgium,
1974), whoseobservationshave beenpublished
recently (9, 10,58).Here Ishow that AandXrev (a recombinant between theRac
prophage
ofE. coli K-12 and X[25])
have functions(Ral
and Lar)whichcanpreventrestrictionby
EcoB and EcoK. Inaddition,
I suggestthattheLaractivity
of
Xrev
isthesameactivity
that isexpressed
in sbcA- mutantsof E. coli(44).
MATERIALS
AND METHODSStrains.Phage and bacterial strainsare described in Table 1. M8521 is strain M5221 ofH. Greer (18) selected to grow on eosin methylene blue (EMB)
plates.
Media.Abroth contained1%tryptone, 0.25%NaCl,
and 0.1% yeast extract; A plate agar and top agar contained,inaddition,1.1and0.65%agar,respectively, but no yeast extract. LB plate agar contained 1% 621
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Listof bacterial andphagestrains
Restriction-Strain tion pheno-modifica- Othermarkers Sourceorreference type
Bacteria M72 M8002 M8521* M5222 M8521(P1) M5222(P1) Q1 Q8002 Q8003 Q8004 Q8100 Q8803 Q8209 Q8302 Q8303 Q8304 QD5003 QA8002 QA8004 QW11 M1742 M18000
MI8000AR
AB1157 JC5519 JC8679 JC8689 JC8690 JC8691 Phage
AbiolOcI857
Abiol0imm2' cl-Abio252cI857 Abio252imm2cI-Aimm43cI6T Ah bio243imm434cI6T Ah80att"°cI857
ASpi-imm22c25(OP)P22
480vir
ArevexbiolOcI857p4
Ab2intlO2red3cI857p4 Plc+CM
K K K K K,P1 K,P1 K
su-lacamtrpamstr-M72(AcIind831) M72(bio255cl857H1) M72(bio247cI857H1) B1+revertant of C600,
080R
K Ql(AcIind831)
K Qi(Aimm434T)
K Ql(Aimm21)
O C600(rm-)
O r-m-gal-met-[803]
B
rB+msB
F+[2027]B Q8209 (Aind831)
B Q8209(Aimm43T)
B Q8209(Aimm21)
K Ymel,suIII+
K
CR63(h"°att"°cI857)AR
K CR63(h'att °imm21)AR
K recA56recB21[JC7507]
K recA- str- su-(gal-A)A
K MI742
(Ab2cI8570am5O1Sam7) K MI8000 madeAR,mal+
K AB1157suII
K recB21recC22 str31 K recB21 recC22 sbcA23 K Rec-,UV8 derivatives
K JC8679 lacking
K ExoVIII activity
E.Signer Thiswork 18 18 This work Thiswork 43
I. Herskowitz I.Herskowitz I. Herskowitz
M.Meselson via F.W.Stahl W. B.Wood viaJ.Engel W. B.Wood via J.Engel This work
This work This work 16 This work This work
J. Clark via E.Signer 21
A.Strathern
This work E.Signer J.Gillen
J.Gillen
J.Gillen J.Gillen
J.Gillen
42
W. F. Dove E.Signer E.Signer S.Hilliker E.Signer This work I. Herskowitz L. Rosner tryptone,0.5%NaCl,0.5%yeast extract,and 1.5%agar.
Low-salt tryptone top agar contained 1% tryptone, 0.6%agar,and10-3 MMgCl2. SMfor phage dilutions has been described previously by Weigle etal. (56) andwassupplemented with 0.01 MMgSO4and 0.01%
gelatin.EMBagarwas asdescribed by Campbell(7),
butwith 0.05% maltose asasugar supplement. OM
medium for scoring A deletion mutations contained
sodium citrateandwas as describedbyManlyetal. (36).
Phagestocks. Phage stocksweremade fromsingle plaquesby the confluent lysis method. The single-plaque isolations were done in the same bacterial
strains used tomakethe stocks.Modifiedphage stocks weremadeinthefollowinghosts:Q8100 forA.0,Q8803 forAh'.0,Ql forA.K, QD5003 forAh'.K, Q8209for
A.B, Q8100(P1)forA.P,andQ1(P1) forA.K,P.
Plating conditions. Plating bacteria were pre-pared daily by dilutingafreshly saturatedovernight
culture 100-fold intoAbrothcontaining 0.2% maltose andgrowingtoadensity of5x
10i
cells per ml(ifAh'phages weretobe used,low-salt tryptone medium was used).Adilutedphage sample (0.1ml)was mixedwith 0.2 mlofplating bacteria, adsorbed at room tempera-turefor20min, mixed with2.5 ml of top agar, and poured onto 1-day-old A medium plates. A top agar
622 TOOTHMAN
on November 10, 2019 by guest
http://jvi.asm.org/
and Aplateagar were usedforA. Low-salttryptone
topagarandAplateagarwereused forh' and
080.
Plateswereincubatedat37°Cunlessotherwise
indi-cated.
Construction of lysogens.Alysogenswereformed
byspottingphageatapproximately108phageperml
onto bacteria in a A agar or LB agar plate. After
incubationat 34°C,the surviving bacteriawere
puri-fied andtested forimmunityand resistance.
To make P1 lysogens ofM5222, M8521, Ql, and
Q8100,0.2-ml portionsof platingbacteriawerepoured
onto LB agar plates containing 2.5 mM CaCl2.
Plc+CMwasspottedontothese bacteriallawns, and
single colonieswerepurified fromthe lysisareas on A
agarplates containing12.5jig of chloramphenicolper
ml.Coloniesof purified isolatesweretestedonEMB
platesfor the ability torestrictAimm4cI.Kand the
failuretorestrictAimm4cI.K,P.
Coinfections.Afreshovernightculturewasdiluted
100-fold into A broth containing 0.2% maltose and
growntoadensityof1 x108to2x 108cells permlat
37°Con areciprocatingshaker. Bacteriawere
centri-fuged and thensuspendedat108cellsperml in0.01M
MgSO4when the helperwashAand in 0.001 M MgSO4
when the helperwashe";0.2-mlportions of bacteria and 0.2-ml portions of helper phage were mixed in conical 12-mil glass centrifuge tubesat afinal
multi-plicity of infection (MOI) of5(unlessotherwise indi-cated) andadsorbedatroomtemperaturefor15min. LB broth (3 ml) was added to each tube and the infectedcells wereaeratedbyshakingat37°Cfor10
min.AsamplewasdilutedintoSM containing
chlo-roform andtitrated for unadsorbed phage. The in-fected cells were centrifuged, 0.2 ml of test phage (MOI, 0.25, unless otherwise indicated) was added,
and the pellet wasblended with a Vortex mixerto
suspendthebacteria.After furtherphage adsorption for 15 min at room temperature, the cells were (i) eitherdiluted 100-fold intoprewarmed LBbroth and
grownforasinglecycle of growthtomeasurephage yieldorplated directly withtheappropriateindicator
bacteria for infectivecenterformationorboth and (ii)
diluted into SM containingCHC13tomeasure
unad-sorbed phage. When several indicator strains were
usedtodetermine the yields of individualphagetypes,
Avirwastitratedon allindicator strainstobesurethat
the phage plating efficiencies were comparable and withintheexperimental samplingerror.
Controlinfections were done in parallel; in these
infections bothhelperandtestphagesweremodified
to determine (i) whether the Ral helper effect was
specificforstimulatingunmodified phage growthand (ii)whetherphageyields under nonrestricting condi-tionswerenormal. Eachcoinfection described below wasdone atleasttwice in thehostindicatedand, in several instances, inadditional bacterialstrains.
Singlecycleofgrowth.M8521and M5222
(lyso-genscarryingdefectiveRal+ and Ral-prophages,
re-spectively)weregrownunderbothnoninducing(33°C) andinducing(40°C)conditions and usedsubsequently atthecorresponding temperaturesto testtheir abili-tiestosupportthe growthof bothmodified and
un-modifiedsuperinfecting phages; 100-fold dilutions of saturatedculturesweregrowntoadensityof 2x 108
cellspermland thensuspendedin0.01 MMgSO4. A
0.2-mlportion of bacteria and0.2 ml ofphagewere mixedat afinalMOI of0.25andadsorbed for20min atroomtemperature.Infectedcellswerediluted 100-foldinto prewarmed LB brothatthe corresponding
temperatures and 100-fold into SMcontaining CHC13
to measure unadsorbed phage. At 10-min intervals
samples were withdrawn, treated with CHC13, and
titrated.Theendpointandphageyieldsof thesingle
cycles of growth were determined when the phage
titersreachedaplateau.
Raleffectonthemodificationof nonreplicat-ingphage. Coinfectionswere
performed
asdescribed above, except that both the helper and test phages wereaddedatMOIs of10,and afteradsorptionof the testphage A antibodywas added atafinal K of45 beforeplatingforinfectivecenters.Isolation of Arev exbiolOcI857.
Ared-gam-phages, such as AbiolOcI857, cannot grow in
recAA-hostsandaresaidtoexhibit the Fec-phenotype (59).
To enrich for Arevin a stock ofAbiolOcI857, phage
weregrownforasingle lyticcycleinthe recA-strain MI742 at anMOI of50. Arev was isolatedata fre-quency of5 x 10-9 from this stock aslarge
plaque
formersontherecA-strainM1742.(The frequencyof isolation ofArevfromastock of
Abio252c1857
treated inparallelwas9.6x10-'.)
Besides theFec+phenotype, ArevexbiolOcI857shared thefollowingpropertieswithpreviouslyisolatedXrevphage:itgrewinP2
lysogens
(60), it grewonOMmediumplates,and it grew much morepoorlythan itsparent(XbiolOcI857)
at43°C.
RESULTS
Ral
function
of X preventsrestriction
ofsuperinfecting bacteriophage.
The data inTable
2areconsistent
with thehypothesis
that Xhasafunction,
Ral,
whichcanprevent
restric-tion ofunmodified
phage
by
thehostspecificity
system.Ral
appeared
tobepresent
after infec-tions withhybrid phages
carrying
the bio252substitution
butnotafter infections withhybrid
phages
carrying
biolO(Table
2, coinfections
1through 4). Preinfection
withXbio252imm2".K
and
Xbio252cI857.K
partially
blocked
restriction ofsuperinfecting
Ximm43.0,
increasing
theyield
of
Ximm4'
by
factorsof143and17,
respectively.
In other
coinfections
(Table
2,
coinfections 5through
8),
bothRal+
Abio252.0 andRal-XbiolO.0
helper
phage
yields
wereapproximately
10-3, indicating
Ral+
and Ral-phages
werere-stricted
tothesameextent.Inaddition,
theRal+
helper
phages
(coinfections
5 and7)
did notblock
restriction
ofsuperinfecting
Ximm434.0.
Thus, restricted phages could not direct thesynthesis
ofRal,
whichmay be thereasonwhy
Xral+.0
phages failtopreventtheirown restric-tion.The
magnitude
of the Raleffect variedwith thegrowth
phase,
the cultureconditions,
thebacterial
strain,
andthegenotype
of the infect-ing phage. Eventherelatively
small Ral effect shown inTable2wouldhaveasizeable selectiveon November 10, 2019 by guest
http://jvi.asm.org/
TABLE 2. Expression of Ralduring coinfection Coinfection' Phage yieldsduringcoinfection
Helpereffect No. Helper phage Yield ofhelper Yield ofXimm43.K
1 Abio252imm21.K 162(88)b 4.3x lo0- (25) 143
2 XbiolOimm21.K 122 6.0x 10-3 2.0
3 Abio252imm'.K 88 5.0X10-2 17
4 XbiolOimmA.K 135 9.0X 10-3 3.0
5 Abio252imm21.0 2x 10-3(9.4X 10-4) 7.0X 10-3 (170) 2.3
6 Abioloimm21.O 4X10-3 1.0X 10-2 3.3
7 Abio252immx.0 2x10-4 5.0x10-3 1.7
8 Abiol0immA.O 1 x10-4 1.0X10-2 3.3
9 None 3.0x 10-3 (160)
aThesuperinfectingphagewas
Ximm43.0.
Coinfectionswereinbacterial strainQl,
asdescribed in thetext. immXhelperphages carried cI857, imm2'phageswerecI-,and thetestphagewasXimm434cI6T.Yields ofhelper phagesweredeterminedbyplatingonQ8003 (rK+ MK+,Aimm43),andyieldsoftestphagesweredeterminedby platingonQ8002(rK'mK+,A)orQ8004(rK+mK+,Aimm2").
bYields in parentheses are from coinfections done in parallel in which the superinfecting test phage (Ximm434.K)wasmodified.
advantageafter many
generations.
Xral.0would nothavetobeabletoprotectitself inorderfor the ral gene to show aselectiveadvantage.
In coinfectionssuch asthosedescribed inTable2, Xral+ in thewild could preventrestriction ofa superinfectingXral+ phage carryingan inappro-priatemodification;
insuchacase,perpetuating
yourkind is thesame asperpetuating yourself.
Ral
activity
doesnotrequire
host Recorphage
Redrecombination functions. In the experiments shown in Table 2, Ralactivity
was assayed by theability
torescuephage carrying
the immunity region of434. Since this region doesnotcarry aknown K restrictionsite, I felt thatRalmightactby
stimulating
recombination of thehelper with restrictedfragments ofDNA ratherthanby rescuing entiregenomes.Table3 shows that Ral didnotrequire
host recombina-tion functionRecA or RecB. PreinfectionwithXral+
(in this case,Xbio252iMm21cI-.K)
in-creased the numberofinfectivecentersformed by superinfecting AbiolOcI857.0 131-fold. Like-wise, Ral didnot actbystimulatingRedrescue ofrestricted fragments of DNA sinceboth the Ral+ helper phageXbio252imm2'
and the test phageXbiolOcI857.0
wereRed-.Ral expression in the prophage state is underimmunity control. To determine which A genes were essential for Ral function and whether Ral was expressed in the prophage state, theAcI857 lysogens carrying the
Hi
dele-tion and the bio substitutions shown in Fig. 1 were infected withXimm434.0
before and after thermalinduction.Aimm434.0
was able to grow in M8521(bio255cI857H1)at 40 but not at34°C
(Table 4), implyingthatthe Ralactivityofthe prophage wasunderimmunity control. Ral ac-tivity was not expressed in M5222(bio247cI857)
ateither 34 or40°C. These results showed thatTABLE 3. Ral doesnotrequire host RecABor phage Red recombinationfunctionsfor activitya
No. of infective centers produced upon coinfectionofQW11 (recA- recB-) Helper phage
Helper Testphage Helper
phg etphage effect Xbio252imm2".K 0.21(0.17)b 6.8x10-4 (0.01) 131 Abiol0imm21.K 0.14 8.0x10-5 1.5
None 5.2 x 10-5 1
a Thesuperinfectingtestphage wasXbiolO.O.Coinfections were performed in strain QW11 asdescribed in the text. Helper phages carried imm"cI;thesuperinfecting phage was XbiolOcI857. Infective centerformation ofAbiolOcI857 was assayed onstrainQ8004 (rK+mK+,XimmM2),whereas helper phage infectivecenters weredeterminedonstrainQ8002 (rK' MK+,A).
'Thenumbers in parentheses arefromcoinfections done
inparallelin whichthesuperinfecting test phage (AbiolO.K)
wasmodified.
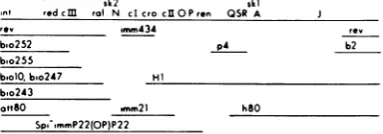
sk2 ski
int redcM ral N cI cro clIOPren QSR A
rev imm434
bro252 p4
boo255
b.olObio247 Hi bo243
ottrO .mm2l hSO
Sprl mmP22(OP)P22
rv b2
FIG. 1. Prophage map ofA indicating relevant sub-stitutionsanddeletions.The regions of DNA deleted bythe variousdeletionsandsubstitutionsare indi-catedbythe black lines. Thefigureisnot drawn to genetic orphysical scale. The bio andatt8O substi-tutionseachcarry a K restriction site, and the mor-phogenesis regionprobably carries three K sites. K restrictionrecognition sites sk2 andsklare indicated onthemap(39).
theRalfunction(s) islocatedtotherightof the bio255 substitution and is atleastpartially de-letedbythe bio247 deletion. ralmay be located to the left of the immunity 21 substitution, as
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.500.264.456.291.365.2] [image:4.500.265.457.457.524.2]TABLE 4. Ralexpression by a defectiveXcI857prophageafterthermal inductiona
Yields of thefollowingsuperinfectingphages be-Bacterialstrain Defective prophage Temp(00) foreandafter thermalinduction ofprophage:
AbiolOimm21.0 AbiolOimm21.K
M8521 bio255cI857H1 33 2.0 x10-4 7.5
40 0.11 (275 33(4.4)
M5222 bio247cI857H1 33 6.4 x 10-4 7.5
40 9.0x 10-4(1.4) 60(8.0)
aSingle
cycles
ofgrowth were accomplished as described in the text. The numbers in parentheses are the increases in plating efficiency at 40 comparedwith the plating efficiency at33°C.Phage yields weredetermined by plating on strain Ql(rK'mK ). The phage yield determinedon strain Ql was 0.72 that determined onstrain Q8100 (r- m-).Abio252imm2' hybrids had Ral activity (Tables 2 and3). Altematively,the Ximm21 phagemay
haveahybrid Ral function partiallyencoded by
the immunity region of phage 21and partially encoded by the ralgeneof A.
In the experiments shown in Table 4, the
progenyphageproducedinthepresenceof Ral
hadan efficiency of plating of0.72 ± 0.06 on a
restrictinghost (Q1)comparedwiththe isogenic nonrestricting hostIQ8100),whereas the control forefficiency ofplating (Xvir.K)plated withan
efficiency of 0.90±0.04.Thisresult is consistent
with thehypothesisthatatleast 80% of the Ral-protectedprogenyarefullymodifiedorthat all
oftheRal-protectedphage lackmodificationat
onesite.
Ral alleviates restriction by EcoK and EcoB butnotby EcoPl. To determine whether Ralcould act on otherrestriction systems, the coinfections shown in Tables 5 and 6were
per-formed. Preinfection of the
rB+mB+
strainQ8209 with the Ral+ helper Xbio252cI857.B increased the yield of superinfecting unmodified Ximm43"cI.0byafactor of 46(Table 5). Clearly,Ral could alleviate EcoB restriction as
effec-tivelyasit could alleviateEcoK restriction.
However, in theexperiments shown in Table 6, under conditions in which Ral alleviated EcoK restrictionsubstantially,nospecific effect of Ral onP1restriction wasobserved. Atahigh
tem-perature, where Ral was expressed in the
M8521(P1) lysogen, there was a 2,750-fold in-creasein theplating efficiencyof
Ximm'.P,
butonlya7.7-foldincrease intheplating efficiency
of
Ximm32.K,
whichwasnotsignificantlydiffer-ent fromthe 2.9-fold effectonthetotally
modi-fiedcontrol(Ximm43'.K,P).
Ral isnotamethylasewithbothK
spec-ificity and B specificity. Fromthe results of
the experimentsshown in Table 4,Iconcluded
thattheprogenyresultingfromaRal-protected
infectionwereK modified. Because Ralprotects DNAfromEcoB,itwasof interesttodetermine
whether Ral was a methylase which acted
in-dependentlyof the host modification and
restric-TABLE 5.
Ral
alleviates E. coli BrestrictionaPhageyieldsofAimm43 during coinfection of anrB+mB+host Helper phage
Helper Test phage Helper Abio252cI857.B 29(11)b 0.46 (5) 46 Abiol0cI857.B 23(25) 0.004(10) 0.4 AcI857.B 43(45) 4.9 (16)
490'
ArevexbiolOcI857.B 9(45) 1.9 (9) l9odNone 0.01 (30)
a Thesuperinfecting testphage was
Ximm43.0.
Coinfec-tions wereperformedinstrain Q8209(rB+mBs)
asdescribed inthe text.Aimm'4 carriedcI6T.Aimm4 phage yields were calculated on strain Q8302(rB' mB+, A), and helper phage yields were determined on strain Q8803(rB+mB+, AimmM4).'Theyieldsinparenthesesareforthecorresponding coin-fections in which thesuperinfecting phage(Aimm4cI.B)was
modified.
'Thehelper effect was duetoRaland Red. dThehelper effectwasduetoLarand Der.
tion system to modify DNA at both K and B sites. Preinfection of the
rK-mK-
hostQ8100
with the Ral+ phage
Xbio252cI857.0
didnot re-sult inBand Kmodifications of thesuperinfect-ing
Ximm4.0
(datanot shown). However, it is conceivable that Ralmethylated
only
the paren-talphage DNA and that thisparental
modifi-cationwassubsequently
lostthrough
rounds ofreplications
under the rKmK
conditions. To observe the effect of Ralonparental phage,
the infections showninTable7 wereperformed.
In theseexperiments,Itriedtodetermine whethersuperinfecting
parental
DNA became B modi-fiedby Ral withoutenlisting
thehelp
of EcoB. Toscorethe modificationacquired
by
parental
phageDNA, infectionswere
performed
inlyso-genshomoimmunetothe
superinfecting phage.
In thisway, DNA
replication
was blockedby
therepressorofthe residentprophage,
whereas DNAmaturation andmorphogenesis
functions weresupplied by
theheteroimmune
helper
phage.In
conclusion,
whentherK+mK+
lysogen
M8002 was
superinfected
withAb2intF
red-cI-p4.K
afterpreinfection
with eitherRal+
orRal-
helper
phage,
theprogenyphages
werenot B
modified,
asthey
failed toform
plaques
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 6. Ral doesnotalleviatePl restriction Efficiencyofplatingof:f P1lysogen Temp (°C)
Aimm434.K
Aimm4.PAimm43.K,P
M8521(P1) 34 6.0x 10-6 4.0x10-5 0.38
40 4.6 x 10-5(7.7)b 0.11 (2,750) 1.1 (2.9)
M5222(P1) 34 3.7x 10-5 6.9x 10-5 0.65
40 7.7 x 10-5 (2.1) 4.9x10-4(7.1) 1.1(1.7)
aEfficiency ofplating
of
Ximm434
onP1
lysogensinthe presence(40°C) andabsence(340C)
of Ral.Aimm434
wasAimm434cI6T. Phageweretitratedasdescribed in thetext.Indicator culturesweregrown andpetriplates were incubatedatthe temperatures indicated.Efficiencyofplatingwasmeasured withrespecttothe titeron strainQ8100 (r- m-).
bThenumbersinparenthesesaretheincreases inplatingefficiencyfor each
phage/host
combinationatthe higher temperature.TABLE 7. Ral isnot amethylase which modifies parentalphage DNA upon injectiona Phage yields fromcoinfections of rK'mK'A lysogen Coinfec- tingphage Helper phage Yield of
superinfecting
phagein thefollowing Yield oftion hosts: helper
r-m- rK MK'
+MB
rB + phage1 Ab2cl-p4.K Abio252imm21.K 0.25 0.32 3.1 x10-4 13
2 Ab2cI-p4.K Abiol0imm21.K 0.32 0.38 <1x10-3 20
3 Ab2cI-p4.B Abio252imm21.K 0.09 0.09 0.02 18
4 Ab2cI-p4.B Abiol0imm21.K 9.5x 10-4 6.4x 10-4 6.4x10-4 22
a Theprocedure for the Ral rescue ofnonreplicating phage is described in the text. The MOIs of both helper phage and test phage were each 10. Helper phages carried imm21cI-; Ab2cI-p4was Ab2intlO2red3cI857p4.
Coinfections were in strainM8002,aAlysogen of bacterial strain M72. (M72 does not exhibit any MOI effect on theability to restrict unmodified phage over a MOI range of 0.25 to 15 [data not shown].)AcI-p4were scored assmallplaquesonthefollowingAimm2' lysogens: strain Q8104 (r- m-), strain Q8004 (rK+ MK+), and strain Q8304 (rB+mB+).The yields of helperphages weredetermined by plating on the A lysogen Q8002 (rK+mK').In addition,XcI-p4werescored on strain MI742, a RecA- host (data not shown). Yields wereequivalent to the yieldsfound with strainQ8004; thus,AcI-p4did not rescue the bioregion of the helper phage.
on an
rBnmB+
host. As maturation ofunrepli-cated
phage
requires
recombinationtoproduce DNAof maturablelength (46),
therewas a ques-tionastowhethertheputative
EcoB modifica-tion of theparental
superinfecting
DNA could havebeen lostby
recombinationwith therepli-cating
helper
DNA.Thatthiswasunlikely
wasshown by additional
experiments
(Table
7,co-infections 3 and 4), in which the test
phage
already
carriedaBmodification. Incoinfection 3,Ab2int-red-cI-p4.B
phage
maturedunderre-stricting
conditionsbut,
inthepresenceofRal,
retainedparentalmodification(oratleastagood fractiondid,astheyields calculatedon the iso-genicrB+mB+andrK+mMK
strainswere0.02and 0.09, respectively). Since parental modificationwasretainedontheRal-protected phage
during
the necessary recombination and maturation steps, it islikely
that any de novo Ral modifi-cation of EcoB sites onAb2int-redc-p4.K
(Table7,coinfection 1) would also have survived subsequent recombination and maturation ifit had actuallyoccurred. Thus,it is unlikelythat Ral is a methylase which can recognize EcoB and EcoK sites
independently
of thehost mod-ification restrictionsystem.A-P22 hybrid ASpi-imm"2
c25(OP)'P22
and 480 do notexpress Ralfunctions. S. typhi-murium has a modification andrestrictionsys-tem (hsdSB) which is allelic to the systems of E. coli K-12 and E. coli B (53). Because the arrangementofgenesinSalmonella phage P22 is like thearrangement in A (4), it wasofinterest
todetermine whether P22 carried a Ral function in a positionhomologoustothe position of Ral in A. TheA-P22 hybrid
ASpi-immP22c25(OP)
P22 carries a substitution of P22 DNA extending from redthrough the ren genes of A (Fig. 1) (22, 50). Since this hybrid carries the P22 erf gene, it was necessary to test Ral activity on a phage whoserestrictedfragmentscould not be rescued by Erf-promotedrecombination (Table 8). The unmodified test phage chosen wasAh'attuo.
cI857.0, which has at least two K restriction sites in DNA segments nonhomologoustotheA-P22 hybrid; one of thesesites(sk480)segregates with h80, andtheother (sk2)segregateswith immu-nity of A (39).Hence, progeny phage were scored asbeing measured by Ral if they had both the immunity of A and the host range of480. Under conditions in which preinfection by Abio. 252imm2cI-.K stimulated theformation of
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.500.64.459.228.322.2]TABLE 8. ASpi-imm2(OP) 22is
phenotpically
Ral 'Infective centers made by thefollowingsuperinfecting phages during coinfections: Helper phage
Wh8att"O.O Aimm434.0.
ASpi-immF22(OP)F22.K 1.3 x 10-3 (0.21) 0.23b(0.60)
Abio252imm21.K 0.23 (0.40) NTc (NT)
AbiolOimm21.K 7.3 x 10-4 (NT) NT (NT)
None 1.1 X 10-3(0.67) 4.0x10-4 (0.90)
a Coinfections were
performed
in bacterialstrain QD5003 as described in thetext. Infective centers of helper phages were determined on strain Q8003 (data not shown, but all were approximately 1). The numbers in parentheses are the yields inparallel coinfections of the corresponding modifiedsuperinfectingphages. Infective centers ofsuperinfecting Ximm4cI6Twere assayed in strainQ8004, and infective centers of superinfectingphageAh'att'cI857were assayed in strainQA8002. bErf-promoted rescue of
imm.
cNT, Not tested.
fectivecentersbysuperinfecting
Ah8°att*cI857.0
200-fold, ASpi-imm 2c25(OP)P hadno effect.
The rescue of
Aimm'.0
(a phage with moreextensivehomology with the A-P22 hybrid) was
probably duetoErfrescueof the immunity434
region of restricted DNA. The lack ofrestriction alleviationby the A-P22 hybrid wasnotdueto
expression of an inhibitor in the P22
immun-ity regionorto insufficientexpression ofaRal
from the P22 leftward promoter, because
Abio252immP`(OP)m,
whichcontains the P22promoter, could supply the Ral function of X during coinfection (datanotshown).
Similarly, another lambdoid phage (480) was
tested for restriction alleviation. In the
experi-mentsshown in Table9, W80didnotexpress a
Ral function. Preinfection with 480vir did not preventrestriction of
Aimm".0
byEcoK,astheyield of
Aimm'
wasnotincreasedby the pres-enceof the40virhelper.Xrev has a restriction alleviation
func-tion, Lar. Arev (isolated as a revertant of Ared gam-, which formed plaques on
RecA-hosts) carries a substitution of foreign DNA
between b2 and cIII which codes for a new
attachment site, as well as an integrase and
recombination function (Fig. 1)(17, 59).Arevhas rescued therecEgenefrom E. coli andexpresses
its product, exonuclease VIII (ExoVIII), upon
infection (13), which gives risetothe
Der'
phe-notype (Redin reverse) and theability of this
"Red"gam-phagetoplateunder RecA-
con-ditions. The recE gene is expressed in sbcA mutants of E. coli (49). sbcA- mutants were
selectedasmitomycin C-resistant, Rec' revert-ants of recB recC- strains (3) and have the
additional phenotype that they exhibit a
sub-stantiallyincreasedplating efficiencyofA.0(44). ThesedatasuggestedthatArevwasprobably
created by arecombination event with E. coli
DNA which picked up the recE gene and a
nearby"int"gene,indicatingthatthe sbcA locus
may be part of a cryptic prophage (17). The
TABLE 9. 480isphenotypically Rala Phage yields on coinfection Helper phage
Yield of helper Yield ofXimm434
4080
105(135) 0.02(12)None 0.04(55)
aThesuperinfectingphagewas
Ximm4.0.
Thecoin-fectionswereperformedinbacterial strainQD5003as described in the text. Yields in parentheses are for
parallel coinfections where thetestphagewas modi-fied. Ximm43 wasXimmmcI6T, and
080
was p80vir. Theyields ofAimm43 were determinedonthe O'R host strain Ql, and080vir
were assayed on strain QD5003.increased plating efficiency of A.0 on sbcA-strains indicates that this cryptic prophage might also express a Larfunctionanalogousto the Ral function of A. To test this hypothesis, plaque-forming phagewereselectedon arecA
-host fromastock of the Ral- phageAbiolOcI857 (see above). This Xrev exbiol0 had the same propertiesaspreviously isolated Arev phages(17, 59): (i) it grew in recA- andpoLA- bacterial strainsand P2lysogens (60), (ii) it
plated
onOM mediumplates containing citrate, indicating that it hadanetdeletion ofphage DNA. (iii) andit grewpoorlyat ahightemperature(430C).
Xrev exbiolOcI857 appeared to have a Lar function which
prevented
restriction ofthe su-perinfecting Ah'bio243imm434.0
phage (Table 10). The numberofrK+mK'
cells infected withXh80bio243imm434.0
whichformedinfectivecen-tersincreased50-foldinthepresenceofthe Arev helperphage, when Xh
imm434
infectivecenterswereselected for(Table 10).Thiswas
probably
aminimum Lar effect since coinfection with the Arevhelper
phage
reduced theprobability
of infective center formationapproximately
five-fold when thesuperinfecting phage
wasmodified (Table 10). TheseAh8'imm'
phagesprobably
retained the bio substitution, as noAh'-revimm
43
recombinantswerescoredasinfectiveon November 10, 2019 by guest
http://jvi.asm.org/
[image:7.500.250.446.234.300.2]TABLE 10. Arevisphenotypically Ral+a
Infective centers and yields producedbythefollowing phagesduringcoinfection
Helperphage Xh`bio243iM4"b
ArevimM41
Infective Yield Infective centers Yield
centers
XrevexbiolOcI857.K 0.05(0.08) 35(27) 0.02(0.02) 0.7(0.7) None 0.001(0.38) 208(116) <5.3x 10-5(<3.1x 10-4)
aThesuperinfectingphagewas
AhJ°bio243imm4
'.0.
Coinfectionswereperformed
asdescribed in thetext. Ah5'bio243imm4' carriedthecI6Tmutation.Thenumbersinparenthesesarefromparallelcoinfectionswhere thesuperinfecting phagewasmodified. Infective centersweredeterminedonrKW
mK'
hosts.bInfective centers were determined on strain QA8002 (AR,
Ah"°cI857).
Plaqueswerevery tinydue to the heteroimmune prophage exclusion exhibited by certain Xh52 prophages against certainAh'
superinfecting phages (P. Toothmanand D.Forbes,unpublisheddata).Noinfective centers weredetectedonstrainMI8000XR (recA ,Alysogen),indicatingthat theAh imm4&4infectivecenters onstrainQA8002carriedbio243 andnot rev.Thehelpereffect was due toLar.
'Infectivecentersandyieldsof revimm43 recombinantswereassayedonstrainMI8000(Rec-, A).Thehelper
effectwasdue toDer.
centers onthe appropriate selection host (data
not shown). It is unlikely that the
Ah8sbio243imm434
progenywererescued by the Der recombination function, unless for someotherreasonEcoKfailed to restrict both thesite
inthe bio substitutionand the site(sk2)between bio243 and imm434. However, if such were the case, Xhsebio243imm43 should have behavedas
ifit had only one restriction site (skl), and it wouldnothave beenrestrictedsoefficiently (i.e.,
only 50-fold insteadof1,000-fold) (39).
Lar and Der haveverydifferenteffectsonthe average burst size of the phages which they
rescue, where averageburst is defined asyield per infective center. The average burst of
Xh8simm434,
Lar-protected phage was large(equivalenttothe burstofthemodifiedcontrols) (Table10). Theaverageburst of theXrevimm43 recombinantsrescuedby Derwassmall, approx-imately 1(Table 10). This differencemayreflect
the times that Lar and Deractduringinfection (early and late, respectively). However, from these data it isimpossible totell whether Der acted before or after Lar or evenwhether Der
actedin thesamecellsasLar.
Table 11 presents further evidence that Lar
andDerareseparatefunctions. Gillenetal. (14)
isolated Rec-UV-sensitive revertants of
recB-recC- sbcA mutants in an attempt toisolate mutations in the recE structural gene. Three
such revertantswhich containedno detectable
ExoVIIIprotein are listed inTable 12. Two of
these (JC8689 and JC8690) simultaneously
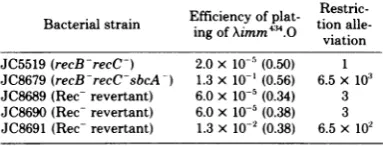
re-verted to the stringent restriction phenotype. Thesemutantsmaybe sbcA+revertantsor
dele-tions of recEandlar.However,JC8691 retained itsrestrictionalleviationactivity despite its loss of ExoVIII. Thus, Lar activity expressed in
sbcA-strains isseparablefrom,oratleast does
notrequire,ExoVIII activity.
TABLE 11. Restriction alleviationactivity ofsbcA-strainsofE. coli isnotduetoExoVIIIactivitya
Bacterial strain EfficiencY
of
plat- tionalle-viation
JC5519(recB-recC-) 2.0x10-5(0.50) 1 JC8679(recB-recC-sbcA-) 1.3x10'(0.56) 6.5x103 JC8689(Rec- revertant) 6.0x10-5(0.34) 3 JC8690(Rec-revertant) 6.0x10-5 (0.38) 3 JC8691(Rec-revertant) 1.3x10-2(0.38) 6.5x102
a Phage were titratedasdescribed in the text.Aimm43 was Aimm4cI6T. The numbers in parentheses are theplating
[image:8.500.267.459.293.366.2]efficienciesofAimm44.K.Restriction alleviation was measured withrespecttotheplating efficiencyonstrainJC5519. Bac-terial strainsJC8689,JC8690,andJC8691 were Rec- revert-antsofstrainJC8679 and containednoenzymaticallyactive ExoVIIIprotein(JaneGillen,personalcommunication).
TABLE 12. Ral isexpressed inAB1U57a
Phageyields during
coinfections Helper
Helper phage Hle
Yield of Yield of effect helper Ximm434
Abio252cI857.K 17(16) 0.02 (9) 20 AbiolOcI857.K 15(20) 0.001(6) 1
ArevexbiolOcI857.Kb 32(32) 1.1(7) 1,100
None 0.001(55)
aThe superinfecting phage wasAimm434.0. Coinfections
wereperformed in strain AB1157 as described in the text.
Yields ofhelper phagesweredeterminedon strain Q8003, and
yieldsofAimm43 weredeterminedon strain Q8002.Aimm3 carriedcI6T.Yields in parenthesisarefrom parallel coinfec-tions wherethesuperinfectingphage wasmodified.
'The coinfectingphagesupplied Derfunction as wellas
Lar.
TheArevisolated by Zissler et al. (59) and the Arevisolated byGottesmanet al. (17) also
sup-plied
Lar during coinfections and alleviated EcoB restriction.Ral
activity
does not require transacti-vation ofhost Lar activity. To test whetheron November 10, 2019 by guest
http://jvi.asm.org/
[image:8.500.267.458.446.529.2]GENETICS
Ral
activity requiredhost Larfunction,the coin-fection experiments describedinTable12were done. The coinfection host was AB1157. The observationsthat sbcA - mutantscannotbe iso-lated inAB1157(49), thatXrev cannot lysogenize AB1157efficiently(17), and thatAB1157is Rac-(35) all suggestedthat AB1157was deletedfor the cryptic prophage containing recE and lar. Recently, Kaiser andMurray (25) have shown that Xrevis formedbyrecombination with the Rac prophage, and these workers have con-firmed that the Rac prophage is deleted in AB1157. AsTable 12shows, bothAbio252cI857 and Xrev exbiolOcI857 express restriction alle-viation activityincoinfectionexperimentsin an AB1157genetic background. Therefore, the re-striction alleviation activity expressedinE. coli K-12 strains after infection byAbio252cI857orXrev
exbiolOcI857 does not result from the de-repression of the hostLarfunctionunder sbcA control.DISCUSSION
Ralexpression is under controlof thecIgene inA and isexpressed early during infection by Xral+-modified phage. Unmodified phagewhich superinfect a Xral+-infected cell aremuch less likelytoberestricted,and theprogenyofthose phage which do escape restriction arefully or almostfully modified.
TheRalfunction ofA canpreventrestriction of unmodified phage DNA by the EcoK and EcoBenzymes.Ral doesnot seem tobe a meth-ylase which can recognize and methylate the
appropriate
bases within theA(N)8TGC
recog-nitionsequence foundinbothEcoK and EcoB sites(26,31,40). If thiswereso,the nonreplicat-ing phage whichmaturein the presence ofRalshould
haveboth EcoK and EcoB modifications(Table
7). Thus, thespecificity
ofmodification whichoccursinthe presenceofRalisthe spec-ificity definedby
thehostmodification and re-striction system.TheobservationthatRalcansavephagefrom restrictioninthe absenceofrecombination func-tions is consistent with the observation of Za-beau et al. (58) that Ral actsto rescue entire genomes, since it protectsnonhomologousphage P2 fromrestrictionatlowMOIs.
Under Ral- conditions, theRec and Red re-combination systemsseemtodiffer in theability torecombinehelper phagewithrestricted frag-ments. Thesimplest interpretationof the data inTable8is that theX-P22
hybrid
doesnothave aRal functionanalogoustothe Ral function ofA,
whichcanprevent restrictionof nonhomolo-gous DNA, at least under the conditions in which the XRal function is active. The Red function of thishybrid (Erf
of P22) caneffi-ciently rescue the immunity region of phage
Aimm434,
indicatingthat Red canrecombine re-stricted fragments andhelper phages. There is no evidence that Rec can perform similarly, sinceinfective centers (which shouldbethe most sensitive indicator of such a phenomenon, as opposed to phage yield) are not increased sig-nificantlyby aXred-ral-
helper,comparedwith nohelperatall. This may be due toexpression of inhibitorsofrecBC nuclease,such asgam,by theunmodifiedtest phage used in these experi-ments,or, morelikely,it is anintrinsic property of the Recrecombinationsystem(perhaps,sim-ply, restricted fragments are better substrates for nuclease digestion than for recombination).
In vitrostudiesonthe kinetics ofmodification andrestriction of unmodifiedDNA byEcoBand EcoKhaveshownthat it ismodified slowly and restrictedrapidly (55). SeveralwaysRal might stimulate modificationattheexpenseof restric-tion are as
follows.
Four stages have been rec-ognizedinthe restriction process. First,S-aden-osylmethionine
is bound to the enzyme which undergoes activation; this form of the enzyme bindsnonspecifically
to DNA. Next, ATP is bound, and theenzyme undergoesaconforma-tionchange, binds
specifically
andtightlytothe unmodified DNA, and, in the continued pres-ence of ATP, restricts the DNA (20, 57). Ral might interactdirectly
with the restrictionen-zyme
complex
toproduce
anr-m+phenocopy,
resembling the hsdR mutant EcoK-18, which forms short-lived
recognition
complexes
that cannotbestabolized by ATP andcannotrestrictunmodified
DNA(DNA.O),
butstill can meth-ylateit (6, 20). Debrouwere etal.(9)
have sug-gested that theRal
effect maybe mediatedby
adirectinteractionwith rho. Evidence for such aninteraction
is allindirect,
but it is an inter-esting model.Invitro,EcoK and EcoB restrict unmodified DNAin the presence of
S-adenosylmethionine,
ATP, and
Mg2+,
butEcoK and EcoBcanmodify
DNA.0
if ATP and Mg2+ are left out of the reactionmixture(19, 37).Thus,potentially,
Ral mightact tostimulate
modificationby
lowering
ATP
concentrations
orMg2+ concentrationsorbothin the
cells.
Anotherpossibility
isthatRal may be aDNA-binding
protein
that interacts with DNA at the EcoB and EcoKrecognition
sitesto change thelocal structure of the DNA and make the DNAa morefavorablesubstrate formodificationoralessfavorablesubstrate for restriction.The Lar
function
expressed by
Arev and sbcA -strains maypreventrestrictioninafashion anal-ogoustothe Ral function of A. The observation that A can rescue the reverse functions(an
in-tegrase, ageneral
recombination functionVOL. 38,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[Der], and a restriction alleviation function
[Lar])
from many strains of E. coli suggestedthat revispartof a lambdoidprophage.
Subse-quently, KaiserandMurray (25) demonstrated
thatrevishomologousto the Racprophage(33,
35), which isvery closeto the sbcA locus (34). Thus, the recE geneis involved in(i)
recombi-nation during transfer ofrac+ by Hfr mating with rac-recB- orrac- recC-recipients (35),
(ii) recombination proficiency in recB-
sbcA-and recC sbcA strains(30, 49),and(iii)
recom-binationproficiency byArev(13).Itis notknown
whether theLar expressedin sbcA- strainsand
byXrevis codedforbyagenedistinctfromrecE.
However,itis clear that
ExoVIII,
whichiscodedforbythe recEgene (29),is notrequiredforLar
activity.The recEgeneproductwitha
molecu-lar weight of 1.4 x
105
may contain separatedomains for theexoVIIIandLaractivities (13).
It remains to be determinedwhether recEand
larareseparategenes.
ACKNOWLEDGMENTS
I thank Ira Herskowitz for histhoughtful tutelage and
providinganatmospherewhereideasandgraduatestudents flourished. I thank Jane Gillen for bacterial strains, Jane
Gillen,Marc vanMontagu,MarcZabeau,andJeffSchellfor
discussion andcommunication oftheirunpublished data,and KimPersons andPaulaBortle forpreparationof the
manu-script.
Thiswork wassupportedbyaPublicHealthService re-searchgrantAI11344to I.HerskowitzandbyaPublicHealth
ServiceBiochemistry TrainingGrant to theDepartmentof
Chemistry,UniversityofOregon.
LITERATURE CITED
1. Adler,S.P., and D.Nathans. 1973. Studies of SV40
DNA. V.Conversion ofcircularto linearSV40DNAby
restriction endonuclease from Escherichia coli B.
Biochim.Biophys.Acta299:177-188.
2. Arber, W., and D. Dussoix. 1962.Host specificityof
DNAproducedbyE.coli.I.Host-controlled modifica-tionofbacteriophagelambda. J.Mol. Biol.5:18-36.
3. Barbour, S., H. Nagaishi, A. Templin, and A. J.
Clark. 1970.Biochemical andgeneticstudies of
recom-binationproficiencyin Escherichiacoli. II. Rec+
re-vertantscausedbyindirectsuppressionof Rec-
muta-tions. Proc.Natl.Acad.Sci.U.S.A.67:128-135.
4. Botstein, D.,R. K. Chan, andC. H. Waddell. 1972.
Genetics ofbacteriophageP22. II. Gene orderandgene function.Virology49:268-282.
5. Boyer,H.W.,andD.Roulland-Dussoix.1969. A
com-plementationanalysisof the restrictionand
modifica-tion ofDNAinEscherichia coli. J.Mol.Biol.
41:459-472.
6. Buhler, R., and R.Yuan. 1978. Characterization ofa
restrictionenzymefromEscherichiacoli Kcarryinga
mutation inthemodificationsubunit. J. Biol.Chem.
253:6756-6760.
7. Campbell, A. 1957. Transduction and segragation in
EscherichiacoliK12.Virology4:366-384.
8. Davison, J.,andF.Brunel.1979.Restriction insensitiv-ityinbacteriophageT5.II. LackofEcoRImodification
inT5+andT5rismutants. J.Virol.29:17-20.
9. Debrouwere,L.,M.VanMontagu,and J.Schell.1980.
TheralgeneofphageA. III.Interferencewith E.coli
ATPdependentfunctions.Mol.Gen.Genet.179:81-88.
10. Debrouwere,L., M.Zabeau, M. VanMontagu,andJ.
Schell.1980.The ralgeneofphageA. II.Isolationand
characterization of ral-deficient mutants. Mol. Gen.
Genet.179:75-80.
11. Dussoix, D., and W. Arber. 1962. Hostspecificity of
DNAproduced byE. coli.II.Controloveracceptance
of DNA frominfecting phagelambda.J. Mol. Biol.5:
37-49.
12.Georgopoulos, C.P., andH. R. Revel. 1971. Studies
withglucosyltransferasemutantsof the T-even bacte-riophages. Virology 44:271-285.
13. Gillen, J. R., A. E. Karu,H.Nagaishi, and A. J. Clark.
1977. Characterizationof thedeoxyribonuclease
deter-minedbylambdareverse asexonucleaseVIIIof E.coli.
J. Mol.Biol.113:27-41.
14.Gillen, J. R., D. K. Willis, and A. J. Clark. 1981.
Geneticanalysisof theRecEpathway of genetic recom-binationin Escherichiacoli K-12. J. Bacteriol. 145:
521-532.
15. Glover, S. W. 1970. Functionalanalysisofhost-specificity
mutantsin Escherichiacoli. Genet.Res.15:237-250.
16.Goldberg,A.R., and M. Howe. 1969. New mutationsin
theScistronofbacteriophage lambdaaffecting host
celllysis. Virology38:200-202.
17.Gottesman,M.M., M. E.Gottesman,S.Gottesman,
and M. Gelbert. 1979. Characterizationof bacterio-phageAreverse as anEscherichiacoliphage carrying
auniqueset of host-derivedrecombinationfunctions.J.
Mol. Biol.88:471-487.
18.Greer,H. 1975.Thekil geneofbacteriophage lambda. Virology66:589-604.
19. Haberman,A.,J.Heywood,andM.Meselson. 1972. DNA modification methylase activity ofEscherichia colirestrictions endonucleases K and P. Proc. Natl.
Acad.Sci.U. S.A.69:3138-3141.
20. Hadi,S.M.,T.A.Bickle,andR. Yuan.1975. TIhe role
ofS-adenosylmethioninein thecleavageof
deoxyribo-nucleicacidbythe restrictionendonucleasefrom Esch-erichiacoli K.J. Biol. Chem.250:4159-4164. 21. Herskowitz,I., andE. R. Signer. 1974. Substitution
mutationsinbacteriophage Xwithnewspecificity for
lategeneexpression. Virology61:112-119.
22. Hilliker, S.,andD.Botstein.1976.Specificityofgenetic
elementscontrolling regulation ofearly functionsin temperate bacteriophage.J. Mol.Biol.160:537-566.
23. Horiuchi,K.,andN.D. Zinder. 1972.Cleavageof bac-teriophagefl DNAbythe restrictionenzymeof
Esch-erichiacoli B. Proc.Natl.Acad.Sci. U.S.A.
69:3220-3224.
24. Hubacek, J.,andS. W.Glover.1970.Complementation analysisoftemperature-sensitivehostspecificity mu-tationsinEscherichiacoli. J.Mol.Biol.50:111-127.
25. Kaiser, K.,andN.E.Murray.1979.Physical
character-izationof the"Racprophage"in E.coli K12.Mol.Gen. Genet.175:159-174.
26. Kan,N.C.,J.A.Lautenberger,M. H.Edgell,andC.
A. Hutchinson.1979.The nucleotidesequence recog-nizedbytheEscherichiacoli K12restrictionand
mod-ificationenzymes. J. Mol. Biol.130:191-209.
27. Kruger,D.H.,C.Schroeder,S.Hansen,andH. A.
Rosenthal.1977. Activeprotection by bacteriophages
T3 andT7againstE.coli B-andK-specificrestriction
of theirDNA.Mol.Gen.Genet.153:99-106.
28. Kuhnlein, U.,S.Linn,andW.Arber.1969.Host
spec-ificity of DNAproduced by Escherichiacoli. XI. In
vitro modificationofphage fd replicativeform. Proc.
Natl.Acad.Sci.U.S.A.63:556-562.
29. Kushner, S.,H.Nagaishi,and A.J.Clark. 1974.
Iso-lationofexonucleaseVIII:theenzymeassociatedwith
the sbcA indirectsuppression. Proc. Natl.Acad. Sci.
U.S.A.71:3593-3597.
30. Kushner,S.R.,H.Nagaishi,A.Templin,and A.J.
Clark. 1971. Genetic recombination in Escherichia
coli:the roleofexonuclease I.Proc. Natl.Acad.Sci.
on November 10, 2019 by guest
http://jvi.asm.org/
U.S.A.69:1366-1370.
31. Lautenberger, J. A., N. C. Kan, D. Lackey, S. Linn, M. H.Edgell, and C. A. Hutchinson. 1978. Recogni-tion site ofEscherichia coli B restriction enzyme on
oXsBl and simian virus 40 DNAs: an interrupted se-quence. Proc. Natl. Acad. Sci. U.S.A.75:2271-2275. 32. Linn, S., and W. Arber. 1968. Host specificity of DNA
produced byEscherichia coli. X. In vitrorestriction of phage FDreplicativeform. Proc. Natl. Acad. Sci. U.S.A. 59:1300-1306.
33. Lloyd, R. G.1974.Thesegregation of the sbcA and Rac phenotypes in an Escherichia coli recB- mutant. Mol. Gen. Genet. 134:249-259.
34. Lloyd, R. G., and S. D. Barbour. 1974. The genetic location of the sbcA gene of E. coli. Mol. Gen. Genet. 134:157-171.
35. Low, B. 1973. Restoration by the rac locus of recombinant formingability in recB- and recC- merozygotes of Esch-erichia coli K12. Mol.Gen. Genet. 122:119-130.
36. Manly, K.F., E. R. Signer, and C. M. Radding. 1969. Nonessential functions of bacteriophage A. Virology 37: 177-188.
37. Meselson, M., and R. Yuan.1968.DNArestriction
en-zymefrom E. coli.Nature (London) 217:1110-1114.
38. Murray,N. E., P. L. Batten, and K. Murray. 1973. Restriction ofbacteriophage A by Escherichia coli K. J. Mol. Biol. 81:395-407.
39. Murray, N.E., P. Manduca de Ritis, and L A. Foster. 1973.DNA targetsfor the Escherichia coli K restriction systemanalyzed genetically in recombinants between phages Phi 80 and lambda. Mol. Gen. Genet.
120:261-281.
40.Ravetch,J.V., K.Horiuchi, and N. D. Zinder. 1978. Nucleotide sequence of the recognition site for the restriction-modification enzyme of Escherichia coli B. Proc. Natl. Acad. Sci.U.S.A.75:2260-2270.
41. Roberts, R. J. 1978. Restriction and modification
en-zymesand theirrecognition sequences. Gene4:183-193. 42.Signer, E. R., K. F.Manly, and M. A. Brunstetter. 1969.Deletion mapping of the cIII-N region of bacte-riophageA. Virology39:137-140.
43. Signer, E. R., and J. Weil. 1968. Recombination in phage A. I. Mutants deficientingeneralrecombination. J. Mol. Biol. 34:261-271.
44.Simmon,V.F.,and S.Lederberg.1972.Degradationof bacteriophage lambdadeoxyribonucleic acid after
re-strictionby Escherichia coli K-12. J. Bacteriol. 112:
161-169.
45. Sommer, R., and H. Schaller. 1971. Nucleotide
se-quenceof therecognitionsiteof theB-specific restric-tionmodification system in E. coli. Mol. Gen. Genet. 168:331-335.
46. Stahl,F.W.,K. D.McMilin,M.M.Stahl,R. E.Malone, and Y.Nozu. 1972. Arole forrecombination inthe
productionof"free-loader" lambdabacteriophage
par-ticles.J. Mol. Biol. 68:57-67.
47. Studier, F. W. 1975. Gene 0.3 of bacteriophage T7 acts to overcome the DNArestriction system of the host. J. Mol. Biol. 94:283-295.
48. Studier, F. W., and N. R. Movva. 1976. SAMase gene of bacteriophage T3 is responsible for overcoming host restriction. J. Virol.19:136-145.
49.Templin, A., S. R. Kushner, and A. J. Clark. 1972. Genetic analysis of mutations indirectly suppressing recB and recC mutations. Genetics72:205-215. 50. Toothman, P., andL.Herskowitz. 1980. Rex-dependent
exclusion of lambdoid phages. II. Detenninants of sen-sitivitytoexclusion. Virology 102:147-160.
51. Toussaint, A. 1976. The DNAmodification function of temperatephageMu-i.Virology 70:17-27.
52. VanOrmondt, H., J. A. Lautenberger, S.Linn,and A.de Waard. 1973. Methylated oligonucleotides de-rived frombacteriophage fd RF-DNA modified in vitro by E. coli Bmodification methylase. FEBSLett.33: 177-180.
53.Van Pel,A., andC. Colson. 1974. DNA restriction and modificationsystemsinSalmonella. II. Genetic com-plementation between the K and B systems of Esche-richia coli and the Salmonellatyphimurium system SB, with thesame chromosomal location. Mol. Gen. Genet. 135:51-60.
54. vonGabain, A., G. S.Hayward,andH.Buyard.1976.
Physical mapping of the hindI11, EcoRI, Sal, Sma restriction endonucleasecleavage fragments from bac-teriophage T5 DNA. Mol.Gen. Genet. 143:279-290.
55. Vovis, G. F., K. Horiuchi, and N. D. Zinder. 1974.
Kinetics ofmethylation of DNA by a restriction endo-nuclease fromEscherichia coli B. Proc. Natl. Acad. Sci. U.S.A. 71:3810-3813.
56.Weigle, J.,M.Meselson,and K.Paigen.1959.Density alterationsassociated withtransducingability inthe bacteriophage A. J. Mol. Biol.1:379-386.
57. Yuan, R., T. A.Bickle,W.Ebbers,andC. Breck.1975.
Multiple steps in DNA recognitionby restriction
en-donuclease from E. coli K. Nature(London)
256:556-560.
58. Zabeau,M., S.Freidman,M. Van Montagu,and J. Schell.1980.The ral gene ofphageA. I.Identification of anon-essential gene that modulates restriction and modification in E. coli. Mol. Gen. Genet. 179:63-74. 59. Zissler, J., E.Signer,and F. Schaefer.1971.The role
ofrecombination ingrowthofbacteriophagelambda. I. Thegamma gene, p.455-468. InA.D.Hershey (ed.), Thebacteriophage lambda. ColdSpringHarbor Labo-ratory,ColdSpring Harbor,N.Y.
60. Zissler,J.,E.Signer,andF.Schaefer.1971.The role ofrecombination ingrowthofbacteriophagelambda. II. Inhibition ofgrowthby prophage P2,p.469-475.In
A.D.Hershey(ed.),Thebacteriophagelambda. Cold Spring HarborLaboratory, ColdSpringHarbor,N.Y.