Vol.44, No. 2 JOURNALOFVIROLOGY, Nov. 1982,P.736-741
0022-538X/82/110736-06$02.00/0
Copyright©1982, American Society for Microbiology
Characterization
of Two Conformational Forms of the Major
DNA-Binding Protein
Encoded by Herpes Simplex Virus 1
DAVID M. KNIPE,* MARGARET P. QUINLAN, AND ANNE E. SPANG
Departmentof Microbiology and Molecular Genetics, Harvard Medical School, Boston, Massachusetts 02115
Received 3 May 1982/Accepted 13 July 1982
We have resolved two electrophoretic species of the major DNA-binding
protein,infected cellpolypeptide 8 (ICP8), encoded by herpes simplex virus1.In
pulse-chase experiments, we observed the conversion of the ICP8a form, the
slower migrating species, to the faster migrating form, ICP8b. Thus, the two
speciesappear toberelatedasprecursor-product. The conversionwasnotdueto
proteolytic cleavage, because higher concentrations of reducing agents in the samplebuffer shifted the faster moving formtotheslowermoving species. Also, thetwoforms have identical peptide patternsasanalyzed bypartialproteolysis in sodium dodecyl sulfate. Thus, the faster moving species appears to be a
conformational isomer containing intramolecular disulfide bonds. The functional
significance of thetwoforms of theprotein is discussed.
Herpessimplex virus (HSV) encodes amajor DNA-binding protein (2, 14, 15, 18)as abetaor
delayed early geneproduct (8, 9). This protein has been called ICP8 (infectedcellpolypeptide 8), VP130 (viral protein of molecular weight 130,000), or ICSP 11/12 (infected cell-specific
protein 11/12) (2, 8, 9, 11, 12, 14, 15, 18). Two
lines ofevidence indicate that theprotein playsa
role in the replication ofviral DNA: (i) mutant
viruseswhich encode adefective DNA-binding
protein or whose temperature-sensitive lesions
mapinornear the ICP8gene are defective for
DNA replication (4, 14); and (ii) monospecific
antiserum to the HSV type 2 (HSV-2) protein inhibits DNA replication in vitro but does not
neutralize the viral DNA polymerase activity (14). The HSV-2 protein ICSP 11/12 has been
reportedtobindmoretightlytosingle-stranded
DNAthantodouble-strandedDNA(16), and the
purified HSV-1 and HSV-2 DNA binding
pro-teins can enhance the denaturation ofa poly-deoxyadenylic acid-polydeoxythymidylic acid duplex (14). In infected cells, the protein is localized throughaseriesofstages to the
nucle-us, where itfinally binds to viral DNA (6, 10, 13). Thus, ICP8 appears to be involved in the
replication of HSV DNAin the cell nucleus.
Noposttranslationalmodifications of this
pro-tein have been reported. It appears to lack
phosphorylation (18). Others havereportedtwo
closelymigratingproteinbands nearthe size of
ICP8 in the bound fraction from a DNA-cellu-lose columnwhen extractsfrom HSV-2-infected
cells wereused(16, 18).Inthiscommunication,
wedefine therelationship between two
electro-phoretic forms of the HSV-1 ICP8 protein
mole-cule.
Weobservedapair ofcloselymigratingbands in the region of a polyacrylamide gel where ICP8, the major DNA-binding protein encoded by HSV, migrates (Fig. 1). Thetwobandswere presentinextractspreparedfrom cells labeledat 4hpostinfection. The molecular weights of the
two bands were determined to be 127,000 and 125,000 by comparison with the migration of proteins of known molecular weight (not
shown). Becausewe believe thesetwobandsto
betwodifferentforms of ICP8,wehave named
the slowermigrating band ICP8aand the faster
migratingband ICP8b.
Thetwo bandswere both presentinextracts
of cells immediately after a 5-min pulse label
(Fig.1).Duringthe 2-h chaseperiod,therewas a
progressive conversion to the ICP8b form so
that by 2 h of chase, the ICP8b form was the predominant form. Figure 2 shows the
micro-densitometertracingof the ICP8 region ofagel from a similar pulse-chase experiment. During
the chaseperiod,the amountof the ICP8aform decreased, and the amount of the ICP8b form increased.Thus, itappearedthat theICP8a form
wasthe precursor to the ICP8b form, although
theconversion was nevercomplete.
The time of expression of the two bands
correspondedtothatforabetaordelayedearly
gene inthatbothwere labeledatmaximalrates
at3 to 6hpostinfection (not shown). Also,the
period of expression of the two species was
extended to at least 10 h postinfection by the incubation of infected cells in medium
contain-736
on November 10, 2019 by guest
http://jvi.asm.org/
NOTES 737
*-- mw. _ _ _
UP _ -:_
q:
:r
:.:u,;m'
ing phosphonoacetate (not shown). This drug blocks viral DNA replication, lowers or
com-pletely inhibits the expression of late proteins, andprolongs the expression of beta proteins(4). Thealphaorimmediate early viral proteinICPO,
knowntomigrate in thesameregionof thegel,
migrated slightly faster than this doublet and
was not labeled with these kinetics. Thus, the twoprotein species, ICP8a andICP8b, possess
Cp theproperties ofabeta protein.
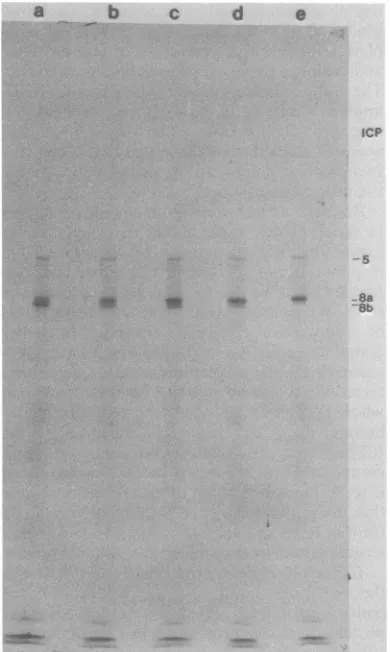
Becausewehad prepared the cellularextracts in the previous experiments in sample buffer containing 0.5% mercaptoethanol before elec--~5 trophoresis, we determined whether the two
6 speciesmight be duetoincomplete reductionof
8a ICP8. We treatedextractswithincreasing levels
8b
iob ofdithiothreitol
(DTT)
in an attempt toensureTlOalla complete reduction of ICP8. The extract
pre-lib pared in 0.5% mercaptoethanol largely
con-15 tained the ICP8b form (Fig. 3, lane a). However,
when DTT was added to the extracts before boiling, the ICP8b form was converted to the ICP8a form. The conversion was also more
complete as higher levels of DTT were added
(Fig. 3, lanes b through e). These results argue -24 that the ICP8bformisanincompletely reduced
25 form of ICP8 and migrates faster duetoafolded
structureheldby intramolecular disulfide bonds.
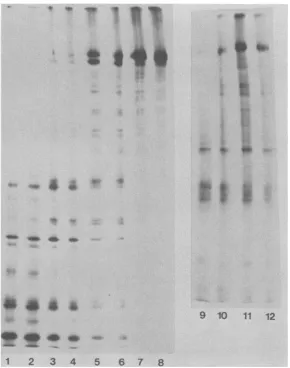
To further examine the relationship between the two polypeptide species, we excised the
bandsfromunfixed, dried gels and subjected the proteinstopartial proteolysis insodiumdodecyl
-36 sulfate by the method of Cleveland et al. (3).
CleavageofICP8aand ICP8bbythe
Staphylo-coccus aureus V8 protease yielded identical
-41 patterns of
peptides
(Fig.
4,
lanes to8).
Diges-tion with elastase yielded a variety of sizes of
peptideproducts,andagain the profiles with the
0
10 20 40 80 120 [image:2.491.52.232.70.528.2]Minutes
ofchase
FIG. 1. Pulse-chase labeling of the two forms of
ICP8. Cultures of Vero cells infected with HSV-1
strain mP (multiplicity ofinfection, 20; reference 5)
werepulse-labeled for 5minat4 hpostinfectionwith
12 ,uCi of[35S]methionine per ml inmethionine-free
minimal essentialmediumsupplementedwith 1%
dia-lyzedcalfserum.Unlabeled methioninewasaddedto
1mM,andcultureswereharvestedatthetimes shown
during the chase period. At the times indicated, the
flaskswereimmersed inanicebath,and the
monolay-ers were washedwithphosphate-buffered saline and dissolved in gel sample buffer (8) containing 0.5%
mercaptoethanol and 0.5 mM phenylmethylsulfonyl
fluoride.Theextractsweresubjectedto electrophore-sis ina9.25%polyacrylamide gel. Anautoradiogram ofthe driedgelisshown.
is 5 30 60
Minutesofchase
FIG. 2. Conversion of ICP8a to ICP8b during a
chase period. Cultures of infectedcells were labeled
andsubjectedtochaseconditionsasdescribed in the
legendto Fig. 1. Thecultures wereharvested atthe
timesshown, andextractswerepreparedand
subject-edtoelectrophoresis. The microdensitometer
(Joyce-Loebl) tracing of the ICP8 speciesfrom theresulting
autoradiogramisshown.
Sb Ob
Sb as
as
VOL.44, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.491.250.446.467.601.2]a b c d e erties of the two forms oftheprotein, we
pre-pared extracts frominfectedcells and
subjected
the proteins to
chromatography
on native and denatured DNA-cellulose underidentical condi-tions. Both species, ICP8a and ICP8b, weredetected in the high salt eluates from the two
jcP columns,
identifying
themasDNA-bindingpro-teins. The proteins eluted fromdouble-stranded DNA-cellulose at 0.3 M NaCl (34% of total
ICP8) and 0.5 M NaCl (22%), but they eluted
from single-stranded DNA-cellulose at 0.5 M
NaCl (59%) and 1.0 M (23%) NaCl concentra-tions
(Fig.
5).
Theproportion
oftheICP8protein
boundto the columnswas
higher
for denatured:
::
[image:3.491.55.250.73.399.2]:*
. .!.-8a DNA(89%
binding)
than fornative DNA(58%
...
::.:3-8b
binding). These results demonstrate that the HSV-1 ICP8 protein binds more tightly and more efficiently to denaturedDNA, as previous-ly demonstrated for the HSV-2 protein (16).A number of posttranslational modifications
ofproteins,
including
phosphorylation,
glycosy-lation, and cleavage, lead to a change in the electrophoretic mobilities ofprotein molecules. In this paper, we have shown that incomplete reduction of the HSV major DNA-binding
pro-tein leads to two electrophoretic forms of the
protein. The faster moving species, ICP8b, is
converted to the slowermoving species,ICP8a, by increasing the concentrations of
reducing
agents in the gel sample buffer.ICP8btherefore apparently contains disulfidebonds that alter the conformation and the mobility of the protein. Because the incompletely reduced species mi-FIG. 3. Conversion of ICP8b to
ICP8a
by reduc- grates faster than the more fully reduced form, tion. A nuclear preparation was prepared from infect- we believe that the incompletely reduced form ed cells by Dounce homogenization followed by wash- contains intramolecular disulfide bonds.Pre-ing
of the nuclei with 1% Nonidet P-40-0.5% sumably, a multimeric formwouldmigrate slow-deoxycholate. The nuclei were dissolved in gel sample er than the monomeric form in the gels. The buffer containing 0.5% mercaptoethanol. The extract primary amino acid sequences of the two poly-was divided into 5 aliquots, andDTT was added to the peptide species appear to be very similarbe-following
concentrations before boiling of the samples cause (i)ICP8b can be converted toICP8a byand electrophoresis: a, no
additions;
b, 10mM;
c, 20 reduction and(i)
thepeptide
profiles
generatedmM;
d, 40mM;
e, 100 mM. An autoradiogram of the by digestionwith
any of fourproteolytic
en-dried gel is
shown.bydgsinwtanoffupreltce-zymes are identical for the two species. Folding and disulfide bond formation could lead to an ICP8a and ICP8b species were identical (Fig. 4, altered conformation that allows the protein to
lanes 9 to 12). The profiles generated by diges- move faster through the gelmatrix oralters the tion of the two species with chymotrypsin and binding of sodium dodecylsulfatetotheprotein. papain were also identical (not shown). The two Several types of evidence suggest that both species therefore appeared to be very similar in forms may exist in the infected cell. (i) We
their primary amino acid sequence. These re- observed a progressive conversionoftheICP8a sults support the idea that the
ICP8a
form repre- form to the ICP8b form, theextent of whichwassents the unfolded form of the protein and that dependent on the time of chase. Thus, within by oxidation it is converted to the disulfide- one polyacrylamide gel, the state oftheprotein bonded, folded form
(ICP8b).
was different whenextracted from cellsat differ-We have previously reported that both forms ent times. (ii) Our preparation of extracts in can be detected in both the cytoplasm and sample buffer containing 0.5% mercaptoethanol nucleus of infected cells (10). Thus, the conver- probably precluded the spontaneous formation sion was not specific to one compartment of the of disulfide bonds (7). (iii)We detectedasimilar infected cell. To further test the functional prop- doublet in the ICP8 region of gels in which the738 NOTES J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL.44, 1982
IX 1: :.:
i-~~~~~~~~~~k*-.:.~ ~ *
,w~~~~~~~~~~~~~~~~~~~
::y*'w e a A
9 10 11 12
wop
1 2 3 4 5 6 7 8
FIG. 4. Partial proteolysis of ICP8a and ICP8bspecies.ICP8a and ICP8b in labeled nuclearextractswere
resolved on9.25% polyacylamide gels. The gels were dried andsubjectedtoautoradiographywithout fixation
because fixation ofgelsled to precipitation ofICP8in a form that could not be electroeluted (M. Quinlan, unpublished observation).Theautoradiogramwasused as atemplateto cut outtheaand b bandsfromthedried
gel.Theremainder oftheprocedurewassimilartothatof Clevelandetal.(3),except that the driedgelbands
weresoaked for 30 min in a solutioncontaining 12.5 mMTris-hydrochloride (pH 6.8),0.1% sodiumdodecyl
sulfate, 1 mMEDTA, and 30 mMDfT,asdescribed by Smith andHightower (17). DUT wasincluded inan
attempt tocompletely reduce the twoproteins and make themequally susceptible toproteases. Shown isa
fluorogramof a15%gel inwhich these two proteins were incubated with various concentrations of protease.
Odd-numberedlanes containedICP8a.Even-numbered lanes containedICP8b.Theamountsof proteaseadded
toeach well were asfollows: 1 and 2, 0.5gLgof S. aureus V8 protease; 3 and 4, 0.1gigof V8protease; 5 and 6,
0.025pgof V8 protease; 7 and8,0.005 ,ugofV8protease; 9 and10,0.1gLgofelastase;11and12,0.025gLgof elas-tase.
residual ammonium persulfate had been
re-moved by preelectrophoresis or in which ex-tracts reacted with N-ethyl maleimide were run
(not shown). Thus, at least a portion of the oxidized form was not due to oxidation of the protein within the gel.
The conversion from the ICP8a form to the
ICP8b formappears not tobedependent on the
cellular location of the protein. Furthermore,
ourexperiments suggest that both forms of the protein can bind tosingle-strandedDNA. At this
time, we cannot correlate this conformational
changewithanystageof maturation or function oftheprotein. We envision the conversion as a progressive oxidation of -the ICP8 molecule, leading to increased disulfide bonding duringits lifetime within the infected cell. The disulfide
bondingmay be facilitatedbythetertiary
struc-NOTES 739
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.491.105.395.76.444.2]740 NOTES
a.a
a b c d e f g
hi
j k I m nFIG. 5. DNA-cellulose chromatography of infected cell extracts. Labeled infected cell extracts were
prepared byamodification of the methods of Alberts and Herrick(1)and Powell andPurifoy(16). Flasks (25
cm')of infected Vero cellswerelabeledfrom4 to5 hpostinfectionwithIY5S]methionineand chasedfor 1 h with
cold methionine. The monolayers were washedwith phosphate-buffered saline, scrapedfrom theflasks, and
frozenat-800C.Thecellswerethawed in buffercontaining20 mMTris-hydrochloride (pH 7.5),10 MMMgCl2,2
mMCaC12, 1lmM EDTA,0.5 MNaCl,1 mM2-mercaptoethanol,and 500gigof bovineserumalbuminperml and
weresonicated for 15s.Thelysateswereincubated with 50jigof DNase Ipermlfort1hat200C. Theextractwas
clarifiedbycentrifugationat27,000 xgfor 15 min EDTAwasaddedto20MM, and theextractsweredialyzed
extensively against D buffer (150 mM NaCl, 20 mM Tris-hydrochloride [pH 7.5], 1 mM EDTA, 1 mM
mercaptoethanol, 10%glycerol).Theextractwasagainclarifiedby centrifugationat27,000xgfor15min. The
supernatantwasloadedonto0.75-mi DNA-cellulosecolumns,washed withDbuffer, andeluted with Dbuffer
containingvarious concentrations of NaCl. Bovineserumalbuminwasaddedto5gig/mlin alleluates,and each
eluatewasdialyzed againstasolutionof 0.15MNaCI.Proteinswererecovered from all fractionsbyprecipitation
with 9 volumes ofacetone.Native and denatured calfthymusDNA-celluloseswereprepared bytheprocedureof
Alberts and Herrick(1)andkindly supplied byWilliamRuyechan,Uniformed ServicesUniversityof the Health
Sciences, Bethesda, Md. The native DNA-cellulose contained approximately 1 mg of nucleic acid per g of
powder,whereas the denatured DNA-cellulose containedapproximately0.75mgof nucleic acidper gofpowder.
Shown isanautoradiogramofagelinwhich variousproteinfractionsweresubjectedtoelectrophoresis.a,Total
cellularlysate; b, extract putonDNA columns;c, pelletfromhigh salt DNaseextraction; d,pelletfollowing
dialysis. Samplesetoiarefrom denatured DNA-cellulose:e,flowthrough; f,0.3MNaCleluate;g,0.5 MNaCl
eluate; h,1.0M NaCleluate; i,4.0 MNaCleluate.Samplesjton arefrom native DNA-cellulose:j,flowthrough;
k,0.3 MNaCleluate; 1,0.5 MNaCleluate;m,1.0MNaCleluate;n,4.0 M NaCl eluate.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.491.103.401.97.453.2]VOL. 44, 1982
ture of the protein. Further work is needed to determine whether thefoldedstructure suggest-edby theseexperimentsisindeed present in the protein in the infected celi and whether it is required for any of the interactions of this pro-tein.
Wethank JohnMekalanos for hiscomments on the manu-script, William Ruyechan forDNA-cellulose and discussion of this work, and RosemaryBaccoforthe preparation of the manuscript.We alsothankAbbott Laboratories for the gift of disodium phosphonoacetate.
This workwassupported by Public HealthService grant CA 26345 from the National Institutes of Health. D.M.K. is a Cancer ResearchScholarof theMassachusettsDivision of the American CancerSociety. M.P.Q. is apredoctoral trainee supported by Public Health Service training grant CA 09031 from the National Institutes of Health.
LITERATURECfTED
1. Alberta,B., and G. Herrk. 1971.DNA-cellulose chroma-tography. MethodsEnzymol. 21D:198-217.
2. Bayllss,G. J., H.-S.Marsden,and J.Hay.1975.Herpes
simplex virus proteins: DNA-binding proteins in infected celUsand in the virus structure. Virology 68:124-134.
3. Cleveand,D.W., S. G.Fscher,M. W.Kbcner,a U.
K.Laenmni.1977. Peptide mapping bylimitedproteolysis
in sodium dodecylsulfate and analysis by gel electropho-resis. J. Biol. Chem.252:1102-1106.
4. Cooky,A. J., D. M.Kalpe,P. C.Jones, and B.Roinnan. 1981. Molecular geneticsof herpes simplexvirus. VII. Characterization ofatemperature-sensitive mutant pro-duced by in vitro mutagenesis and defective in DNA synthesis and accumulation ofgamma polypeptides. J. Virol.37:191-206.
5. Ejervito, P. M., E. D. Keff, and B. Rolzman. 1968.
Characterization of herpes simplexvirusstrainsdiffering intheir effectsonsocialbehavior of infected cells. J. Gen. Virol.2:357-364.
6. Fenwck,M.L., M. J.Walker,and J. M.Petkevkh. 1978.
On the associationof virus proteins with the nuclei ofcells infected with herpessimplex virus. J. Gen. Virol. 39:519-529.
7. Henning,R.,R.J.Miner,K.Reske, B. A.Cu am,
andG. M. Edehnan. 1976.Subunit structure, cell surface orientation and partial amino acid sequences of murine histocompatibilities. Proc. Natl. Acad. Sci. U.S.A. 73:118-122.
8. Honess, R. W., and B. Romnan. 1973. Proteins specified by herpes simplex virus. XI. Identification and relative molar ratesof synthesis of structural and non-structural herpes viruspolypeptides in the infected cell. J. Virol. 12:1347-1365.
9. Hons, R.- W., and B. Rolnan. 1974. Regulation of herpesvirusmacromolecular synthesis. I. Cascade regula-tion of the- synthesis of three groups of viral proteins. J. Virol. 14:8-19.
10. Knipe, D., and A.Spang. 1982. Definition of a series of stages in theassociationof twoherpesviralproteinswith the cellnucleus.J. Virol. 43:314-324.
11. Marsden, H. S., N. D. Stow, V. G. Preston, M. C.
Tlnbury,and N. M.Wkike. 1978. Physical mapping of
herpes simplex virus-induced polypeptides. J. Virol. 28:624-642.
12. Morse, L. S., L.Pereira, B.Rolznian,andP. A.Schaffer. 1978.Anatomyof HSVDNA. X.Mapping of viral genes by analysis ofpolypeptides and functions specified by HSV-1 xHSV-2 recombinants.J. Virol. 26:389-410.
13. Perder,L., M.H.Wolff,M.Fenwlck,and B.Rolznan.
1977.Regulation of herpesvirus macromolecular synthe-sis.V.Propertiesof alphapolypeptides made in HSV-1 and HSV-2infected cells.Virology77:733-749. 14. PoweDl, K. L., E.Lkter,and D. J. M.Purlfoy.1981.
Non-structural proteins ofherpes simplex virus. II. Major
virus-specified DNA-binding protein. J. Virol. 39:894-902.
15. Powell,K. L., and D.J.M.Purlfoy. 1976.DNA-binding proteins ofcellsinfectedbyherpes simplexvirus type 1 and type 2.Intervirology7:225-239.
16. Prfoy,D.J.M., and K. L.Powell. DNA-binding
pro-teins inducedby herpessimplex virustype 2 in HEp-2 cells. J. Virol. 19-.717-731.
17. SmIth,G. W., and L. E. Hightower.1981. Identification of the Pproteins and otherdisulfide-linkedand phosphory-lated proteins of Newcastle disease virus. J. Virol. 37:256-267.
18. WUlox,K.W., A.Kohn, E.Sklyanskaya,and B.Rolman.
1980. Herpes simplexvirus phosphoproteins. I. Phos-phate cycles on andoffsomeviral polypeptidesand can altertheiraffinity forDNA. J.Virol. 33:167-182.
NOTES 741
on November 10, 2019 by guest
http://jvi.asm.org/