II.
Analysis
of
Intracellular
Development and the Structure of Very
Fast-Sedimenting
DNA
BORRIES KEMPER* AND DENNIS T. BROWN
InstitutfurGenetik der UniversitatzuKoin, 5000 Koin 41, Federal Republic of Germany Received for publication26November 1975
Withtheexception of mutantsingene 49,all mutantsinphageT4defectivein
theprocessof headfillingaccumulate a normal replicative DNA intermediate of
200S. Mutants in gene 49 producea veryfast-sedimenting (VFS)DNA withs
values ofgreaterthan 1,OOOS. The intracellular development of the VFS-DNA
generatedingene 49-defective phage-infected cells was followed by sedimenta-tionanalysis of crude lysatesonneutral sucrosegradients. It wasobserved that
theproductionof a 200S replicativeintermediate is onestep in thedevelopment
of VFS-DNA.Afterrestoring permissiveconditions thedevelopment of the
VFS-DNAcanbereversed, but the 200S form is notregeneratedunder these condi-tions. The process of head filling can take place from the VFS-DNA under
permissive conditions. Fromthe absence of other components in the VFS-DNA
complexes, its high resistance toshearing, its resistance against the attack of
the single-strand-specific nuclease S1, and from itsappearance in the electron microscope, acomplexstructureoftightlypackedDNA isinferred.The demon-stration by the electron microscope of branched DNA structures sometimes
closely relatedto partially filled headsis taken in support of the idea that the process of head fillingin gene 49-defective phage-infected cells is blocked by
somesterichindrance inthe DNA. Inlightof theseresults, the role of gene 49 is
discussedas a control function for the clearance ofthese structures. Afixation
procedure forcross-linking ofgene49-defective headstotheVFS-DNA allowed
ustostudy progressivestages intheprocessof headfilling.Electronmicroscopic evidence is presented which suggests that during the initial events the DNA
accumulatesinthevertexes of the head.
Mutantsingene 49of phage T4aredeficient
intheprocessof headfilling. As aconsequence,
partially filled heads (14, 19, 25) and
unma-tured very fast-sedimenting (VFS) DNA accu-mulate inthe infectedcell (9). As shown in the
precedingpaper (13)the VFS-DNAseems tobe
boundtothe membrane duringthe lifecycleof
the mutant but can be isolated free of
mem-brane or other material without changing a characteristic sedimentation rate of greater than 1,OOQS. These s values differ markedly from thereported 200S for the regular replica-tiveintermediate, which was observed in wild-type or maturation-defective phage-infected cells with afunctioning gene 49 (7, 8, 10). In the
following, experiments are reported that were done to investigate the relationship between the 200S and the VFS-DNA in vivo and to further analyze the structure of the VFS-DNA
complexes by biochemical, physical, and elec-tronmicroscopic methods.
MATERIALS AND METHODS
Cells and bacteriophage. In addition to the strainsof Escherichia coliandphage T4 which were described earlier (13), the phage mutant strains
listed in Table 1 were used. All strains were ob-tained from W. B. Wood. Double mutants were con-structed by standard procedures in our laboratory.
Chemicals and enzymes. Ethidiumbromide was obtained from Calbiochem. Formaldehyde was pur-chased as a crystalline solid (paraformaldehyde) fromPolyscience. The powder was dissolved in
wa-ter to afinal concentration of 2% (wt/vol) and neu-tralized by the addition of NaOH at 60 C.
Polyethyl-eneglycol6000wasfrom Serva. [3H]thymidine (spe-cific activity, 25 Ci/mmol) and [L-'4C]leucine (spe-cific activity, 250 mCi/mmol) werepurchased from Amersham Buchler. Single-strand-specific nuclease S1 (30) was a giftof F. Schmidt from our depart-ment.
Assay conditions for nuclease S1. A total assay volume of 300 ,ul contained 22.5 ,umol of CH3COOH-CH3COONa, pH 5.0, 75 Amol ofNaCl, 25 nmol of ZnSO4, and 10 nmol ofheat-denatured DNA and
1000
on November 10, 2019 by guest
http://jvi.asm.org/
FUNCTION OF GENE 49 OF PHAGE T4. II. 1001
enzyme. Underthese conditions1
Al
of theenzymepreparationdegraded75 ugofheat-denaturedDNA at37Cwithin30min. T4DNAwas aswelldegraded
asDNA from othersources.
Isolation ofgene49-defective phage heads. The
isolation of ['4C]leucine-labeled gene 49-defective
heads from Brij 58 lysates (13)followed the
proce-dure of Luftigetal.(25).Accordingtothese authors, the heads havean s value of350S. Different
num-berswerereported by other authors (19).Thegene
49-defective heads isolated from theBrij 58lysates
have an s value of 4105 in our experiments, as
concluded from their cosedimentation with intact
phagelambda(410S). Thevariationsof thesvalue
reflect a variability of the head constituents, as
demonstrated by Laemmlietal.(19),whowereable to show that the DNAcontent of theheads is de-pendentonthe lysis procedure.
Lysis procedures. The lysis ofphage-infected cells
was achieved as previously described (13). In the case of the polyethylene glycol-stabilized lysates,
theprocedure of Laemmlietal. (19)wasadaptedto ourstandard lysisprocedure. Infected cells(4x 109) weresedimentedat4 C bycentrifugation and
resus-pendedin1mlof 10-2Mphosphate bufferatpH7.1.
Anequal volume of lysis buffer containing 10-2M
phosphate buffer, pH7.1,20%polyethylene glycol,2 X 10-3 M KCN,0.6MNaCl,2x 10-3 M EDTA, and 2 mgof lysozyme perml wasadded. After 15 minat 0Ccell lysiswasinduced by the addition of Brij58 aspreviously described (13).
Cross-linking procedure and isolation of the
cross-linked product. A combination ofa sucrose
gradientcontainingformaldehydeasacross-linking agentandaCsCl block gradient fortheseparationof thecross-linkedproductswasused in thesame
cen-trifugation tube. A density block gradient was
formed from1.5mleach of solutionscontainingCsCl at1.8, 1.5,and1.3g/ml.On top of thisalinear 5-ml
sucrosegradient containing35to15%sucroseand4 to0%formaldehyde in 0.1 Mphosphate buffer, pH 7.1,0.3 MNaCl,and10-3MEDTA(11)wasformed
byaBuchlermixingdevice. A 1.5-ml amount of 15% sucrose in the described buffer without
formalde-hydewaslayeredontop;thiswasfollowedby1mlof
the polyethyleneglycol-high salt-stabilized lysate. Centrifugationsweredone in theSW41 Ti rotorfor 90 minat4C and 17,000rpminaBeckman
ultra-centrifuge.
Electron microscopy. Insitulysisofthe infected E. coli cells was carried out by the procedure of
Laemmliet al. (21). The purified DNA complexes
and DNA complexes with attached viral capsids
werevisualizedin theelectronmicroscopebya
mod-ification of the procedure of Kleinschmidtand Zahn
(15).Thegradient-purified structureswere first
di-alyzed against0.5 M ammonium acetate, 0.001 M EDTA (pH 8.0). The suspension was then made 5
,ug/ml in cytochrome c and spread on a distilled water hypophaseatroom temperature. The
mono-layer of proteinand DNAwaspickedupon carbon-coated grids that had been made hydrophylic by glow discharge. The mountedcomplexeswere
posi-tively stained by dipping in a 2% aqueous uranyl
acetatesolution followedbyrepeated washingwith
TABLE 1. Bacteriophagestrainsa
Mutant Gene Phenotype
amN66 16 2008DNA, emptyheads
amNG63O 17 200S DNA, emptyheads amB17 23 2005 DNA, no heads
tsC9 49 VFS-DNA,partially filled heads
a Thestrains listed arespecified by their original
selec-tionnumber (first column) and the number of the mutant gene (second column). The phenotype in respect to the production of DNA intermediates and head structures is given in the last column.
distilled water. The stained specimens were exam-ined in the electron microscope utilizing dark-field optics. All specimens were photographed in a Sie-mens 101electronmicroscope.
Other techniques. The following techniques were described earlier (13): preparation and lysis of phage-infected cells, isolation of VFS-DNA or 200S DNAfromphage-infected cells, labeling of intracel-lular phage DNA, neutral and alkaline sucrose gra-dient analyses, and determination of trichloroacetic acid-insolubleradioactivity.
RESULTS
DevelopmentofVFS-DNAinvivo. To follow
the intracellular generationofVFS-DNA,
non-permissive cells were infected with T4 phage
carrying an amber mutation in gene 49
(amE727). The newly synthesized DNA was
pulse-labeled from 8 to 12 min after infection
with tritiated thymidine. At different times
afterinfection,aliquotswereremoved from the culture and gently lysed by the lysozyme-Brij
58 technique (13). The sedimentation behavior of the DNA was analyzed on neutral sucrose
gradients. Purified phageT4and empty heads
isolated from a gene 49 mutant were run in
parallel tubes, marking the positions of 1,OOOS and410S, respectively.
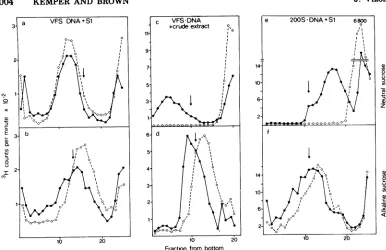
The results in Fig. lademonstrate a drastic
change in the sedimentation rate of early la-beled DNA. At 8 minafter infection the main
portion of the label is found in a position
be-tweenthe twomarkers. With increasing time
thesvalues (calculated for themainfractionof the peak) change from 1,OOOS at 10 min to
approximately 1,500S and 1,800S at 14 and 19 min, respectively. From 19 to 29 minthe shift towardshighers values isreduced and broad-eningof thepeak becomes apparent, which in-dicates an increasing heterogeneity of the DNA.
Forcomparison with the mutantingene 49,a mutant in gene 23 (amB17) has been tested. This mutant is defective in the production of
headsbutwild type forgene 49function, andit was shown that it causes the accumulation of the regular replicative 200S DNA in the
in-fected nonpermissive host (8, 10). The results
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.503.253.445.74.130.2]0
a)
C:
E
¢Y)
0-T
CO)
10005
gene49 gene49tS gene 23 gene49tS-gene23
FIG. 1. Sedimentation analysis of intracellular mutantphageDNA. E. coli B23 wasinfected withfive phage per celland incubatedat37 or 42Cinthecaseof temperature-sensitivemutantphage.ThephageDNA
waspulse-labeled from 8 to 12 min after infection with [:3H]thymidine. Cells infected with
temperature-sensitive mutants wereincubated at42Cfor30minbeforetheywereshiftedto apermissivetemperatureof
28C.Atthetimesindicatedinthefigure,aliquotswerewithdrawn andlysed bythe describedtechnique (13). The sedimentation behaviorof thephageDNAwasanalyzedon neutral10 to30%sucrosegradientsinan
SW50.1 rotor at17,000 rpm and 4Cfor30 min. TheDNA content ofeach fraction was determined by trichloroacetic acid precipitation. Phage labeled in their DNA content with ['4C]thymidine or gene 49-defective heads labeled with['4C]leucinewereaddedtothesamples before centrifugationtomark thepositions ofl,OOOS and 410S,respectively. Thefollowing phagemutants wereusedfor infection:(a)amE727(gene49); (b) tsC9 (gene 49); (c) amBl7(gene23);(d) C9ts-amB17 (gene49-gene 23).
showninFig. lcindicate that theearly labeled
gene23-defective DNA sediments between the twomarkersthroughout the timeunder inves-tigation. At 29 min after infection, a shoulder
toward the 1,OOOS position is detectable. This seems to be due to the production of a small number of complete phage, as suggested by a fourfold increase in the titer of viable phage between 19 to 20 min. We have no final expla-nationfor the difference between the reported s value of 200S for the gene 23-defective DNA and the much higher s value(>410S)obtained in our experiments. The same result was ob-tained whenboth DNAs, prior to sedimentation
analyses, were purified from the lysates by CsCl blockgradientcentrifugation. Samples of identical density from position I of the gradient
(for detailsseeprecedingpaper [13])were
cho-sen for comparative study, thus reducing the
possibility of differencesin the amountof con-taminatingsubstances. Although other
expla-nations arenotexcluded,it isconceivablethat the more gentle lysozyme-Brij 58 lysis proce-dure better preserves the in vivo structure of the complexes, leading to higher s values. In
ordernot toconfuse thediscussion onthis sub-ject, wecontinue toreferto DNAsthat, under
ourconditions, sedimentinthe position of gene 23 DNA as "200S DNA" until this problem is investigatedinmoredetail. From the compari-sonof the development of the DNA complexes in cells infected by gene 23- and 49-defective phage,weconclude that the development of the VFS-DNA of gene 49-defective phage at early
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.68.460.54.371.2]FUNCTION OF GENE 49 OF PHAGE T4. H. 1003
times passes a stage in which the DNA shows
the same sedimentation rate as the gene
23-defective 200S DNA. Whether these DNAs of the two mutant strains are identical in other respects was notinvestigated.
Whathappens to the VFS-DNA complexes if
permissive conditionsarerestored? A
tempera-ture-sensitive mutation in gene 49 (tsC9) was
used for this investigation. Although the
com-plex formationof tsC9 DNA is not as fast as in
amB727-infected cells (probably due to leaki-ness of the ts mutation), the isolated DNA
shows all characteristics of the VFS-DNA
re-ported earlier (13). The results of temperature
shift experiments are demonstratedin Fig. lb
and show that upon restoration of permissive
conditions the VFS-DNA is almost
quantita-tively shifted from 1,400S to a lower value of
1,00Sat theposition of intact phage.
Approxi-mately50%ofthe label in this position proved toberesistant to DNase I, which is taken as an
indicationofsuccessfulpackaging (23). The rest
of the DNA remains DNase sensitive but
co-sediments with intact phage. The number of
viablephage is 30- to 50-fold increased. No 200S material isproduced, if compared with a control
sample keptat 42 C afterthe shift. The same
results are obtained when chloramphenicol is given 2 min before the shift, indicating that concomitantproteinsynthesis is not a require-mentforsuccessfulheadfilling or DNA cutting
after restoration ofpermissive conditions (25;
data notshown).
Itwasreported that theVFS-DNA could be convertedinto a200Sformunder the influence ofactive P49-containing crude cell extracts in vitro (9). Ifthis conversion is afast and
tran-sient step in vivo, we might have missed the
200S product inour experiment. Wetherefore tried to block the ability of the temperature-sensitive mutation to propagate phage at low
temperature by combining the tsC9 mutation
with an amber mutation in the gene for the
main head protein gene 23, amB17. At high
temperature we expect the gene 49-defective phenotype and the production of VFS-DNA. Afterashiftto low temperature, P49 is reacti-vated andcleavage of the VFS-DNA could
oc-cur,but theblockinheadassemblyshould then lead to an accumulation of intermediates
be-tween VFS-DNA and packagedDNA. The
re-sultsarepresentinFig. ld.Besides somevery
limited cleavage of the VFS-DNA, no
signifi-cantproduction of200S or smaller-sized DNA canbeobserved.
Fromtheseresultsweconcludethatthe gene
49-defective VFS-DNA can directly serve as a
substrate for packaging and the formation of 200S DNA is not required. The results also
indicatethat in the absence of active head fill-ingthe reactivation of P49 is not sufficient to cut VFS-DNA into 200S or unit length DNA molecules in vivo.
Resistance of the VFS-DNA against enzy-matic cleavage and hydrodynamic shear.
Ex-perimentsby Huberman (12) strongly suggest
that each cell infected with a 200S
DNA-pro-ducing phage mutant gives rise to only one
complex containingthe DNA of awhole burst.
Our observation that the rate and yield of DNAsynthesis is essentially the same in
200S-or VFS-DNA-producing strains (data not
shown) favors the idea that the increase in
sedimentation value of the VFS-DNAiscaused
by an increase in compactness rather than by
variationsin theamountof DNAper complex. As was shown in a preceding report (13), no
substances other than DNA werefound inthe
VFS-DNA complexes and a direct DNA-DNA
interaction was suggested as a reason for
for-mation of the VFS-DNA. This could occur by
incomplete recombinational eventslinking
dif-ferent parts of the DNA molecules (1, 2) and tightening the complex structure.
Recombinant structures frequently contain
single-stranded regions (1, 2), which upon
cleavage should result in a separation of the DNA genomes involved. If such regions were present in the VFS-DNA it should be possible
to cleave them with single-strand-specific
nu-clease
Si
invitro(30),resultingin anunfoldingof thecomplexes. As showninFig.2ahowever,
a 15-fold excess of thatamount of theenzyme necessary tocompletelyconvertthe heat-dena-turedgene49-defective DNA into
trichloroace-tic acid-soluble material did not change the sedimentation behavior of the native gene
49-defective DNA, asjudged fromneutralsucrose
gradient analysis. The analysisonalkaline su-crose gradients demonstrates the introduction ofapproximatelyonebreakontheaverage per
single-stranded molecule (Fig. 2b).
Thesedimentation velocity of20(0S DNAwas
drasticallyreduced if treated with nuclease
Si,
thus corroborating the results of Frankel (7). The number of
Si-introduced,
single-strandbreaks, however, was not significantly
differ-entfrom the numberof breaks introducedinto
VFS-DNA(Fig.2eandf).This resultis
surpris-ing because the introduction ofapproximately
the same number ofsingle-strandbreaks hasa
very different effect on the two DNA
com-plexes.
In contrast, the VFS-DNA can be rapidly
degraded after the exposure to crude extracts containing P49(Fig. 2c; 9) withoutintroducing
more than one break per single-strand mole-cule (Fig. 2d). Control experiments show that VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
CM
x
E
Q)
0L
en
0
-CO
(n 2
-5
z
a) U) 0
U)
-c -I
Fractonfrom bottom
FIG. 2. TreatmentofVFS-DNAor200S DNA with nucleaseSlorP49containingcrude extracts. A 6-nmol
amount of[3H]thymidine-labeled, native gene49-defective VFS-DNA, orgene23-defective 200S DNA was
treated withanamountof single-strand-specificnucleaseSlsufficientto convert 90 nmol of heat-denatured T4-DNA intotrichloroacetic acid-soluble material under the assay conditions described in Materials and Methods. Afterthetreatment the DNAswere analyzedon neutral (a and e) oralkaline (b and
t)
sucrosegradients bytrichloroacetic acidprecipitation. Anothersample ofnative gene49-defective VFS-DNA was
treated withacrudeextractofT4wild-type-infectedcellsasdescribedbyFrankel et al.(9), and theproducts
wereanalyzedonneutral and alkalinesucrosegradientsasdescribed above (c andd).The arrow points to the positionofl,000Sin(a),(c),and(e)andtotheposition of70Sfull-lengthsingle-stranded T4 DNA in (b), (d), and(f). Symbols: (O) distributionofthe DNA beforethetreatments; (0)distributionofthe DNA after the
treatments.
crude extracts from gene 49-defective phage-infected cells donotshow this effect. This signi-fiesadifference between theactionofSianda gene 49-controlled factor(s) on the VFS-DNA. Weconclude that the VFS-DNA contains
sin-gle-strandedregionscleaved by theenzymeSi.
However,thesesites seemofnoobvious impor-tance for thecomplexstructure.
If the DNA molecules are -more tightly
packedinthe VFS-DNAcomplexes thanusual,
one could expect an increased resistance
against shear forces in comparison to less
tightly packed DNA like the 200S material. Thisinfact is true, as isshown by our analysis ofthesedimentation behaviorofVFS-DNA and
200SDNAbefore and after theexposureto mild shear forces bypipetting.
Morethan 50% ofthe VFS-DNA still exists in aVFS form (Fig. 3aandb),whereas all of the 200S DNA is reduced in itssedimentationvalue
(Fig. 3cand d). On thecontrary, the s value of the VFS-DNAiseven enhanced after the
shear-ingprocedure,probablyindicatingachange in
the complex structure. We think that loosely
arranged DNA strands protruding froma com-pact center of the complex (see below) can
re-duce thesedimentation velocityina parachute-like manner. Preferential removal of these
strands by shear would then havean accelerat-ingeffectonthesedimentation of therestofthe VFS-DNA complex.
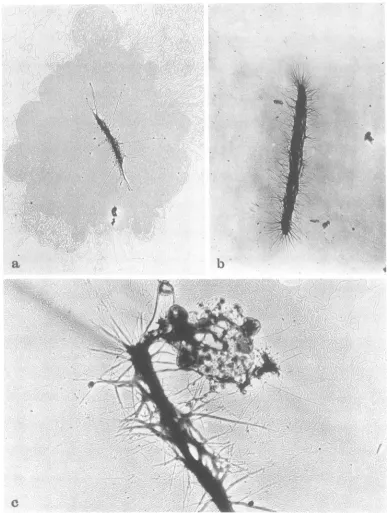
Visualization of gene 49-defective DNA.
Visualizationof released and membrane-bound
DNA inthe electron microscope was achieved
by taking DNA samples after CsCl gradient centrifugation(13)and dialyzingand spreading
themby the Kleinschmidt technique(15).
Typi-cal results obtained with DNA isolated at 30 minafter infectionareshowninFig. 4b and c. Bothpreparationsshow the DNA in large
com-plexes. The outer regions of these complexes
consist of loosely arranged individual DNA
molecules, whereasthecenters are highly com-pact,exhibitingatypical network structurein
which DNA molecules seem to be twisted around each other, forning strands of higher
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.67.453.58.308.2]FUNCTION OF GENE 49 OF PHAGE T4. II.
order. The membrane-bound DNA (M-DNA) (13) contains pieces ofmembrane and/or cell wall material predominantly attached to the center part (Fig. 4b). This portion of the M-DNA can be removed by treatment with or-ganicsolvents, which resultsintheconversion
of M-DNA into released DNA (13; electron mi-croscopicobservation not shown).
The network structure of the center, which develops quite early after infection (see next section), is typical for gene 49-defective DNA and is rarely seen in complexes from other mu-tantsdefective in DNA packaging (12). A typi-cal 200S DNA isolated at 15 min after infection from gene 23-defective phage(amB17) is shown
inFig. 4a. Its developmental stage is compara-bletothe gene 49-defective DNAshown in Fig.
5b. Thecenterof the gene23-defective DNA is
smaller and the DNA molecules are more
widely spread than in the gene 49-defective DNA.
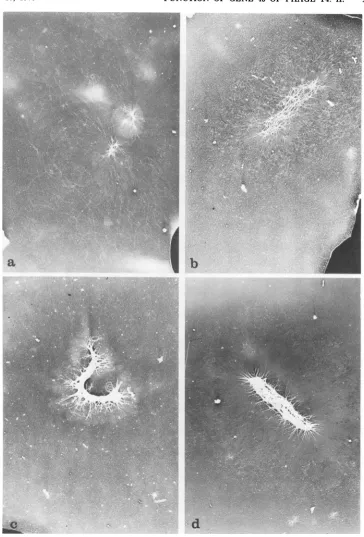
Development of gene 49-defective DNA as observed in the electron microscope. The S values of the VFS-DNA increase with time after infection (Fig. 1), andwewanted to know how this change is correlated with the structure of the complex. Therefore, samples were taken
at10, 15, 25,and35 minafter infection, and the isolated VFS-DNA wasanalyzed in the electron
microscope. The results are shown in Fig. 5.
One can see that with increasing time the
amountof DNA percomplexincreases while its center portion develops its typical network
structure.At30minafter infection the average sizeof thecenter(4by0.7,Lm)iscomparableto
the average size of the cellby which the
com-plexissurrounded. The fact that the DNA con-tentof the infected cellconserves theshapeof theenvelopedespite the lysis and isolation pro-cedure is taken as additional evidence for strong interactions between individual DNA
molecules within thecomplex.
In situ lysis of cells infected with gene
49-defective phage. It has been reported that,
after the removal of water by exposure to
ethanolic solutions, the DNA collapses into a
compactform ofhigherorder (16, 17). Some of these forns resemble theobservedstructuresof the gene 49-defective DNA(cf. plateIain
refer-ence 16). Since the DNA in our preparations
wasexposedto 90%ethanolafterspreading,the
visualized network structures of the centers
could be an artefact induced by the method. Therefore, we tried to reduce the danger of precipitation by eliminating the ethanol step andusingthe in situlysis techniquefor
visual-ization ofthe DNA (21). A
temperature-sensi-tive mutation (tsC9) was used, which
permit-ted us to follow thedevelopmentofthe
intracel-a) CONTROL 2100 5x SHEAR
I, I71 it
0
o5
3-10 20 10 20
[image:6.503.255.448.72.274.2]Fraction from bottom
FIG. 3. Shearing of VFS-DNA and 200S DNA. A sample of gene 49-defective VFS-DNA or gene
23-defective 200S DNA was sheared by pipetting the DNAfive times with an automatic Eppendorfpipette. The DNA was analyzed before and after the very mildshearingprocedure on neutral10 to 30%
su-crosegradients. Trichloroacetic acid-precipitable ra-dioactivity was determined. The arrow in (A) and (B) indicates the position of the marker phage (1,OOOS), whereas in (C) and (D) it points to the positionof marker heads(410S).
lular DNA after restoring permissive condi-tions. Cellswere infectedat 42 Cin theusual way,and the culture was splitinto twoportions
at30minafter infection. Onesamplewaskept
at 42 C, whereas thesecondsamplewas
trans-ferredto28 C.Atdifferenttimesafter the shift
aliquots wereremoved from both cultures and prepared for the lysis procedure by placing them into 0.01% OS04 (final concentration). Typical results are shown in Fig. 6. Pictures weretaken from samples isolated after30 min at 42C (Fig. 6a)orafter30 min at 42 Cplusan
additional30min at28 C (Fig. 6b).The prepa-ration of thesampleatnonpermissive tempera-ture shows empty heads, cell wall material,
andanetworkstructurespreadoverthe entire
areaof thelysedcell. Thisstructureresembles the center of the DNA complexes seen before (Fig. 5). No significant change in its
appear-ancehas beenobservedup to 60 minafter infec-tion at 42 C. The preparation of the cells that were shifted at 30 min to the permissive
tem-peratilre
and kept there for an additional 30 min has filled heads and does not show thecomplexDNA structures seen in thenonshifted cells.
The DNA structures observed in these cell
preparations are notrestrictedtocells infected
VOL. 18, 1976 1005
on November 10, 2019 by guest
http://jvi.asm.org/
;
* No
e lb~~~~
a
IL
_ V
b
..
C
FIG. 4. Visualization of 200S DNA and VFS-DNA in the electron microscope. The DNA from phage-infectedcellswasisolatedbythedescribedmethod(13),spread bythetechniqueof Kleinschmidtetal. (15),
and then visualized inthe electron microscope bybright-field illumination. (a)Membrane-free 200S DNA from T4amBI7(gene23)-infected cells isolatedat 15minafter infectionat37C.Magnification, x 8,500.(b) Membrane-free VFS-DNA from T4 amE727 (gene 49)-infected cells isolated at 35 min after infection. Magnification, x 8,500. (c) Membrane-bound VFS-DNAfromthesamepreparationas in (b).
Magnifica-tion, x20,000.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.65.452.75.588.2]FUNCTION OF GENE 49 OF PHAGE T4. II. 1007
b
C
d
FIG. 5. Developmentalstagesof theVFS-DNA asvisualized by the electron microscope.E. coli B23 was
infected withT4 amE727 (gene49),andtheDNAwasisolatedat(a)10,(b)15, (c)25, and(d)35 minafter infection. The membrane-free portion of each preparationwasvisualizedintheelectronmicroscopeby dark-fieldillumination. Magnification, x 7,000.
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.69.433.67.604.2]a
b
FIG. 6. Insitulysisofgene49-defective phage-infected cells. E. coli B23 was infected with T4 tsC9 (gene
49) andgrown at 42C for30min. An aliquot was removed and analyzed by the in situlysisprocedure (21) (a). The restofthe culture was then shifted to thepermissive temperature of28 C and kept there for another 30 min, before the cells were analyzed in the same way as above (b). Magnification, (a) x 30,000; and (b)
x 77,000. L,lump.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.503.78.438.74.610.2]FUNCTION OF GENE 49 OF PHAGE T4. II.
with a gene 49-defective mutant. To a lesser extentthey were also seen in gene 23-defective (amB17) phage-infected cells. Furthermore, the proceduredoes notallowareliable distinc-tion between the intracellular forms of VFS-DNAand200S DNA.
It is worth noting that almost all the prepara-tions of cells infected with gene 49-defective phage show a higher than usual number of lumps at bothtemperatures. Lumps consistof
uncleaved, randomly aggregated P23 and their production is caused by mutations in gene 31 (18). Recently, Vandersclice et al. (29) pre-sented evidence that the concentration of P31 is reduced in cells infected with gene 49-defective phage, whereas the amount ofuncleaved P23
was increased. The observed appearance of lumps in this study is consistent with their results.
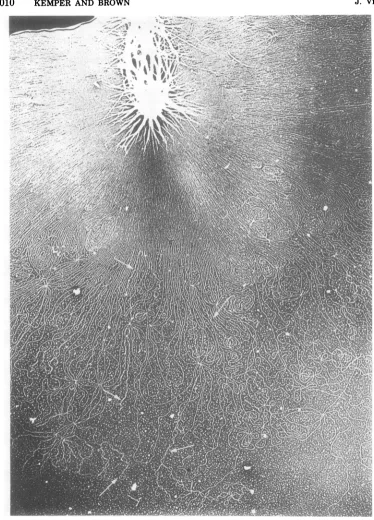
Supercoiled VFS-DNA. Evidence was
pre-sentedthat the chromosome of E. coli is orga-nized into a definite number of supercoiled loops (6, 27, 32), and itwouldbeinterestingto
knowif this is the general way to maintain the order ofoversized DNA molecules. Therefore,
weanalyzeddifferent developmental stages of the gene 49-defective DNA for supercoiled
mol-eculesinthe electron microscope. In only a few instances were we able to demonstrate super-coiledregions inthe DNA(Fig. 7). No correla-tionbetween the number of supercoils and the
developmental stage of the DNA could be
found.
This analysis is necessarily limited to the outer regions of the complexes, because the inner part is too tightly packed and details
cannotbe seen by the electron microscope. To
test whether the compact centers contain
su-pertwisted DNA, we exposed VFS-DNA to varying concentrations to ethidium bromide andanalyzedthesedimentation behavior of the
complexeson neutral sucrose gradients inthe presence of thedrug. Concentrations of ethid-iumbromide between 2and 160 ,g/ml had no
effectonthesedimentationbehavior. Although this result isnot astrongargument againstthe presence of supercoils inthe complex, itdoes, however, exclude the possibility that such
structures are ofimportance for the
arrange-mentof the DNAinthe VFScomplexes. Relationship between the VFS-DNA and head filling. We have shown that VFS-DNA
canbeusedas asubstrate forheadfillingwhen permissive conditionsarerestored. Other
work-ersdemonstratedtheaccumulationofpartially
filled headsingene 49-defectivelysates (14, 19, 25). The observation that these heads cosedi-ment with the gene 49-defective DNA was
taken as evidence for a close association
be-tweenthe partially filled heads and unmatured DNA (19, 23).
Our VFS-DNA preparations did not show anyheadsattachedwhen we usedthe described
standard isolation procedure (13). However, when the DNA content of the heads was stabi-lized by polyethylene glycol and high salt
ac-cordingtotheprocedure of Laemmli et al. (19), and when this treatment was followed by a
cross-linking reaction (11), we were able to
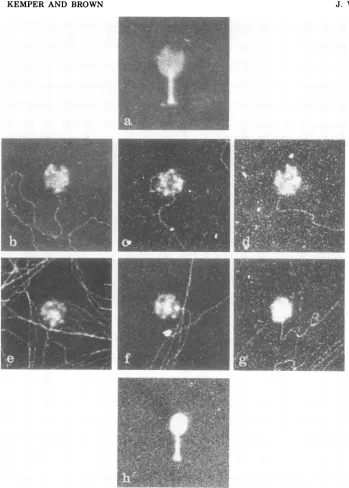
demonstrate directly the close contact of the gene 49-defective headsto the VFS-DNA com-plexes. In Fig.8 agene49-defective VFS-DNA complex is shown which containsanumber of headsattached to protruding DNA molecules. Examples for a terminal connection between theheads and the DNA are showninFig. 9.An
examination of T4 heads cross-linked to the
DNAcomplexin gene49-defective mutants
re-vealed heads in a variety of partially filled
states (Fig. 9b-g). Some heads were nearly empty, whereas others contained relatively large amounts of DNA. We believe that all of the complex-associated heads are attached to
the DNA by virture of the continuity of the DNA seenoutside the head and the DNA inside the viral head. Support for this notion is ob-tained from studies carried out in an identical fashion with mutations in genes 16 and 17.
Although these mutations arealso defectivein
DNA packaging and large numbers of empty
heads areproduced (19, 24), these heads could
not be cross-linked to and purified with the viral DNA usingourtechniques.Therefore, the accidental trapping ofphageheadsinthe gene 49-defective DNA complex is ruled out. Our resultsfindsome supportinthework of Wun-derli and Kellenberger (H. Wunderli, personal
communication), whoobtained, bythin section-ing of phage-infected cells, incompletely full
heads, which represent either particles into whichonly part ofafullDNAcomplementhas
beenpackedorfull particlesof which apart of
theDNAhadescapedby artefact.
Theuseofpositive stainingwith
uranyl
ace-tatefollowedby dark-fieldelectron
microscopy
hasallowedour
study
tobeextendedtoincludesomepreliminaryobservationsontheprocess of
packagingof the T4 DNAintovirus heads dur-ing morphogenesis. The DNA in these
posi-tively stained preparations binds
uranyl
ace-tatemuchbetter than
protein,
anditispossible
to determine the amount anddisposition
of DNA packaged into the virus heads(Fig.
9). Emptyheadscontrastverypoorly
by
the proce-dure, as is evidentby
examination ofghosted
intact virions (Fig. 9a), whereas
similarly
pre-pared and stained
complete
phages
have veryhighlycontrastedheadsindark-field
illumina-VOL. 18, 1976 1009
on November 10, 2019 by guest
http://jvi.asm.org/
1010 KEMPER AND BROWN
w
~~~~~~~~~~~~~~-1.'
..
-\-4,,
i
.C
-9A /
/
FIG. 7. Demonstration of supercoiled portions in the VFS-DNA. VFS-DNA isolated as membrane-free
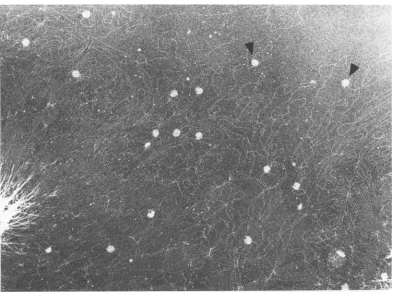
DNAfromT4amE727 (gene49)-infected cellswasspreadandvisualizedin theelectronmicroscopeunder dark-fieldconditions.The arrowspointtosupercoiled portionsoftheDNAcomplex.Magnification, x20,000.
tion (Fig. 9h).
It is of particular interest that those heads containingrelatively smallamounts of DNA in our experiments did not have this DNA
uni-formly distributed inside the heads. These
areas of DNA concentration wereperipherally
locatedasthough associated with somecapsid structural component, perhapsthe vertexes.
Theisolatedgene49-defectiveheadsthat
ex-hibit a DNA content of 10 to 50% cannot be
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.503.65.439.62.582.2]FUNCTION OF GENE 49 OF PHAGE T4. II. 1011
FIG. 8. Demonstrationofattached heads to the VFS-DNA. A sample of gene 49-defectivephage-infected cellswaslysedat 30 minafterinfection by the polyethyleneglycol-high saltproceduredescribedinMaterials andMethods. The heads werecross-linked to the DNA by formaldehyde treatment, and the complexes were spread and then visualized in the electron microscope. Arrows pointto twoexamples of heads terminally attachedtoDNA molecules. Magnification, x90,000.
filled to completion in the absence of gene 49
product (P49) in vivo. Fromthe observed
nu-cleolytic function of P49 (9; this paper) it was
suggested that some steric hindrance of the DNA substrate might be the reasonfor thelack ofcomplete head filling inthe mutant (5). We therefore began a search for some unusual
structures intheVFS-DNA, and itwaspossible
to demonstrate the appearance of branched
DNAmolecules sometimessituated close to the vertex of anattached head (Fig. 10). Although
aclear decisioncannotbemade with our
pres-ent knowledge, these structures can be
inter-pretedasrecombinantstructures (cf. reference 2, Table 2A, Plate Ia, b, Plate Ila, b). The observation that each branch of the Y-shaped
structures shows the same diameter reduces theprobability that these branches are
stimu-lated byakinked, folded, ortwisted portion of
the molecule. If it were true that uncleaved recombinantstructuresaccumulateinthegene 49-defectiveDNA, leadingtothegenerationof VFS-DNA complexes, it would be conceivable
thatthe same structurescaninterferewith the
process ofheadfilling.
DISCUSSION
VFS-DNA complexes, which accumulate in
cellsinfected with gene 49-defective phage un-der nonpermissive conditions, were isolated, and their structures were analyzed
enzymati-callyandintheelectronmicroscope.
With increasingtimeafter infectionatypical
structureofincreasingcomplexityisdeveloped.
Each complex characteristically consists of a center of tangled strands containing many DNAmolecules and a surrounding area of more looselyarrangedDNAloopsand free ends (Fig. 4). Atpresent, it isdifficult todecide whether the compactcenterportionof thecomplex rep-resents the true structure ofthe DNA in vivo. It cannot be excluded that the structure is
artificially generated during the preparation,
which includes an ethanolic washing step through which the DNA could be collapsed
intotheobservedcomplexstructureby
precipi-tation (16, 17). The in situ lysis procedure,
which avoids theethanol step, leadsto similar resultsregarding the network structureof the DNA. But, again, we cannot be certain that VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.503.55.450.75.369.2]FIG. 9. Electron microscopic examinationofpartiallyfilledDNA-attached heads. The isolation procedure of VFS-DNA carrying cross-linkedgene49-defective headswasthesame asdescribedinthelegendtoFig.8.
Examples ofattached heads were selected forfurther magnifications, and a sequence ofheads with an
increasing amountof DNA wasselected (b-g).Osmotically shocked wild-typephage (a) andcomplete phage (h) were taken as controls and demonstrate the DNA-free and the completely filled state ofthe phage. Magnification, x 90,000.
the danger ofprecipitating the DNA is com- trated form, whichmust be differentfromthe
pletely eliminated. Regardless ofthese objec- organization of the 200S DNA since it is only tions, however,wemayconclude thatthe VFS- generated intheabsence ofP49.
DNA is arranged in a locally highly concen- After isolation,thecomplexesareverystable
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.503.85.434.61.549.2]FUNCTION OF GENE 49 OF PHAGE T4. II. 1013
and they maintain nearly the shape and the size of the cell (Fig. 4b and 5b-d). The s value, which has been taken as a measure for the
integrity of thecomplexes, is hardly changed by further manipulations (13). The stability against shear isremarkable and differs greatly from the observed lability of the regular 200S DNA(cf. reference 7). This tightly packed DNA inside the cell can be reactivated for the process ofhead filling, as was shown by the analysis of thefate of the DNAinvivo (Fig. 1).
Whatcauses thedifference betweenthe VFS-DNA and the 200S DNA? We believe that a
direct DNA-DNA interaction isinvolvedinthe
compact structureofthe VFS-DNA whichis not
realized in the 200S form. Such interaction
could be achieved by recombination between individual DNA molecules if the recombinant
structures are not completely cleaved in the VFS-DNA, thusleading tostable linkages
be-tween individual DNA strands. It also could explain why headfillingdoesnotgoto
comple-tionifone assumesbranched recombinant mol-ecules blocking the entrance of the head. The demonstration of branched DNA structures in
front of attached heads (Fig.10) is in supportof this idea. Gene49function would benecessary to cleave these structures. The observation of enhanced repair synthesis after the
reactiva-tionof P49invivo (22)could beinterpreted as
theresult of this cleavage.
Ifthis model were true, why cantsP49 not
reverse the development of VFS-DNA in the absence of concomitant head filling after per-missiveconditions have beenrestored?
Packag-ingof the VFS-DNA,however,can occur inthis
mutant after shift to permissive temperature withoutsynthesis of proteins,providedall
com-ponents needed for head fillingarepresent. It
seemsconceivableto usthatsomehead-related
structure(s)isneededtoallow tsP49toactonly
onthe DNA molecules thatare actively being packaged. Thefindingsthat thesamets muta-tion (tsC9) does not completely prevent the
generation of VFS-DNA underpermissive
con-ditions (9) and thatextractsfromcellsinfected with this mutant at permissive conditions do not cleave VFS-DNA in vitro (9;
unpublished
data) fit the model.
The fact that notallhead-attachedmolecules
thatcanbeclearly followedover adistance ofat
least several head lengths show these struc-tures is not a strong argument against the model. It is known that the partially filled headsare very labile (19) and theirDNA con-tent is largely dependent on the lysis proce-dure.Therefore,itisconceivablethat theheads loose some ofthealready packed DNA before fixation, thus moving away from the
interfer-ing structure. Improved lysis procedures are needed to investigate this point.
The observation that the single-strand-spe-cificnuclease Si does not cleave the VFS-DNA complexes whereas P49 does so efficiently in vitro (9) does not rule out recombinational
structuresasacomplex buildingagent, because
the single-stranded regions of the hypothetical DNA linkscouldbe filled in by repair enzymes, orfor some other reasons they could be simply notaccessible to the nucleaseS1. The recombi-nant structures studied in T4-infected cells by Broker etal. (1, 2) were generated in the
ab-sence of DNA replication, and the observed single-stranded regions of the viral DNA may be dueto aspecial situation.
Analternative to the recombinational model would beamodelinwhichan unknown
mecha-nism forunwinding the DNA exists, postulat-ing afunction of P49 as an untwisting activity able torelease supertwists of higher order from the DNA. This type of supertwists should be different from the regular ethidium
bromide-sensitive supertwists, which were notdetected
as a main structural component in the VFS-DNA. From our present knowledge of the func-tionof untwisting activities (4, 31) this alterna-tivepossibility seems unlikely to us.
Besides the study of the questions of how the phage DNA is keptinpackagable order inside the cell and how the phage chromosomes are unraveled, it is likewise important to learn how the DNA is arranged inside the head as the
processofhead fillinggoes on.
Thedescribed procedure of cross-linking
par-tially filled headstothe DNA hassome promis-ingpotential in thisrespect, because itallows thestudy of distinct stages during head filling. Thedemonstrated accumulation of the DNAin
locally separated, distinct portions inside the head during early packagingis aninteresting observation. Theperipherallylocalized
concen-tration of thephage DNA within the partially filled capside suggests a preferential
associa-tionof the genome with somespecific areasof the headduring filling. It is possible that the headfillingprocessisinitiatedby aggregation ofviral DNA in the vertexes of thecapsid. A similar model for packaging of DNA in the capsid of adenovirions has been proposed by
Brown etal. (3). These authors havesuggested
that the adenovirus genome is concentrated into 12 DNA protein-containing spheres that areassociatedwith the12 vertexesofthe intact
capsid.
At the present time we cannot decide whetherthe DNAarrangementisduetosome
specificinteraction between the DNAand head protein(s) or ifit is the result oflimited free
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
.iX
t.
-.< Ig
9i
---4
FIG. 10. Electron microscopic examination of branched DNAstructures. The isolation of VFS-DNA with cross-linked, attached gene 49-defective heads was done asdescribed in the legend to Fig. 8. Threeexamplesof branched DNA structures closely situated to the terminally attached heads are shown (a, b, c). The accompanyingdrawings show the interpretation ofeach structure. Magnification, x 76,000. In(d)and (e) twoexamples ofbranched DNA structures without a visible correlation topartially filledheads are shown.
Magnification, x120,000.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:15.503.84.429.67.605.2]FUNCTION OF GENE 49 OF PHAGE T4. IIL 1015
space available at the early stages of
packag-ing,where most of the head interior seems to be occupied by a protein core released
concomi-tantly withDNA packaging by proteolytic
deg-radation (26), giving room for more DNA (19,
20, 28). Studies are in progress in our labora-tory tofurtherinvestigate these questions.
ACKNOWLEDGMENTS
We gratefully acknowledge the help of H. J. Hirsch during the initial work of the electron microscopic analyses, and we aregrateful to E. Kellenberger and H. Wunderli, who made their results available to us before publication.
This work has been supported by the Deutsche For-schungsgemeinschaft through SFB 74.
LITERATURE CITED
1. Broker, T. R. 1973. An electron microscopic analysis of pathwaysforbacteriophage T4 DNA recombination. J.Mol. Biol. 81:1-16.
2. Broker, T. R., and I. R.Lehman. 1971. Branched DNA molecules: intermediates in T4 recombination. J. Mol.Biol. 60:131-149.
3. Brown, D. T., M. Westphal, B. T. Burlingham, U. Winterhoff, and W. Doerfler. 1975. Structure and composition of theadenovirus type 2 core. J. Virol. 16:366-387.
4. Champoux, J.J., and R. Dulbecco. 1972. An activity from mammalian cells that untwists superhelical DNA-A possible swivel for DNA replication. Proc. Natl. Acad. Sci. U.S.A. 69:143-146.
5. Casjens, S., and J. King.1975. Virusassembly.Annu. Rev.Biochem.44:555-611.
6. Delius, H., andH.Worcel.1973. Electron microscopic studies on the folded chromosome of Escherichia coli. ColdSpringHarborSymp.Quant.Biol. 38:53-58. 7. Frankel, F. R. 1966.Studiesonthe nature of
replicat-ing DNA inT4-infected Escherichia coli. J. Mol. Biol. 18:127-143.
8. Frankel,F. R. 1968.DNAreplicationafter T4 infection. Cold SpringHarborSymp. Quant.Biol. 33:485-493. 9. Frankel,F.R.,M. L.Batcheler,and C. K. Clark.1971.
The role of gene 49 in DNA replication andhead morphogenesis in bacteriophage T4. J. Mol. Biol. 62:439-463.
10. Fujisawa,H., and T. Minagawa. 1971. Genetic control of the DNA maturation in the process ofphage
mor-phogenesis.Virology45:289-291.
11. Harrison, D. P., D. T. Brown, and U.C. Bode. 1973. Thelambda head-tailjoining reaction:purification, properties andstructureofbiologicallyactiveheads and tails.J. Mol. Biol. 79:437449.
12. Huberman, J. A. 1968. Visualization of replicating
mammalian and T4bacteriophageDNA.ColdSpring HarborSymp.Quant. Biol. 33:509-524.
13. Kemper, B., andE.Janz.1975. Functionof gene49 in bacteriophageT4. I. Isolationand biochemical char-acterization of veryfast-sedimentingDNA. J. Virol.
18:992-999.
14. King,J. 1968. Assembly of the tail ofbacteriophage T4. J. Mol. Biol. 32:231-262.
15. Kleinschmidt, A., and R. K. Zahn. 1959.Ober Desoxy-ribonucleinsaure Molekeln in Protein-Mischfilmen. Z.Naturforsch. Ser. B 14:770-779.
16. Lang, D. 1969. Collapse of single DNA molecules in ethanol. J. Mol. Biol. 46:209.
17. Lang, D. 1973. Regular superstructures of purified DNA in ethanolicsolutions. J. Mol. Biol. 78:247-254. 18. Laemmli, U. K., F. Beguin, and G.
Gujer-Kellenber-ger. 1970. A factor preventing the major head protein ofbacteriophage T4 from random aggregation. J. Mol. Biol. 47:69-85.
19. Laemmli, U.K., and M. Favre. 1973. Maturation of the head ofbacteriophage T4. I. DNA packaging events. J. Mol. Biol. 80:575-599.
20. Laemmli, U. K., and R. A. Johnson. 1973. Maturation of thehead ofbacteriophage T4. II. Head-related, aberrantT-particles.J.Mol.Biol. 80:601-611. 21. Laemmli, U.K.,E.Molbert, M. Showe, and E.
Kellen-berger. 1970.Form-determining function of the genes required for the assembly of thehead of bacterio-phage T4. J. Mol. Biol. 49:99-113.
22. Laemmli, U. K., N. Teaff, and J. D'Ambrosia. 1974. Maturation of thehead of bacteriophage T4. III. DNA
packagingintopreformedheads. J. Mol. Biol. 88:749-765.
23. Luftig, R. B., and C. Ganz. 1972. Bacteriophage T4 headmorphogenesis.II.Studies on the maturation of gene49-defective head intermediates. J. Virol. 9:377-389.
24. Luftig, R. B., and C. Ganz. 1972. Bacteriophage T4 head morphogenesis. IV.Comparison of gene 16-, 17-, and 49-defectiveheadstructures.J. Virol. 10:545-554. 25. Luftig, R. B., W. B. Wood, and R. Okinaka. 1971. Bacteriophage T4 head morphogenesis. On the
na-tureofgene49-defectiveheadsand their roleas inter-mediates. J. Mol. Biol. 57:555-573.
26. Onorato, L., and M. K.Showe.1975.Gene21 protein-dependent proteolysis in vitro ofpurified gene 22 productofbacteriophageT4. J. Mol.Biol. 92:395-412. 27. Pettijohn, D. E., and R. Hecht. 1973. RNA molecules bound to the folded bacterial genome stabilized DNA folds and segregate domains ofsupercoiling. Cold Spring HarborSymp. Quant.Biol. 38:31-41. 28. Showe, M.K.,and L. W. Black.1973.Assemblycoreof
bacteriophage T4: an intermediateinhead formation. Nature(London)New Biol. 242:70-75.
29. Vandersclice, R. W., and C. D.Yegian.1974.The iden-tification of latebacteriophageT4proteinsonsodium dodecyl sulfatepolyacrylamide gels. Virology 60:265-275.
30. Vogt,V. M. 1973.Purificationandfurtherpropertiesof single-strand-specific nuclease fromAspergillus ory-zae.Eur. J. Biochem. 33:192-200.
31. Wang, J. C. 1971. Interaction between DNA andan
Escherichiacoliproteincw.J.Mol. Biol. 55:523-533. 32. Worcel, A., E. Burgi, J. Robinton, and C. L. Carlson.
1973. Studies on the folded chromosome of Esche-richia coli. Cold Spring HarborSymp. Quant. Biol. 38:43-51.
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/