Copyright© 1976 AmericanSociety forMicrobiology Printed in U-SA.
Bidirectional Replication of Adenovirus Type 2 DNA
MARSHALL S. HORWITZ
Department of Microbiology-Immunology, CellBiology, and Pediatrics, Albert Einstein College of Medicine, Bronx,New York 10461
Received for publication 11 November 1975
After short periods of labeling
with[3H]thymidine,
recently completedadeno-virus DNA
molecules
wereisolated
and cleaved with
restrictionendonucleases.
The strands (heavy and light) of
mostof the
restrictionendonuclease fragments
wereseparated. The
patternof
labeling clearly shows
an asymmetryof
radioac-tivity
onthe isolated strands of each
restrictionendonuclease piece. The data is
consistent with
replication
proceeding
inthe
5' to 3'direction
oneach strand.
Thus, there
is an initiation pointplaced
at or neareach
end of the molecule.
Adenovirus
type 2 DNA, a linear moleculeof 23 x 106daltons, replicates
in the nucleusandproduces approximately
100,000copies/cell
dur-inglytic infection
(9, 10).The
duplex
DNAhas
nonpermuted sequences (6)
and inverted
termi-nal
redundancy
atends
(8, 34),which
areiden-tical for
100 to 140nucleotide
pairs (1).The
inverted terminal redundancy allows the
for-mation
of single-strand
(ss)circles by base
pair-ing
between both ends ofthe denatured
ssmole-cules. Since the ends of adenovirus duplex DNA
are
identical,
these
molecules
cannotbe
con-verted
tocovalently linked double-strand
(ds)
circles like those formed
by
bacteriophage
lambda DNA
(10, 34). Duringreplication,
ssmolecules larger than the
genomehave
notbeen demonstrated
(12, 29, 30);therefore,
nocovalent addition of
progeny DNA toparental
molecules
occurs.There
aresmall pieces of
ade-novirus
DNA
similar
to"Okazaki fragments"
(20),
which
canbe dissociated from the
replica-tion
complex by alkaline denaturation
(2, 12,31, 33).
During
normal viral
replication
it is
notknown if all regions of the
genome arefirst
polymerized
intoOkazaki
fragments,
which
aresubsequently
joined. Replicating
molecules
have
asignificantly
higher
buoyant
density
than
parental
viral DNA
(27, 29). Further
evi-dence has shown that the
density
shift
iscaused
by extensive
ssDNAregions and
notby
RNA-primer
fragments
asreported
for
polyoma
DNAreplication (16).
Recently,
anumber
of
models
of
replication, which include bidirectional
growth,
have beenproposed for adenovirus
DNA.Sussenbach
andco-workers have
pre-sented
datathat
replication
starts attheright
end
(AT
rich) by displacing
theparental heavy
strand
with continuouspolymerization
inthe 5' to3'direction
onthe
light-strand template
(7).
After
adelay, replication
starts onthe
dis-placed
strand at variousinternal
points
ormaystartat
the
3'end of
the template molecule in apattern
of
continuous growth in the oppositedirection
to that on the first strand.Experi-ments
from
our laboratory, in whichadenovi-rusDNA
half-molecules
were produced byme-chanical shearing, have shown
that there isbidirectional growth
with two terminationsites,
oneonthe left and another
on the righthalf
of the
molecule (13).The
presentstudy extends these findings byusing
two restrictionendonucleases,
whichhave allowed
us to examine nine regionsof theDNA. The method
todetermine the
originand
terminus
of DNA
replication
issimilar
tothat
employed by Dintzis
(5) toanalyze the direction
of
replication
of
polypeptides and
morerecently
by
Danna and
Nathans
(4) toisolate the
originand
direction of replication
of simian virus 40DNA. When
aradioactive
precursor is added toa system
synthesizing DNA, the
label entersreplicating molecules
at a growing point that is at adifferent site
ineach molecule. This
as-sumes
that
the labeling procedure does
notchange
the
rateof
DNAsynthesis by
synchro-nizing
molecules
at anyphase
of the replicationcycle.
The
molecules,
which
arecompleted
dur-ing
labeling periods shorter than the total
syn-thesis
timeof the
macromolecule,
will be
pref-erentially
labeled
atthe terminus.
Therefore,
when
completed molecules
areeffectively
sepa-rated from
replicating molecules, the
amountof
radioactivity
invarious
regions
of the
com-pleted molecules
will reflect theorigin anddi-rection
of
synthesis.
In this report the pattern
of
labeling of
dsDNAindicates
highest
specific activity
atbothends of the
DNA,
which is consistent with atermination site at
each end of
themolecule.
While
thismanuscript
wasinpreparation,
sim-ilar results
werereported
(23, 28).
However, examination
of
thesingle-strands
307
on November 10, 2019 by guest
http://jvi.asm.org/
308 HORWITZ
of each
of these
DNAfragments
has shown that the heavy and light strands labelasynchro-nously. This difference in labeling ismost
con-sistent with replication in which each strand is
synthesized in the 5'to3' direction starting at
opposite ends of the molecule.
(A preliminary report of these results was
presented at the Cold Spring Harbor Tumor
Virus Meeting, Cold Spring Harbor, N. Y.,
Au-gust, 1975.)
MATERIALS AND METHODS
Cells and viruses. Thesourceof HeLacells, ade-novirus type 2,andthe conditions of infection have been previously described (17). All experiments weredone in suspensionculturesataninput multi-plicity of 4,000virions/cell (100 to200PFU/cell).
Radioactive labeling of cells. At 18h postinfec-tion,theinfected HeLa cellswerecentrifuged for2 min at 1,500 rpm in anInternationalPRJcentrifuge
andresuspendedat aconcentration of 107cells/mlin
Eagle spinner medium with 5% fetal calf serum. After 5 min of temperature equilibrationin a water bath at 37 C, the cells were radioactively labeled
with [3H]thymidine at 0.25 mCi/ml (40 to 60 Ci/
mmol). The incorporation of radioactivity was stoppedby dilutingthe cellsin 7volumes ofice-cold Earle salts, rapidly centrifuging the cells at 1,500 rpm, andadding 0.2%sodium dodecyl sulfatein0.01 MTris-EDTA (pH 7.4). Usingtheseconditions, the incorporation ofradioactivity was linear for 60 min without anyappreciablelagatthebeginningof this interval (14).
Purificationof viral DNA. Intact [3H]thymidine-labeled viral DNA was purified from cells after pre-cipitating large-molecular-weight cell DNA by a modification of the Hirt procedure (11, 29, 30). Cells (3 x 107)weresuspended in 2 ml of 0.01 M Tris, 0.01 M EDTA (pH 7.4) at 0 C. Sodium dodecyl sulfate (0.2%) and 500 ,ugof Pronase(preincubated at 37 C
for2htodigestanyresidual nucleases) wereadded,
and the mixturewasincubated for 15 min at 30 C. The volume was increased to 9 ml by the addition of the Tris-EDTA buffer, which contained 1% sodium dodecyl sulfate. After a 5-min incubation at 30 C, NaCl wasadded to a final concentration of 1 M. The solutionwasleftat 4C for16h,andthe precipitate wasremoved bycentrifugationat12,000 rpmfor 20 mininaSpincoangle 30 rotor. By processing 1.5 x 107to3 x 107cellsina finalvolumeof10ml,80%of newlyreplicated adenovirus DNA was recovered in the Hirt supernatant. Viral DNAwasprecipitated from the supernatant with2volumes of ethanol. The DNA, redissolvedin 1mlof 0.01x SSC (SSC = 0.15 MNaCl+0.015 Msodium citrate), was centrifuged inanSW27 rotorof the Spinco ultracentrifuge (16 h at 22,000 rpm) on 16-ml, 5 to 20% neutralsucrose gradients containing 1 M NaCl, 0.01 M phosphate buffer, and 0.01 M EDTA. The 31S fractions were
pooled, dialyzed against 0.3M NaCl, 0.01 M Tris,
0.01 M EDTA (pH 8.1), and loaded onto 2-ml col-umnsofbenzoyl-naphthoyl-DEAE-cellulose
(BND-cellulose). The ds DNA waseluted with 1 MNaCl, and the DNAcontaining any ssregion waseluted with 1 MNaCl and 2% caffeine inthe same Tris-EDTA buffer (29).The appropriate column fractions
were precipitated with 2 volumes of ethanol and
redissolved in 0.01 x SSC.
['4C]thymidine-labeled viral DNA, which was used as auniformly labeled marker, was purified by disrupting virionwhichhad beenbandedtwice on CsCldensitygradients(12).
Restriction endonucleases. The enzymes from both Escherichia coli(EcoRI)andHaemophilus par-ainfluenzae (HpaI) were purified from bacterial strainsobtainedfrom the ColdSpringHarbor Labo-ratory. The bacteria were grown and the enzymes
were purified as previously described (19, 25). All
endonucleasedigestswereincubatedfor4hat37C in 10 mM Tris-hydrochloride (pH 7.4) with 10 mM
MgCl2, 6 mM KCl, and 1 mM dithiothreitol. The
reaction wasstopped in 0.04 MEDTA; the solution wasadjusted to a final concentration of10%sucrose and 0.1% bromophenol blue. The DNA fragments wereseparated by electrophoresis on cylindrical gels (1.6by 35 cm)of1.4%agarose inTris-EDTA-acetate (TEA = 40mMTris-hydrochloride, 1 mM EDTA, 5 mM sodium acetate) buffer for 16 h at 100 V. The gelswerestained in thesameTEAbuffer containing
0.5 ,ugofethidiumbromide perml, and thebands
werevisualized with aUV-lightsource(25). For separation of the strands ofeach restriction endonuclease fragment ofDNA, the ds fragments were cut outof the agarosegelafterstaining with ethidiumbromide and visualizationbyminimal ex-posure to UV irradiation. The short cyclindrical pieceof gel wasplaced into aglassscintillation vial with10mlof 0.2MNaOH for 2.5 h at room tempera-ture. The NaOH was decanted, and the gel was soaked for a further 2 h in TEA electrophoresis buffer at 0 C. Thegelslicewasplacedback intothe
electrophoresis tube, anda new column ofgelwas
polymerizedovertheoriginalslice. The agarose was
pouredafterequilibrationat60C.After the
polym-erization, thegeltubewasinverted,and the
electro-phoresis wasperformed under identical conditions
as used for the original separation of the ds frag-ments.The gel werestainedwith ethidiumbromide
andthe single strands were visualized. The
sepa-ratedstrands were cut from the gel, and the radioac-tivity wascounted.
Radioactivityinthe gel slices was quantitated by
remelting the agarose in an autoclave and then adding 10mlofscintillation fluid [1 part Triton, 2 partstoluene, 5 g of 2,5-diphenyloxazole (PPO) per liter, and 50 mg of1,4-bis-(5-phenyloxazolyl)benzene (POPOP) per liter], which had been heated to 50 C. The samples, which were shaken immediately after the addition of scintillation fluid, were cooled to room temperatureandcounted in the ambient tem-peraturescintillation counter(Beckman LS 230).
Reagents.
[3Hlthymidine
(40 to 60 Ci/mmol) and['4C]thymidine
(57mCi/mmol) were purchased from Schwarz BioResearch, Inc. Agarose was obtained from Sargent Welch Co. BND-cellulose was pur-chased from Serva.J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS DNA
309
RESULTS
Isolation of recently completed viral DNA
molecules. Eighteen hours postinfection, cells
wereradioactively labeled with [3H]thymidine
and viral DNA wasseparated from host DNA
by using the Hirt procedure.The Hirt
superna-tant, containing replicating and completed
viral DNA molecules, wascentrifugedon
neu-tral sucrosegradients as shown in Fig. 1. The
completed molecules (31S) sediment to the
re-gion in the gradient that peaks atfraction 14.
The heterogeneous population of replicating
molecules sediment faster andappearbetween
fractions1and14(29).The completedmolecules
were further purified free of replicating
mole-')
t0
0. C-)
I
u0
10
C-)
I0 15 20 top
WCTION
NUMBER
FIG. 1. Sedimentation velocitygradients to
sepa-rate replicating from completed viral DNA. Pulse-labeled DNAisolated from the Hirtsupernatantwas
centrifugedonneutralsucrosegradientsasdescribed
inMaterialsandMethods. The gradientswere
frac-tionated into 0.75-mlaliquots, and the radioactivity
wasdeterminedby removing20plfromeachfraction
forquantitatingthetrichloroacetic acid-precipitable DNA.Fractionsintheregionof31S-completedviral DNA (fraction14)werepooledasdesignatedby the
bar,andthe DNAwasdialyzedagainst 0.3M NaCl,
0.01 M EDTA, and 0.01 M Tris (pH 8.1) before chromatographyonBND-cellulose(Fig. 2).Symbols:
0,5-minpulse; 0,15-minpulse.
cules
onBND-cellulose, which eliminates any
DNAwith
ssregions. The results of
BND-cellu-lose chromatography of the 5-, 10-,
15-,and
240-minsamples
areshown in Fig. 2. The first peaktoelute
(fraction 2)
containsthe
completely
dsmolecules, and
thesecond peak
(fraction
8)rep-resents any
molecules with
ssregions.Greater
than
99.3%of
DNA, purified from virion
andsimilarly treated, elutes
inthe ds region. When
DNA
from the virion
isdenatured by boiling
before
chromatography,
100%of the DNA
sub-sequently elutes in fraction
8,the
ssregion
(data
notshown). With the
shortest pulse
timesexamined (5 min),
mostof the DNA from
the31S region
of
the gradient has
some ss regions,which
are onmolecules that have
justinitiated
replication.
By 15min,
the
31S region of thegradient has
morelabeled
ds than
ssmolecules,
and
by
4h
76%of the
labeled
DNA is inds
molecules.
Determination
ofspecific activity of dsre-gions of
pulse-labeled
DNA. Theds[3H]-
thym-idine-labeled DNA from fraction
2(Fig. 2) wasmixed with
uniformly labeled
[14C]thymidine-containing DNA isolated from the adenovirion.
The DNAs
weredigested either with the
re-striction
endonuclease EcoRI
or amixture of
this
enzymeand that derived from
HpaI. The
patterns
of
digestion for EcoRI and
HpaI
have
been elucidated
(18). The
pertinent restriction
endonuclease
pieces
aredrawn
toscale
onthe
abscissa of
Fig.
4.The EcoRI
pieces
arede-signated
incapital letters (B
toF) and the
HpaI
fragments
aredesignated
inlowercase
letters
(e, c,f,
a)."C-" and "E-" refer
toEcoRI
fragments further
digested by
HpaI with
theloss
of
approximately
1.5%of the genome.
The results of
atypical EcoRI
+HpaI
restric-tion
endonuclease
digest
of the mixture of
[3H]DNA
pulse-labeled for
5 minand the
[U-'4C]viral DNA
areshown in
Fig.
3.Peaks with
the
higher
ratios of 3H- to'4C-labeled DNA
include
"c,"
"C-,"
and
"e,"
which
are nearboth
ends of the molecules
(see
Fig. 4).
A
summaryof results obtained from
5-,
10-,
and 15-min
pulse-labeled DNAs
areshown
inFig.
4.The
lowest
specific
activity
(3H/14C)
is inthe
region
of fractions
"a"and
"B" nearthe
centerof the
molecule,
and the
specific activity
increases
to-ward both
ends. As
expected,
differences in the
specific activity
of the individualpieces
aregreatest
for the
5-mindigest,
less
at 10min,
and
minororabsentby
15min.This
patternof
specific
activity
clearly
showsbidirectional
growth
of the
molecule,
but
anumber of
quite
different models could
explain
this
data.
To
differentiate between
numerouson November 10, 2019 by guest
http://jvi.asm.org/
[image:3.509.63.248.242.519.2]310 HORWITZ
.10
Ca.2
I
~~~~~~~~~~~~~~~~~~~~~5
5 10 5 10
F R A C T I O N N U M B E R
FIG. 2.BND-cellulosechromatography toseparate replicatingfromcompletedviral DNA. Two-milliliter columnsofBND-cellulosewerepreparedin2.5-mldisposableplastic syringes by wettingthe cellulose in 0.3 M
NaCl, 0.01 MEDTA, 0.01 M Tris buffer (pH 8.1) containing 20% ethanol. The ethanolwas removed by
centrifugation ofthe resin immediatelyafter suspension ofthe cellulose. The BND was poured into the columns andextensivelywashed with thesamebufferwithout ethanol until theoptical densityof theeffluent at260nmwasless than 0.05. TheDNA, which had beendialyzed againstthe columnbuffer,wasloadedonto the BNDand washed with 12 mlofthesamebuffer.Elutionofds DNAwasachieved with 12 mlofthe column
buffertowhich NaCI had been addedtoafinalconcentrationof1M.Aliquots (2 ml)werecollectedduringthe
elation,and
50-pJ
sampleswereremovedforquantitationofthe DNAineachfraction.The columnwasthenwashed with 12 mlof the latter buffer,towhich2%caffeine(designated bythe arrow) had been added. Thess DNA elated after the addition of caffeine andwas quantitated by taking
50-pl
aliquotsfor radioactivity counting. The ds DNA, elatedinfraction 2,wasprecipitatedwith2volumesofethanol. (A) *,5min; 0,10 min.(B) A,15 min; A, 240 min.ties,
it was necessary toknow if
replication
wassimilar
onboth strands. Sharp
etal.
(24)have
shown that the
denatured heavy (H) and light
(L)
strands ofmost
of the restriction
endonucle-asefragments
canbe successfully separated
onagarose
gels.
Usingthe alkaline denaturation
technique for
DNAembedded
inagarose
(seeabove),
wehave been able
to separate H and Lstrands from the EcoRI
"B."
"C," "D," "E,"
and"F" piecesand the HpaI "e" and "c" pieces.
Determination of specific activity of ss
re-gions
of pulse-labeled DNA. The ratio of 3H(pulse)-
to 14C(uniformly)-labeled
DNA wasde-termined for the separated
strands ofeachre-striction
endonuclease
piece isolated fromre-cently completed duplex
DNA asdescribed
inMaterials and
Methods. The
extent ofstrand
separation
for several ofthe fragments is shown
inFig. 5.
The strand separation of DNA
dena-tured within the
agarosegel
is
superior
todena-turation in
solution
notonly
because of the
relative
speed
of the former
technique
but
also
because
of
the
decreased
amountof renatured
DNA
found
after
electrophoresis.
In
mostex-periments,
asinthe
oneshown
inFig.
5,there
is norenatured
DNAdetectable by
ethidiumbromide staining. The
assignmentof
strandspecificity (H
orL) for the isolated strands
fol-lows the designation of Sharp
etal.
(24). Thefaster
movingband for the EcoRI "B." "C,"
"D,"
"E," and "F" and the slower
band for HpaI "e"and "c"
(P.
A.Sharp, personal
communication)belong
tothe
samestrand, which
isthe heavy
strand
inalkaline
CsCl gradients. The
HpaI "a"fragment does
not separate into H and Lstrands
under
anyconditions
tried thus far.J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.509.106.414.53.339.2]oc B C- D
I, I E-FeI fI
go
6
a-10 20 30 40 50
F R AC T I 0 N N U M B E R
FIG. 3. Agarose gel electrophoresis of a restriction endonuclease digest of the5-minpulse-labeled DNA. The ds viral DNA from fraction 2 (Fig. 2), which was extracted from cellspulse-labeled for5 min with
[3H]thymidine,wasmixed with
["4C]thymidine-containing
DNApurifiedfrom adenovirions. The mixturewasdigested with EcoRI andHpaIfor4h asdescribedinMaterialsand Methods. Afterelectrophoresisofthe DNA on1.4%agarosecolumns for 16 h at 100 V, the gel was stained withethidium bromide and the DNA bands werelocated. The gel was sliced into aliquots starting with acutbetween the 'a" and Sc" piecestoinsurethat these closely migrating fractions were in separate gel slices. The gels were melted, and theradioactivitywas quantitated by scintillation counting. The designation of the restriction endonuclease fragments isas de-scribed in thetext. Thelocationofthefragments and theirrelative sizes areshown on the abscissa ofFig. 4.
Fragments"E-" and "F' donotseparatein thissystem.
In Fig.
6 and 7, a summary of the specific
activities of the isolated strands is shown for
pulse-labeling
periods of 5 and 10
min,
respec-tively. It
isclear that the specific activity
isquite
different for the corresponding regions on
both
strands, and this suggests that the
Lstrand
(0)
isreplicating from left to right and
the
Hstrand
(0)
from right to left.
Single-strand
data for the "e" fragment at the extreme
left end of the molecule is not shown in the
figures
but
ispresented
inTable 1. The high
specific activity on the
Lstrand
of the "e"
frag-.
ment and
the
lowspecific activity
onthe
Hstrand do not continue the trend of labeling
derived from quantitating replication
on theother
96%of
the molecule.
DISCUSSION
The data
is most consistentwith
bidirec-tional
growth,
which isasymmetrical
oneachof the strands of
adenovirus type 2 DNA. The
light strand appears
toinitiate
ator
nearthe
left hand end and replicate continuously to the
right. This corresponds to growth
in the 5' to 3'direction
according tothe
data ofSharp
etal.,
whoassigned
the 5' end ofthe
heavy
strand tothe right (24).
Incontrast, the
heavy
strandinitiates
onthe
right and
replicates toward the
lefthand end. This model of
replication does
notrequire
Okazaki
fragments
tosuccessfully
propagate
eitherchain,
although
severalinves-tigators have
reported the
presence of theseintermediates.
There
appears
to be adiscrepancy
oflabeling
at
the lefthand
end of the
molecule, which
wasdetected by
examining
theHpaI
"e"piece
repre-senting
4%of the
DNA.The
light
"e"strand
had more
radioactivity
thanexpected for
anorigin of
replication.
This could
occurbecause
the
originof replication
containsnucleotides
311
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.509.112.403.61.346.2]312
HORWITZ
such
asRNA serving a primer function. Since
the RNA would
have
tobe
excised and replaced
with
deoxynucleotides,
this
region
may be
la-beled by
thymidine with kinetics similar
to aterminus. This would
occur ifthe RNA
wereremoved toward the end of the
replication
onthat
particular strand.
Another possible
explanation
is that the
rep-1.2*
0
o
-
0.-e c f a B F D E C
RESTRICTION ENDONUCLEASE FRAGMENT
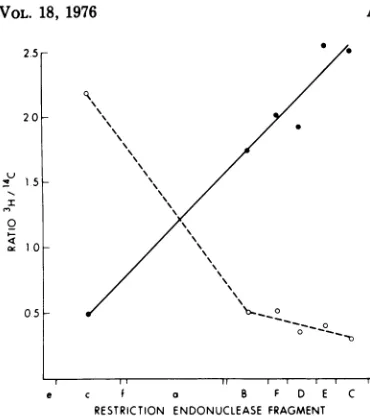
FIG. 4. Orderoflabeling ofselectedregionsofds adenovirus DNA. From data suchas thatinFig.3, ratios ofpulse-labeled[3H]DNAtouniformly labeled
['4C]DNA
were calculated. Included are the data from the 5-, 10-, and15-min pulse-labeling periodsobtainedeither from digests usingEcoRI alone
(frag-mentsB, F,D,E, C) ortogether with theHpaI
en-zyme (fragments e, c, f, a). This results from the
three time points are normalized to give identical ratios for the "e" fragment. Symbols: 0, 5 min; 0, 10min; A, 15 min.
licating
molecule
is acircle,
and
that
replica-tion
continues
beyond
the
molecular end to
anextent
of
4%of the genome.
Although
linear
adenovirus DNA molecules have terminal
re-dundancy,
it is
ofthe
inverted
type,
which
does
3
Or-25k
20
0 15
1 0
05
e c f a 8 F D E C
RESTRICTION ENDONUCLEASE FRAGMENT
FIG. 6. Order
of labeling
(5min) ofselectedre-gions of
ssadenovirusDNA. DNA waslabeled with[3H]thymidine for
5mininadenovirus-infectedcellsat18hpostinfection. The recently completed
duplex
viral DNA was purified and digested either with EcoRI
for fragments B, F,
D, E,andCorwith EcoRI+
HpaI for fragments
e, c, f, and a; the DNA waselectrophoresed
onagarosegels
as describedinFig.
3.Selectedfragments
wereprocessedtoseparatethe H and Lstrandsaccordingtothe descriptioninFig.
5 andinMaterials andMethods.The specificactivityof
thepulse-labeled
DNA(PH)
in relation touni-formly
labeled DNA (14C) wasdeterminedand plot-tedinrelationtoits mappositionontheadenovirus chromosome.Symbols: *,
Lstrand; 0,H strand.t
c,
r..>
I
R
Hpo.
e
[image:6.509.267.451.137.317.2]Hpii
or
FIG. 5. Strand separation ofrestrictionendonuclease fragments. Pulse-labeled DNA
([3H]thymidine)
was mixedwithuniformly-labeled DNA(['4C]thymidine)
anddigested eitherwithEcoRIorHpaI.
Thepieceswere separated onagarose gels, stained, andcutfrom thegel.After soaking thegelslices in 0.2 MNaOHto denature the DNA, the endonuclease fragments werererun onneutralagarosegelsasdescribedinMaterials andMethods. The results ofstrand separationareshown forseveralof thefragments.AlthoughtheHpaI"a" piecedoes notseparate intoHandLstrands, theHpaI "c"and the EcoRI"B"fragmentseparate intotwobands. (For otherfragments successfully separated by this technique, see text).
III
I~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.509.62.248.159.360.2] [image:6.509.66.457.472.584.2]qS 20h
u
I
0
or 10
05K
e c f B F D E C RESTRICTION ENDONUCLEASEFRAGMENT
[image:7.509.62.247.56.265.2]FIG. 7. Order of labeling (10 min) of selected re-gionsof ss adenovirus DNA. Infected cells were la-beled with [3H]thymidine for 10 min in the same experiment as described in Fig. 6. The DNA was processed and quantitated exactly as described for the 5-min sample.
TABLE 1. Specific activitiesof the heavy and light strandsseparated from the
HpaI
"e"fragment after5-or10-minlabeling
5min 10min
Strand
"e "c" "e" "C"
Heavy 2.3 3.1 0.8 2.2
Light 7.8 0.5 6.5 0.5
aThe "e" fragment wasobtained from the same
DNAs as shown in Fig. 6 and 7. The "e" heavy strandcorrespondstothe DNAplottedas
(0)
inFig. 6 and 7, and the specific activities (pulse-labeledPH]DNA/uniformly labeled [14C]DNA) shown were
calculated asforthefigures. Data for the "c" frag-mentfrom Fig. 6 and 7 are shown for comparison.
not
allow the usual
typesof DNA-DNA
interac-tions
atthe ends
tofacilitate circularization of
duplex DNA. Recently,
aprotein that holds the
ends of
adenovirus DNA
together
has
been
pur-ified with the
DNAfrom virions (22).
However,
no
circular
forms have
yetbeen
recognized
asintermediates in adenovirusDNA
synthesis.
It
isalso
possible
thatthere
is anonspecific
5'-exonuclease
digesting
small
portions of
theDNA. If
these
regions wererepaired,
thymi-dine label would
appearatthe
ends of
thecom-pleted
DNAmolecules. Of the three
proposed
explanations,
only
the model
of areplicating
circle would also
explain
the
specific activity
ofthe "e"
heavy strand, which is lower than
ex-pected for its position next tothe "c"
fragment.
Similar data is not yet available from the
righthand end of the molecule. The HpaI "g"
fragment,which is 1.4% of the genome, is at the
right end. Although the "g" band was
visual-ized with ethidium bromide staining
and
ap-peared in
fraction
44(Fig. 3),the "g"radioactiv-ity wasalways superimposed on a background
of counts in
fractions
45through
47, whichmakes
exactcalculations difficult. We have not yet been able to separate the strands of the "g" fragment and are approaching the quantitationof counts on
each of the strands by
hybridiza-tion of duplex "g" fragment with isolated
strands ofthe EcoRI "C" fragment.
It is alsopossible that labeling at the
molecu-lar ends may be complicated by an inability to
separate newly initiated molecules with very
short
replicating
regions from the pool of ds,31S completed molecules. It is difficult to
de-sign
acontrol
experimentfor
thispossibility.
The models of adenovirus replication as a
linear
molecule fail to provide a mechanism forthe synthesis of the
5'ends of the
DNA. If RNA is a primer in this system, there would not be away to fill
the
gap atthe ends
uponthe
removalof
RNA, because all
theknown
DNA polymer-asesrequire
aprimer nucleotide
sequencebe-fore
elongation
can occur.This
problem
hasbeen solved by the formation of
concatameresfor thereplication of a molecule such as
bacteri-ophage T7, which
replicates
as a linear DNA(32).
Concatameres, which
arejoined
moleculeslonger than
unitlength, allow the end of
onemolecule
to serve as aprimer
for another.
No suchcovalently
linked concatameres have beenfound
during adenovirus
DNAreplication,
al-though small quantities of viral DNA
may sedi-mentfaster than
genomelength
onalkaline
gradients
(3).These larger molecules
havenever
been shown
tobe labeled with the
kinet-ics
expected of replicating intermediates,
arereported
tobe linked
tohost cell
DNA, and
maybe important
inthe
integration of viral
DNA inhost chromosomes even during the lytic
cycle.
The data
presented in this report are consist-entwith
oneof the models
for bidirectionalreplication,
originally reported by Sussenbach et al.(27).
However,
ourobservations
are notconsistent withSussenbach's proposal that
rep-lication
onthe displaced strandoften
begins atone or several
internal
initiation sites.Al-though
ourdata
do not specify on whichmolec-ular end the first
round of displacementsynthe-sis occurs,
Sussenbach
proposed thatreplica-tion
always
began at the right end by displac-ing the heavy parental strand. His observationIf1 I- - T
18,
2.5r
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.509.61.255.367.432.2]314
HORWITZ
of the displacement of
only the heavy strand
has been
questioned by several
other
investiga-tors (15, 28).
This
asymmetrical
model
of bidirectional
replication
for
adenovirus DNA
issimilar
tomodels
reported for the replication of
mitochon-drial DNA (21).
Another similarity between
these
twosystems is
the
relative resistance of
the
replication of both DNAs to
inhibitors of
protein
synthesis (14, 26).
The
uncoupling
of
DNA
synthesis from its
usual strict
dependence
onnew
protein
synthesis may
depend
inpart
onthe
displacement model of replication
shared
by
both mitochondrial and adenovirus DNAs.
ACKNOWLEDGMENTS
This investigation wassupported by Public Health Ser-vice grant CA-11502from the National Cancer Institute. Marshall S. Horwitz is the recipient ofa Public Health Service CareerDevelopmentAward from theNational Can-cerInstitute(1K04CA-35554).
Iwish to thank Arthur Davino for experttechnical as-sistance,JerardHurwitzforhelpful discussions,and Ste-phen Baum,Susan Horwitz, and Matthew Scharff for criti-calreadingof the manuscript. Ialsowish tothank Julian Panforproviding EcoRI enzyme for some of the pilotstudies and MartinFarber for furnishing the bacterial strainsused toproducethe restrictionendonucleases.
LITERATURE CITED
1. Arrand, J. R., W. Keller, and R. J. Roberts. 1974. Extent ofterminalrepetition inadenovirus2DNA. ColdSpring Harbor Symp. Quant.Biol. 39:401-407. 2. Bellet, A. J. D.,andH.B.Younghusband.1972.
Repli-cation of the DNAof chick embryo lethal orphan virus. J.Mol. Biol.72:691-709.
3. Burger,H., and W. Doerfler.1975.Intracellular forms ofadenovirusDNA. III. Integration of theDNAof adenovirustype 2 intohost DNA inproductively in-fectedcells. J. Virol. 13:975-992.
4. Danna, K. J., and D. Nathans. 1972. Bidirectional repli-cationofsimian virus 40DNA. Proc.Natl. Acad. Sci. U.S.A. 69:309-310.
5. Dintzis,H. 1961. Assemblyofpeptidechains of hemo-globin.Proc.Natl.Acad. Sci.U.S.A. 57:247-261. 6. Doerfler,W.,and A. K.Kleinschmidt.1970.
Denatura-tion pattern of the DNA of adenovirus type 2 as
determined by electron microscopy. J. Mol. Biol. 50:579-593.
7. Ellens,D.J.,J. D.Sussenbach, and H.S.Janz. 1974. Studiesonthe mechanism ofreplicationof adenovi-rus DNA. III. Electron microscopy of replicating DNA.Virology 61:427-442.
8. Garon, C. F., K. W. Berry, and J. A. Rose. 1972. A unique form of terminalredundancy in adenovirus DNA molecules. Proc. Natl. Acad. Sci. U.S.A. 69:2391-2395.
9. Green, M. 1962. Studies on the biosynthesis of viral DNA. Cold Spring Harbor Symp. Quant. Biol. 27:219-233.
10. Green, M., M. Pifia, R. Kimes, P. C. Wensink, L. A. MacHattie, and C. A. Thomas, Jr. 1967. Adenovirus DNA. I. Molecular weight and conformation. Proc. Natl.Acad. Sci. U.S.A. 57:1302-1309.
11. Hirt, B. 1967. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 26:365-369.
J. VIROL. 12. Horwitz, M.S. 1971. Intermediates in the synthesis of type 2 adenovirus deoxyribonucleic acid. J. Virol. 8:675-683.
13. Horwitz, M. S. 1974. Location of the origin of DNA replication in adenovirus type 2. J. Virol. 13:1046-1054.
14. Horwitz, M. S., C. Brayton, and S. G. Baum. 1973. Synthesis of type 2adenovirus DNA in the presence ofcycloheximide.J. Virol. 11:544-551.
15. Lavelle, G., C. Patch, G. Khoury, and J. Rose. 1975. Isolation and partial characterization of single-stranded adenoviral DNA produced during synthesis ofadenovirus type 2 DNA. J. Virol. 16:775-782. 16. Magnusson, G., V. Pigiet, E. L. Winnacker, R.
Abrams, and P. Reichard. 1973. RNA-linked DNA fragments during polyoma DNA replication. Proc. Natl.Acad. Sci. U.S.A. 70:412-415.
17. Maizel, J. V., Jr., D. 0.White, and M. D. Scharff. 1968. Thepolypeptidesofadenovirus.I.Evidence for multi-ple protein components in the virion and a compari-son of type 2, 7a and 12.Virology 36:115-125. 18. Mulder, C.,J. R. Arrand, H. Delius, W. Keller, U.
Pettersson, R. J. Roberts, and P. A. Sharp. 1974. Cleavage maps of DNA from adenovirus types 2 and 5 by restriction endonucleases EcoRI and HpaI. Cold SpringHarborSymp.Quant. Biol. 39:397-400. 19. Mulder, C., and H.Delius.1972.Specificityof the break
produced by restriction endonuclease R1 in simian virus 40 DNA as revealed by partial denaturation mapping. Proc.Natl. Acad.Sci.U.S.A.69:3215-3219. 20. Okazaki, R. T., T.Okazaki, K.Sakabe, K. Sugimoto, R. Kainung, A. Sugino, and N. Iwatsuki. 1968. In vitromechanism of DNA chaingrowth. Cold Spring Harbor Symp. Quant. Biol. 33:129-143.
21. Robberson, D. L., H. Kasamatsu, and J. Vinograd. 1972. Replication of mitochondrial DNA. Circular replicative intermediates in mouse L cells. Proc. Natl. Acad. Sci. U.S.A.69:737-741.
22. Robinson,A. J., H. B.Younghusband, and A.J. D. Bellett. 1973.AcircularDNA-protein complexfrom adenoviruses.Virology56:54-69.
23. Schilling,R., B.Weingartner, andE. L. Winnacker. 1975.Adenovirustype 2 DNAreplication.II.Termini of DNAreplication.J.Virol. 16:767-774.
24. Sharp,P. A., P. H. Gallimore, and S. J. Flint. 1974. Mapping of adenovirus2RNA sequences inlyrically infected cellsandtransformed cell lines. ColdSpring Harbor Symp.Quant.Biol.39:457474.
25. Sharp,P.A., B.Sugden,and J.Sambrook. 1973. Detec-tion of two restricDetec-tionendonuclease activities in Hae-mophilus parainfluenzae using analytical agarose ethidium bromide electrophoresis. Biochemistry 12:3055-3063.
26. Storrie, B., and G. Attardi. 1972. Expression of the mitochondrialgenome inHeLa cells. XIII. Effect of selective inhibition ofcytoplasmicormitochondrial protein synthesis onmitochondrial nucleic acid syn-thesis. J. Mol. Biol. 71:177-199.
27. Susenbach,J. S., P. C. VanderVliet, D. J. Ellens, and H.S.Janz. 1972. Linear intermediates in the replica-tionof adenovirus DNA. Nature (London) New Biol. 239:4749.
28. Tolun, A., and U. Pettersson. 1975. Termination sites for adenovirus type 2 DNA replication. J. Virol. 16:759-766.
29. Vander Eb, A. J. 1973.Intermediates in type 5 adenovi-rusDNAreplication. Virology 51:11-23.
30. VanderVliet,P.C., and J.S.Sussenbach. 1972. The mechanism ofadenovirus DNA synthesis in isolated nuclei. Eur. J. Biochem. 30:584-592.
31. VlIak,J.M., T. H.Rozin,and J.S. Sussenbach. 1975. Studies on the mechanism ofreplicationof
on November 10, 2019 by guest
http://jvi.asm.org/
rusDNA. IV. Discontinuous DNA chain propagation. Virology 63:168-175.
32. Watson, J. D. 1972. Origin of concatemeric T7 DNA. Nature(London) New Biol. 239:197-201.
33. Winnacker,E.L. 1975.Adenovirus type 2 DNA
replica-)ENOVIRUS REPLICATION
315
tion. I.Evidence for discontinuousDNAsynthesis. J. Virol. 15:744-758.
34. Wolfson, J.,and D.Dressier.1972.Adenovirus-2DNA contains aninverted terminal repetition. Proc. Natl. Acad. Sci.U.S.A.69:3054-3057.

![FIG. 3.Fragments[3H]thymidine,scribedonquantitateddigestedweretheseThe 1.4% Agarose gel electrophoresis of a restriction endonuclease digest of the 5-min pulse-labeled DNA](https://thumb-us.123doks.com/thumbv2/123dok_us/1561322.108746/5.509.112.403.61.346/fragments-thymidine-scribedonquantitateddigestedwerethesethe-agarose-electrophoresis-restriction-endonuclease-labeled.webp)
![FIG. 6.formlychromosome.Hofted3.5EcoRIgionselectrophoresed[3H]thymidineatviral+ and Selected HpaI and 18 Order of labeling (5 min) of selected re- of ss adenovirus DNA](https://thumb-us.123doks.com/thumbv2/123dok_us/1561322.108746/6.509.62.248.159.360/formlychromosome-hofted-ecorigionselectrophoresed-thymidineatviral-selected-labeling-selected-adenovirus.webp)