JOUIRNAL OFVIROLOGY, June 1972, p. 930-937 Copyright @,1972 AmcricanSociety for Microbiology
Vol. 9, No. 6 PrinitedinU.S.A.
Structure
aiid Fuiiction
of
the
Polypeptides
in
Simian
Virus
40
II.
Transcription
of
Subviral
Deoxynucleoprotein
Complexes
In
Vitro
ENG-SHANG HUANG, MEIHAN NONOYAMA, AND JOSEPH S. PAGANO
Department oJ Bacteriology aticl Immnunology andtc Departnlent ofMelicine, School oJ Medicine, Uniiversiti of North Cciroliia, CliapelHill, North Carolina 27514
Receivedforpublication 21 January 1972
A deoxynucleoprotein complex (DNP-1) isolated from simian virus 40 (SV40)
after disruption of the virus in an alkaline buffer contains the viral
deoxyribonu-cleic acid (DNA) and four minor structural polypeptides. Dissociation of DNP-I
by equilibrium centrifugation in CsCl yields a complex (DNP-II) that contains a
smallamount ofpolypeptide tightly bound to the viral DNA. Studies ofthe
tem-plate activity ofthese deoxynucleoprotein complexes in vitro with Escherichia coli transcriptase show that therate oftranscription of DNP-I and DNP-IL is 30 and
80%,,
respectively, compared with that of deproteinized SV40 DNA component I.Indimethylsulfoxide gradients, thecomplementary ribonucleic acid (cRNA)
syn-thesized from DNP-I is one-third to one-half the size ofthe cRNA species from DNA-I and DNP-IL. Competition hybridization experiments show that with the E. coli transcriptase only a portion (about one-half) ofthe SV40 genome is
tran-scribed with DNP-I astemplate, whereas mostorallof the genomeis transcribed
with DNP-II as template. The template activity of the deoxynucleoprotein
com-plexeswithahighlyactiveform lI ribonucleicacid polymerase prepared from SV40-infected permissivecells follows similartranscription kinetics. The results indicate
that structural nucleoproteins of SV40 bind nonrandomly to the viral DNA and
effect thetranscriptionofsomesubsetof itssequencesinvitro. The biological significance of
deoxyribonu-cleic acid (DNA)-bound proteins of mamma-lian chromation (10, 19, 23), plant chromatin
(3, 6, 10), and Escherichia coli colicinogenic factor (4, 9) has been extensively investigated. Several lines of evidencethat the
binding
of chro-mosomal proteinsto DNA inactivates transcrip-tionsuggest that these proteins act as regulatorymolecules in gene expression (3, 10, 23). In the
colicinogenic factor system, the protein of the
DNA-protein relaxation complex may act as an endonuclease and open a single-strand nick to
initiateDNA replication (9).
These observations led us toinvestigatethe pos-sible role in the regulation of simian virus 40
(SV40) geneexpressionoftheviralstructural pro-teinsasociatedinthevirion with the viral DNA.
There is increasing evidence for the existence of
nucleoproteinsin SV40andpolyomavirus (2, 11, 15;P. M.FrearsonandL. V.Crawford, inpress).
We show here the template activity of SV40
deoxynucleoprotein complexes in vitro with DNA-dependent ribonucleic acid (RNA) poly-merases ofbacterial and mammalian origin. The
data sLIggest that SV40 may carry with it
struc-tural components thatexert anonrandom limita-tion on transcription so that a particular subset rather than the full complement of viral DNA sequencescanbetranscribed.
NIATERIALS AND METHODS
Cellsandvirus.Themethods forcultivation,
purifi-cation, isotopic labeling, andalkaline degradation of
virus areinprecedingreports (11, 15, 22).
SV40DNA and DNAcomponentI.SV40 DNAwas
extracted from purified virions bythephenol-sodium
dodecyl sulfate (SDS) method (21).SupercoiledSV40
DNA (conponent 1) was separated from SV40open
circles (component II) with ethidium bromide (100-200,ug/ml) andequilibrium densitycentrifugation in
CsCl with an initial density of 1.56 g,'cm3 in the
Spinco T-65 rotor at 43,000rev,'min for 48 hr (22).
The band corresponding to DNA component I was
collected; the ethidium bromide was removed by
repeated extraction with isoamyl alcohol until there
was no color in thewater phase. The DNA solution
was dialyzedagainst
tris(hydroxymethyl)aminometh-ane (Tris)-buffered isotonic saline solution, pH 7.4
(TBS).
Preparation of subviral deoxynucleoprotein cot' 930
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 DEOXYNUCLEOPROTEIN COMPLEXES. IX plexes. Two types ofdeoxynucleoprotein complexes,
DNP-I and DNP-II,consistingof the viral DNA and structural virion polypeptides were isolated from degradedvirus (15).
Purification of DNA-dependent RNA polymerase fromE. coli. DNA-dependent RNApolymerase
con-tainingo-factor wasisolated from E.coli, strainQ13,
by the methodof Burgess(7).Enzyme ofhigh specific
activity (400 units/mgprotein,200units/ml) from low salt followed by high salt-glycerol gradientswas used for thepolymerization assay.
Transcription assay with E. coli DNA-dependent
RNA polymerase (transcriptase). The stock reaction mixture (2.5 X concentration) for the enzyme
con-tained 0.1 M Tris-hydrochloride, pH 7.9, 0.025 M
MgCl2, 0.25 mM
ethylenediaminetetraacetic
acid (EDTA), 0.25 mMdithiothreitol(DTT),and 0.375 mM uridine triphosphate (UTP), guanosine triphosphate(GTP), cytidine triphosphate (CTP), and adenosine triphosphate (ATP) (7). For the assay,0.1 ml of the stockmixture,0.01 mlofE.coli RNApolymerase,0.1 ml of templatecontaining 1.0 ,ug of DNA, 0.01 ml of 3H-UTP
(100,4Ci/ml,
25Ci/mmole), and 0.03 mlof distilledwater tomake thevolume0.25mlwere incu-bated ina37C waterbathfor 15, 30,45, or60 min. The mixtures were precipitated with 0.05 ml100%/-0
trichloroacetic acidcontaining0.2 Msodium
pyrophos-phate, collected on a membrane filter (Bac-T-flex,
Schleicher and Schuell), and washed eight times by filtrationwith5% trichloroacetic acidcontaining0.02 Msodium pyrophosphate.After drying, the filterswere
counted intoluenescintillationfluid.
Synthesis of virus complementaryRNAinvitro. The synthesis of 3H-UTP-labeled viral complementary
RNA (cRNA) wascarried out byslightly modifying
themethod ofWestphaland Dulbecco (31). To 0.25 mCi of3H-UTP (25Ci/mmole) lyophilizedina3-ml
ampoule wereadded0.1 mlof the stockreaction
mix-turewithout coldUTP, 0.03ml of E. coli RNA
poly-merase, 0.1 ml of SV40 DNA component I (100 ,ug/ml), or the equivalent quantity ofnucleoprotein complex, and 20,ulitersofdistilledwater. After incu-bation for 100 min in a 37 C water bath, 20 Ag of
deoxyribonuclease (Worthington)wasaddedtodigest
the DNA (1 hrat 37 C) and stop the reaction. The mixtureswerethentreated with SDSat afinal concen-tration of
1%<7;o
and chromatographed on Sephadex G-50(mediumgrade;bedvolume, 5ml;diameter,0.5cm; flow rate, 0.5 ml/min) in O.1X SSC containing 0.01% SDS. Thefirst peak thatelutedwas collected, extracted three times with chloroform, and dialyzed
against thiee changes of0.1X SSCcontaining 0.01%
SDSfor24hrat4C.Cold viral cRNAwassynthesized
as above except that cold nucleotide triphosphates
wereusedin the reaction mixture.
RNA-DNAhybridizationandcompetitive hybridiza-tion. SV40 DNAcomponentI wastreated with deoxy-ribonuclease (31) and denatured by heating in boiling
water for 15 min and then quicklychilled inanice bath. The denatured DNA(diluted to 0.01,g in 3 ml
of6X SSC) was immobilized on membrane filters
that hadbeen soaked in 6X SSC and thenwashed in 40 ml of 6X SSC.Thefilters weredried inavacuum
at roomtemperature overnight and at 80 C for 2 hr.
The hybridization ofRNA to immobilized
dena-tured DNA was by the method of Gillespie and Spiegelman (13). The DNA filters wereimmersed in scintillation vials in 1.0 ml of 6X SSCcontaining0.05 gg of 3H-cRNAtranscribed from SV40DNA compo-nent I (4X 105 counts/min;specific activity, 8 X 106
countspermin perMg) or, in thecompetition experi-ments, together with graded amounts of cold cRNA fromDNP-I, DNP-II, or DNA-I.
Two variations of the competitive-hybridization technique were employed. (i) After the first hybridiza-tion with 0.05 Mug of 3H-cRNA of DNA-I and increas-ing amounts of cold competing cRNA, the filters containing denatured viral DNAwererinsed with four changes of 2X SSC and washed by suction filtration
oneachside with50ml of 2X SSC. The filters were thentreatedat 37Cfor1 hrwith pancreatic ribonu-clease(20ug/ml) thathad beenpreheatedat80 C for
15mintoinactivate contaminating deoxyribonuclease (13, 31). Thefilterswereagain rinsed four times with 2X SSC and washed byfiltration (17). The 3H-cRNA from DNA-I (0.05 Mg) was then added for a second hybridization in the presence of 1.0 mg of yeast RNA which wasreprecipitated with alcohol and0.1%OSDS. The filters were then subjected to the same washing andribonucleasedigestion procedure. Finally the
fil-terswererewashedoneachsidewith 2X SSC by
suc-tion filtration and were dried, and the radioactivity
wascounted.
(ii) After hybridization with amounts of hot and cold cRNA and denatured DNA comparable to those in(i),thefilterswerewashed with fourchanges of 2X SSC and heated ina70 Cwaterbath for 10 minwith 6X SSCto removeremainingnonspecifically
hybrid-izing materials. The filters were rinsed with four
changesof2X SSC andsubjectedtothesame ribonu-cleasetreatmentand finalwashing proceduresas in (i) withoutasecondhybridization.
Preparation of mammalian DNA-dependent RNA
polymerasefrom SV40-infected monkey kidney cells.
Confluentmonkeykidneycells (MA-134) growing in 30 roller bottles (surface area, 1,000 cm2) were in-fected with SV40 (50 plaque-forming units/cell).
Twenty-four hours later, the cells were rinsed twice with ice-cold Hanks solution (BSS), removed from thebottles with glassbeadsorrubberpolicemen, col-lected bycentrifugation, and washed twice with cold BSS.
The purification of DNA-dependent
RNA-poly-meraseessentiallyfollowed the method ofB. Sugden andW.Keller(personalcommu/iicatioti), inwhich the enzymes were eluted from the cells and purified
through ammonium sulfate fractionation,
diethyl-aminoethyl (DEAE)-cellulose chromatography, and
centrifugation in a low salt-glycerol gradient. RNA
polymeraseform II (RNAP-II) (25, 26) was purified byfurthercentrifugationina5to
20%,-
sucrosegradi-entinahigh-saltbuffer.
Invitro mammalianDNA-dependent RNA polymer-asetranscription assay. The reaction mixture is modi-fied from the formula of Sugden and Sambrook (28). The stock reactionmixture (2.5X concentration) for the assaycontained0.2MTris-hydrochloride, pH 7.9, 0.01 M MgCl2, 0.005MMnCI2,0.25 mM DTT, 0.08 M
KCl, 1.25 mgofbovine serumalbumin per ml, 0.25
931
VOL.9, 1972
on November 10, 2019 by guest
http://jvi.asm.org/
HUANG, NONOYAMA, AND PAGANO
mM EDTA, 0.125 M (NH4)2SO4, 10 ,uCi of 3H-UTP perml, and 0.6 mmATP,GTP,and CTP.
The standard assay system consisted of 0.1 ml of stockreaction mixture, 0.1 ml of template containing 4,g of DNA, 20 ,uliters ofRNAP-1L from SV40-infected MA-134 cells, and 2 mm adenosine-3 ',5'-monophosphoric acid (cAMP). The rest of the proce-dure followed the E. coli system as described above.
RESULTS
Subviral deoxynucleoprotein complexes. Purified SV40virionscanbedissociatedin alkalinebuffers
atpH 10.5 into asoluble proteinand a
deoxynu-cleoprotein complex containing the viral DNA and the fourminor ofthesixstructural viral
poly-peptides (15), followedbyvelocitysedimentation
insucrosegradients. DNP-I canin turn be
disso-ciated into solubleprotein and another
deoxynu-cleoprotein complex (DNP-II) by equilibrium density centrifugation in CsCl. DNP-IL contains mainly the viral DNA and a small remnant of
viral protein, apparently chiefly VP3, tightly boundtotheDNA anddifficult todetect (15).
Transcription in vitro of deoxynucleoprotein complexes with E. coli DNA-dependent RNA polymerase. We examined the kinetics of
tran-scriptionofDNP-I, DNP-II, and SV40 DNA com-ponent I (supercoiled DNA). The concentration of DNA was assayed by the diphenylamine
method (8) and verifiedby extinctioncoefficients and theratio of absorbanciesat260 and 280nm
(30).
DNP-I,
DNP-II, and SV40DNA-I, diluted withTBS tocontain equalamounts of DNA (1.0,g) wereusedastemplates.Asindicated inFig.1, the rateof
incorporation
of soluble 3H-UTP into acid-precipitable materials with DNP-I (Fig. la,fractions
10-12)
astemplate
is only about30%0
of the rate obtained with DNA component I.
With DNP-IItherate is about
80%,o.
Theresultsshowclearly thatDNA componentI andDNP-I
have a different template activity. Removal of
protein from DNP-I to form DNP-II increased thetemplate
activity,
and the difference inactivity
between DNA-Iand DNP-IIis not so obvious.
Themorerapidlysedimenting material (Fig.
la,
fractions 15-17) in the gradient used to isolate
DNP-I contained on electrophoresis much more viral coat protein (VP1 and 2) than did
DNP-I;
in otherwords, the degradation of the virus was
less complete and the template activity of this material waslower.
Sedimentation properties of cRNA species syn-thesized in vitro. After 30min of
synthesis,
SDS was addedto thereaction mixtures to a concen-tration of0.1%tostop thereaction. The mixturesimmediately were sedimented in a
sucrose-dimethylsulfoxide
gradient
to avoiddegradation
of the RNA.
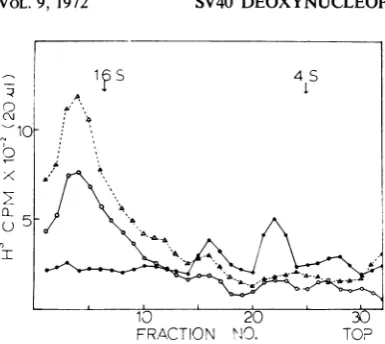
Figure
2 shows the sedimentation.4 yuulieL,.
DNP-I DNP-IL
DNA-I
15 10 5tOp0 5 1
DP0to
-.0X4
I Fc10 12 3
12
o
P~~~~~~~~~c
16 -1715
30
~
45603
[image:3.495.268.456.79.318.2]TIME VINUTES
FIG. 1. Kinieticsof ribonlcleoside triphosphate
inicor-porationt
withSV40 DNA-I, DNP-I, or DNP-II asthetemplates. DNP-I was
obtainied
bydegradationz
ofthe virusinanalkalinebuffer,pH 10.5,for 15hr,followedby
sedimenitationi
in a sucrosegradienzt.
DNP-Il wasproducedby dissociation of DNP-I
durinig
equilibriumdentsity cenitrifugationl
in CsCl (seereferenice
16 fordetails.) The stock
reactionz
mixture(0.1 ml),0.1mlofthe different templates niormalized to 1.0
Mg
of DNA, 0.01 mlofE. colitrantscriptase,
0.01mlof3H-UTP (100yuCi/ml),
and 0.03mlof distilledwaterwere mixedanld
incubated at 37 C. After different time intervals, the incorporated material was precipitated with ice-cold trichloroacetic acid
anid
collectedon membranefilters.Tlhe
amounlt
of3H-UTP incorporated with DNA-Iastemplate, (0 *);DNP-IIas template
(0
---0),DNP-I
(fractioni
10-12) astemnplate
(O--O), frac-tioII 15 (A -A); and 16 anid 17 (A A) astemplates.
profiles ofthe cRNAspeciessynthesizedinvitro.
The RNA transcribed from DNP-I sediments
more slowly than the cRNA from DNA-I and
DNP-II.
The cRNA transcribed from DNP-IIshows no significant difference from that of
DNA-I.
The peaks of cRNA from DNP-I arelocatedatapproximatelythe6and 10S
positions;
the cRNA species of DNP-I and DNA-I are at 18S.
Other preparations were analyzed by
velocity
sedimentation in 0.1 % SDS-sucrose
gradients
after chromatography of the cRNA
species
through Sephadex G-50; thecRNA fromDNP-I sedimented in a broad peak of about 6S,
932 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 DEOXYNUCLEOPROTEIN COMPLEXES. II
FRACTION N1O. TOP30
FIG. 2. Sedimentation analysis in a dimethyl sul-foxide (DMSO) gradienit ofthe cRNA species
synthe-sizedin vitro withDNP-I, DNP-II, and DNA compo-inent I astemplates. The3H-cRNA speciessynthesized for 30 minfrom DNA-I, DNP-I, andDNP-IH were
sedimentedin a2.5 to10% sucrose gradientpreparedin
100% DMSOmodifiedfromTonegawaetal.(29). After
24 hrof centrifugation at 45,000 rev/minintheSpinco SW 50.1 rotor, 20
piliters
ofsample fromeachfractionof thegradient were driedonglass filtersandwashed byfiltrationwith5%trichloroacetic acid with 0.02M 50-diumpyrophosphate, dried, andcoutnted. The
sedimen-tation isfrom righttoleft: 23, 16, and4S RNAfrom
E.coliwereusedasmarkersin aseparate tube.
Distri-bution ofDNA-I cRNA (AL); DNP-I cRNA (@);
DNP-11 cRNA (0).
compared with 16 to 18S for cRNAspeciesfrom
DNP-II andDNA I.
Competition study between the cRNA
species
of DNA component I,DNP-I,andDNP-II. Toexam-ine whether the binding of the DNA-associated viralstructural
proteins (VP3,
4, 5,6)
totheviralDNAresults in specific or randomlimitation of
transcription,
competitive hybridization
studiesweredone.
We first determined the amount of 3H-cRNA
ofDNA-IneededtosaturatetheamountofDNA usedinthecompetitive
hybridization experiments.
Theresults, shown in Fig. 3, indicated that0.05
,g
of 3H-cRNAproduced
close to maximalhybridization; application ofthreetimesasmuch cRNA(0.15Mg) caused hybridization ofonly 24%
more counts.
WecalculatedtheamountofDNAimmobilized
in membrane filtersbylabelingit with
'4C-thymi-dine.Thespecificactivityof'4C-SV40DNA-I was
7,333 counts per min per
Mg.
Theaverage counts of DNA usedforhybridizationwere72countsperminperfilter. Thepercentage of DNA remaining immobilized on thefilters was 82.6%
(±
3.3%) ofthe DNA applied; after22 hrof competition hybridization there was no significant loss of DNAfrom thefilters.Figure4 shows theresults of two competition studies. Cold eRNA synthesized in vitro from
DNP-I, DNP-II, or DNA-I was used in compe-tition against3H-cRNAof DNA componentI. In the presenceof 0.5
,ug
of cold cRNA fromDNP-I,
when theratio of cold cRNA to 3H-cRNA was 10, about half (51.2%) of the control counts remained on the filters (Fig. 4A). With cold cRNA from DNP-II or DNA-I, the counts
re-maining bound to thefilters were about
12%o
ofthe control counts. The results obtained with
method (ii) (Fig. 4B) with a single simultaneous hybridization confirmed the results showninFig.
4A. These dataindicate that with DNP-Ias
tem-plate only half of the SV40 genome sequences were transcribed with E. coli transcriptase; in
contrast with DNP-II as template, most of the DNA was transcribed. These data support the
hypothesis that theproteins ofDNP-Iarebound
nonrandomly to theviral DNA and that bound areas orblockedinitiation sites (E. coli
transcript-ase) allow the transcription ofonly about half of the SV40 genome.
In the converse experiment, cold cRNA from DNA-I in competition with labeled cRNA from
DNP-I prevented thehybridizationof morethan 95%of thecountshybridizing in the control filters
toDNA-I intheabsenceofcold cRNA.
Purffication
andtemplate specificityofRNAP-IIfrom SV40-infected cells. RNA polymerase form
I (nucleolar enzyme) and formII (nucleoplasmic enzyme) (25, 26) obtained after DEAE-cellulose column elutionwerepooled and centrifugedin a
2
x
O
0I
0.05
3.10 0.15H3 c-RNA
(1ig)
FIG. 3. Determination of thesaturation levelof
3H-cRNA synthesizedfrom SV40DNA-IforSV40DNA. Aconstanit
amoun1t
ofdenaturedSV40 DNA (0.01plg)
wasimmobilizedonntitrocellulosemembranefilters. Thle
filters wereimmersedin6X SSCwithgradedamounts
of3H-cRNA (8 X 106 countsperminper,ug)
syntthe-sizedfrom SV40 DNA-I, 1.0 mg ofyeast RNA, and 0.1% SDS. Hybridizationwascarriedoutfor22hras
in method ii (see text). With 0.05
pAg
of3H-cRNA inthe reaction, thenumberofcounts perminute that
hy-bridizedisclosetotheplatealuofthe curve.
10---7
0
v
933 VOL.9, 1972
1
n
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.495.48.241.60.232.2] [image:4.495.256.443.426.561.2]HUANG, NONOYAMA, AND PAGANO
100
A
60-er)
40
-20
<=:
o
--g
°
0.1
0.2
0.3
0.4
C 5
z
620-00
0.1
0.2
0.3
0.4
0.5
0
0.1
0.2
0.3
0.4
0.5
cold
c-RNA
(Aug)
FIG. 4. Competitive hybridizationt ofcRNA species
synthesizedin vitrofromDNA-I,DNP-I, andDNP-II.
Thefilters withconistantamountsofimmobilized DNA wereimmersedin1mlof 6X SSC containing 3H-cRNA (0.05jig; specific activity, 8 X 106 countsperminper I jig) synithesized from SV40 DNA-I, graded amounlts
ofcold cRNAfrom DNP-I, DNP-II, orDNA-I, Img of yeast RNA, and0.1% SDS. The hiybridizationi ex-periments were performed as described in Materials and Methods (i) and (ii) (see text). A, After thefirst
competitive hybridization, thefilters weresubjectedto
washing andribonucleasedigestiont;0.05 jAgof3H-cRNA ofDNA-Iwasthentaddedforthe secondhybridizationi.
B, After a single competitive hybridization thefilters werewashed and heatedat70Cfor10mm inplaceof
the seconidhybridization. The counits of3H-cRNA
hy-bridizedto immobilized SV40 DNA in the absenceof
cold competing cRNA wereusedas the 100%
hybrid-ization control; inexperimenits A anidB, the control countswere13,516anld13,402 counts/minl,respectively.
Thecounts onfilters withoutSV40 DNA were58 and 56counts/min in A and B, respectively. ColdcRNA synthesizedfrom DNP-J (0);coldcRNAsynzthesized
fromDNP-II(A);coldcRNAsynithesizedfromDNA-I
(0i)
low-salt-glycerol
gradient.
Twocomponents wereresolved (Fig.
5A);
RNAP-Imigrates
asadimer,
and RNAP-IIasa monomer (J. Sambrook, per-sonal communication). With denatured calf
thy-mus DNA as the
template,
theactivity
ofRNAP-IIfromSV40-infectedcellsisatleast four timesasgreatasthatof RNAP-I.
After the fractionswere sedimented ina
high-salt gradient,onedistinct RNAP-II peak appeared
(Fig. SB). Theenzymeobtained from this gradient was free from detectable host cell DNA or
con-tamination with viral DNA. Table 1 shows the template specificity of RNAP-II. The results indi-cate that in vitro RNAP-II prefers denatured DNA as a template. IfcAMP was added tothe reaction mixture, there was a slight increase in 3H-UTP incorporation when native SV40 DNA componentIwasthetemplate.
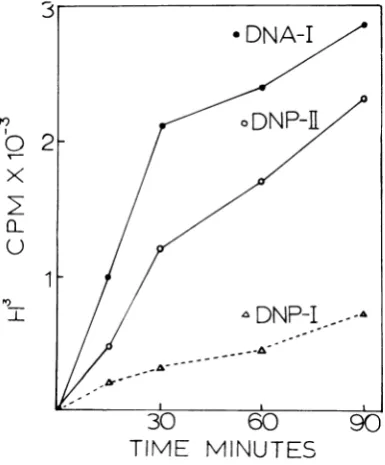
Transcription of deoxynucleoprotein complexes in vitro with mammalian RNA polymerase. RNAP-II (16, 25, 26) from permissive cells
infectedwith SV40wasusedforin vitro
transcrip-tion assays.DNA-I, DNP-I, or DNP-II
contain-ing equal amounts of DNA (4
,ug/0.1
ml) wereused as the templates. The rates of transcription with DNP-I and DNP-IIare28 and82%,
respec-tively, compared withthatof DNA componentI
(Fig. 6). The results with mammalian enzyme
from infected cellsarereproducible andare
com-parable to the results with the E. coli enzyme.
Withboth types ofenzymes, there was a
signifi-cantreduction in transcription when DNP-Iwas
the template and a slight reduction when
DNP-IIwas thetemplate.
DISCUSSION
Deoxynucleoprotein complex I isolated from alkali-degraded SV40isactive inDNA-dependent RNA synthesis with the transcriptase fromE.coli
we
II2
-1II
/ 1
.'I
I
I
.-
I...
_ r)
J . '\
e)OffO><>t)d°
X- * *sx
1VS 1 5 .- , 15
T
[image:5.495.60.249.75.344.2] [image:5.495.265.452.419.559.2]F T. T?-L
...
FIG. 5. Purificationandsedimentationofmammalian RNApolymerases. Polymerase forms I andIIeluted
from a DEAE-cellulose column were pooled,
precipi-tated, anld dissolved in buffer. A, The enzymes were
centrifugedinalow-salt20to50%glycerol gradientat
26,000rev/mimlinaSpinco SW27rotorfor24 hrat4 C. B, RNAP-II was isolatedfrom fractionis 9, 10,and 11
ofa low-salt-glycerol gradient, precipitated, dialyzed,
dissolvedinahigh-salt buffer,andthencentrifugedina
high-salt5to20%Osucrosegradienttinthe SW 27rotor at26,000 rev/minfor30 hr.
934
J.VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
SV40 DEOXYNUCLEOPROTEIN COMPLEXES. IX TABLE 1. Templatespecificity of mammalian
DNA-dependent RNA polymerase (RNAP-II) iso-lated from S V40-infected MA-134 monkey
kidney cellsa
Templateb
Amt of _
DNA NativeSV40
used as Denatured Native DNAtiven I
tem- calfthymus calfthymus mponent I
plate DNA DNA
(pg) (without (without (with (without
cAMP) cAMP) CAMp)d cAMP)d
30 15,689c 1,630
10
11),733
2,9494 16,402 2,054 3,954 3,396
1 13,664 975 1,388 716 a Method of B. Sugden andW. Keller (personal communication). Abbreviations: DNA,
deoxy-ribonucleic acid; RNA, deoxy-ribonucleic acid; SV40,
simian virus 40; cAMP, adenosine-3',
5'-mono-phosphoric acid. Enzyme control, 143 counts/
min; 30 gg of denatured calfthymus DNA con-trol, 143 counts/min; 30jugofnativecalfthymus
DNA, 72 counts/min; 4Ag of SV40 DNA 1, 132 counts/min.
IThe amount of RNAP-I in all assays is 20
,lAiters.
c3H-Uridine triphosphate incorporation,
ex-pressed in counts per minute.
d Finalconcentration of2.0mmoles/ml.
andanRNA
polymerase
from mammalian cells.However, the
template
activity
of DNP-Iismuchlessthanthatofan
equal
amountofdeproteinized
SV40 DNA component I. The decrease appears tobe due toviral
proteins
boundtotheDNA;
thetemplate
activity
increases after the removal ofmost ofthe
protein
from DNP-Iby
CsClcentrif-ugation. DNA-cRNA
competition hybridization
experiments
show that the cRNAsynthesized
invitro with DNP-I as
template hybridized
withonly
a fraction of the viral DNA transcribable with the E. coli enzyme. This indicates that the viralprotein
preventstranscription
ofasubset of theviral DNA sequences either bybinding
with itorblockingE.colitranscription
initiation sitesorboth. WithDNP-IIasthe
template,
transcrip-tionwas not asreduced, andwecouldnot assess the small differences in the transcribed RNA
species in the
competition
hybridization
experi-ments.
Thereare technicalpoints worthconsideration.
Lucas andGinsberg (17)forhybridization
inhibi-tion testsappliedtoimmobilized viral DNA cold messenger RNA (mRNA) from infected cells, treated withribonuclease and washedextensively,
and thenapplied3H-cRNA of viral DNA. We first assessed the amount of 3H-cRNA synthesized
fromDNA-I needed to saturate theimmobilized
viral DNA and then used two different methods toascertainboth the existence of differentcRNA
species astranscribed fromDNP-I, DNP-II,and DNA-I and also the percentage of the genome
transcribed in eachcRNA.
Inthe first method weapplied cold cRNA from DNP-I, DNP-II, or DNA-I together with 3H-cRNA fromDNA-I, treated it with ribonuclease, washeditextensively,andthenapplied additional
3H-cRNA from DNA-I so as toinsuresaturation of any remaining unbound sites. In the second method, we simply applied a mixture of cold cRNA and 3H-cRNA from the appropriate
sources and in varying ratios simultaneously. In both cases, up to a10-foldexcessofthe cold
com-peting cRNAwasused;wepresumedthat thiswas a saturating amount as aplateauwasreachedin the competitive effect (Fig. 4). The results were
virtually identical. For analysis of these in vitro products,where theprecise quantities of the reac-tants are known, we suggest that simultaneous
application of both kinds of cRNA allows for a
quantitative competitive effect, whereas
applica-tion of the cRNA species sequentially does not
allow competition for binding to sites on the
DNA, but onlyindicatesthe final percentages of
the genome to which the different RNA species
bind. With the sequential
technique,
there is a3
*DNA-I
b
2-~N-T_
U
~~~~~DNP-1
TIME
MINUTES
FIG. 6. Kinetics oftranscription in vitro ofSV40
DNA-I,DNP-I, andDNP-IIwith a mammalian RNA
polymerase. The mammalianRNApolymerase II assay isdescribedinthetext. Theamountof3H-UTP
incor-porated withDNA-I (-),DNP-I (A),orDNP-IIas
template (0).
VOL. 9, 1972 935
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.495.250.443.376.609.2]HUANG, NONOYAMA, AND PAGANO
dangerthateven aminor contaminationwith, for
example,
"late" RNAsequences in thepredomi-nately "early" cold RNA appliedfirst will mask
real differencesbased on the relativepopulations oftwokinds ofRNAspecies.
When chromatin from animal or plant tissues
is used as a template for synthesizing cRNA in
vitro, only a part of theDNAis transcribed (6,
10, 19). Chromosomal proteins persisting in a
complexwiththe DNA make specificportions of
it unavailable for transcription and account for
this effect. SV40DNA-bound protein may play a
similar role. However, the existence of viral
deoxynucleoprotein complexes and their
func-tional significance in vivoarestillopenquestions.
Tovalidate this concept, it would be necessary to find in permissive infected cells virus-specific
mRNA corresponding to thein vitrotranscripts ofDNP-I and DNP-II. If in lytic infection the
early viralgenes areexpressed beforeviralDNA
replication and the other late genes after DNA
replication (20,24, 27,29), thentheblockto
tran-scription
of lategenes islifted so that the entiregenomeis ultimately expressed in lytic systems. In contrast, in transformed systems such as SV 3T3cells, theentiregenomeispresent but the late
functions arenever expressed. Sauer andKidwai (27) haveshownthat thesections of viralgenome
transcribed in SV40-transformedGMK (18) cells
are twice as large as in SV40-transformed 3T3
cells. Therefore, Oda and Dulbecco (20) and Sauerand Kidwai(27) invoketwotranscriptional control
mechanisms:
theblocktotranscription of lategenesequences thatoccursinpermissive
cells beforeviralDNAreplication
isduetosome prop-erty of the virus, whereas in thetwo systems of transformed cells cited thepersistent
block totranscription ofmostlate genes is based on
cel-lularproperties.
A deoxynucleoprotein complex similar to
DNP-I might be released in vivo after
un-coating ofthe virus. Then, after removal ofthe
capsid,
thenucleoprotein
byitselforincoopera-tion with some other factor
might
repress lategenefunctions and
permit
onlytheexpression
ofearly genes. After this, the internal viralprotein is removed from the viral DNA, perhaps by an
early gene product. Viral DNA
replication
andlate gene expression are then favored. There is another
possibility. Newly synthesized
viralpro-teinmaybindtonewly
synthesized
viralDNA attheearlygenesitessothat thelate genes involved withvirus maturation continuetobefavored. This
possibilityisunlikely, however,
if,
as Alonietal. (1) showed, early mRNA is present throughouttheinfectiouscycle.
The mammalianRNA polymeraseusedin this
work
(RNAP-II)
was highly active,especially
when denaturedDNA was used as template, its
specific activity being as high or higher than the E. coli polymerase. A template preference for
native DNA may depend on recovery of some
factor lost during DEAE-cellulose chromatog-raphy(B.Sugden and W.Keller,personal commu-nication). Since the binding sites ofE.coli enzyme
and mammalian enzyme may not be identical,
with three to nine sites for the bacterial enzyme butperhapsonly one for themammalian enzyme
(14),
the useofRNA polymeraseofmammalianorigin may be crucial. Viral nucleoprotein may
onlyblock a specific initiationsite for
transcrip-tion, which could
still
allow theE.colipolymeraseto initiate nonorthodox transcription and yield cRNAthat mightapproximatebut not match the
corresponding mRNA ininfected
cells.
The highactivity of the newly available mammalian polymerases
will
allow synthesis of adequate3H-cRNA
fromDNP-I
for competitivehybridi-zation experiments with "early" mRNA from
SV40-infected cells.
ACKNOWLEDGMENTS
We thank Norman S.-T. Chen for assistance and Joseph
SambrookandRobertWeinberg for helpful criticism. B. Sugden
andW.Keller generously gave details for the preparation of the
mammalian RNA polymerases.
This workwassupportedby a grant to theUniversityof North
Carolinafrom theAmericanCancerSociety(IN15-LInstitutional
Grant),by Public HealthServicegrant 5SO0 FR05406from the
Division of Research Facilities and Resources, and by grant
VC-48 from the American Cancer Society. Joseph S. Pagano holds Public Health Service Research Career Development Award 5 K04 Al 13516from theNational Institute of Allergy
andInfectious Diseases.
LITERATURE CITED
1. Aloni, Y.. E. Winocour, and L. Sachs. 1968.
Characteriza-tionof thesimian virus40-specificRNAinvirus-yielding
and transformed cells. J. Mol. Biol. 31:415-429.
2. Barban, S., and R. S. Goor. 1971. Structural proteins of
simian virus 40. J. Virol. 7:198-203.
3. Bekhor, I.,G. M. Kung, and J. Bonner. 1969.
Sequence-specific interaction of DNA and chromosomie protein.
J. Mol.Biol.39:351-364.
4. Blair,D.G.,D. B.Clewell,D.J.Sheratt, andD. R.Helsinki.
1971. Strand-specific supercoiled DNA-protein relaxation
complex: comparison of the complexes of bacterial
plas-mids Col Ei and Col E2. Proc. Nat. Acad. Sci. U.S.A.
68:210-214.
5. Bonner, J.,andR.C.Huang.1963.Propertiesof chromosomal
nucleohistone. J. Mol. Biol. 6:169-174.
6. Bonner,J.,R.C.Huang,andR.V.Gilden. 1963.
Chromo-somally directedprotein synthesis. Proc. Nat. Acad. Sci.
U.S.A.50:893-900.
7. Burgess, R.R.1969.Anewmethodfor thelargescale
puri-fication of E. colideoxyribonucleicacid-dependent
ribo-nucleic acidpolymerase. J. Biol. Chem. 244:6160-6167. 8. Burton,K.1956.Astudyof theconditions andmeclhanismof
the diphenylamine reaction for the colorimetric
estima-tion ofdeoxyribonucleicacid. Biochem. J. 62:315-323. 9. Clewell, D. B.,and D. R. Helsinki. 1969. SuLpercoiled
cir-cular DNA-protein complex in Escherichia coli:
purifica-tion and inducedconversiontoanopencircular DNA form. Proc.Nat. Acad. Sci.U.S.A. 62:1159-1166.
936 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 DEOXYNUCLEOPROTEIN COMPLEXES. II
10. Dahmus,M.E.,and J. Bonner. 1970. Nucleoproteinin
regu-lation ofgenefunction. Fed. Proc. 29:1255-1260.
11.Estes, M. K., E.-S. Huang,andJ. S. Pagano. 1971. Struc-turalpolypeptidesofsimian virus40.J. Virol. 7:635-641. 12. Frenster, J. H.,V. G. Allfrey, and A. E.Mirsky. 1963. Re-pressed and active chromatin isolated from interphase lymphocytes. Proc. Nat. Acad.Sci. U.S.A. 50:1026-1032. 13. Gillespie,D., and S. Spiegelman. 1965.Aquantitativeassay
for DNA-RNA hybrids with DNA immobilized on a
membrane. J. Mol. Biol. 12:829-842.
14. Herzberg,M., andE.Winocour. 1970. Simianvirus40
deoxy-ribonucleic acid transcription in vitro: binding and
tran-scriptionpatternwithamammalianribonucleicacid
poly-merase. J. Virol. 6:667-676.
15. Huang,E.-S.,M.K. Estes, andJ.S.Pagano. 1972.Structure
and function of the polypeptides in simianvirus 40. 1.
Existence of subviral deoxynucleoprotein complexes. J.
Virol.9:923-929.
16. Keller, W., andR. Goor. 1970. Mammalian RNA
poly-merase:structural and functional properties. ColdSpring
HarborSymp. Quant.Biol. 35:671-680.
17. Lucas,J. L.,and H. S. Ginsberg. 1971.Synthesis of
virus-specific ribonucleicacid inKBcells infected withtype2
adenovirus.J. Virol.8:203-213.
18. Koprowski, H.,F. C.Jensen,and Z.Steplewski. 1967.
Ac-tivation ofproduction ofinfectioustumorvirusSV40in
heterokaryon cultures. Proc. Nat. Acad. Sci. U.S.A. 58:
127-133.
19. Marushige, K.,andJ.Bonner. 1966.Templateproperties of
liver chromatin. J. Mol. Biol. 15:160-174.
20. Oda, K.,and R. Dulbecco. 1968.Regulationoftranscription
ofSV40DNA inproductivelyinfected and intransformed cells. Proc. Nat. Acad. Sci. U.S.A. 60:525-532.
21. Pagano, J. S. 1969. Assay ofinfectious deoxyribonucleic
acid, p. 184-197. In K. Habel and N. Salzman (ed),
Basic techniques in virology. Academic Press Inc., New
York.
22. Pagano, J. S., and C. Hutchison III. 1971. Small circular
viral DNA;preparationandanalysis ofSV40 andOX174
DNA,p. 79-123. In H. Koprowski and K. Maramorosch
(ed.), Methodsin virology, vol. 5. Academic Press Inc.,
New York.
23. Paul,J.,and R.S. Gilmour. 1968. Organ-specific restriction
oftranscription in mammalian chromatin. J. Mol. Biol.
34:305-316.
24. Rapp,F.,J.L.Melnick, andT.Kitahara. 1965.Tumor and virus antigens ofSimian Virus40:differentialinhibition of synthesis by cytosine arabinoside. Science 147:625-627.
25. Roeder, R. G.,andW. J. Rutter. 1969. Multiple forms of
DNA-dependent RNApolymerasein eukarytic organisms.
Nature (London)224:234-237.
26. Roeder, R. G., andW. J.Rutter.1970. Specificnucleolar and
nucleoplasmic RNA polymerases. Proc. Nat. Acad. Sci.
U.S.A.65:675-682.
27. Sauer,G.,and J. R.Kidwai. 1968.The transcriptionofthe
SV40 genome in productively infected and transformed
cells. Proc. Nat. Acad. Sci. U.S.A. 61:1256-1263.
28. Sugden, B., and J. Sambrook. 1970. RNApolymerase from
HeLa cells. ColdSpring HarborSymp. Quant. Biol. 35:
663-670.
29. Tonegawa,S.,G.Walter, A.Bernardini,and R.Dulbecco.
1970. Transcription of the SV40genomein transformed
cells andduring lyticinfection. ColdSpringHarborSymp. Quant.Biol. 35:823-831.
30. Warburg,O.,and W.Christian. 1942. Isolierungund
kris-tallisation das Garungsferments enolase. Biochem. Z. 310:384-421.
31.Westphal, H., and R.Dulbecco. 1968. ViralDNA in
poly-oma- andSV40-transformed cell lines Proc. Nat. Acad,
Sci. 59:1158-1165.
VOL. 9, 1972