Copyright©1970 American Societv for Microbiology Printedin U.S.A.
Membrane Binding
of
Input
Arbovirus Ribonucleic
Acid: Effect
of
Interferon
orCycloheximide
ROBERT M. FRIEDMAN AND T. SREEVALSAN
Laboratory of Pathology, NationalCancerInstitute,NationalInstitutesofHealth,
Bethesda, Maryland 20014,and Department of Microbiology, SchoolofMedicine andDentistry,Georgetown University, Washington,D.C.20007
Received for publication 9 March 1970
By 1hr afterinfection, 36%of input Semliki Forest virus ribonucleic acid(RNA) which was cellassociatedwas found in amembrane structure. This structure had
many similarities to the membrane-associated replication complex (MRC) which had previously been identified in arbovirus infections. Interferontreatment didnot
affect theassociation of viral RNA with the MRCstructure,butcycloheximide treat-mentinhibited it.
In a previously published report, the fate of
input SemlikiForestvirus (SFV) ribonucleic acid
(RNA) wasstudied. Under normalconditions, a
portion ofthe42Svirion RNA (7) was, by 1 hr
afterinfectionofchick cells, alteredto 30 and 20S
viral RNA forms. The latter was ribonuclease
resistant. In cells treated with interferon before
infection, or with cycloheximide or puromycin
during
infection,
viral RNA probably wasun-coated,asevidencedby the formationof26S viral
RNA,aformneverfound in the virion, but little
or no 20S
ribonuclease-resistant
RNA was formed. It wasconcluded thatallthreetreatmentswere similar in that they inhibited the
transcrip-tion ofinput viral RNA, thereby preventing the
formation of an RNA complementary tothe
in-put strand (6).
The results of a more detailed study are
re-portedinthispaper. The
input
SFVRNArapidlybecame associated with a membranous
cyto-plasmicstructure which hadmany of the
proper-ties of a replication complex, a structure which hasbeen
previously
described inSindbisvirusandpoliovirus infections (9, 15). The replication
com-plex was membrane
associated,
as was that ofSindbis virus
(15).
A difference wasnoted,how-ever, in the effects of treatment with
cyclohexi-mide during the course of infection or with interferon before infection. In
cycloheximide-treated cells, entry ofviral RNA into the
mem-brane-associatedreplication complex (MRC)was
decreased, whereas in interferon-treated cells no
effectonthisstepwasnoted.
MATERIALS AND METHODS
Viruses. Pools of SFV and chick cell monolayer
cultures were prepared by previously described
methods (8). An SFVpoolwasprepared in the
pres-enceof 10,uCi/mleachoftritiatedadenosine (19Ci/
mmole) and tritiated uridine (20 Ci/mmole). The viruswas partially purified bythe methodofCheng
(1) with theaddition ofaterminal bandingina15to 60% sucrosedensity gradient.Radioactivitywas con-centratedintwofractions at adensity of1.20g/cm3. Infectivity [22 X 10"1 plaque-forming units (PFU)/ ml] washighest in these fractions. Theinfectivityto
radioactivity ratiowas2.4 X 10' PFU per count per min.Allreportedresultswereconfirmedatleastonce
with another pool ofpurified 3H-SFV and, in most
cases,withapool ofthecloselyrelatedSindbis virus. Infection procedure.Monolayers of chickcells
con-tainingabout 2.5 X 107cellswereincubatedat37C
for1 hrwith 0.5
;tg
ofactinomycinD perml and thenfor 1 hrat4 Cwith 0.1 mlof3H-SFVat avirus:cell multiplicity of2:1. After this period, 5 mlofwarm
Eagle'smediumwith10%fetalcalfserum wasadded
for1hrat37C.
CeU fractionation. Theprocedureemployed forcell
fractionation was based on previously described methods (15). After 1 hr at 37 C, the infected cells
were washed five times with iced 0.85% NaCl and
scraped intoasmallvolumeofthesaline.After
sedi-mentation at800 X gin thecold,thecellswere
re-suspended in 1 ml of 0.01 M tris(hydroxymethyl)-aminomethane (Tris), pH 7.2, and 0.001 M
ethylene-diaminetetraacetic acid (EDTA) andallowedtoswell
for30minbeforehomogenizationinaDounce
homog-enizer. After homogenization, the cell extract was
centrifuged at 800 X g for 5 min, the fluidwas
re-moved, andthesedimentwasresuspendedin 1 mlof Tris-EDTA and again sedimented. The supematant fluidswere combined and sedimentedat 10,000 X g for 15min. The supernatant from this sedimentation
was the postmitochondrial fraction. The pellet was
gently resuspended in 1.0mlofTris-EDTA, and the differential sedimentation procedure was repeated.
The postmitochondrial supematants werecombined,
and thefinalpelletwasresuspendedin0.5mlof Tris-169
on November 11, 2019 by guest
http://jvi.asm.org/
EDTA. Thesuspensionswerethenlayeredovera
dis-continuous sucrose gradientin Tris-EDTA prepared
as follows: 0.4 ml of 50% sucrose, 0.4 ml of 45% sucrose,0.4 ml of 42.5% sucrose, 1.1 ml of 40%
sucrose, 1.1 mlof25% sucrose, and 1.1 mlof15%
sucrose. Sedimentation in the SW-50 head wasfor 30 min at 131,000 X g. The gradients were collected,
and radioactivity was estimated by previously
de-scribed methods (7).
RNA was extractedfromthe mitochondrial frac-tion by an sodium dodecyl sulfate (SDS)-phenol
methodpreviously described(7).Itwassedimentedin a6 to 30% sucrosegradient for 1 hrat300,000 X g
inanSW-65 rotor.
Interferon. Partially purified interferon was pre-pared anddonatedbyKarl Fantes (3). The
prepara-tion contained 10,000 international units of chick interferonand 179 ,ugofproteinper ml.
Incorporation of radioactivity. Incorporation of radioactivityfrom viral RNA intocell fractionswas
estimated by precipitation and washing with 2.5%
perchloric acid. The washed precipitates were then
solubilized with 0.33 N NaOH and counted. Protein
contentwasestimated bythemethodofLowryetal.
(10).
Sarkosylfractionation of the mitochondrial fraction. ThemethodofTremblayetal. (16) wasemployedto
isolate a cell membrane-viral RNA complex as
follows:2.5mlof15%sucrosein 0.01 M Tris(pH 7.2),
0.1MKCl,and0.01MMgCl2waslayeredover2.5ml
of40% sucroseinthe same
buffer,
and 0.1 mlof themitochondrial fraction was layered above the 15%
sucrose. Sodium lauroyl sarcosinate was added to
themitochondrial fractionto afinalconcentration of 0.1%,and the upperlayerwasgentlymixed.Thetube was immediately centrifuged in an SW-50 rotor at
51,000 X g for 30 min. The band of
magnesium-Sarkosyl crystals which collectedattheinterface ofthe sucrose solutions was harvested by puncturing the sideofthe tube.
RESULTS
Membraneassociatedinputviral RNA. Tostudy
the distribution of
radioactivity
after infectionwith 3H-SFV, chickcell
monolayers
wereinfectedwith a partially purified pool of 3H-SFV as
de-scribed. After1 hrat37C, thecells werewashed
and fractionated into nuclear and cytoplasmic extracts. Only 16% of the radioactivity applied
was adsorbed to the cells, even atthe low
mul-tiplicityofinfection and smallvolumeused(Table
1). Of theradioactivity associated with thecells,
about 90% waspresent in the cytoplasmic
frac-tion. Of this radioactivity, 36% was associated
with the mitochondrial fraction and 64%with the postmitochondrial fraction.
Whenthe mitochondrialandpostmitochondrial
fractions were analyzed in discontinuous 15 to
50% sucrosegradients(Fig. 1),mostofthe
radio-activity
present in themitochondrial fractionwas [image:2.492.258.452.91.207.2]found to sediment at a density of 1.18 (41%
TABLE 1. Distribution of radioactivity after infec-tion of chick fibroblasts with tritiated Semliki
Forest virus (3H-SFV)
Acid-pre-Determination cipitablecounts recovered
Total radioactivity added... 38,300
Washedoff cells... 31,200 Adsorbedto cell... 6,120
Nuclearfraction... 680
Cytoplasmicfraction... 5,300
Postmitochondrial fraction... 3,300
Mitochondrial fraction... 1,850
sucrose).Very little radioactivity was found at the
topof thegradient (Fig. 1A).On theother hand,
in the postmitochondrial fraction the reverse
situationwasseen. Most oftheradioactivity was
present at the top ofthe gradient, and a relatively minorfraction was present at its midpoint (Fig.
1B).
We were interested in thenature ofthe
radio-activity in the mitochondrial fraction and
sedi-mentationat adensityof1.18 since, later in
infec-tion, this region of the gradient in extracts of
Sindbis virus-infected cells has been shownto
con-tainan MRC with the replicative intermediate
form of viral RNAand the viral RNA-dependent
polymerase(15). The radioactivityofthis fraction
(Fig. IA) was probably not due to unecipsed
virus or to nucleocapsids with theirouter
mem-branecoatremoved. The peak of viral infectivity,
although notsharp, did notcoincide withthat of
the radioactivity (Fig. IA). Also, although the
uncoated viral RNA is completely ribonuclease
resistant, that of the fraction under study was
only partially resistant (Fig. 2). Moreover, when
artificial mixtures were prepared from the
cyto-plasm of SFV-infected cells and 3H-SFV,
3H-SFV core or nucleocapsid, or the purified viral
RNA forms (42, 26, and 20S RNA), only the
3H-SFV showed any tendency to adhere to the
mitochondrial fraction (Fig. 3); however, that
3H-SFV which did adhere tended to band at a
higher density than did the bulk ofthe
radioac-tivity examined in Fig. IA.Thenucleocapsidand
purifiedviral RNA formsremainedwiththe
post-mitocondrial fraction.
These resultssuggested to us that thebulk of
theradioactivity sedimentingwith the
mitochon-drial fraction was not due to
unecipsed
virus.Extraction of radioactive RNA fromthis fraction
tended to confirm this notion (Fig. 4). The
ex-tracted RNA was polydisperse with a peak at
about 20S. It was partially resistant to
ribonu-cleasetreatment. OftheRNA forms ofSFV,these
properties are most similar to those of the
on November 11, 2019 by guest
http://jvi.asm.org/
U.
CL
2,
200
r-C)
mnr
CO
20=
c),
:0n 0rtl
mc~ O ou
0 10 20 30
[image:3.492.254.451.69.219.2]FRACTiON NUMBER
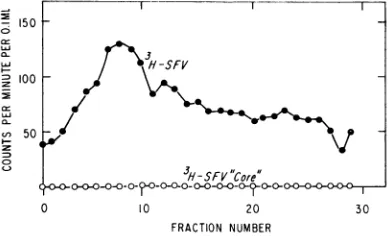
FIG. 1. Analysisin a discontinuous sucrose density
gradient of cytoplasmic radioactivity after infection
with 3H-SFV. Gradient fractions from the (A)
mito-chondrial or (B) postmitochondrial fraction were
analyzed for radioactivity and, in the case of the
mitochondrial fraction, infectivity. The top of the gradient in this andsubsequentfiguresis to the right.
tially double-stranded replicative intermediate (RI) form (5). No significant amount of 42S RNA, the RNA of the virion,wasfound. These
results do notrule outthe possibility that some
26Ssingle-strandedviral RNAwaspresentinthe mitochondrialfraction since the method of
analy-sis employed does not separate this form
com-pletely from the RI form (7). The bulk ofthe RNA, however, would appear to be in the RI form.
Tostudy thekinetics of the entry of viral RNA into the MRC, RNA was extracted from the
mitochondrial fraction ofinfected cells atearlier
time periods. Cells were washed five times after
virusadsorptionat4C,andmediumat37 Cwas
added. Mitochondrialfractions wereprepared at
5 and 20 min afterwarming. RNAwasthen
ex-tracted from these fractions and analyzed on sucrose density gradients. The results (Fig. 5)
showed that 5 min after warming, a peak
ap-pearedat 42S, correspondingtothe RNA of the
virion. Some slower sedimenting material was
alreadypresent. At20minafterwarming,mostof
- 150 C
: f~~~rec/ed 0-0--0 X
0 10 20 30
FRACTION NUMBER
F1G. 2. Ribonuclease resistance of radioactivity associated with the mitochondrial fraction after infec-tion with 3H-SFV. A mitochondrial fraction was pre-pared as in Fig. 1. One-half of the fraction was sedi-mented as in Fig. 1, and the rest was treated with ribo-nuclease (1 /2g/ml, 20min, 0 C) before sedimentation.
2-.150 ~~I T
-
100<vs
LI
3/-SSFV
"Core"iooneot-oO-o
-0 10 20 30
FRACTION NUMBER
FIG. 3. Sedimentation in themitochondrial fraction of radioactivity .from artificial mixtures containing 3H-SFV or 3H-SFV cores or 3H-SFV-RNA forms (42, 26, or 20SRNA). Preparations containing these were mixed with a cytoplasmic extract from SFV-infected chick cellsafter homogenization. Mitochondrial fractions were then prepared as in Fig. 1 and analyzed on 15 to 50% discontinuoussucrose density gradients.
NVo
radioactivity was found to be associated with the mitochondrial fractions from the mixtures of cytoplasm with 3H-SFV cores (as shown) or 3H-SFV-RNAforms (notshown).the labeled RNA was of the slower sedimenting species, probably, as at 1 hr after warming (Fig. 4), in the RI form. This result suggested that some 42S RNA rapidly became membrane-asso-ciated and was then converted to a replicating
RNAstructure.
Finally, additional results indicated that the radioactive fraction found in Fig. 1 was not due to uncoated 3H-SFV. In infection of chick cells with radioactive infectious RNA of the closely related Sindbis virus, the radioactivity of the
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.492.254.449.297.415.2]28S 18S
/0
,O Rlbo ucleose-frea/ed
C'
1 .
ICP0
O/d E 00i0o ,o OooX°> '
I T-'T -T
20 FRACTION NUMBER
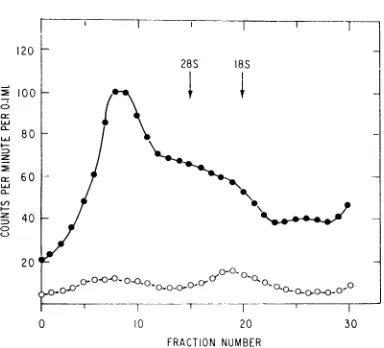
FIG. 4. Analysis ofRNA extractedfrom the
mito-chondrial fraction of3H-SFV-infectedcells. Cellswere
infected with 3H-SFV, and after I hr mitochondrial
fractions were preparedasdescribed in the legendto
Fig. 1. RNA was extracted in 0.1 M NaCl by an
SDS-phenol methodandanalyzed (300,000X g,1hr)
on6to30%sucrosedenisity gradients.Thedesignations
18S and 28S specify the peaks ofadded ribosomal RNA markers. One-half ofthemitochondrialfraction
was treatedin0.1Am NaCl with I ,ug ofriboniuclease permlfor20minat37 Cbeforesedimentation.
z 100
0
28S I8S z 75
uJ
z
,0
8
2
50s
LU
z 0 8 H;S
[image:4.492.52.247.64.250.2]FRACTION NUMBER
FIG. 5. Kinetics ofappearance ofviral RNAforms in the mitochondrialfractionzof 3H-SFV-infectedcells. Cellswereinfectedasdescribed inthelegendtoFig.4.
After 5 or 20 min, mitochondrial extracts were
pre-pared and RNAwasextractedfromthese.Analysisby sucrose density gradients wasalso carried out as
de-scribedinthelegendtoFig.4.Symbols: *,RNA
ex-tractedafter5min; 0,RNAextractedafter20 min.
mitochondrial fraction sedimented in the same mannerastheresult showninFig.IA(T. Sreeval-san,inpreparation). Also, analysisofthespecific activity of thevirus employed in theexperiment
shown in Fig. 1A (see above) demonstrated that
the titer of infectious virus recovered could not
account for the level ofradioactivity seen in the fractions containingthehighestlevels oftritium. Therefore,the association of viral RNAwith a specific cell fraction (Fig. IA) was probably not due toan artifactual interaction of viral RNA with cell membranesduringthepreparationof cy-toplasmic extracts since mixing of viral RNA forms with such extracts failed to cause such a
specific association. Additional evidence in favor
of the idea thatinputviralRNAwaspresentin an MRCwasdetermined in experiments employing
Sarkosyl magnesium precipitation ofmembrane
fractions.Ithas beendemonstrated that functional
bacterialdeoxyribonucleic acid (DNA) is associ-ated withamembrane and the complex may be
precipitated by magnesium with Sarkosyl (16).
Theprecipitation isspecificformembrane-bound functionalDNA,asDNAor RNAforms
artifici-ally mixed with cellextracts are notprecipitated.
When the mitochondrial fraction of cells infected
with3H-SFVwas precipitated with Sarkosyl and
magnesiumandtheprecipitate bandedat a15and
40% sucrose interface, most of theradioactivity
recoveredwasfound in theinterface band (Table 2). The counts present in the postmitochondrial
fraction or in artificial mixtures of viral RNA
forms and cell sap did not band atthe interface
whensubjectedtothesametreatment.
Theresults discussedsofarsuggested that, 1 hr
after productive infection with 3H-SFV, a small percentageof the
input
viral RNAwasspecificallyassociated with a membrane component of the
mitochondrial fraction.By 1 hr afterwarming to
37C, mostof this RNA wasprobably inthe RI form. This membrane component had, therefore,
someof thecharacteristics ofaviralMRC. Effectofcycloheximideorinterferon on
associa-tionininputRNAwithcellmembrane. Tostudythe
effect of cycloheximide or interferon on input
viral RNA, chick cells were treated with 1,000
units/ml of
partially
purified chick interferonbeforeinfectionwith 3H-SFV, or, with 100
Ag
ofcycloheximide perml during infection. The dose
of interferon employedinhibited virusgrowthby
more than 200-fold; the dose of cycloheximide
inhibited protein synthesis by94%. The effect of
these treatments on the distribution of cyto-plasmicradioactivitywasstudied (Table 3). In the control cells, the distribution of cytoplasmic
radioactivitybetween themitochondrial and
post-mitochondrial fractionswassimilartothat shown in Table 1. When thespecificactivityofthe
radio-activity in these fractions was studied, however,
morethan2.5 times the activity of the postmito-chondrial fraction was present in the mitochon-drial fraction. Inthe cells treated with
interferon,
the distribution ofradioactivity
wasapproxi-matelythesame asin thecontrol. Incells treated
120
0 100
Z 80
X 60
4z
(-- 40)
20
0
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.492.55.246.372.505.2]TABLE 2. Distribution of radioactivity after sedi-mentation of a sodium lauroyl
sarcosinate-treated mitochondrial fraction
Acid-pre-Determination cipitablecounts recovered
Total radioactivity added... 1,055
Top component.. 90
[image:5.492.252.443.61.353.2]Interface precipitate... 816
TABLE 3. Distribution ofcytoplasmic radioactivity after infection of chick fibroblasts with
tritiated Semliki Forest virus
Ratio of Total Specific specific radio- activity activities
fraction acti - (counts
(Mito-Cytoplasmic
frraction
aecotvit
pe min chondrial(ecounret
(counts/pepg fraction: of postmito-mm) protein) chondrial fraction)Control
Mitochondrial ... 8,339 11.9 2.6:1 Postmitochondrial. 17,596 4.65
Interferon-treated
Mitochondrial... 8,425 14.0 2.4:1 Postmitochondrial. 13,443 5.85
Cycloheximide-treated
Mitochondrial... 3,288 4.50 0.66:1
Postmitochondrial.. 24, 726 6.82
withcycloheximide during infection, however, the bulk of the specific activity was in the
postmito-chondrialfraction.
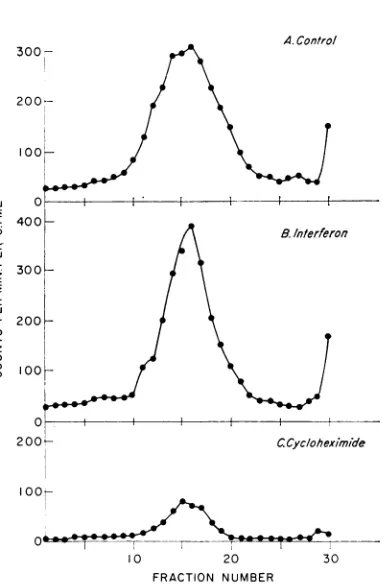
These findingswereconfirmed insucrose
den-sity sedimentation studies on the mitochondrial
fraction (Fig. 6). Again, the control containeda
peak ofradioactivity sedimentingat adensity of
1.18 (Fig. 6A).Asexpected, the interferon-treated
cells contained a similar, but usually an even
sharper, peak, asshowninFig.6B. On theother
hand, in cells treated with cycloheximide during
infection, thepeak atthis densitywasvery much
decreased (Fig. 6C). Therefore, inputviral RNA
appearedto enterareplication complex (RC) -like structure after interferon treatment, butthis step
was inhibited in the presence ofcycloheximide.
Onepointof interestwasthenatureofthe viral RNA present in the RC-like structure in
inter-feron-treated cells. As shown in Fig. 4, under
ordinary conditions the predominant RNA
species present in the mitochondrial fraction of control cells after 1 hr was the RI form. It has
also been shown that, in cells treated with high
concentrations of interferon, little or no
ribo-nuclease-resistant RNA was formed from input
LdJ
a-z
n
z
0
U
A. Control
I
I10 20
FRACTION NUMBER
30
FIG. 6. Effectofinterferon orcycloheximide treat-ment on association ofinput 3H-SFV with
mitochon-drialfractionis. (A) Untreated cells; (B) cells treated
with 1,000 unitsof interferon permlfor 14 hrbefore
infection; (C)cellstreated with 100,ug ofcycloheximide
per ml during infectiont. The cells were infectedwithl
3H-SFV and, after I hr, mitochondrialfractions were
prepared and analyzed on discontinuous 15 to 50%
sucrosedensitygradients asdescribed in thelegendto
Fig. 1.
SFV RNA (6).Thepresenceof viralRNA inthe MRCstructureofinterferon-treated cells seemed
paradoxical. RNAwas,therefore,extracted from the mitochondrial fraction of interferon-treated cells. Theresults(Fig. 7)showthat themajor peak
of RNA presentwasin the 42S form, inmarked
contrast tothepredominant205 peakinthe
con-trol cells (Fig. 4). In addition, as expected, little or noribonuclease-resistant RNA could be
dem-onstrated in the mitochondrial fraction of the interferon-treated cells (Fig. 7). In this respect, the interferon-treated cells after 1 hr resembled
the control cellsveryearlyafterwarmingto37 C
(Fig. 5).
DISCUSSION
The results indicated that inuntreated,infected cells a small fraction of the input 3H-SFV RNA entersanMRCstructure. Most, if notall,ofthis
VOL.
173
oo.-n
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.492.42.238.161.386.2]100
C-w 80
c 60
2 40
:D
-o-oQ'0O-°- °~oo_ oo- °'
_-0 10 20
[image:6.492.62.254.66.240.2]FRACTION NUMBER
FIG. 7. Analysis ofRNA extracted from the
mito-chondrial fraction of 3H-SFV-infected cells which had
been treated with interferon. Cells were infected with
3H-SFV after 14 hr oftreatment with 1,000 units of interferon per ml. Mitochondrial fractions were
pre-pared, and RNA was extractedfrom them and
ana-lyzedasinFig. 4.
RNA is in the RI form (5), suggesting that the
structure is indeed associated with virus RNA synthesis.Later ininfection, functionalviralRNA
polymerase is present in a very similar structure
(15).
Results from cycloheximide-treated cells sug-gest that entry ofinputRNA into this structure
requires protein synthesis, since this viral func-tionisinhibited. Onthe otherhand, no such
in-hibition was seen in interferon-treated cells, but
the viral RNA present in the MRC structure
remained for themostpart inthe 42S form pres-entin the virion.
The resultswith interferon indicate that,
what-everthe site ofits actionis, this must liebeyond thestepinwhichtheinputRNAisintegrated into the MRC structure. Interferon treatment has no
effectonviraluncoating (2),exceptinthecaseof
vacciniavirusin which thisispartially dependent
on viral protein synthesis (11). Since treatment
with 1,000 units of interferonper ml appears to
block translation of input RNA in the
SFV-infected cell (4), the integration step probably
doesnotrequire theviral RNApolymerase. This conclusion was strengthened by findings with a
temperature-sensitive mutant of the closely
re-latedSindbisvirus, TS-6. The temperature-sensi-tive step ofTS-6 isrelatedto viralRNA
polym-eraseproduction, but TS-6 RNAhasbeen found toentertheRC-likestructure atpermissive(27 C)
andnonpermissive (39 C) temperatures. Atboth
temperatures, however, cycloheximide treatment
blocks this step (T. Sreevalsan, in preparation).
In all cases studied, therefore, the action of
cycloheximidewas to inhibitviral RNAentryin the MRCstructure; thenatureofthe protein
re-quired for this function is ofsome interest. The
currentconceptofinterferon action is that
trans-lation of viral, butnotcellular,messengerRNAis
blocked ininterferon-treatedcells (14). The pres-entresults, taken together,suggest thatitmaybe
thesynthesis ofacellprotein which is requiredfor
theintegration ofinput viral RNA into the
RC-like structure. If the latter hypothesis is correct, someofthe properties ofthis cell proteincan be
deduced. The protein would not be one
specifi-cally induced by virus infection, since the virus replicates quite well in actinomycin D-treated cells. Also the protein must have a fairly rapid
turnover rate,aconclusion suggestedby the result
obtained with cycloheximide. The existence ofa
host proteincontrolling virus replication in C-M cells persistently infected with mumps virus was
recently reported. This protein alsoseemstohave the properties noted above (13).
An alternative explanation for our
observa-tions would be that the integration step requires viral protein synthesis andthat cycloheximide isa more efficientinhibitor of virus protein synthesis
than isinterferon. In the lattercase,residual virus
proteinsynthesis ininterferon-treatedcellswould
be responsible forthe integrationstep. Interferon hasbeen showntoinhibitsome stepsinthe
syn-thesis of viral RNA more effectively than others (12).
LITERATURE CITED
1. Cheng,P. Y. 1961. Purification, size, and morphology ofa
mosquito-borne animal virus, Semliki Forest virus.Virology 14:124-131.
2. DeSomer, P., A. Prinzie,P.Denys, Jr.,and E.Schonne.1962. Mechanism of action ofinterferon. I. Relationship with viralribonucleic acid.Virology 16:63-70.
3. Fantes, K.H. 1965.Furtherpurificationof chickinterferon. Nature(London)207:1298.
4. Friedman,R. M. 1968. Inhibition ofarbovirusprotein syn-thesisbyinterferon. J. Virol.2:1081-1085.
5. Friedman,R.M. 1968.Replicative intermediate ofan arbo-virus. J. Virol. 2:547-552.
6. Friedman,R.M.,K. H.Fantes,H. B.Levy,andW.B.Carter.
1967. Interferon actionon parental Semliki Forest virus
ribonucleic acid.J. Virol.1:1168-1173.
7. Friedman, R. M., H.B.Levy,andW.B. Carter. 1966.
Replica-tion of Semliki Forest virus.Three forms of viral RNA pro-ducedduringinfection. Proc. Nat. Acad. Sci.U.S.A. 56: 440-446.
8. Friedman, R. M., and J. A.Sonnabend. 1965. Inhibitionof inteferon action by puromycin.J. Immunol. 95:696-703. 9. Girard, M.,D.Baltimore,and J.E.Darnell.1967. The
polio-virus replication complex: site forsynthesis ofpoliovirus
RNA. J. Mol. Biol. 24:59-74.
10. Lowry,0.H.,N.J.Rosebough,A.L.Farr,and R.J.Randall.
1951.Proteinmeasurementwith the Folinphenolreagent. J. Biol. Chem. 193:265-275.
11. Magee, W. E.,S. Levine,0.V.Miller,andR. D.Hamilton.
on November 11, 2019 by guest
http://jvi.asm.org/
1968.Inhibitionbyinterferonof theuncoatingof vaccinia virus.Virology 35:505-511.
12. Mecs, E., J.A.Sonnabend,E.M.Martin,and K. H.Fantes.
1967. The effect ofinterferononthesynthesis of RNA in chickcells infected with Semliki Forest virus. J. Gen. Virol. 1:25-40.
13. Northrop,R.L.1969.EffectofpuromycinandactinomycinD
onapersistentmumpsvirusinfection in vitro. J. Virol. 4: 133-140.
14. Sonnabend,J.A.,andR. M. Friedman. 1967.Mechanismsof interferonaction,p.202-231. In N. B. Finter (ed.), Inter-ferons. North-Holland Publishing Co., Amsterdam. 15. Sreevalsan, T., and F. H. Yin. 1969. Sindbis virus-induced viral
ribonucleic acid polymerase. J. Virol.3:599-604. 16.Tremblay,G.Y., M. J.Daniels, and M.Schaechter. 1969.
Iso-lation ofa cell membrane-DNA-nascent RNA complex
from bacteria.J. Mol. Biol.40:65-76.