Vol. 38, No. 2 JOURNAL OFVIROLOGY, May1981,p.504-517
0022-538X/81/050504-14$02.00/0
Vesicular
Stomatitis Virus mRNA and
Inhibition of
Translation
of Cellular mRNA-Is There a P Function in
Vesicular Stomatitis Virus?
HARVEYF. LODISH*AND MARY PORTER
Department of Biology, Massachusetts Institute of Technology,Cambridge, Massachusetts 02139 Received 15December1980/Accepted4February1981
Infectionof animalcellsbyvesicular stomatitis virus(VSV) results in inhibition
of translation of cellular mRNA. We showed previously that, in BHK cells
infected by the Glasgow isolate of VSV Indiana, this is duetocompetition during
the initiationstepofproteinsynthesis of viral and cellular mRNA fora constant,
limiting number of ribosomes. We show herethat infection of thesamecellswith
the San Juan isolate of VSV resulted in a morerapid shutoff of host protein
synthesis and that this was paralleled by a more rapid accumulation of viral
mRNA.Extendingourconclusion that shutoff is duetomRNAcompetition,we
show further that the average size ofpolysomes translating viral and cellular
nmRNA wasthreefold smaller in cellsinfectedby VSV San Juan than by VSV
Glasgow,which, inturn, wasabout one-half that of uninfectedcells.In allcases,
cellular andviral mRNA's which encoded the same-sized polypeptideswerefound
onthesame-sizedpolysomes,aresultindicatingthat theefficiency of translation
of bothtypesofmRNA's is about thesamein the infected cell.Also, therewasno
preferential sequestration of viralorcellular mRNA's in ribonucleoprotein
par-ticles.Additionalcorrelations between the levels of viral mRNA's and the
inhi-bition ofproteinsynthesiscamefromstudies of three otherwild-type VSV strains
andalso fromstudies with Vero and Lcells. Inparticular,therateof shutoff of
L-cellproteinsynthesisafter infectionbyanyVSV isolatewasslower than thatin
BHK cells, and this was correlated withaslowerrate of accumulation of viral
mRNA. VSVtemperature-sensitive mutantswhich synthesized, atthe
nonper-missive temperature,noVSV mRNAfailed toinhibitsynthesisofcellular
pro-teins. Stanners andco-workers (C. P. Stanners, A. M. Francoeur, and T. Lam,
Cell 11:273-281,1977)claimed that VSVmutantRlinhibitedsynthesis of Lcell
protein synthesis less rapidly than did its parent wild-type strain HR. They
concluded that this effectwasdueto amutation inanunspecified VSV protein,
"P."Wefound, in bothLand BHK cells, that Rl infection resulted inaslightly
slower inhibition ofcellular mRNA translation than did HRinfection and that
this was correlated with aslightly reducedaccumulation of VSV mRNA. The
level of VSV mRNA, rather thananyspecific VSV protein, appeared tobe the
keyfactorin
determining
therateofshutoff of host protein synthesis.Infection of mammaliancellsbyvesicular
sto-matitis virus(VSV) results ininhibitionof syn-thesisofcellularprotein and RNA andeventual cell death. Neither replication of the genomic RNA norproduction ofinfectious virus is nec-essary for theinhibitionofcellularprotein
syn-thesis;however, sometranscriptionofthe
nega-tive-strandedviralgenome into mRNA is
essen-tial(3,4,6-8).Much,ifnot all, of theinhibition
ofcellular proteinsynthesis ismanifestedat the
translational level: after infection cellular
mRNAremainsintact and fullyfunctionalin in
vitro translationsystems, yet istranslatedin the
cellsat adecreasingrate (2).
Atleast three different mechanisms for this
translationalcontrol have beenproposed. Nuss
etal. suggested thatVSV mRNAinitiates
pro-tein synthesis severalfoldmore efficiently than
does cellular mRNA, and thus out-competes
mRNA for ribosomes (9). Stanners et al. (12) have isolated a mutant ofthe HR (Winnipeg)
wild-type isolateof theIndiana serotype ofVSV.
Infection ofL cells with this mutant does not
resultintherapid,precipitous inhibition of
pro-tein synthesisobserved with the HRwild-type
strain. Theyproposed thataspecific viralgene
product,termedP, isrequiredfor thisinhibition
andthat themutant,Rl,is P-.
504
on November 10, 2019 by guest
http://jvi.asm.org/
IS VSV?
More recently, we showed that in growing
BHKcells over 80% of most of thepredominant
species of cellular mRNA are bound to
poly-somes, and over 60% remainonpolysomes 4 h
after VSV infection (2). The average size of
polysomestranslating individual cellular mRNA
isreduced about two- to threefold after infection.
Forexample, in uninfectedcells, actin
(molecu-lar weight, 42,000) mRNA is found
predomi-nantly onpolysomes with12ribosomes;4hafter
infection it isfound onfivesomes, the same size
aspolysomes that aretranslating VSV N
(mo-lecularweight 52,000) and M(molecularweight
35,000) mRNA. We concluded that the
inhibi-tion ofcellular protein synthesis after VSV
in-fection isdue, in large measure, to competition
for ribosomes by alarge excess of viral mRNA.
Theefficiency of initiation of translation on
cel-lular and viral mRNA's is about the same in
infectedcells; cellular ribosomes are simply
dis-tributedamong more mRNA's (about threefold
more) than are present in growing cells.About
20 to 30% of each of the predominant cellular
andviral mRNA's are present in
ribonucleopro-tein (RNP) particles in infected cells and are
presumably inactive in protein synthesis. There
is no preferential sequestration of cellular or
viral mRNA's in RNPs after infection (2). All of the above-mentioned studies were done with different isolates ofVSV Indiana and in
different cultured cells.Aspart of asystematic
approach to the resolution of thesedifferences,
we have studied the effects of four commonly
used wild-type VSV strains and also the Rl mutant on three of the cell linesused in these
studies. Ourresults indicate that the rate and
extentofinhibition isafunction of both the cell
line and virus used. In allcasestested,however,
therewas agoodcorrelation between the
accu-mulation of viral mRNA and the extent of
inhi-bition of translation of cellular mRNA, thus
extendingourconclusionsonthe mechanism of
translational control to other cell line-VSV
strain combinations. Our studies also suggest
that Rlmaynotbeasimplemutantof the HR
wild-typestrain.
MATERIALS AND METHODS
Virus strains. The Glasgow isolate ofVSV was obtainedfrom RobinWeiss(ImperialCancer Research Fund,London),whooriginallyobtainedit fromCraig Pringle(InstituteofVirology,Glasgow) (3). The San Juan isolate of VSVwasobtained from Robert Laz-zarini(NationalInstitutes ofHealth),who obtained it from MarthaStampferandAliceHuangin 1972. The Mudd-Summers strain was also obtained from Dr. Lazzarini. According to him (personal communica-tion), itwasfirstusedatHarvard and thenpassedon
insuccessiontoDrs.Carver, Marcus,Summers,Mudd, andHolland beforereachinghim.StrainsHR andRl
aredescribed by Stanners et al. (12) and were obtained from C. Stanners (Ontario Cancer Institute). All vi-ruses used in these studies were grown on Vero cells.
Animalcelis and infections. Growth of suspen-sion BHKcells and monolayer Verocells and proce-dures for infection with VSV weredetailed in previous papers (2, 3). Mouse L cells were obtained from C. Stanners and were cultured as monolayers in minimal essential a medium (GIBCO no. 410-1900) with 7% calf serum and 3% fetal calf serum.Infection of these with VSV used a protocol identical to that for Vero cells. In all cases, 10PFU of VSV per cell was used.
Labeling ofinfected cells with[3'Smethionine
andanalysis of proteins by sodium dodecyl sul-fate (SDS)-gel electrophoresis. Procedure for la-belingandelectrophoresis have been fully described inpreviouspublications (2, 3).
Isolation of total cytoplasmic RNA. A previous publication detailed our procedures for isolation of polysomes and polysomal RNA from BHKcellsand forcell-free protein synthesis (2). The procedure de-veloped for extraction of RNA from BHKcellswas used withonlyminormodificationsfor monolayer L cells.About 5x 107cellsin three 150-mm-diameter plates were chilled on ice and washed several times withphosphate-bufferedsaline.Thecells were scraped off theplatesintophosphate-buffered saline and then treated as in theprotocol for suspension cells (2). Note that total cytoplasmic RNA, not selected for polya-denylatedcomponents,wasused.Thus, mRNA activ-itiesof RNAfragmentsarenormalized to totalcellular RNA, i.e., mostlyrRNA,whichisproportional to the number of cellsfractionated.
RESULTS
Virus andcell strains. Much of this study
is a comparison of the effects of five strains of
VSV onthree different host cells. Tracing the
historyof thecommonlaboratorystrainsofVSV Indiana is only slightly less complicated than
followingthegenealogyofOld Testament
patri-archs,but our bestattemptispresentedin
Ma-terials andMethods.Strain Rl is a
temperature-resistantrevertantoftemperature-sensitive (ts)
mutantT1023, which is itselfreported to be a ts
mutant of the HR strainwith alesionin
com-plementation group I (12). The BHK culture
used wasgrowninsuspension, whereas the Vero
andL-cellculturesweremaintained andinfected
as monolayers. Analysis by one-dimensional
SDS-gels of the [3S]methionine-labeled
pro-teinssynthesizedbythese (uninfected) cells
re-veals fewsimilarities except for actin,which is
inall cases the mostpredominant cellprotein.
Thus, to facilitate comparison ofdifferent cell
lines,we have focused onthe translation ofactin
mRNA. After infection of BHK cells by the
Glasgow isolateofVSV,inhibition ofactin
syn-thesis parallels, we have shown, that of most cellularproteins (2).
BHKcellsinfectedby differentstrains of VSV. Infection of BHK cells with four ofthe VOL. 38,1981
on November 10, 2019 by guest
http://jvi.asm.org/
506 LODISH AND PORTER
five virus strains tested (Glasgow,
Mudd-Sum-mers,HR, andR1) resulted in similar kinetics of
inhibition of synthesis of actin andofmost
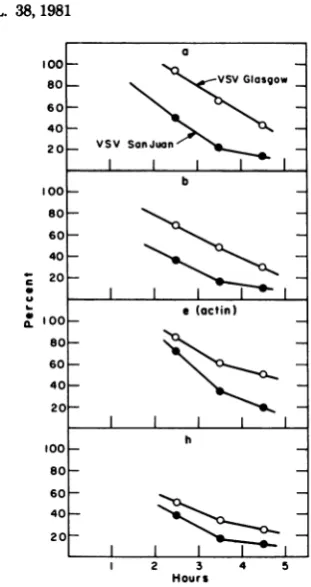
cel-lular proteins (Fig. 1 and 2). At 2.5 h after
infection, the rate of actin synthesiswas 60 to
75% that of mock-infected cells, andat4h itwas
25 to30%. Infectionby VSV SanJuan,by con-trast,resulted in amuchmore rapidinhibition
of synthesis of predominant cellular proteins
(Fig. 2-4); actinsynthesiswasreducedto45%of
the control value at 2.5 h after infection, 1 h
sooner than after infection by the other VSV
isolates.Inhibition ofsynthesisofother
predom-inant cellular proteins also occurred about 1 h
sooner after infection by VSV San Juan than
VSVGlasgow (Fig. 4).In allcases,by5h
post-infection actin synthesis wasless than 10% of
controllevels (datanotshown).Thisresult,
to-gether with microscope studies on these cells,
indicated thatover90%,andprobablyover98%,
wereinfected with VSV.
The inhibition of cellular protein synthesis
after infection ofBHKcellsby VSVGlasgow is
due, wehave shown (2), tothe competition of
cellular and viral mRNA for theconstant,
lim-iting numberof functional ribosomes. Were this
toapplytoinfection of BHK cellsby VSV San
Juan, thenonewouldexpect a morerapidrate
ofaccumulation of viral mRNA after infection
by VSV San Juan. Figure 3 andTable 1 show
that thiswasthecase.Inthis study,aconstant,
limitingamountofcytoplasmic RNAwas
trans-lated inawheatgermcell-freesystem.The
one-dimensional gelanalysis of Fig.3(lanes11to18)
makestheimportantpoint that all translatable
cellularmRNA's remained intact andfunctional
after infectionby both VSV Glasgow and VSV
San Juan. Permicrogram of cellular RNA,
prep-arations from infected and uninfected cells
di-rected the synthesis of equal amounts of all
predominantcellular polypeptides.Many of the
translation products of cellular mRNA
comi-gratedwith predominantprotein species
synthe-sizedby the growingcells.Anotableexample is
actin (polypeptide y), but conclusive
identifica-tion ofthesecellularspecies must awaitpeptide
mapping. The additional translation activity
present in RNA from infectedcellsdirected
syn-thesis of the five VSV structural proteins, plus
afew otherpolypeptides which may have been
incompleteordegraded products of these. The
amounts ofboth total translatable mRNA and viral mRNAincreased steadily from 2.5 h after
infection, and did so about 1 h earlier after
infection by VSV SanJuan than byVSV
Glas-gow (Table 1).Therefore,the differential
accu-mulation of viral mRNA paralleled the
differ-entialinhibition ofhostprotein synthesis by the
twostrains. The increased levelof VSVmRNA's
inSanJuan-infectedcells wasparalleledby an
c
- -~~~~d
I
-~~~N/NS
BHsactin C
2.5(aeh -)o 4.5h(lns713hferifcin (Lane 6, 7,l
andly3)
Mockprtinfecnted;s(1
and8)vsvGlasgow; (2and9)VSV SanJuan; (3 and10) VSV Mudd-Summers;(4and11)VSVHR; (5and12) VSV Rl. Samples ofthe cell lysates were analyzed by electrophoresis throughalinear10 to15% polyacryl-amidegel containing SDS. Shown is a
radioauto-grainofthe driedgel. Theuppercaselettersonthe
rightdenoteVSVproteins,andthelowercaseletters indicatepredominant species ofcellularpolypeptides. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
a.
40-20_
2 Hours
FIG. 2. Synthesis of actin inBHI different VSVstrains. Theradioau
were scannedwith a microdensitoa
ditions where the darkeningof the proportional to the amount ofrac
Theamountofradioactivityinthe(
relativetothatofmock-infectedcel function of time after infection. Alt inFig.1,therewas noreduction ofo
Ihafter infection ofanyofthestraii
(0) VSV SanJuan (SJ): (-O-) VS (A) VSV HR (HR); (0) VSVMud (--O--) VSVRI (Rl).
accelerated rate of synthesis oi comparedwithinfectionbyVSV
parelanes 4and7and lanes 5a] TheamountofviralmRNAs3 infection of BHKcellsbyVSVs
Rlwassimilartothat in VSV G
cells (Table 2). Togetherwitho
1and2) thatthekinetics of inhil protein synthesisarethesame,t withourconclusion that host shu a function of the amount of tr mRNA.
Is the increasedlevel of viral infected by VSV San Juan, r4
Glasgow, indeedthe causeofti hibition of translation of cellu]
answerthisquestion,westudied distribution of the predom mRNA's and of theviral mRNA tion.Figure5 shows that83% of3
ribosomeswere onpolysomesin and 75% were on polysomes af eithervirus. However,theaver
somes was smallerin the VSVSa cells than in those infected by
I thesewere,inturn,smaller than those in mock-Cells infected cells. At this time the rate ofprotein
synthesis,
as measured by incorporation of [S]methionine,
relative to that ofuninfectedcellswas82% after VSV San Juan infectionand
87% after VSVGlasgowinfection.Thus,mostof
the polysomes in the infected cells must have
been functional. Theprofiles in Fig. 5a,
there-fore, indicate that thesamenumber of functional
ribosomesaredistributedamong moremRNA's
(i.e., viralplus cellular) after infection by VSV
Glasgow, andyet more after infection by VSV
\\ sHR San Juan.
\ MS
Considerably
moreinformationis obtainedby\ GL
analyzing
the translationproducts directedby RI- RNA from individualpolysome and RNPfrac-tions onalong polyacrylamide gradient gel (2).
SJ By quantitating with a microdensitometer the amountof eachpolypeptide synthesizedby each i i
gradient
fraction,
it ispossible
todetermine thesubcellular localization of many predominant
cellularmRNA's encoding proteins of molecular
Kcellsifected by weights from 20,000 to 70,000 (Fig. 6).
tgramsof
Fig.
I With few exceptions (proteins/i,
8, and D infhlmwas strictly
Fig.
3), in uninfectedcells over 85% of thetrans-dioactive
protein. latable mRNA encoding the 12 predominant actinpolypeptide, cellularproteins
was bound topolysomes
(Fig.
fIs, is plotted as a6;
see also reference2).
Ingeneral,
the size of 'hough not shown polysomes translatinganygivencellularmRNA actinsynthesis at wasproportionalto the size of theproteinprod-n-sused.Symbols: uct.ActinmRNA (y;molecularweight, 43,000)
VGlasgow
(GL):
was localized predominantly in fractionscon-'d-Summers
(NS);
taining
12 ribosomes, and 0 (molecular weight,31,000) mRNAwas localized inpolysome
frac-tionscontaining6to 8,whereasmRNA's
encod-fVSV proteins ing proteins 1, ic, X, and ,u (molecular weight,
rGlasgow(com- about20,000)wereenriched in fractions
contain-nd8ofFig. 3). ing5ribosomes.
ynthesized
after Three hours after infectionby VSV Glasgow,;trains HR4 and actin mRNA waslocalized on hexasomes, 50%
'lasgow-infected the sizeofpolysomes translatingactin mRNA in
urfinding (Fig. growing cells.
Importantly,
viral and cellular)ition
ofcellular mRNA's encoding proteins of about the same,hisisconsistent size (N,52,000daltons; M,35,000daltons; actin,
itoff isprimarily 42,000 daltons: and0,31,000daltons)were
trans-anslatable viral latedonpolysomes containingthesamenumber
ofribosomes,inthiscasefourtosix(Fig.6). We
mRNA incells take thisasevidence that therateof
polypeptide
elative to VSV chaininitiation,relativetoelongation,ontypical
ie enhanced in- viral and cellular mRNA's is about the same.
lar mRNA? To The polysomal mRNA's encoding the smaller
Ithesubcellular proteins (I,K,
X,
and,u) were enriched instruc-iinant cellular turescontainingtwo tothreeribosomes,avalue
k3h after infec- significantly smaller than the fivesomes which
IH-labeled
BHK translate these mRNA's ingrowing
cells.uninfectedcells, The sizeof
polysomes translating
allpredom-terinfection by inant cellular and viral mRNA's was much age size ofpoly- smaller after infection byVSV San Juan than
inJuan-infected by VSV Glasgow. Actin mRNA, to cite one
r VSV Glasgow: example,waslocalized
predominantly
todi-andVOL. 38,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.500.48.244.61.291.2]-G
-a
_R
N/NS
-
y
- E
I
M
~.
m-'I
-K
-I"
1
2 3 4
5
6
8 9
10
11
12
13
1415
1E
Uninf.
VSV
VSV
Unninf
VSV
VSVGlasgow
San
Glasgow
San
Juan
Juan
Cell
-free Translation
508
a-d
actin e
Cells
on November 10, 2019 by guest
http://jvi.asm.org/
IS THERE A P FUNCTION IN VSV? 509
100_
80 VSV Glasgow
60_
40-20 VSV SonJuan
b
100
60
40
-c 20 I
, e (actin)
800
60
-
40-
20-1
h
100
soo-80__
60
40-20
F
2 3 4 5
Hours
FIG. 4. Synthesis ofBHKproteinsafterinfection with VSVGlasgowor VSV San Juan. The autora-diogram of Fig. 3 (lanes 1-9) was scanned with a
microdensitometer. Theareas underthepeaks
cor-respondingtofour predominantcellular proteins,a,
b,c(actin),andh, weredetermined and normalized tothevalueof mock-infectedcells.
trisomes, only 17 to 25% the size ofpolysomes translatingactin mRNA ingrowingcells.Again,
theviral and cellular mRNA'sencoding proteins of thesamesize (N, M, actin,and 6)were trans-latedonpolysomes containingthesamenumber
ofribosomes, heretwo tothree.
Togetherwith the data ofFig. 3 and4, the study inFig.6indicatedthat the reduction after infection in the numberofribosomestranslating each moleculeofactin mRNA and otherspecies of cellularmRNAwas areflection, primarily,of
the increased amount of total translatable
mRNAspecies (predominantlyviral mRNA)in thecell.Virtually thesamenumber ofribosomes were distributed among a larger number of
mRNAmolecules.Thus, the twofold increase in
the totalamountoftranslatablemRNA 3 hafter
infection by VSVGlasgow observed in this
ex-perimentwasreflectedinatwofold reduction in
thenumber ofribosomesperactin mRNA. The
fivefoldincrease foundinthis experimentafter VSV San Juan infectionresulted in a
propor-tionately larger reduction inthe number of
ri-bosomes translating actin mRNA, and thus in the rate of actin synthesis. In neithercase did
there appear to be preferential translation of viral mRNA's or of cellular mRNA's in the
infectedcells,norwastherepreferential
seques-tration of particular cellular RNAs intoinactive
RNPs;about thesamefractions of viral mRNA's
and typicalcellular mRNA'swerein RNPs
(Ta-ble3).
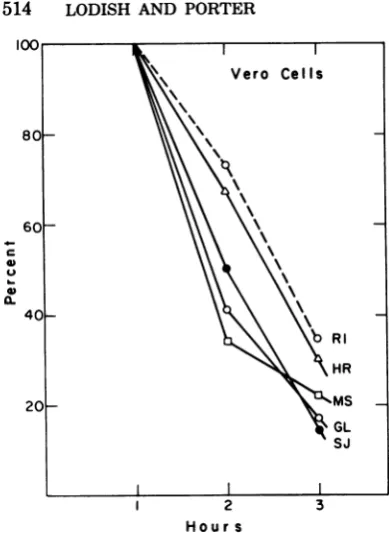
Vero cells.Inhibition ofcellular protein
syn-thesis byall strains of VSV occurredata
some-what fasterrateinVero cells than in BHKcells
(Fig. 7 and 8). The rate of synthesis of total protein 3 h afterinfectionwasgreaterthan 80% of that obtained inuninfected cells (seefigure
legends). Depending on the virus strain, actin
synthesiswasreducedto15 to30% of the control value by 3h. In allcases,by 5 h actin synthesis wasless than 10% ofnornal, andallofthecells
hadrounded andpartly detached from the plate. Thus, wellover 90% of the cells were infected.
Note, however, that the relativeefficiencyofthe different VSV strainstoinhibit cellularprotein synthesis differed in the two types ofcells. In
BHKcells, VSVSan Juaninhibited more
rap-idly than did VSV Glasgow, whereas in Vero cells, both inhibitedatabout thesamerate(Fig.
8). However, in both Vero and BHK cells,
shutoffby VSV HR and Rl strainswasslightly
slower than by VSV Glasgow (compare Fig. 8 and 2). Because the differences among VSV
strainswaslesspronouncedin Verocells thanin
BHKcells,we didnotattempttocorrelatethe
different profiles of shutoffof Veroproteinsby
FIG. 3. Gelanalysis ofproteins synthesizedininfectedBHKcells andofcell-freetranslationproductsof RNAfrom infectedcells. A linear 10to 15%gradientpolyacrylamide gelwas used. (Lanes 1-9) Proteins
synthesizedduringa30-minlabeling periodwith[3S]methionine, beginningatthe indicated time.(1) Mock-infectedcells,2.5h; (2)3.5h; (3)4.5h; (4) VSVGlasgow-infected cells,2.5h; (5)3.5h;(6)4.5h; (7) VSV San Juan-infected cells,2.5h; (8)3.5h;(9)4.5 h.AsinFig. 1,the lower-caselettersrepresentpredominantspecies ofhostproteins. (Lanes10-18)Proteinssynthesizedinawheatgermcell-freesystemusing2 gofthe indicated
RNAsper25-,dreaction.A5-,ulsample ofthereactionwasanalyzed. Table1presentsthe totalamountof
acid-precipitable radioactivity incorporatedperreaction, as well astherelative amountofVSVproteins made. (10)RNAfrommock-infected cells,2.5h; (11)3.5h; (12)4.5h; (13)RNAfrom VSV-Glasgow-infected cells,2.5h;(14)3.5h;(15)4.5h; (16)RNAfromVSV SanJuan-infected cells,2.5h;(17)3.5h; (18)4.5 h. The
Greek lettersontherightdenotepredominanttranslationproducts ofcellularmRNA,and thecapitalletters
arethe VSV structuralproteins.
r
a
I
VOL. 38,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.500.67.223.64.358.2]510 LODISH AND PORTER
the various strains(whicharereproducible)with
the levels of VSV mRNA's.
MouseLcells.Therateof inhibition of
cel-lular proteinsynthesis after VSVinfectionwas
much slower in L cells (Fig. 9 and 10) and in
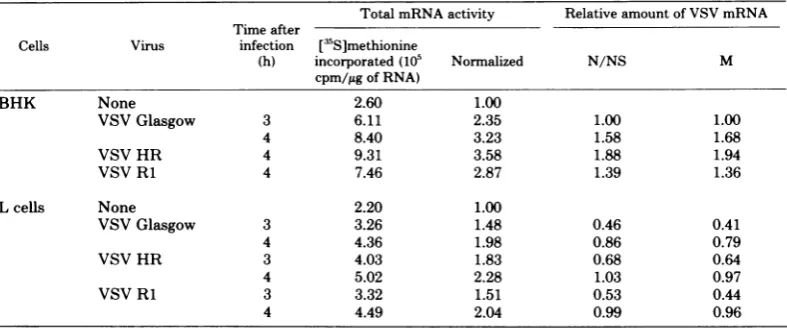
TABLE 1. Messenger activityin RNAfrom VSV-infected BHK celisa
Relative amt of mRNA activity VSV mRNA
Cell
5['S]methi-onine incor-
Normal-porated (105 °zed N/NS M cpm/,ug of
RNA)
Uninfected 4.18 1.00 VSVGlasgow
2.5h 4.98 1.19 1.00 1.00
3.5h 6.99 1.67 3.50 3.62
4.5 h 13.86 3.31 11.50 12.10
VSV San Juan
2.5 h 8.41 2.01 4.85 5.31
3.5 h 12.98 3.10 10.85 11.10
4.5 h 20.59 4.92 19.63 21.60
'Total cellular RNA was extracted from BHK cellsatthe indicatedtimes after infection. Wheatgermcell-freeprotein synthesis reactions(25Ll) containing 1,2, or4itgoftotal cytoplasmic RNAwereused,andduplicate5-,ulsampleswere
assayed foracid-precipitableradioactivity.For allRNAs,the amount of[3S]methionineincorporatedintoacid-precipitable radioactivity wasproportionaltotheamountof RNAadded; shown incolumn 2isthe totalamountof'S radioactivity incorporated in the entire 25-1lreaction. Incolumn3, the results are normalizedto themRNAactivityof RNAfrom uninfected cells. Each of these reactionswasanalyzed by SDS-gelelectrophoresis,andtheamountofN/NS and Mproteins wasdeterminedfrom the area under thepeakof the micro-densitometer scan of theradioautogram. These valuesare
normalizedtothe value characteristic of RNA from cells 2.5 h after infectionbyVSVGlasgow.
either Vero or BHK cells. At 3 h, synthesis of
actinin Lcellswas70to 100% ofthat of
unin-fectedcells, dependingontheVSV strain used.
In all ofourexperiments, shutoff of actin
syn-thesis afterinfectionof LcellsbyVSV Rlwas
slightlyless rapid than after infection by VSV
HR, althoughthis difference wasnotobserved
in thetwoother cell lines tested (Fig. 2 and8). Inallcases,the totalamountofproteinsynthesis
atanytime up to 5 h ofinfection was greater
than 90% that of uninfected cells (data not
shown).
Two typesof comparisonsindicated thatthe
shutoffofsynthesisofL-cellproteinwas
corre-latedwiththe levelof VSVmRNA. First, VSV
mRNA accumulated to a slightly lower level
after infection of L cells by VSV Rl than by
VSV HR (Table 2); this corresponded to a
slightlyslowerrate ofinhibition ofL-cellactin
synthesis by VSV Rl. Second, with all three
VSV strains tested (Glasgow, HR, and Rl),
much less translatable VSV mRNA
accumu-lated afterinfectionin L cells than inBHK cells (Table 2); this correlatedwellwith thereduced
shutoff observed in Lcells.Note that theamount oftranslatable mRNA (per milligram of cyto-plasmicRNA) recovered from uninfected Lcells
and BHK cellswasthesame and thatthe
pre-dominantspecies oftranslatable L-cell mRNA
also remainedfullyfunctional after VSV infec-tion(Fig. 11).
VSV ts mutants. As another approach to
investigating the role of VSV mRNA in
inhibi-tion of cellular proteinsynthesis,weusedtsVSV
mutants in each of the five complementation
groups (10, 13). All of the mutants used were
TABLE 2. Messenger activity in RNA from VSV-infected L cells and BHKceilsa
Total mRNA activity Relative amount of VSV mRNA Time after
Cells Virus infection [3S]methionine
(h) incorporated(105 Normalized N/NS M
cpm/igofRNA)
BHK None 2.60 1.00
VSVGlasgow 3 6.11 2.35 1.00 1.00
4 8.40 3.23 1.58 1.68
VSV HR 4 9.31 3.58 1.88 1.94
VSVRi 4 7.46 2.87 1.39 1.36
Lcels None 2.20 1.00
VSVGlasgow 3 3.26 1.48 0.46 0.41
4 4.36 1.98 0.86 0.79
VSVHR 3 4.03 1.83 0.68 0.64
4 5.02 2.28 1.03 0.97
VSVRi 3 3.32 1.51 0.53 0.44
4 4.49 2.04 0.99 0.96
aAnalysis of the total
cellular
RNAswas detailed in the legend to Table 1. Values for total mRNA activityarenormalizedtothatobtained with RNA fromuninfected BHK or Lcells.In all cases, the amount of viralN/ NS andMmRNAactivity is nornalizedto that obtained from RNAisolated3 hafterinfectionof BHK cells withVSVGlasgow.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.500.56.249.165.313.2] [image:7.500.60.454.467.631.2]IS THERE A P FUNCTION IN 511
VSV Son Juan
.3
c
2_
L1a,
VSV GlasgowUninfected
z
b
l
2 3 4 5 6 7 L 9 10 I 4VSV SanJuan
3-VSV Glasgow a V
0
o-'2
Uninfected w VS G oPolyribosome
i * Size
I I 1018 6 5 4 31 2 I1 5 10 15 20 25 30 35
Fraction
FIG. 5. Subcellular localizationofribosomes and translatable mRNA in growing BHK cells and in cells 3 h after infectionby VSVGlasgow and VSV San Juan.Asampleof6xplO BHKcellswasgrown for20hinthepresenceof1.5mCiofa'HJuridine (The RadiochemicalCentre)and harvestedby centrifuga-tion. One-thirdofthe cells were mock infected, and one-third each wereinfectedwithVSVGlasgow and VSV San Juan. Three hoursafter infection, a post-nuclearsupernatant fr-omthe cultureswasprepared
andcentrifuged througha 15 to30%(wt/vol)sucrose
gradient,asdetaiedpreviously(2).Shown inpanel b isthe 3Hradioactivityper50-ldportion offractions from thegradient ofthe extractfrom uninfectedor
infectedcells. The sizeofpolyribosomes was deter-mined by extrapolation oftheprofile ofan extract
fr-om rabbit reticulocytes analyzed in parallel. As
indicatedbythebrackets, 1.0mlfromthreeadjacent
fr-actions was pooled. RNA wats then isolated and
translated in a wheat germn cell-fr-ee system. Two differentamountsofRNAwereused,generallyabout 1.5 and3.0%ofthe total RNArecoveredfr-om each
derived fromtheGlasgow isolateofVSV. Three
of themutantsmade littleor no VSVmRNAat
39.50C,
the nonpermissive temperature:tsG114(I),
tsG22(ll);
andtsG41(IV).Infectionof BHK cells (Fig. 12) or Vero cells (data not shown) at39.5°Cbythesemutantshad noeffect at all on the rate ofsynthesis ofpredominantcellularproteins. Shutoffofcellularproteinsdid occur, by contrast, after infection by the two
RNA' mutantsused: tsG33(III) and tsL513(V)
(Fig.12).Thelowerrate of VSVprotein
synthe-sis intsL514(V)-infected
cells,
relativetoinfec-tion by the wild type, was a consequence of
increased cell death; at 3 h these celLs
synthe-sized
wild-type
levels of VSV proteins (14).Again, there was an excellent correlation
be-tweensynthesis ofVSV mRNA andinhibition
oftranslation of
cellular
mRNA. Acorollary ofthisfinding is that the infecting particles
them-selves,atleastup to alevelof 10PFU/cell,had
noeffectonmRNAtranslation.
DISCUSSION
Dependingontheisolate of VSV Indiana and
onthe host cell used, therearemarked
differ-ences in the kinetics of inhibition of cellular
proteinsynthesis. In all casestested, this
inhi-bition was manifest at the translational level:
theamountoftranslatablemRNA's for the
pre-dominant cellular proteins which can be
ex-tracted from the cells remainsunchangedafter
infection, whereas therateofsynthesisof
cellu-larproteins decreasessteadily.
Weemphasize again (2) thatwehaveusedin
vitrosynthesisof discrete viral and cellular
pro-teinsas ameasureof theamountof
correspond-ingmRNA in theinfected cells.Especiallyinthe
caseof VSVRNAs, thistechniqueis
preferable
to the morecommonly used
hybridization,
gel
separation,or isotopeincorporation techniques
toquantitateVSVmRNA,since itmeasuresthe
biologically relevant
parameter-its ability
todirect protein
synthesis.
Not allVSV-specific
polyadenylated RNAs are functional mRNA's,
and thus physical measurements canseriously
overestimate the amount of VSV mRNA. A
fraction. Within this range, protein synthesis was
invariablyproportionaltotheamountofRNAadded (cf.reference 2). Plottedontheordinateinpanelais thetotal amountoftranslatable mRNA perpooled
fraction:this is theproductofcpm
of[3SJmethionine
incorporated/cpm of
[3HJRNA
added and totalamountof
[3H]RNA
in thepooledfractions. Since recoveries of[3H]RNAwereverysimilarinall frac-tionsandaveraged85%,thecorrectionfordifferential recovery of[3H]RNA in the different fractions is small anddoesnotaffecttheposition ofanyofthe datapoints shownbymorethan10%.VOL. 38,1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.500.44.235.71.462.2]512 LODISH AND PORTER
c 0 0
.-LL
z
E
120 100
80
60 40 20
80
40
80
40
120
80
40
mRNA Fraction
l I I I 1
18 12 9 6 4.5 3 2 I RNP 18 12 9 6 45 3 2 I RNP
Polysome Size
FIG. 6. Subcellularlocalizationof specificcellular and viral mRNA. RNAfromdifferent fractions ofthe polysomegradientof Fig.5 wastranslated inawheatgermcell-freesystem,and theproductswereresolved byelectrophoresis through a10to15%gradientpolyacrylamide gel(cf. Fig. 3).In all cases, anamountof recoveredRNA,equivalentto0.76% ofthe total RNA present in the initialpooledfraction,wasaddedto a 25-ulcell-freereaction. Threedifferent exposuresofthegel ofthetranslationproducts werescanned witha Joyce-Loeblmicrodensitometer,andtheareasofthepolypeptidebandsindicated(cf. Fig. 3)weredetermined. A1-h exposurewasusedforthe VSV G and Mprotein; a24-or48-h exposurewasusedforcalculationof cellular bands.Theareas(in arbitrary units)shownontheordinatearenormalizedtotheequivalentofa 24-h exposureofthefilm; thus,the valuesforthedifferentpolypeptidesareproportionaltotheirrelativeextents
of synthesisin the wheatgermextract. The average sizeofthegradientfractions fromwhich theRNAwas isolatedwastakenfromFig.4.
significant fraction of normal VSV 13 to 18S polyadenylated RNAs possess the 5' termini
pppAandpppGininfectedcelLs. Thesearenot
foundonpolysomes andarepresumably inactive asmessengers (11).Incellsinfectedatthe
non-permissivetemperaturebytsmutantsdefective inthe Mprotein, there isavastoverproduction
of VSVpolyadenylatedRNAs(1, 5).Gel electro-phoresisandhybridizationstudies indicatethat they are indistinguishable from nonnal VSV
mRNA (1, 5). However, they are
undermeth-ylated andare notfoundonpolyribosomes (5).
Previously,westudied the mechanism of this
inhibition in BHKcellsinfected by the Glasgow
isolate of VSV Indiana (2). At 4 h the total
amountoftranslatable mRNA (viral plus cellu-lar) was about threefold that of growing cells.
Most species of cellularand viral mRNA were
localizedonpolysomes.Infection did result in a
reduction(about threefold)inthe rate of initia-tion oftranslationofallpredominantspeciesof
cellular mRNA, but translation ofcellularand
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.500.67.461.60.437.2]TABLE 3. Fractionofviraland cellular mRNA's in RNP particlesa
Fraction of mRNA
mRNA Uninfected VSVGlasgow- VSV San Juan-cells infectedcells infected cells
a 0.19 0.14 0.20
Y 0.07 0.14 0.30
( 0.18 0.20 0.29
0.16 0.18 0.29
Kc 0.15 0.31 0.26
A 0.17 0.34 0.28
N/NS 0.16 0.39
M 0.13 0.37
aThe
data in Fig. 6 were used to calculate the fractionof mRNA activity for each of six cellular and two viral mRNA's which is localized in the RNP faction(gradient fractions32 to37;pools11and12).viral mRNA'swhich encoded for thesame-size
proteins occurredon the same size of
polyribo-somes.We concludedthat the rates ofinitiation
ofprotein synthesisontypicalcellularandVSV
mRNA'sarethesame and that the large amount
ofviralmRNA synthesizedafterinfection com-peteswithcellularmRNAforaconstant number
of ribosomes. Our results are in disagreement
with the prediction of Nusset al. (9), who
con-cluded,onthe basis ofthe effects ofhypertonic
treatment ofcells, that VSVmRNA istranslated
inpreferencetohost mRNA. However, neither
themechanism of inhibition ofproteinsynthesis
by hypertonic treatment nor the relevance of
these results to mRNA translation in nornal
media isatall clear.
Thepresentresults extendourconclusionsto additional VSV strain-host cell combinations.
First, inhibition of cellular proteinsynthesis is
dependent on synthesis of VSV mRNA. After
infection at the nonpermissive temperatureby
VSV tsmutants defectiveinsynthesis of VSV
mRNA,therewas noinhibitionatall of
synthe-sisof cellularproteins (Fig. 12). Identical results
were obtained withmutantsdefectivein the N
(complementation group IV),NS (groupII), or
L(groupI)proteins. Mutants defectivein the M
orG protein, bycontrast,made closetonormal
levels of VSV mRNA and inhibited synthesis of
cellular proteins with normal kinetics (Fig. 12
and datanotshown).
Second, inBHKcells, threeof the four VSV
isolates tested [Mudd-Summers, HR
(Winni-peg),and
R]
inhibitedsynthesisofcellularpro-teinsatabout thesame rate asdid theGlasgow
strainofVSVIndiana.Similarly,theamountof
VSV mRNA accumulated after infection by
these strains was approximately the same, as was the rate of VSV protein synthesis in the
infected cells (Fig. 1). Bycontrast, infectionof
BHK cells by the San Juan isolate of VSV
resulted ininhibition of synthesis of allcellular
proteins, and actin inparticular, about45 to60
min earlier than did infection by the other
vi--N/NS
-actin
-M
78 9 10 11 12 13 14
!
C,2
e5
6;Z
A-
CFIG. 7. Gel analysis ofproteins synthesized in Verocellsinfected bydifferentstrainsofVSV. Sam-plesofinfectedormock-infected Vero cellswere la-beledfor30minwith[3S]methioninebeginningat2 h(odd-numberedlanes)or3h(even-numberedlanes) after infection. Analysisofthe samples was as de-scribed in thelegendtoFig.1.Inparenthesesisgiven the total amount of
[OSJmethionine
incorporated intoprotein3hafterinfectionrelativetothe valueofuninfectedcells. (1 and2) Uninfectedcells(1.00); (3
and 4) infected by VSV Glasgow (0.90); (5 and 6) infected by VSV San Juan(0.85); (7and8) infected byVSVMudd-Summers(0.81);(9and10)infectedby VSVHR (0.81);(11and12)infected byVSVR1(0.91). 38,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.500.250.442.124.547.2]00
I1
ruses (Fig. 2 and 4). This inhibition wascorre-tXVero CelIlIs latedwithan increase in therateof
accumula-VtO Cellsdtionof VSVmRNA and inanaccelerationof the
rateofVSVproteinsynthesis: thesameamount
80L
ofVSVmnRNA
was accumulated inthe cell45min sooner after infection by VSV San Juan
thanbyVSVGlasgow. (Thehistoryof the
lab-oratorystrain of VSV SanJuan is highly rele-vant[A. Huang,personal communication]. The
60\ strain, as originally obtained in 1963 from the
American TypeCulture collection, formed
rela-tively smallplaques andyieldedastock ofvirus
of about 108 PFU/ml. To select for a variant
ICL| \ \ \\ _ which grew to ahigher titer, Dr.Huangpassaged
40\ R thevirusinCHOcells17timesinsuccessionat
RI
\\tlow multiplicities of infection. After this, the
HR virus was cloned repeatedly. The yield of
infec-tious virus in a
single-step
growth
curve was20 MS increased about 10-foldby this enrichment
pro-GL cedure, and it is apparent thata more rapidly
Si replicatingvariant [at least in some cells]was
selected. The differences between this isolate
and the other VSV wild-type strains is most
1 2 3 pronounced in the BHKcellline.) The effects of
Ho u r s this increased amount of inRNAonthe
[image:11.500.59.254.60.327.2]subcel-lular distribution ofcellular (and viral) mRNA
FIG. 8. Synthesis of actin in Vero cellsinfectedby are incomplete accord with our
conclusion
that different strains of VSV. The gel profile shown in viraland cellular mRNA'scompete on anequal Fig. 7 wasanalyzedasdetailed in Fig.2.Plotted is basis during the initiation step ofprotein syn-theamountof radioactivityin the actinpeptide, rel-..
.
a native to thatofmock-infected cells, at several times thesis for a
limiting
number ofribosomal sub-after infection. Although not shown, there is no unts. At 3 h after infection by either virus, at change in the rate ofactin synthesisIh afterinfection least 75% ofcellularribosomesremain in poly-by any VSVstrain. Symbols are as in Fig. 2. somes,andthetotalrateof totalproteinsynthe-L Cells
M
N/NS ctin
2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
hr | 2hr 3hr | 4hr | 5hr
FIG. 9. Gelanalysis ofproteins synthesized in L cells by different strains ofVSV.Samples of mock-infected orVSV-infected cellswerelabeledfor 30 min with[3S]methioninebeginning at1h(lanes1-6),2h (7-12), 3 h (13-18),4h (19-24),or5h (25-30)after infection. Total cellular protein was analyzed byelectrophoresis througha10%polyacrylamide gel containing SDS; shown is a radioautogram of the dried gel. (Lanes 1, 7, 13, 19,21,25,27)Uninfected cells; (2, 8, 14, 20,26) infected by VSV Glasgow; (3, 9, 15) infected by VSV San Juan; (4, 10, 16, 22, 28)infected by VSV Mudd-Summers; (5, 11, 17, 23, 29) infected by VSV HR; (6, 12, 18, 24,30) infected by VSV RI.
J. VIROL.
514 LODISH AND PORTER
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.500.65.456.419.599.2]IS THERE A P FUNCTION IN VSV? 515
L
G N/NS -actin
-M
[image:12.500.50.243.74.240.2]2 3 4 5 6 7 8 9 10 11 12
FIG. 10. Cell-free translation products of RNA from VSV-infectedLcells.Proteins were synthesized inawheatgermcell-freesynthesisreaction, using 1 pgoftheindicated RNAs per25-,ilreaction. A
5-jtl
sampleof the reactionwasanalyzed on a 10 poly-acrylamide gel containing SDS. Table 2 presents the total amountofacid-precipitableradioactivity incor-porated perreaction,aswellasthe relative amounts of VSVproteins produced. (Lane 1) No RNA; (2) RNAfrommock-infectedLcells,3h;(3)RNAfrom VSVGlasgow-infected L cells, 3 h; (4) RNA from VSVHR-infectedLcells,3h; (5) RNAfrom VSVR1-infected Lcells, 3h; (6) RNAfrommock-infected L cells, 4 h; (7) RNA from VSV Glasgow-infected L cells,4h;(8) RNAfrom VSVHR-infectedLcells,4 h; (9) RNAfrom VSVRI-infected Lcells, 4h; (10) RNAfromadifferent preparation ofVSV Glasgow-infectedL cells,4h; (11)RNAfrom VSV Glasgow-infected BHKcells,4h; (12)RNAfrom VSVGlasgow-infected BHKcells,3h.sis is atleast 80% that of
giowing
cells. Thus,the vastmajority of ribosomesarefunctioning
in protein synthesis. However, the averagesize
of polysomes translating cellular and viral
mRNA's ismuchsmaller intheVSV San
Juan-infectedBHKcells.After infectionbyVSV
Glas-gow,mRNA's fortwotypicalcellularproteinsof
molecularweight of 30,000to40,000, 0 andactin,
are found to be pentasomes, the same size as
polysomeswhicharetranslatingVSV N(52,000)
and M (35,000) mRNA. This is 40 to 50% the
size ofpolysomeonwhich isfound actin mRNA
in growing cells, a result consistent with the twofold increase intotal amount of functional mRNA per cell. In the VSV San Juan-infected
cells, by contrast, actin, 0, N, and M mRNA's
arealllocalizedtodi-ortrisomes-one-sixth the
number of ribosomes per actinor0 mRNA found
ingrowing cells.Thiscorrespondstothe five-to
sixfoldincrease in total translatable mRNA per cell observed in thisexperiment.
Although ourresults on infection ofmouseL
cells by VSV are different in several respects
fromthoseusing BHK cells, theyareconsistent
with the notion that host shutoff is a
conse-quenceofmRNAcompetition for limiting
ribo-somes. First, for all VSV strains studied the kinetics of hostshutoffareslower in L cells than inBHK cells:about1hin the case ofinhibition
oftranslationofactin mRNA. Thisisparalleled
by a correspondingly slower rate of
accumula-tion ofVSV mRNA. Inparticular, the amount
ofVSV mRNA found3 h afterinfection of BHK
cells with VSVGlasgowis about the same as at 4h afterinfectionof L cells by thesamevirus
preparation (Table2).
Anadditional correlation emerges from a
com-parisonoftheinfectionby L cells by VSV strains
HR4 and Rl. AsStanners et al. (12) reported,
inhibition of cell protein synthesis isless after
Rlinfection than after HRinfection. However,
theconsistentdifferencewefind(Fig. 11) isnot
as marked as that previously described (10).
Nonetheless, thereducedlevel of VSVmRNA
which accumulates by 3 h in Rl-infected cells
relativetoHR4-infected cells correlates with the
reducedinhibitionofactinmRNAtranslation.
Stannersetal. (12)foundamuchmorerapid
inhibition ofL-cell protein synthesis after
infec-tion by the HR strain thanwe have observed.
Since we have used cell linesand virus grown
fromstocksprovidedby C. P.Stanners,wehave
L Cell s I0oo
80k
r-60
a)
a. H
401-
201-1 2 3 4 5
Hou r
FIG. 11. Synthesis of actin in L cells infected by four strains of VSV. The gelradioautogramof the gel in Fig.9 wasanalyzedasdetailedin thelegendto Fig.2.Symbolsarealsodescribed there.
38,
HF
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.500.252.448.378.627.2]234 5 67 8
b~~\/ V
'V
5
no explanation for this difference, except that
we are culturing thecells asmonolayers. Also, in allofourstudieswe takecare that the cells
aregrowinglogarithmicallyatthetimeof
infec-tion; thephysiologicalstateof the host cellcan
L affect the response to VSV infection (12). We
have confirmed the finding of Stanners et al.
that inhibition ofL-cellprotein synthesis is less
rapidafter infectionby strain Rl thanby HR. However, wequestion their assumption that Rl
is, in fact, a single-step mutant derived from
strain HR, and thus the conclusion that the
differences in kinetics of shutoff of L cells
be-tweenHR4 andRl aredue toamutation in a
single viralgenefunction, P. Recall that Rlwas
isolatedas atemperature-resistant revertantof
tsT1023(I), which issupposedtobea mutantof
strain HR. We havefoundconsistently that the
Rl Mprotein migrates 10% fasteronSDS-gels
thandoes the HRMprotein. This istruefor M
proteinssynthesized inBHK, L, and Vero cells
(Fig. 1, 7,and 9). It is also thecasefor HR and
-0 Rl M
proteins
synthesized
inawheatgerm
cell-free system (Fig. 10). It isdifficult to imagine
howasingle-step mutation could result insuch
alarge shift ingelmobility.Clarification of this
point willrequireisolation of P- mutations
di-rectly fromaknownwild-type strain.
- N / N S We do not know, in detail, why the kinetics of
inhibition of cellular proteins synthesis varies
-ac
ti|
n with the host cell and with the strain of VSVused. Thereis,asnotedabove,agoodcorrelation
between the kinetics of host shutoff and the
accumulation of viralmRNA, but it isnotclear
why the rate of accumulation of viral mRNA
should besodependentonthe host cell line and
:M on the strain of VSV. Our preliminary studies
indicate that the inhibitionof actin synthesis 3
hafter infection ofBHKcells by10 or 50 PFU/
cellisnotsignificantly(10%)different,so
differ-entialadsorptionorpenetration ofthevirionsis
unlikelytobeinvolved. As noted in thetext, in all cases all of the cells are infected by VSV.
Whereas a VSV virion contains its own RNA
polymerase activity which will, in vitro, direct
synthesis of all five VSV mRNA's, it is not
known whether, in the infected cells, specific
[image:13.500.69.253.58.603.2]host proteins (which might be present also in the virionin minute amounts) are also essential, nor is it known how the intracellular levels of
FIG. 12. Gelanalysis synthesisofproteins synthe-sizedinBHKcellsinfectedat40.0°Cbytsmutantsof VSV. As inFig. 1, cellswereinfectedby the indicated mutantsandlabeled with[3S]methionine4hafter infectionat40°C. Electrophoresisofthe labeled pro-tein utilized at 10 to 15%gradient polyacrylamide containing SDS.(Lanes1and2) Uninfected cells; (3) infection bywild-typeVSVGlasgow;(4) infection by tsG114 (I): (5)infection bytsG22(II); (6)infection by tsG33(III);(7)infection by tsG41(IV); (8)infection by tsL513(V).
516
on November 10, 2019 by guest
http://jvi.asm.org/
A P
ribonucleoside triphosphatesorSAdoMetinthe
cells might influence therateof viral RNA
tran-scription.
Taken together, our results indicate that in-hibitionof cellularprotein synthesisisprimarily a consequence ofcompetition by viral and
cel-lular mRNAfor a constant limiting number of
ribosomes. Shutoffclearlyisdependenton
syn-thesis ofviral mRNA. However,ourdataonthe
subcellular distribution of mRNA's after
infec-tion andonthecorrelation between the levels of
viral mRNA and inhibition of cellular mRNA
translationareonly semiquantitative. It is
diffi-culttoeliminaterigorously other factors which
could reduce the rateoftranslation ofcellular
mRNA's after infection. Previously we listed
someof these: (i) reduction in therateof
poly-peptide chainelongation, (ii) death ofacertain
fraction of cells, and (iii) sequestration into
RNPs of a significant fraction (about 25%) of
cellspecies of cellular and viral mRNA's. These
mRNA's could be localized only to the dead,
inactive cells. Whatever the mechanismfor
in-crease inthelevel of RNPs afterinfection, it is
significant that there is no difference in the
extentofsequestration oftypicalviral and
cel-lular mRNAspecies.
ACKNOWLEDGMENT
Thisworkwassupported byPublicHealthService grant AI-08814-13from the National Institutes of Health.
LITERATURE CiTED
1. Clinton, G.M., S. P.Little,F. S. Hagen,and A. S. Huang.1978.The matrixproteinof vesicular stomatitis virusregulatestranscription. Cell15:1455-1462. 2. Lodish,H.F.,and M. Porter. 1980. Translational control
ofproteinsynthesisafter infectionbyvesicular stoma-titis virus. J. Virol. 36:719-733.
3. Lodish, H. F., and R. A.Weiss. 1979. Selective isolation of mutants of vesicular stomatitis virus defective in production of the viral glycoprotein. J. Virol. 30:177-189.
4. Marcus, P.I., M. J.Sekellick, L Johnson, and R. A. Lazzarini.1977. Cellkillingby viruses. V. Transcribing defective interfering particles of VSV function ascell killing particles. Virology 82:242-246.
5. Martinet, C., A. Combard, C. Printz-Ane, and P. Printz. 1979. Envelope proteinsreplicationof VSV: in vivo effects ofRNA' temperature-sensitive mutations on viral RNAsynthesis. J.Virol. 29:123-133. 6. Marvaldi,J., J. Lucas-Lenard, M.Sekellick,and P.
I.Marcus. 1977. Cellkilling by viruses. IV. Cell killing and protein synthesis inhibition by VSV require the same gene functions. Virology 79:267-280.
7. Marvaldi, J., M. J. Sekellick, P. I. Marcus, and J. Lucas-Lenard. 1978. Inhibition of mouse L-cell protein synthesis by ultraviolet-irradiated VSV requires tran-scription. Virology 84:127-133.
8. McAllister, P. E., R. R. Wagner. 1976. Differential inhibition of host protein synthesis inL-cells infected with RNA- temperature-sensitive mutants of vesicular stomatitis virus. J.Virol.18:550-558.
9. Nuss, D. L, H. Opperman, and G. Koch. 1975. Selective blockage of initiation of host protein synthesis in RNA-virus infected cells. Proc. Natl. Acad. Sci. U.S.A. 72: 1258-1262.
10. Pringle, C. R. 1977. Genetics of rhabdoviruses, p. 239-290.In H. Fraenkel-Conrat and R. R. Wagner (ed.), Comprehensive virology, vol. 9. Plenum Publishing Corp., New York.
11. Rose,J. K. 1975. Heterogeneous 5'-terminal structures occur on vesicular stomatitis virus mRNAs. J. Biol. Chem. 250:8098-8104.
12. Stanners,C. P., A. M. Francoeur, and T. Lam. 1977. Analysis of a VSV mutant with attenuated cytopatho-genicity: mutation in viral function, P, forinhibitionof protein synthesis.Cell11:273-281.
13. Wagner, R. R. 1975. Reproduction of rhabdoviruses, p. 1-94. In H. Fraenkel-Conrat and R. R. Wagner, (ed.), Comprehensive virology, vol. 4. Plenum Publishing Corp., New York.
14. Zilberstein,A., M. D. Snider,M.Porter, and H. F. Lodish. 1980. Mutants of vesicular stomatitisvirus blocked at different stages in maturation of the viral glycoprotein. Cell 21:417-427.
on November 10, 2019 by guest
http://jvi.asm.org/