Copyrighti 1974 AmericanSocietyforMicrobiology PrintedinU.S.A.
Viral
Proteins Formed
in
aCell-Free
RABBIT Reticulocyte
System
Programmed with
RNA from
aTemperature-Sensitive
Mutant
of
Sindbis
Virus
RANIERI CANCEDDA,I RICHARD SWANSON, AND MILTON J. SCHLESINGER
Department of Microbiology, Divisionof Biologyand Biomedical Sciences, Washington University, St.Louis,Missouri63110
Received for publication 29April 1974
Viral messengerRNA was isolated from BHK cells infected with a
tempera-ture-sensitive mutant of Sindbis virus and was further purified using an
oligo(dT) column. Addition of this mRNA cell-freeextracts from rabbit reticulo-cytesledtoformation ofdiscrete authentic viral capsid protein when thereaction
wasperformedat 29 C. However,thissameprotein-synthesizingsystemfailedto
make discrete viralcapsid when incubated with the viral RNAat39C. Instead, larger-molecular-weight polypeptides that contained the viral capsid peptide sequenceswere produced. The inabilityto makeaseparateviral capside protein in vitroatelevatedtemperaturesby the mRNA fromthismutantexactlymimics
the phenotype of this ts mutant in viral-infected cells. Three mechanisms are
discussed that might accountforatemperature-sensitive release of capsid. One of these is based on a model in which there are multiple sites for initiation of translation ofpolypeptides ona polycistronic viral mRNA.
Among a variety of temperature-sensitive
mutantsofSindbisvirusthat have been charac-terized are a group that fails to make virion proteins atthe nonpermissivetemperature even though viral RNA formation appears normal (3). Extracts of cells infected with these mu-tants at 40 C show the presence of a large polypeptideofmolecular weight -130,000 that
is not detectable at lower temperatures or in
cellsinfected with wild-typevirusatthehigher temperature (18). Tryptic peptide fingerprints of this large protein clearly indicated that it
contained those amino acidsequences that are
normallyfound inthe three individual Sindbis
virion proteins (20). Thus, the mutational de-fect prevented the normal processing of the
Sindbis'viral protein. Primarily onthe basisof
thephenotypeof thiskindof temperature-sensi-tive mutant, investigators studying Sindbis
virus formation have postulated that the
nor-mal pathway of synthesis of virion proteins consists of translation from a polycistronic
mRNA in which initiation ofsynthesis occurs only atthe 5' terminus of thenucleic acid and
the individual proteins arise by a
post-transla-tional proteolytic cleavage (16, 18). Otherdata from analysis ofRNA and proteins in Sindbis
IPresentaddress: 2ndFacultyofMedicine,2nd Institute
ofBiochemistry, UniversityofNaples,Naples,Italy.
virus-infected cellsgenerallyareconsistent with
this model if one presumesthat theproteolytic
cleavagesoccurduring translationof theprotein onthepolyribosome. Furthermore, there is now
substantial evidence that this model describes
the mechanism for viral protein synthesis in
cells infected withpicornaviruses (5, 10, 11, 23).
We initiated studiesontheinvitroformation of Sindbis viral proteins in cell-free extracts
programmed with viral RNAs obtained from
Sindbis-infected BHK cells in order to obtain evidence for posttranslation proteolysis and to
gain additional information about Sindbis viral protein synthesis. Thus far, wehaveshown that the addition of a 26S species of viral RNA to
rabbit reticulocyte extracts leads to formation of the viral capsid protein, but therewas little
or no evidence for an involvement ofa
proteo-lyticenzyme in theprocess(6, 7). Furthermore,
there were only limited amountsoftranslation
other than the capsid cistron, although varying
the components of the rabbit reticulocyte sys-tem aswellasthesourceof the viral RNA does
allow for larger-molecular-weight proteins to
form. We expected that in vitro translation of themRNAfromatemperature-sensitive mutant
unable to make capsid or discrete
envelope
protein at the nonpermissive temperaturewould provide further insight into our under-standingofSindbis viral protein
synthesis.
664
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
Chemicals. Pactamycin was generously provided by D.Rekosh, MassachusettsInstitute ofTechnology. Itwaspreparedasamillimolarsolution inmillimolar acetic acid anddilutedtogiveafinalconcentrationof
3x 10-1 M in the reaction mixture. Guanidinewas an
ultrapurepreparationfromSchwartz-Mann (Orange-burg, N.Y.); ureasolutionswereshakenwith Amber-lite MB-3(Mallinckrodt Chemicals, St. Louis, Mo.) (10 g/liter of4Murea) beforeuse.Allotherreagents, including [35S ]methionine and [3HJuridine, were identicaltothose describedpreviously (6).
Virus and cell preparations. The ts-2 Sindbis
virus mutant wasprovided by Boyce Burge,
Massa-chusetts Institute of Technology. Virions were iso-lated fromculture fluidsofBHK-infectedcellsgrown in roller bottles at 30C. Purification of virus and isolation and purification of RNA from virus and viral-infected cells were identical to the methods described (6, 7), except that infected cells were harvested after 13.5 h at 30 C. A profile of the electrophoretic mobilityinanacrylamide-agarose gel
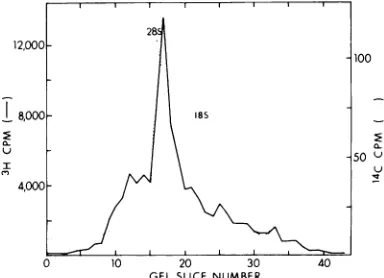
of themRNAused in this work ispresentedinFig.1.
Invitroproteinsynthesissystemandisolation of products.All in vitroexperimentswith RNA from the
ts-2 virus and viral-infectedcellswereperformed with
the cell-free system II prepared from lysed rabbit
reticulocytes. Details of thispreparation, the compo-nentsof thereactionmixture, and the analysisof the proteins formed are described elsewhere (6). For preparation of the tryptic peptide fingerprints, sam-ples from the reaction mixtures were precipitated directlywith 9 volumes ofcoldacetoneafterreduction andalkylation, andnodialysiswasperformed.
RESULTS
Effect of temperature on in vitro protein
synthesisby viral RNAs. RNAspartially
puri-fied from BHKcells infected with either the ts-2 Sindbis virus mutantorwild-type virus
stimu-12,000
1100
78,000 as18
50
x~~~~~~~~~~~~0u
In
GELSLICE NUMBER
FIG. 1. Partiallypurifiedpreparation of viral RNA
from BHK cells infected with ts-2at 30C.
Electro-phoresis was performed on a 1.8% polyacrylamide,
0.5% agarose gel (24). Symbols: -, 'H counts per minuteof viral RNA;...1Ccountsperminuteof18S
and28Sribosqmal marker RNA.
lated the incorporationof [35S]methionine into
protein incell-freeextractsfromrabbit reticulo-cytes(Table 1).Stimulationby added RNAwas
observed inreactionsincubated at 29and39 C, andtheamountof35S-labeled protein formedat
the two temperatures was not significantly
different. For reactions containing the ts-2
RNA, however, thepattern ofproteins revealed
in sodium dodecyl sulfate
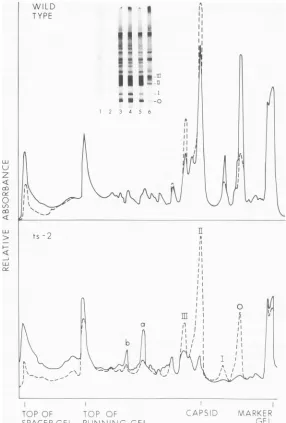
(SDS)-polyacryl-amide slab gels formedatthetwotemperatures was noticeably different (Fig. 2). We have shown previously that band II is identical to
virion capsid protein and noted that bands I
and III contain capsid peptides (6). Both the gel autoradiogram and densitometric tracings of reactions at 39 C containing ts-2 RNA showed a large decrease in the amount of
capsid and asignificant increase in two
higher-molecular-weight bands (Fig. 2aand b) and in
material that stayedatthetop ofthe5%spacer gel. Some variations were also observed in
capsid polypeptides formed by wild-type viral RNA withmoreofthe smallerproteins appear-ing. The shift to larger-molecular-weight
pro-teinsin incubations at 39 C with the ts-2 RNA was confirmed by an analysis of the reaction mixtures after rate-zonal centrifugations in a urea-sucrose gradient (Fig. 3).
Analysis of protein formed by ts-2 RNA at 29and39 C.Despite the marked disappearance of capsid polypeptides from the 39-C reaction mixturecontainingts-2RNA,wecould show by
apeptidefingerprintofatrypticdigestionofthe total reaction mixturethat thecapsidcistron of
ts-2 RNA was translated (Fig. 4). In fact, the fingerprints fromthe29- and39-Cmixtures are indistinguishable and both show nine peptides that are absent from the fingerprint of the reaction mixture containing no viral RNA. Of the nine, sevencanbe relatedtopeptidesofthe isolated viral capsid (Fig.4C). Thetwo intense spots in fingerprints A, B, and D ofFig. 4 are
TABLE 1. Stimulationof35Sincorporationinto
protein
35Sincorporatedinto
Speciesof RNA gaddedto protein(X
10'3
SpeciesofRNA reactiona counts/minm)
29C 39C
Wild-type viral 13 123.2 134.8 ts-2viral ... 15 113.7 102.3
None 63.0 56.0
aTotal volume of reactionmixture was 50
Mliters,
andreaction was stopped after 60min. Components of reaction mixture are noted in ref. 6.° Measured on 5Mlitersofreaction mixture.
14,1974
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.41.233.463.602.2] [image:2.493.247.439.513.618.2]SCHLESINGER
WILD a I
TYPE
su
--i4 _5 z
v : I f!
a-I
I_
aa*t_
-, 4 4 6
AI II II
II
-o
ii
II
11
ts~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
I
l~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~I l l~~~~~~~~~~~~~~~~~~~~~~~~I
<
=
X< _ 1z, X f
>~~~~~~~~~~~i
CF TL'. KEP
SPACER OEL RUINN'NOOGEL
G--FIG. 2. Patternof proteinsformed invitro with wild-type andts-2 viralRNAs.Samples aredescribedin
Table 1.Insetshows autoradiogram of slab gel electropherogram.Samples 1 and2,No addedRNA;3and4,
wild-type viral RNA;5and6, ts-2viral RNA. Samples1,3,and5wereincubatedat29C;samples2, 4,and 6
were incubated at 39C. Autoradiogram was scanned in a Gilford gel-scanning device to provide the
densitometertracingsshown.Upperpanel:...Sample3,wildtype,29C;-,sample4,wild-type,39C.Lower
panel:. Sample5, ts-2, 29C;-,sample6, ts-2, 39C.
believed to be unincorporated [35S]methionine and methionine sulfoxide. The reaction
mix-tures were not dialyzed before acetone
precipi-tation and digestion, and not all the
unincor-porated [35S ]methionine was removed by this
procedure.
Characteristics of the ts-2 mutation. The
data from the experiments described above
suggest thatat leastoneapparent defect in the
translation ofts-2 RNA is an inability to form discrete capsid polypeptides. One possible
ex-planation for this is that the ts-2 capsid is
abnormalinstructureandat39 Caggregates to
higher molecular-weight forms.To testthis, we
treated the reaction mixtures with 5 M guani-dine hydrochloride immediatelyafter the reac-tion and displayed the products on a
sucrose-ureagradient.Theresultswereidenticaltothat
666
J.VIROL.LLJ
Lu
z
co
Uz
QlY
0 Uf) co
LuJ
LBJ
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.118.404.72.495.2]noted in Fig. 3. Wealso used slab gels contain-ing 4M urea and0.1% SDS forelectrophoresis
as well as heating samples to 100 C in SDS before loading in the gels. In no casecould we
observe the capsid or the capsid-like protein bands in the 39-C reaction mixtures with ts-2
RNA.
The ts-2 mutation doesnot affect the initia-tion ofprotein synthesis onthe basis ofresults with pactamycin, an inhibitor of initiation of polypeptide chains in eukaryoticcells(8).
Addi-tion ofpactamycin 2minafter initiationat29C
followed byanimmediate shiftto39 Cgavethe
same amount of inhibition as noted when the
incubation was contained at 29 C (Table 2). A slab gel electropherogram of these reaction mixtures showed that capsid was absent from the 39-C incubation but present after a 29-C
incubation. Thus, the ts-2 defect was still ex-pressed even though initiation of synthesis had been inhibited at39C. The initiation of synthe-sis by ts-2 RNAat 39 C did not inhibit subse-quent translation at 29 C (Table 2), and a slab gel electropherogram showed a strong capsid protein band.
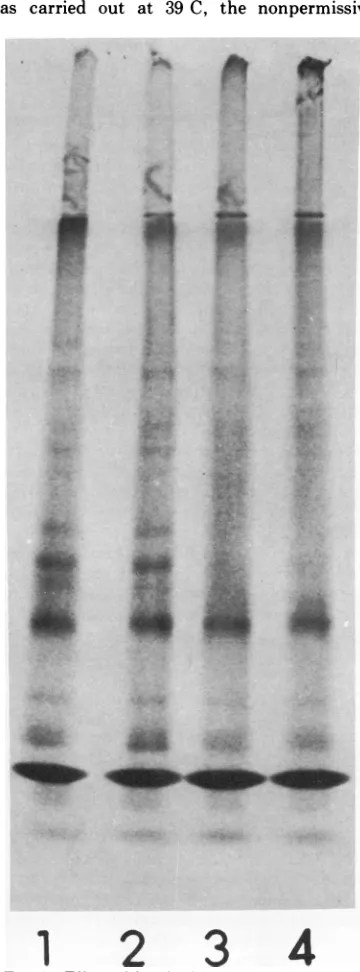
Evidence for a protease activity. In an experiment designed to look for an unusual
aggregation of the ts-2 polypeptides at higher
temperatures, areaction mixture that had been incubated for 60 min at 29 C was treated with RNaseto stop synthesis and placed at39 C for
an additional 60min. An SDS-polyacrylamide
gel pattern of this reaction mixture showed a disappearance of the faster-moving capsid-related band ILL (Fig. 5), suggesting aprotease activity in therabbit reticulocyte cell-free sys-tem. The protease seemed to be relatively specific in that capsid and the lower-molecular-weight band appeared stable. Wearecurrently
investigatingthisproteaseactivitytodetermine whether it is viralspecificand whether it playsa meaningful role in the Sindbis virus protein formation invitro.
DISCUSSION
The expression of the ts-2 mutation by viral RNA in a cell-free protein synthesis system is shown by results presented heretobe remarka-bly similar to the phenotype of ts-2 virus-infected cells. In both cases, discrete viral
capsid protein virtually disappears at the
non-permissive temperature and larger, abnormal polypeptides accumulate. The largets-2protein
found in vivo contains aminoacidsequencesof
the viral capsid (20), and the tryptic peptide
fingerprints presented in this work show that
the polypeptides made in vitro from ts-2 RNA at 39 C also have capsid peptides. Thus, the
aE-u
n
FRACTION NUMBER Top
FIG. 3. Rate-zonalcentrifugationin aurea-sucrose
gradient of proteins formed in vitroby ts-2 viral RNA.
Arrowrefers to theposition ofE. coli
beta-galactosi-dasesubunit (molecularweight = 135,000). Symbols:
-, 29-C reactionmixture; ...39-C reactionmixture;
- -, endogenous reaction mixture at 39 C with no
addedRNA.
mutational defect in ts-2 blocks formation of a
discrete capsid protein. We can propose three
ways in which this might happen: (i) an amino
acid substitution resulting from a missense
rnutation distorts the growing polypeptide chain sothatthesite forproteolytic cleavageto
produce capsid protein is blocked; (ii) the mutation causes theprotease required for
cap-sid appearance to become temperature
sensi-tive; (iii) the mutation alters the viral mRNA structure so that the translation ofcapsid cis-tron becomes directly linked to the translation
ofthe viral envelope cistrons.
Our datapresented here and elsewhereonthe invitro formation ofSindbis viral proteins can
be interpreted to support each of the above proposals. Furthermore, the in vivo data that
has been accumulated in several laboratories
from experiments on Sindbis virus replication
are unable to clearly distinguish between these three hypotheses. We cite here the arguments
pro and con for the three possible modes of action ofthe ts-2mutation.
(i) A defective misfolded polypeptide chain
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.250.437.64.355.2]'4
A
I
I'f
1'
*S
B
2
4
6
9
0 #6
8
J7
10
3
4
5
p0
S
0
C
2
4
i
'44
D
$
5
"aa
FIG. 4. Tryptic peptide fingerprints. (A)Incubation with ts-2 viralRNA at29C; (B) incubation with ts-2
viral RNAat39C; (C) isolatedcapsidproteinbandfrominvitro reaction withwild-typeviral RNA incubated
at30C; (D)reactionmixture withnoadded viralRNA,incubationat39C.Thesameamountof[35S]peptides (-106counts/min) wasusedforfingerprints A, B,and D.Sampleswereappliedatthelowerleftcornerofthe
papers; electrophoresis was in theupwards direction at pH3.5 followed by descending chromatography in
butanol-acetic acid-water(2:0.5:2.5) (totheright in eachpanel). Seereference20foradditional details.
(9
6
87
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.54.458.77.593.2]TABLE 2. Effect ofpactamycinonin vitro translation of ts-2 RNAa
Incubation temp
RNA Pretreatment (C)
29 39
Wild type None 142.9 89.4
ts-2 None 132.6 102.1
ts-2 2.5minat39C 134.2 NDb
ts-2 Pactamycinat0min 7.9 ND ts-2 Pactamycinat2min 36.1 ND
at 29C
ts-2 Pactamycinat2min ND 34.8 at29C, then shift
to39 C
aFigures are 35S counts per minute incorporated
into protein after 60 min of incubation; they have been multipliedby10-3.
bND, Notdone.
thatcannqa 1eardtQjjrionproteins. This
modelexplains t invivodata
showing7that
a ts-2 protein cannot be chased into virionpro-teinswhen infected cellsarelabeledat39C and shiftedto 29 C (18). It also canaccount forthe disappearance of the shorter capsid-related
polypeptides madeinvitro (bands0and1,Fig.
2) if weassumedthey areformedbya protease action.Furthermore,therearenumerous
exam-ples of missense mutations in which single
amino acidsubstitutionscanproduce profound
changes in a protein's conformation (19).
How-ever, a misfolded precursor polypeptide cannot
readily explain the ability of ts-2 mutants to complement Sindbis virus mutants with defec-tive envelope proteins (2). According to this model, a ts-2 viral RNA could not make a
discrete virion envelope protein at 39C, and thus a ts-2 mutant should not be able to
complementmutantswith alteredenvelope
pro-teins.
The model also predicts that, in vitro, the capsid anditsrelatedpolypeptidesarereleased
at the permissive temperature by a protease. Although we have presented data here for a proteaseactivity, ourearlierdata failedtoshow
aneffectoncapsid formation when the protease
inhibitors phenylmethane sulfonyl fluoride and
L-1-tosylamide-2-phenylethyl chloromethyl ke-tone (TPCK) were present in high concentra-tions (6).
(ii A
teMDerature-sensitiv'-
uiral
-codedp)ro-ea A ts-2 mutationin aproteasecan explain e in vivocomplementation noted above, and
it can also explain why none of the shorter capsid-related polypeptides appear at 39C in vitro. Butthis model predictsthat thisprotease
is aproduct of the in vitrosynthesisat29 C. We
have noevidencethat the protease observedin our system is viral coded. Furthermore, the reactiondemonstratingthisproteolytic activity
was carried out at 39 C, the nonpermissive
i.
;.4.
-4,
J
Iik
1. *:FIG. 5. Effect ofincubation at 39C onts-2 prod-uctsformed at 29C. After60min at29C, reaction mixtureswereprovided with 5,ilitersofRNase A (50
I.g/ml)and incubated60minat39C. (1) Wild-type
RNA at 29C; (2) ts-2 RNA at 29C; (3) wild-type RNAat29 C +60minat39 C; (4) ts-2 RNA at 29C +60minat39C.Equal amounts of 3"S radioactivity wereadded to each slot of the gel.
',Y.
C...'.) .1
0004...
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.255.435.106.592.2]CANCEDDA, SWANSON,
temperature for ts-2. The appearance of the cant change in the levels of viral envelope
different capsid-related polypeptides in vitro proteins (24).
(bands I and III, Fig. 2) was affected mostly by But the suppression of normal termination by the relative efficiency of the protein-synthesiz- ts-2fails to account forcomplementationforthe ing system with longer polypeptides formed same reasons citedabove, unlessone proposed
when moresoluble cellularenzymes wereadded that a low level ofinternal initiation occurs at
(7). This result is not easily explainable by a the envelope proteincistrons of the ts-2 mRNA protease model. at39 C.Toexplain the disappearanceinvitro of
(iii) in the the lower-molecular-weight capsid-related
pro-tr iNtire.
lthugmolst
oft Temper- teins by ts-2 RNA at 39 C, we would suggest re-sensitive mutations examined lead to al- that these latter proteins all terminate in theterations in protein structure, there is at least same manner but that they initiatetranslation
one report ofatemperature-sensitive mutation within the cistron. There is good evidence that
t in an RNA. A single base changein the tyrosyl initiations within a cistron can occur in bacte-tRNA of Escherichia coli affects the function of rial mRNA (14). Noevidenceisavailable thata
thetRNA athightemperature(22). The postu- similartype of reaction occurs inmRNA'sfrom lated change here in the viral mRNA would eukaryotic cells; however, Morrison and Lodish suppress a normal termination signal and per- (15) showed that extracts from eukaryotic cells mit continued translation. The major criticism can recognize the initiation sequences of inter-of this model is that it challenges the basic nal cistrons in a polycistronic bacteriophage
mechanism proposed for production of Sindbis RNA.
virion proteins because it presumes thatcapsid This modelis themostsuitableforexplaining
protein translation is independent ofthetrans- the unusual and remarkable ability of the
tlation of envelope proteins. According to this cell-free protein-synthesizing systems de-model, ribosomes could load at the initiation scribed here and in other reports
(6,
7) to sitesof at leasttwocistronsinthepolycistronic selectively form large amounts ofcapsid.
WemRNA in a manner analogous to that demon- hadsuggestedthat formation of
capsid
invitrostrated for the cistrons in E. coli operons (25) was aresultofa
specific
termination of transla-and inthe cistrons of the E. coli RNA bacterio- tion,andweproposehere thatthissamemecha-phages (12, 13). nism operates in the viral-infected cell. The
j Amodelproposingindependenttranslationof presence of internal initiation sites
containing
Sindbis viral RNA cistrons can explain several RNA regions with
secondary
structure could pieces of data that are inconsistent with the also explain why the 26S Sindbis RNA maybe single-site initiation mechanism forviral RNA found in a "33S" form and why thisspecies
translation. For example, itcan accountforthe makeslarger
sizepolypeptides
invitro than does non-stoichiometric levels of the virion proteins the26SRNA(7).
Ina"33S"form,
theseregions
formed in the infectedcell;there istwo tothree ofsecondary
structurewould be convertedtoantimes more capsid piotein than
envelope
pro- extendedconformation,
thereby
leading
toade-teins in infected cells (21). This model also creased
mobility
ofthe 26S RNA inpolyacryl-explains the inability to accumulate a ts-2 amide
gels
and to agreater
probability
for protein when amino acidanaloguesareaddedtoread-through
translation rather thentermina-Sindbis-infected cells
(18),
and itexplains
the tion and initiation ofnewtranslation.
inabilitytoclearlyshowanaccumulationof ts-2 It is worth
noting again
that attempts toprotein when protease inhibitors wereaddedto
reproduce
theexperimental
results demonstrat-infected cells (16).Theonereportclaiming
that ing asingle
site of initiationon aviralpolycis-TPCK ledto increased amountsoflarger poly- tronic mRNA for
poliovirus
andenceph-peptides actually showed very little increase in
alomyocarditis
virus have failed for Sindbislarger-molecular-weight
protein
in contrast toa virus. These include theuseofproteaseinhibi-great decrease in theamount of virionproteins tors, amino acid
analogues,
and very short(16). Ourexperiments withlow levels ofTPCK pulses of radioactive
isotopes
to infected cells. added for short periodsoftime before additionAnd,
in contrast to Sindbisvirus,
thepicor-oflabel failed to show accumulation ofa ts-2 naviruses make
equimolar
amounts of viral protein whenanalyzed bySDS-polyacrylamide
proteins encodedby
the viral mRNA. Some slabgels (E.Duda,unpublished data).
Amodelexperiments
with Semliki Forestvirus,
atogavi-invoking separate sites of initiation for
capsid
rusclosely
related toSindbis,
havesuggested
andenvelope proteincanalso
explain
a50%lossindependent
sites oftranslation for virion pro-ofcapsidproteinincells infectedwith defective teins(9),
although
larger-molecular-weight
pre-Sindbisviruseventhough therewas no
signifi-
cursors have been detected (4). These precur-670on November 10, 2019 by guest
http://jvi.asm.org/
sors may be related to envelope proteins which were affecteddifferently fromcapsidwhen
can-avaninewassubstituted forargininein Semliki
Forest virus-infected cells (17). Finally, the in
vitrotranslationofSindbis viral RNAyieldsan authentic capsid protein in a cell-free system from ascites cells that produces
mainly
prema-turely terminated polypeptides fromenceph-alomyocarditis viral mRNA (1). It may well be
that thepolycistronic mRNAencodingSindbis
virus proteins functions
differently
than that ofthepicornavirusbecauseaportionof the mRNA codes forproteins thataredirectedintothe host cell membrane. Clearly, more intensive studies with the in vitrosystemand with inhibitors and analogues in vivo are needed to testthis model ofmultiple initiations on an animal virus
poly-cistronic mRNA.
Duringthepreparationof thismanuscript,we
learned that D. T. Simmons and J. H. Strauss
(J. Mol. Biol., in press) have also successfully translated in vitrothe 26S RNA isolated from cells infected with a temperature-sensitive
Sindbis mutant that is similar in its properties to the ts-2 mutant. Their results are similarto ours inthatcapsidproteinisnotformedat37 C but isfound at 27 C. However, theyreport the
presence of discrete
larger-molecular-weight
proteins equivalent in size to the B-1 and ts-2 proteinsin theirinvitro reactionproducts.The
former appeared at 27 C, whereas the latter appeared at 37 C. We have been unable to
detect these large proteinsasdiscrete bands in
polyacrylamidegels; instead, wefind a contin-uum ofradioactive material at the top ofour
slab gels. However, recent preliminary results suggest that thelargerdiscreteproteinsmade in
our in vitro reaction at 37Cusingts-226S RNA (Fig. 2a and b) contain capsid protein tryptic peptides.
ACKNOWLEDGMENTS
This research wassupported by a Public Health Service grant(CA14311-01) from the NationalCancer Institute and GB38657fromthe NationalScience Foundation.
LITERATURE CITED
1. Aviv, H.,I.Boime, and P. Leder. 1971. Protein synthesis directedbyencephalomyocarditisvirusmRNA: prop-erties of a transferRNA-dependentsystem. Proc. Nat. Acad.Sci. U.S.A.68:2303-2307.
2. Burge,B.W., andE. R.Pfefferkorn.1966. Complementa-tionbetweentemperature-sensitive mutants of Sindbis virus. Virology 30:214-223.
3. Burge, B. W., and E. R. Pfefferkorn. 1968. Functional defects of temperature-sensitive mutants of Sindbis virus.J.Mol.Biol.35:193-205.
4. Burrell, C. J., S. M. Martin, and P. D. Cooper. 1970. Post-translational cleavage of virus polypeptides in
arbovirus-infected cells. J.Gen. Virol.6:319-323.
5. Butterworth, B. E.,L. Hall, C. M.Stoltzfus, and R. R. Rueckert. 1971. Virus-specific proteinssynthesizedin encephalomyocarditis virus-infected HeLa cells. Proc. Nat. Acad. Sci.U.S.A. 68:3083-3087.
6. Cancedda,R.,andM. J. Schlesinger,1974.Formationof
Sindbis virus capsid protein in mammalian cell-free
extracts programmed with viral RNA. Proc. Nat. Acad. Sci.U.S.A.71:1843-1847.
7. Cancedda.R.,R.Swanson,andM. J.Schlesinger, 1974.
Effects of different RNAs and components of the cell-free system on in vitrosynthesisofSindbis viral
proteins.J. Virol. 14:652-663.
8. Cohen, L. B., I. H. Goldberg, and A. E. Herner. 1969.
Inhibition by pactamycin oftheinitiation ofprotein
synthesis. Effecton the 30S ribosomal subunit.
Bio-chemistry8:1312-1326.
9. Friedman, R. M. 1969. Primary gene products of an
arbovirus. Biochem. Biophys. Res. Commun. 37:369-373.
10. Holland,J.J.,andE.D.Kiehn.1968.Specificcleavageof viralproteinsasstepsinthe synthesis andmaturation of enteroviruses. Proc. Nat. Acad. Sci. U.S.A. 60:1015-1022.
11. Jacobson, M. F., and D. Baltimore. 1968. Polypeptide
cleavagesin theformationofpoliovirusproteins. Proc.
Nat. Acad. Sci.U.S.A.61:77-84.
12. Jeppesen. P.G. N.,J. A. Steitz,R. F. Gesteland, andP.
F. Spahr. 1970. Gene orderin thebacteriophage R17
RNA: 5'-Aprotein-coat protein-synthetase-3'. Nature
(London)226:230-237.
13. Lodish,H.1970.Secondarystructureofbacteriophage f2 ribonucleic acid and the initiation ofin vitroprotein synthesis. J. Mol. Biol.50:689-702.
14. Miller,J. H.1974.GUG and UUGareinitiationcondons
in vivo. Cell 1:73-76.
15. Morrison,T. G., and H. F. Lodish. 1973. Translationof
bacteriophage Q,B RNA by cytoplasmic extracts of
mammalian cells. Proc. Nat. Acad. Sci. U.S.A. 70:315-319.
16. Pfefferkorn, E. R., and M. K. Boyle. 1972. Selective
inhibition of the synthesisofSindbis virionproteins by
aninhibitor ofchymotrvpsin. J. Virol. 9:187-188.
17. Ranki, M. 1972. Nucleocapsid and envelope protein of
Semliki Forestvirus as affectedby canavanine.J.Gen. Virol. 15:59-67.
18. Scheele, C. M., and E. R. Pfefferkorn. 1970. Virus-specific proteins synthesized in cells infected with RNA+ temperature-sensitivemutants ofSindbis virus. J. Virol. 5:329-337.
19. Schlesinger, M. J. 1970. Genetic probes of enzyme
structure, p.241-266.In P.Bover (ed.),The enzymes, vol. 1,3rd ed. AcademicPress Inc., NewYork. 20. Schlesinger, M. J., and S. Schlesinger. 1973. Large
molecular-weight precursors of Sindbis virus proteins. J. Virol. 11:1013-1016.
21. Schlesinger, S.,andM.J.Schlesinger.1972.Formationof Sindbis virus protein.Identification of a precursorfor oneofthe envelope proteins.J.Virol.10:925-932.
22. Smith, J.D., L.Barnett, S.Brenner, and R. L. Russell.
1970.Moremutanttyrosinetransferribonucleicacids. J. Mol. Biol.54:1-14.
23. Summers, D. F.,and J. V.Maizel,Jr.1968.Evidencefor large precursorproteins in poliovirus synthesis. Proc. Nat.Acad.Sci. U.S.A. 59:966-971.
24. Weiss, B.,andS. Schlesinger. 1973. Defective interfering
passagesofSindbis virus:chemical composition, bio-logical activity, and mode of interference. J. Virol. 12:862-871.
25. Zakin, H., C. Yanofsky, and C. L.Squires. 1974. Regu-lated in vitrosynthesis ofEscherichia colitryptophan operon messenger ribonucleic acid and enzymes. J. Biol. Chem.249:465-475.
on November 10, 2019 by guest
http://jvi.asm.org/