0022-538X/79/02-0612/12$02.00/0
Intracellular
Forms of
Simian Virus
40
Nucleoprotein
Complexes
I. Methods of Isolation and Characterization
in CV-1
Cells
R. FERNANDEZ-MUNOZ,tM.COCA-PRADOS,ANDM.-T. HSU*
TheRockefeller University, New York,New York 10021
Received forpublication10August1978
Anewmethodwasdeveloped for isolation of intracellular forms of simian virus
40 (SV40) nucleoprotein complexes from SV40-infected CV-1 cells late in the
infectious cycle. Incontrast totheTriton extractionmethod, which yields onlya
60-70S complex, this newprocedure yielded three forms of SV40 nucleoprotein
complexes: complex I, complexII, and thematurevirion(V). The three
nucleo-protein complexes differed in physicalaswellasbiochemicalproperties.Complex
I, whichis onlya small portion of the total SV40 nucleoprotein complexes late
duringinfection, wasactive in synthesizing both SV40-specific DNA and RNA.
Pulse-labeling experimentssuggestthefollowingmetabolicpathway: I-+ II-+V.
Conversion ofcomplex ItoII occurredshortly after the completion of SV40 DNA
replication and resulted in the inactivation of thebiosynthetic activities ofI.
Simian virus 40 (SV40) chromatin isolated
fromlytically infectedcells exhibits many
prop-erties similar to those of eucaryotic cell
chro-matin. It iscomposedmainlyof a histone-DNA
complex arranged in the nucleosome structure
(8) and is associated with both DNA (19) and
RNA (6)polymerase activities.
We areinterested inusing SV40 chromatinas
a model system for studying the relation
be-tween the structure and function ofeucaryotic
chromatin. During our initial study of SV40 chromatin isolatedby the Tritonextraction
pro-cedure (7), we werepuzzledby thefactthat no
mature virions were everrecovered in the
ex-tract evenwhen mature virions could be isolated
by banding infected cell lysate in a CsCl
gra-dient. Furtherinvestigationshowed that virions
aswellasmanyintracellularnucleoprotein (NP) complexeswere disrupted byTriton treatment to yield a single species sedimenting at about
60S. This observationpromptedus todevelopa
newisolationprocedure which preserves the
in-tracellular forms of the SV40 NP complexes.
With the new procedure three forms ofSV40
NP complexes were isolated. A minor
compo-nent, complex I,sedimentingatabout70S,was
found to beactively synthesizing SV40RNA and
DNA. In addition, a more condensed form of
SV40 chromatin (complex II),which is derived
from complex Ishortlyafter the completion of DNA replication, was isolated as well as the
maturevirion.
tPresent address: Virologia (Microbiologia) Centro "Ra-monyCajal," Madrid-34,Spain.
In thepresent communication wedescribe the
method forisolating these intracellular forms of
SV40 NPcomplexes and the initial
characteri-zation of their physical and biochemical
prop-erties.
MATERIALS AND METHODS
Cell and virus. TheSVS strain ofSV40was used to infect CV-1 cells. The description of virus, the growth of cells and virus, and the infection procedure have beenreported (10).
Extraction of SV40 complexes. Two alternative methodswereusedfor extraction ofSV40 complexes. (i) For total cell extract,48hafterinfection, cells were washedoncewith TDbuffer (25 mM Tris-hydrochlo-ride, pH 7.4, 0.136 M NaCl, 7 mM KCl, 0.7 mM
Na2HPO4)andwerescraped off the plate into2mlof
cold hypotonic buffer (25 mM Tris, pH 7.9, 1 mM
MgCl2, 0.4 mM CaCl2, 0.5 mM dithiothreitol). The
cellswerebrokenby homogenization (30 strokes) in a tightly fittingglass Dounce homogenizer. The nuclei werespun downby centrifugation in an IEC PR-6000 centrifuge at 2,000 rpm for 5 min. The supernatant was thenlayeredon asucrose gradient for isolation of SV40 NP complexes. (ii) For nuclear extract, cells werewashedonce with TD bufferand were scraped into TD buffer at107 cells per ml. The cells were spun down(1,500rpm,3min) and resuspended inTDbuffer at 107 cells per ml, and the detergent Nonidet P-40 (Shell) wasadded to 0.5%. Afterbriefmixing, nuclei werespundown(2,000 rpm, 5min)and resuspended in 2mlof cold TDbuffer,and thenucleiwere homog-enizedbyDouncehomogenization (30strokes).Nuclei werespundown asbefore, and the nuclearextract was analyzed byusinga sucrosegradient.
Sedimentation velocity analysis of SV40 NP complexes.A5to20% sucrose gradientwasusedto 612
on November 10, 2019 by guest
http://jvi.asm.org/
analyze the properties of SV40 complexes. In later experiments we used a 5 to 40% sucrose gradient, which gives better resolution of different forms of SV40 complexes. The buffer usedisindicated in the figurelegends. The conditions for centrifugation were: SW40 rotor, 37,000 rpm,70min,4°C.
Glutaraldehyde fixation of NP complexes and determination of buoyant density. Fractions from asucrose gradient of SV40complex were fixed with glutaraldehyde according to the procedure of Balti-moreandHuang (1). Fixed samples were layered onto apreformedCsCl gradient (1.2 to 1.6 g/ml in 0.01 M Tris-hydrochloride, pH 7.9,0.01MEDTA, 0.1% Triton X-100) and centrifuged in an SW65 rotor at 32,000 rpmfor12hat20°C. Fractions, 100jl,werecollected intomicrotiterplates. A drop of mineral oil was then added to each fraction to prevent evaporation. The density of fractionswasdetermined by the refractive index.
Unfixedsampleswereanalyzed in thesamegradient exceptthata1-ml cushion ofa1.7-g/ml CsCl solution wasincluded. Unfixed samples were also analyzed in
a44%(wt/vol)metrizamide(Nyegaard)gradient in50
mM Tris-hydrochloride, pH 7.4.The gradients were spunat32,000 rpmfor60hat20°C inanSW65 rotor. Gradientswerefractionated asin the CsCl gradient, and the density (p) of metrizamide solutionwas deter-mined from therefractive index (n), using the formula:
p(5°C) =3.453 n(20°C) -3.601.
Gelelectrophoresis.Low-salt agarosegel
electro-phoresis foranalyzing the mobility of SV40 complex was performed according to Varshavsky et al. (20). SV40complexeswereelectrophoresedina0.4, 1.0,or 2%agarose tubegel (12 by0.7cm)containing 10mM Tris,pH 7.4,at 50V for9h. Afterelectrophoresis the gelswere eitherstained with0.5,ug of ethidium bro-mide per ml or0.2% Coomassie brilliant blueorthe
gels were sliced into 2-mm slices for analyzing the radioactivity. Agarose gelelectrophoresis for analyzing supercoiled DNAwasperformed ina2% agarose slab gel containing40 mM Tris-30mM NaH2PO4-1 mM EDTA,pH 7.8, according to theprocedure of Shure and Vinograd (17). SV40 DNA or DNA fragments
obtained after micrococcal nuclease digestion were analyzed in either 1.4or2.5% agarose underthe con-ditionsdescribedpreviously(10).
Protein samples were analyzed by electrophoresis
ona 14%polyacrylamide-sodium dodecylsulfate slab
gelasdescribedby Laemmli (13).
DNA, RNA, and protein labeling, extraction, and RNA-DNAhybridization.SV40DNA was iso-lated from cells infected at low multiplicity as de-scribed previously (10). The DNA was labeled with 200,iCi of['H]thymidine (NewEngland Nuclear, 20 Ci/mmol) perplate,and theradioactivitywaschased bywashingthecells with fresh medium andincubating them in the medium containing 100 mM thymidine (Calbiochem) for1h. Fordouble-labeling experiments, SV40DNA waslabeledwith 20,ICiof['4C]thymidine
(New England Nuclear,40to60mCi/mmol) per plate. Topreparepulse-labeled RNA, cellswereincubated with1mCiof[:3H]uridine(NewEngland Nuclear,>25 Ci/mmol) perml for1min andimmediately quenched onice with thesimultaneousaddition of 10 mlof ice-cold TDbuffer. The RNA was isolated fromSV40NP
complexes after sucrose gradient fractionation by ex-traction withphenol(saturated with buffer containing 0.1MNaCl,0.01MTris, pH 7.4, 0.01 M EDTA, 0.1% sodium dodecyl sulfate) and chloroform-iso-amyl alcohol (24:1). Conditions for the hybridization oflabeled RNAtotheSV40DNAfilteraredescribed inreference12.
Labeled protein was obtained from cells labeled
with [3H]lysine (New England Nuclear, 60 to 80
Ci/mmol),50,uCi/ml,for24to 48hpostinfection. To isolate CV-1cell histone proteins, monolayer cell cul-tures werewashed twice with10ml of TD buffer, and the cellswerescraped off the plates into TD buffer andcollected by low-speed centrifugation. The histone proteinwasextracted from thecellpellet with0.25N H2SO4at4°Cfollowed by further extraction with 0.4 NH2SO4. The two extracts were combined,10volumes of coldacetonewasadded, and the protein was precip-itated at-20°C. (Calf thymus histones were a gift of G. Vidali.)
In vitro assayof RNA polymerase activities. Endogenous RNA polymerase activities associated with SV40 complexes were assayed according to the conditions of Green and Brooks (6). In vitro transcrip-tionof SV40complexes, with Escherichia coli RNA polymerase (purified according to the procedure of Burgess and Jendrisak[3]),wasperformed in40mM Tris (pH 7.9), 10mM MgCl2, 150 mMKCl, 0.4 mM
K2HPO4, 0.1 mM dithiothreitol, 0.15mM each ATP,
GTP, and CTP, and10,iCiof[3H]UTP(NewEngland Nuclear, 35 to50Ci/mmol) per mlat37°C for30min. Micrococcalnucleasedigestion. SV40 complexes isolated after sucrose gradientfractionation were
di-gestedwith500U ofmicrococcal nuclease
(Worthing-ton) perml for10min inbuffercontaining25mMTris (pH 7.5)-50mM KCl-10 mM NaCl-10mM
mercap-toethanol-10 mM MgCl2-1 mM CaCl2-0.15 mM spermine-0.5 mM spermidine. The reaction was stoppedby the addition of sodium dodecylsulfateto 0.5% and EDTA to 25 mM, and the DNA was ex-tracted with phenol-chloroform-isoamyl alcohol (24: 1)andprecipitated with2volumes of ethanol.
Electronmicroscopy.Adropofsolution contain-ingSV40complexes in TD bufferorin0.1M Tris(pH 7.4) was placedon a sheetofParafilm. Immediately after the drop was placed on Parafilm, grids coated with pallodion were touched to the drop for5 sand washed in distilledwaterfor5s.Alternatively,adrop ofsolutioncontainingcomplexeswasplacedon agrid for5 s, and theexcess liquidwasremoved withfilter
paper.Gridswereshadowed withplatinum-palladium
(80:20) with orwithout priorstainingin 5 x 10-5 M uranylacetatein90%ethanol. The method for spread-ing of SV40 DNA in 50% formamide or the SV40 complexesin 50 to70%formamide has been described (11).
RESULTS
Isolation of SV40 DNA-protein
com-plexes.Previously,the isolation of intracellular
SV40 NPcomplexeshasbeen achievedbyusing
Triton extraction method (7). This extraction
methodyields asingle SV40DNAprotein
com-plexsedimentingatabout 60-70S in thesucrose
on November 10, 2019 by guest
http://jvi.asm.org/
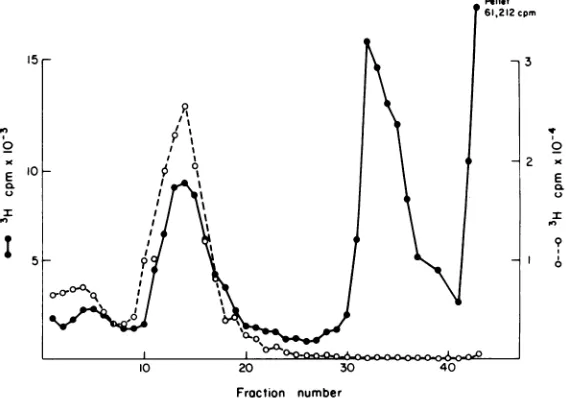
gradient(Fig. 1). However, repeated efforts with thismethodto recover maturevirions fromcells late after infection failed, even when a large
amountof virion could be isolated fromsimilar
cultures afterCsCl gradient fractionation of
ex-tracts obtained from cells lysed by repeated
freeze-thawing. The absence ofany mature vir-ion in the Tritonextractstrongly suggeststhat the intracellular forms ofSV40 NP complexes
are disrupted during isolation. Further investi-gation showed that40to50% of [3H]thymidine-labeled virions purified on aCsCl gradient are
converted into a species sedimenting at 60S
whencoextracted with unlabeled SV40-infected cells by the Triton method. The disruption is dependent on the presence of Triton-treated cellularmaterial becauseno disruptionwas ob-served whenpurified virus alonewas extracted with Triton. These observations indicate that the 60-70S complex extracted by the Triton method could be derived from many different forms of NP complexes, includingmature
viri-ons.To avoid thedisruption of intracellular NP complexes,a newisolationprocedurewas devel-oped. Briefly, SV40-infected cells in hypotonic buffer or nuclei in isotonic buffer obtained by Nonidet P-40lysis ofSV40-infected cells were
homogenizedin atightly fitting Dounce homog-enizer (see Materials andMethods).SV40
com-plexesselectivelyleakedoutfromthenucleiand
could be separated from cellular chromatin by low-speed centrifugation. SV40 complexes
ex-15
E 10_ i
I
'
tracted bythis new procedure exhibitvery
dif-ferentprofilesinthesucrosegradient fromthose
obtained by the Triton method (Fig. 1 and 2).
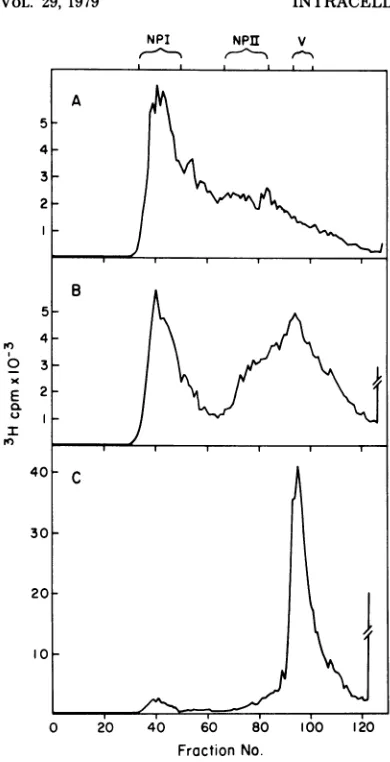
When analyzed ina 5 to20% sucrose gradient,
major components were observed sedimenting
at70S (NP-I) and 180-200S (Fig.1). The heavier
speciescanbe furtherresolved intotwo
compo-nentswhen thecomplexesarepulse-labeledand
analyzed in5 to40%sucrosegradients (Fig. 2).
The material sedimenting at about 200S is
mostly SV40 virionasdefinedbythecriteriaof
electron microscopic morphology and CsCl
gra-dientbanding (virion bandsat a density of1.35
g/ml). The heterogeneousmaterial sedimenting
betweenNP-Iand SV40 virions (V) was opera-tionallydesignatedNP-II.
Thepossibility that SV40 complexes obtained by thenew procedure are due toartifacts
gen-erated during the isolation procedure was
ex-amined in a series of control experiments. 3H-labeled NP-I, NP-II, or virions were added to unlabeled SV40-infected cells and extracted by using the newprocedure. If NP-I orNP-IIwas
derived from virions during homogenization, then coextraction of labeled virion with unla-beled infected cells shouldyield labeled material sedimentingatNP-Iand NP-II positions. Such resultswere notobtained intwotrials.Similarly,
NP-II could be derived from NP-I by
aggrega-tion with itselforwithother cellularcomponents
during extraction. Coextraction of labeledNP-I withunlabeled infectedcells demonstrated that
Pellet
61,212 cpm
3
0 2"x E
IL
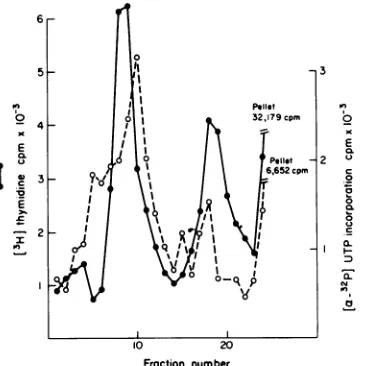
Fraction number
FIG. 1. Sucrosegradientanalysis of SV40chromatinisolatedby the Tritonextraction method(0) and by thenew method asdescribed in the text (0). Cells infected withSV40werelabeledwith 200IuCiof
PH]-thymidineper 100-mmdishofcellsfrom36 to48hpostinfection. SV40chromatinwasextractedandanalyzed
ina5 to20%1csucrosegradient containing50mMTris, pH 7.9,inanSW40rotor(37,000rpm, 60min,4°C).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.129.412.430.629.2]NPI NPfl V
f-11 r-N
0 20 40 60 80 100 120
Fraction No.
FIG. 2. Five to 40%o sucrosegradient analysis of
SV40 DNAprotein complexes extractedby the new
method(see text). SV40-infectedcellswere
pulse-la-beled with [3H]thymidine for (a)30min, (b) 90min,
or (c) 24h, with the endpoint of labeling at48 h postinfection. SV40 complexes were extracted from infectedcellsbyDouncehomogenizationinhypotonic solution andanalyzedina5to40%osucrosegradient
containing 2 mM Tris, pH 7.4, in an SW40 rotor
(37,000 rpm, 70min, 40C). Gradients were
fraction-atedfrom the top into100-julfractions. Similar
pro-files were obtained when the buffer in the sucrose
gradientwasisotonic TDbuffer. In TDbuffer NP-I
sediments slightly faster, whereas virions sediment slightlyslower.
this is notthe case. Similarargumentsindicate
that NP-I couldnotbe derived fromNP-II
dur-ing extraction. These experimentsdemonstrate
thatthe isolated NPcomplexesretain their
char-acteristic sedimentation properties upon
reex-tractionand are not artifacts of the extraction
procedure; therefore, they probably represent
genuineintracellular forms ofSV40NPs.
The efficiency ofextracting SV40 complexes
by the newprocedurewasestimatedasfollows. SV40-infected cellswerelabeled for 24h (24 to
48 h postinfection) with[3H]thymidine. The
ra-dioactive label present in the three classes of
SV40 complexes was compared with the label
present in the Hirt extract of the same nuclei
remaining after prior extraction ofcomplexes.
The ratio of label present in theisolated
com-plexes to that in the Hirt extraction is 1.1 x 106
cpm/1.0 x 106 cpm or about 1, indicating that
theextraction efficiency is about 50%. Analysis of the material present in the two extracts
showed that more than 98% of the material
present in the threeclasses of complexesisSV40
DNA, whereas, at most, 50% ofthematerial in
the Hirt extract is SV40 circles. This suggests
that theefficiency of extractionmustbeatleast 50% and perhaps as much as 70%. In another experiment, nuclei collected after Dounce ho-mogenizationwerereextracted in Triton
extrac-tion buffer, and the extract was analyzed on a
sucrosegradient. No 70Scomplexwasobserved
inthe sucrose gradient, suggesting that
extrac-tion with the newprocedure is at least as effi-cientasTriton extraction.
The extraction of[3H]thymidine-labeled NP complexes from total cells and from nuclei
pre-pared from Nonidet P-40-lysed cells yields the
same profile in thesucrose gradient. However, in thewhole-cell extractthere is extensive
con-tamination of NP-Iby cytoplasmic material,
es-pecially ribosomes. These contaminations were
mostly eliminated by extracting SV40complexes from the isolated nuclei obtained bylysing in-fected cells with Nonidet P-40. Figure 3shows
thesucrosegradient profileofanuclearextract
labeled with
[3H]thymidine
and[3H]lysine
from24to 48hpostinfection and with 32P from36 to
48hpostinfection. Host protein contamination
was greatly reduced (Fig. 3). Complexes ob-tainedfromnuclearextract weretherefore used foranalyzingitsprotein compositions.
Properties of SV40 NP complexes. The physical as well as biochemical properties of SV40 complexes are summarized in Table 1.
Theyare discussedbelow.
(i)Physical properties.Thefollowing phys-ical properties of SV40 complexes isolated in
sucrosegradientwereanalyzed.
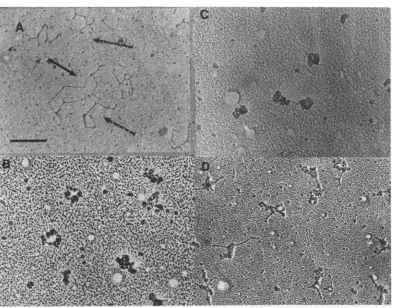
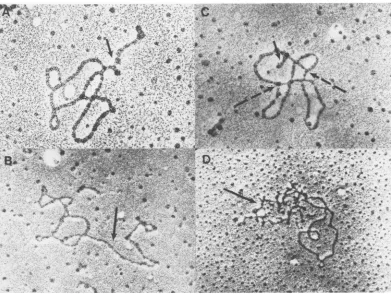
(a) Morphology ofSV40complexeswas
ana-lyzed by electronmicroscopy.Different fractions
in the sucrosegradient containingisotonicbuffer
corresponding to NP-I, NP-II, and V were
di-luted in either isotonic TD bufferor0.1MTris,
pH 7.4, and deposited on the electron
micro-scopegridsasdescribed inMaterials and
Meth-ods. Whensampleswerepreparedin TDbuffer
and extensive surface tension during sample
o
x
E
I
Iv,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.46.241.65.446.2]E
I?
'0
'E
-J-I?
oo 0 4 0 8 o l2
'0
x~~~~~Foto No.
E
a-0 40 r60 80 100 120
Fraction No.
FIG. 3. Profile ofSV40 complexes in a 5 to 40%O
sucrosegradientlabeled with(a) [3H]thymidineand
(b) [:IH]lysine from 24to 48 hpostinfection and(c) with 12Pfrom36to48 hpostinfection.SV40complexes
were extractedfrom the nuclei of infected cells as
described in thetext. Comparetheprofile of12-h32P
labelingwith thatof12-h['Hithymidine labeling,as
shown inFig. 13e.
preparation was avoided, NP-I was seen as a
"beaded" nucleosomalstructure(Fig.4B). In the
same method ofsample preparation, NP-II
ap-pearedas a morecondensed structure (Fig.4C).
Whenthesampleswerepreparedin 0.1 M Tris
(pH 7.4), NP-I was reproducibly observed to
consist of thin DNA-protein fiber about 80
Athick. Mostof the NP-Icomplex spreadunder
this condition showed a supercoiled structure
(Fig. 4A, dashed arrows) similar to that of the
free SV40 DNA. Occasionallyrelaxed open
cir-clescanbe observed (Fig. 4A,solidarrow).The contourlength oftheseopencirclesisabout 1.5
Am, similar to that of free DNA. In contrast,
NP-II, when spreadunder thesame conditions,
showedareproduciblymore compact structure
(Fig. 4D).Occasionally, thin fiber similartothat
observed in NP-Isamples could beseenaspart
of theNP-IImolecule. These observations
dem-onstrate that NP-I and NP-IIhave a different
conformation, whichcanbeeasilydifferentiated
inthe electronmicroscope.
(b) Buoyant density of SV40 complexes was
analyzedinCsClgradientsafterglutaraldehyde
fixation or in a metrizamide gradient without
fixation. NP-I has adensity of1.45 g/mlinthe
CsCl gradient, whereas NP-II and the virion
have densities of 1.35 g/ml (Fig. 5). Although fixed NP-II and the virion have thesamedensity
in CsCl, unfixed NP-II is disrupted into free protein and DNAcomponentsinaCsCl gradient
whereasthematurevirionisnot.Inmetrizamide
gradientanalysis, NP-I and NP-II have thesame
density, 1.18 to 1.20 g/ml, whereas the mature
virion has a density of 1.26 g/ml (Fig. 5). The
secondcomponent ofdensity1.2g/ml in Fig. 5C
is due to the presence of NP-Il material in a
viruspeakinthe sucrosegradient. Thepresence
ofNP-II material in the virus peak isalso
ob-served by electron microscopy as described
aboveandby analyzing unfixed materialin the
virus peak intheCsCl gradient.
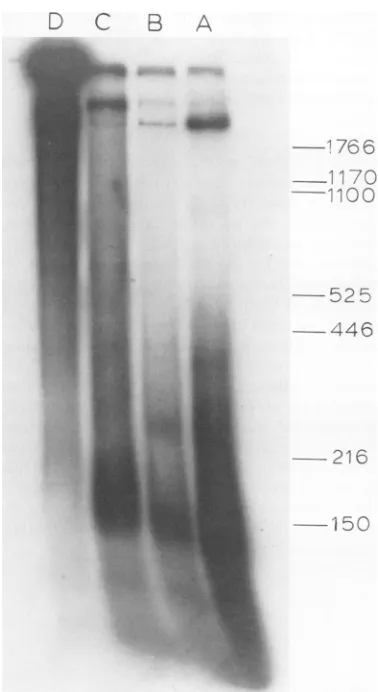
(c) Electrophoresis of SV40 complexes in a
1.0% agarose gel shows that NP-I, NP-II, and
virion have different mobilities (Fig. 6). NP-I and NP-II have broad distribution in the gel. However, the ratio of[3H]lysine to
["4C]thymi-dine label remains approximately constant
throughout the band, indicating that the
breadthofthe band is not theresult of
dissocia-tion of proteins during electrophoresis, but
ratherdue totheinherentheterogeneity of the
samples.
(ii)Biochemical characteristics. TheDNA of SV40 NP complexes is mainly supercoiled
DNA(R90%),asdetermined by electron
micros-copy and agarosegelelectrophoresis. The
num-ber ofsuperhelicalturnsintheDNA ofall three complexesappears to be the same aswhen
an-alyzed by agarosegel electrophoresis (14). The
DNA is arranged in a nucleosome structure in
TABLE 1. Physicalandbiochemicalproperties of SV40 complexes
Property NP-I NP-II Virion Sedimentation coef- 70S 180S 210S
ficient
Density (CsCl; 1.45 1.35 1.35
jg/ml)
Density (metriza- 1.18 1.18 1.26 mide;,ug/ml)
Morphology (elec- Open Condensed Virion tron microscopy)
Replication activity Yes No No invivo
Transcription activ- Yes No No ity in vivo
Replication activity Yes No No invitro
Transcription activ- Yes Variable No ityinvitro
Supercoil density Same Same Same (DNA)
Micrococcal nuclease Nucleosome Nucleosome digestion
HistoneHi Yes Yes No
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.275.466.484.669.2]'-C
15E
Pli,~~
FIG. 4. Electron micrographs ofSV40 NP complexes.SV40 NP complexes obtained from a sucrose gradient in TDbuffer were diluted into TD buffer (B, C) or 0.1 M Tris, pH 7.4 (A, D). Pallodion-coated grids were touched to adrop of complexsolutionfor 5 s. After excess solution was removed, the grids were shadowed
withplatinum-palladium(80:20). (A, B) NP-I; (C, D) NP-II.
NP-I and NP-II as manifested by micrococcus nuclease digestion (Fig. 7). A resistant core of about 150 nucleotides inlength was observed. This result is similar tothatobtained for
poly-omavirusby Ponderetal. (16).
(a) Preliminary analysis of theprotein
com-ponents of SV40 complexes is given in Fig. 8.
NP-I is composed mainly of the five groupsof histones and material migrating in the same
position as VP-1 protein of SV40 virus in a
sodiumdodecylsulfate-polyacrylamide gel.The majorproteinsin NP-II, however, arethe viral capsidproteins VP-1, VP-2, and VP-3. Histone proteinsarealsopresentbut in loweramounts.
Histones, exceptfor
Hi,
arepresent in thema-ture virion. Thepresence ofHiinNP-Iand
NP-II but not in the mature virion has also been
confirmed bythe acid-urea gel electrophoresis technique (15; M. Coca-Prados, G. Vidali, and
M.-T.Hsu,manuscriptinpreparation). Further
characterization of the protein components of
theSV40 NP complexesand their modification
willbereportedelsewhere.
(b) Endogenous RNA and DNA polymerase
activities associated withSV40complexeswere
analyzed byinvivo andinvitroincorporation of RNAorDNAprecursors, Toanalyze the in vivo transcription activities,SV40-infected cellswere pulse-labeledwith[3H]uridinefor1min at37°C.
(Aseparatebiochemicalanalysis of 1-min
pulse-labeled RNA showed thattheyrepresent
grow-ing RNA chains [5].) SV40complexeswerethen extracted fromtotalcells andseparated ina 5 to
20% sucrose gradient. Labeled RNAs were
ex-tracted from each fraction of the gradient and assayed for SV40-specific RNA by hybridizing
to anSV40 DNA filter. Figure 9 shows that
1-minlabeledSV40 RNA is associated withNP-I
and notwith NP-Il or virion. The endogenous
RNApolymeraseactivities werealsoassayedin
vitro for the incorporation of
[3H]UTP
ora-[32P]UTP
into acid-precipitable material. NP-Icanbereproducibly foundtobe active in
tran-scription (Fig. 10). NP-II, on the other hand,
gave low but variable activities. At present we
donotknow thecauseof suchvariabilityinthe
activity of NP-II. Thespecificityof the in vitro
transcriptionofNP-Iwasassayed by hybridizing
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.56.450.73.381.2]CSCI Metrizomide taining 40 or 20% formamide, elongating RNA
a a' could beseen associated with SV40 DNA. The
-Ao A 6
frequency
ofobserving
such structuresis aboutl 50, 1% of the total SV40 DNA scored on the grid.
2- l 14513 , This
frequency
remainedapproximately
the A 140 1.2- 135
140
[l2 ;-1O-
same even when thesample
was diluted to the1 v-o / \ 1 35 t' - 2 extent that no more than twoSV40DNAs could
beseenin thesame
grid
areaina300-meshgrid.
I b b ^ This
suggests
that the structures observed are° 8- 1\ 0.8 °
unlikely
tobe the result of thesuperimposition
6
E- -- 06 E of free RNA and DNA on the
grid.
TreatmentCL6 w'1.-50 1.3 I' 0,6 o
-Q
S>>°}45 [12
1 l , _-°' Q of nuclei with RNase beforespreading
elimi-4-
Itw-
140 11 OA-0 natedtheseSV40 DNA-associated chains,dem-WI 2 1.35 0.2 I
onstrating
that these chainsareRNAinnature.The extended RNA chain in 70% formamide
c c' 20 spreading (Fig.
11A,
B, and C) could be seen to15- 1 A
collapse
intosecondary
structures in 50%form-s50o 3 A 115 amide
(Fig.
liD, arrow), atypical
behavior of 10o 145 12 / -e 1 single-strandedRNA. An RNA chain almost as+-
. 140 11long
as theSV40
genomelength
could beob-5- 135 10 05 served
(Fig.
liB).
Rarely,
replicating
molecules. with RNA chain could also be observed (Fig.
0 10 20 30 40 10 20 30 40 llC).
[image:7.505.67.253.80.322.2]Sedimentation
FIG. 5. Analysis of density of SV40 complexes. a b c d SV40 complexes were labeled with [3H]thymidine
from 24 to 48 hpostinfection, as in Fig. 2c. Fractions corresponding to complexes I and II and virion (see Fig. 2c) wereanalyzed ina44% (wt/vol) metrizamide gradient in 50 mM Tris, pH 7.4 (a', b', c') or in a preformed CsCl gradient after fixation of SV40 com-plexes with glutaraldehyde (a, b, c). Solid curves representradioactivity. Dashed curves represent den-sities.(a,a') NP-I; (b, b') NP-II; (c, c') virion.
labeledproduct to SV40 DNAfilters.About 50%
of the label became hybridized to SV40 DNA
filters after 24 h of incubation at 65°C and
treatmentof thefilters extensively withRNases.
Underthesameconditions, labeled
complemen-tary RNA transcribed from SV40 supercoiled
DNA by Escherichia coli polymerase
hybrid-ized tothe extentof60%. Thus, most, ifnotall,
ofthe RNAtranscribedinvitrorepresents viral
sequences. Since almost nohost DNA was
ob-served in the DNA extracted fromNP-I complex
when analyzed in the electron microscope, we
believe thatthe lower efficiency ofhybridization
of RNA transcribed from NP-I as compared to
complementary RNA is probably not due to
transcription of host DNA.However, the nature
of in vitrotranscripts as to its sizes andlocations
on theSV40genome has not been characterized.
SV40transcriptionintermediates in SV40-in- FIG. 6. Low-salt agarose gel
electrophoresis
offected cells could be observed in the electron SV40 complexes. SV40 complexes corresponding to
microscope. When nuclei ofSV40-infected cells fractions 42 (a), 61 (b), 78 (c), and 96
(d)
in Fig. 2cwereadded to a solution containing either 70% wereelectrophoresed in0.4%agarose in 10 mMTris
(Fig.I1A, B, and C) or 50%(Fig.liD)formamide (pH 7.4)at 50 V for 9 h. The gel is stained with 0.5
andimmediatelyspread ontoahypophasecon- pgofethidiumbromideperml.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.292.437.324.608.2]D
C
B
A
FIG. 7. Microccocal nuclease digesti complexes. 32P-labeled SV40 complexes ingtothose inFig.6(approximately1 l digested with500 Uof microccocalnuc for 10 min at 37°C. The reaction was adding EDTA to0.1M and sodium do to0.5%.DNAwasextractedbyphenolan(
andanalyzedin a2.5% agarosegel. (A,
(B, C)complexII; (D)virion.
DNA-synthesizing activities of
plexeswerealsoanalyzed. Pulse-labe for 10 min with [3H]thymidine de that the majorityofDNA-synthesiz is associated withNP-I (Fig. 12).AE
clusionwasobtainedby assayingin
poration of[3H]TTP (datanotshow
Relationship among SV40NPc
Todeterminewhether theremightt
sor-productrelationship amongthe ofSV40complexes,weperformeda I
experiment (see Fig. 13). As descr
ously,shortpulses with[3H]thymid belNP-I.Achase for1h with100ml\ thymidineresulted in theappearanc
the NP-II region.This result suggests that
NP-II isderivedfrom NP-I. Since NP-IL and virions
are not very well separated in the 5 to 20%
sucrose gradient, we further characterized the
complex in the NP-II-virion region by a CsCl
gradient. Mature virions band at a density of
_ 766 1.34
g/ml,
whereas SV40complexes
aredisso-1~ii
70 ciated into DNA and proteincomponents,which_I
0 band at densities of 1.7 and 1.2 g/ml,respec-100 tively. Such an analysis was carried out using
SV40 complexes pulse-labeled with
[3H]thymi-dine fordifferent lengths oftime,with the end
point oflabeling always at 48 h postinfection.
5 25 The
complexes
extractedwerefirstseparated
in-446 H2b
H4 H2a H3 VP3 H1 VP2 VP1
150~~~~5
0
L)
3
-o ~ b
ion of SV40 >1 2-
correspond-to 5Ag)were 1 leaseperml
stopped by r?
decyl sulfate CD
dchloroform a
ComplexI E
-015-
A
SV40
com-lingin vivo
monstrated .ing activity
similar
con-vitro
incor-rn).
omplexes.
:e a
precur-three forms
pulse-chase
ibed
previ-ine only
la-dunlabeled
eoflabelin
Fraction No
FIG. 8. Sodiumdodecylsulfate-polyacrylamidegel
(14%o) electrophoresis analysis of proteins in SV40 chromatin. (a) Complex I; (b) complex II; (c) virion. Proteins were labeled with['Hilysinefrom 24 to 48 hpostinfection. Gelwasstained with0.2%oCoomassie brilliant blue(inserts), slicedinto 1-mm slices, and counted (solidcurves).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.505.53.242.78.424.2] [image:8.505.256.449.241.608.2]620
1000
S
E
I 500 400 300 200 100
A
\1
A4
F'
lIt
i
It 'II I'
4
-
I\
I
<d
600 500 400 300 N
200 c
E
100 Li
I 1:
Pellet 10 20 24
Fraction-FIG. 9. Analysis ofnascentRNA associated with SV40complexes. SV40-infected CV-1 cells(5 x 10) were pulse-labeled for 1 min with [3H]uridine (1 mCi/mlof medium). SV40 complexeswereextracted from cells by Douncehomogenization of cells in hy-potonic solution andanalyzedina 5 to20%osucrose gradient. RNAs from each fraction were extracted withphenol andchloroformandhybridizedtoSV40 DNAfilters (1
pwg/filter).
Symbols: (0) totalincorpo-ration;(0)hybridizedcounts.
a 5 to 20% sucrose gradient into two general
regions, I andII (seeFig. 13a-e). Complexesin
region II which include both NP-II and SV40 virionswerefurtheranalyzedin a 1.2 to1.6-g/ml linear CsClgradient witha 1.7-g/ml CsCl
cush-iontotrapthe freeDNA(Fig. 13f-g). In the 30-min label, most of the label appears in NP-I
(Fig. 13a),whereasduring the12-hlabelingmost
oflabel appears inthematurevirion and NP-II (Fig. 13e and j). These results and the pulse-chaseexperimentsuggest thefollowing
biosyn-theticpathway for SV40complexes:
' NP-I->NP-II-- virion.
Appearance of label in the NP-IIregionis rather
rapid (within30minof thepulse-labeling time,
see Fig. 2), whereas the SV40 virion is labeled
only after 1h ofpulse-labeling. Sincethe time
requiredforoneround ofSV40 DNAreplication
is about 10 to 15 min, this result implies that
shortlyafter thecompletionof DNAreplication
NP-Iis converted intoNP-II.
Additional experiments have shown that a
similar kinetic relationship could be
demon-strated as early as 16 to 17 h after infection,
shortlyafter theonsetof viralDNAreplication
(datanotshown). However,the amountof
NP-II and virion is muchreduced relative toNP-I
ascomparedwiththatat 48hpostinfection.
DISCUSSION
Using a new isolation procedure, we have demonstrated that thereare atleast three forms ofintracellular SV40NP complexes. That they
are unlikely to be artifacts generated during isolation procedures is shown by the various
controls and by the differential labeling ofthe
complexes. In contrast, extraction with Triton
convertsall the intracellularcomplexes,
includ-ing virions,into the60S-70S form (NP-I).Since
wehaveshownthatNP-Irepresents only a small
portion oftheintracellular SV40 NP complexes late during theinfection cycle, the60S-70S SV40 complexisolatedby the Triton method actually is composed mainly of material derived from
NP-Il andmaturevirions,which are not active inbiosynthesis of SV40 RNA and DNA. Thus, webelieve the isolationprocedure is superiorto
the Triton extraction method inpreserving the
structuresofSV40NPcomplexes and for
study-ing theirbiological functions.
E
0
E I. E f
7
o
E
c
0
.-a
0 0
a- I-0
a.
[image:9.505.91.237.79.315.2]Fraction number
FIG. 10. In vitroincorporation ofa-["P]UTP into
acid-precipitableRNA bySV40complexes.
SV40-in-fected cellswerelabeledwith[3H]thymidine from44
to 48hafter infection.SV40complexes extractedfrom totalcellsby Douncehomogenization ofcells in hy-potonic bufferwereseparated ina 5 to20%osucrose
gradientcontaining50 mM Tris(pH 7.9),50mMKCI, and 0.5 mMdithiothreitol. Fractions collectedfrom
the sucrosegradientwereassayedfortheactivity of
incorporation ofa-[32P]UTP into acid-precipitable
RNA. Symbols: (0)
a-[32P]UTP
incorporation; (0)[3H]thymidine incorporation. Pelletrepresents
con-taminatingnuclei.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.505.275.458.372.555.2], *' ZrXDD , '
FIG.4 11. Electron micrographs of SV40 transcription complexes. Nuclei from SV40-infected cellswere
iAn4 tZ~~~~~~~~~~~~4 f . V
4*~~~~~~~~~~~~~~ ~ ~ ~ ~,~*~ ~ ~
iDicAtemplaenascntRNAchains. Indark fohrmamidealuysprseRnt.chInsmayreprnextendedNconolymeation whereasa
replicatingmolecule. Thetworeplicating forksareindicatedbybrokenarrows.
The three formsof SV40NPcomplexes differ
in both physical and biochemical properties.
NP-Iis active in thesynthesis of SV40DNA and
RNA. The DNA in NP-I is more sensitive to
DNase Idigestion thanNP-Il (E. Derman and
M.-T. Hsu,unpublishedobservations). Analysis
of both its physicalandbiochemical properties
suggests that NP-I is the active chromatin of
SV40. Because it can be isolated
uncontami-natedby the other forms ofSV40NPcomplexes
andbythecellularchromatinandbecauseof its
simple genetic complexity, webelieve that
NP-I representsanexcellent modelsystemfor
study-ing thestructures and functions of active
chro-matinineucaryotic cells.
Incontrast toreportsby otherworkers,SV40
NP-Icomplex wasobservedasauniform 8-mm
DNA-protein fiber when spread in 0.1 M Tris,
pH 7.4.Thisstructuremaybe derived from the
nucleosomalstructure by unfoldingthe
nucleo-somes as aresult of charge repulsion between
DNA-protein fibers in a low-salt environment.
Variability of chromatin conformation observed
in the electron microscope as a result of the
difference inmethods and buffersused for
pre-paring samplesiswell known (17). In our
expe-rience, not only are salt concentration and pH
importantfactors inspreadingSV40complexes,
but surface tension of the sample is a major
factorcontributingtothevariabilityof structure
observed in the electron microscope. This
vari-abilitynotwithstanding,theSV40NP-Iand NP-IIcomplexes, when spread under identical
con-ditions, showedreproducibly recognizable
differ-ences in morphologyunder the electron micro-scope.
The major protein components present in
complex I are composed of histones anda
pro-teinthatcomigrates with VP1. At presentwedo
not know the function of VP1 in NP-I. The
possibility that it is aggregated with NP-I during
isolation cannot be excluded. The presence of
the histone Hi fraction in SV40 NP complex
extracted by the Triton method has been
re-ported by Varshavsky et al. (20) but not by
others (2, 14). Our results show thatHi is
pres-VOL. 29, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.505.50.441.81.375.2]1000
V
a-9
C
* 0
to
o
vs 0
I o#
._-._-I
E on
Fraction number
FIG. 12. Pulse-chase of SV40 complexes with
[3H]thymidine.Atotalof1O SV40-infected cellswere
pulse-labeledwith[3H]thymidine (1mCi/5 mlof
me-dium)for10min (0) and chasedwith 100mM
thy-midine for 1 h (0). SV40 complexes in total cell extract wereanalyzedina5to20%osucrosegradient
containing 50 mM Tris, pH 7.9. Pellet represents contaminating cell nucleiasanalyzedwith the
elec-tronmicroscope.
ent in NP-I and NP-II but not in virion. This
implies that during SV40 infection Hi first
be-comesassociatedwith the uncoated virus and is
later removed when SV40 virus is assembled.
Furtheranalysis is necessary tounderstandthe
functional role ofHiinthe virusinfection
proc-ess.
The histones present in NP-I complex are
highlymodifiedbyphosphorylationand
acetyla-tion. In addition, highly phosphorylated and
acetylated non-histone proteins were also
ob-served (Coca-Pradosetal., manuscript in
prep-aration).
Therole of NP-II in theSV40 infection cycle
isstill unknown. It is derived from NP-I within
20min aftercompletion of DNA replication of
NP-I. At present we do not know what is the
control process that determines the conversion
of NP-I toNP-II with the concomitant change
of conformation and inactivation of the
biosyn-thetic activities ofSV40chromatin. Perhaps it
istriggeredbythe addition of viral coatproteins
Fractions
FIG. 13. Labeling kinetics of SV40 DNApresentin SV40 complexes. SV40-infected cells were pulse-la-beledwith[:H]thymidinefor30min (a,f),90min (b, g),3h(c, h),5h(d,i),or 12h (e,j). Theendpointof labelingperiodswasalways at48h after infection. SV40complexes obtained in total cell extracts were analyzedfirst ina 5to20%osucrosegradient contain-ing 50 mM Tris,pH 7.4. Portions ofthe gradient labeled"II"werepooledandlayeredon a 1.42-g/ml
CsClgradient with a 1-ml cushionof1.7g ofCsCl per ml. Thegradientwasspunat33,000 rpmfor20h in a type 65 rotor. Sucrose gradients (a-c); CsCl gradient(f-j).Sedimentation isfrom lefttoright. In theCsClgradientprofile,thevirionbands at the top ofthegradient(f-j,left),whereasfreeDNA molecules sediment to the bottomofthegradient(f-j,right).
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.505.73.263.72.352.2] [image:11.505.273.463.79.521.2](e.g.,VP3 whichreplaces Hi histones) which are
presentin large quantity in NP-II. These added
proteins mustbe in arelatively loosestructure
sinceNP-II has the same densityas NP-I in a
metrizamide gradientand its DNA is accessible
to micrococcusnucleasedigestion. Study of the
mechanism ofNP-I toNP-II conversion should yield information as tohow the geneactivity of chromatin is regulatedatthe level of the struc-tureof chromatin.
The buoyant density of NP-II is similar to
thatofNP-Ichromatin in the metrizamide
gra-dient whereas inthe CsClgradient it is similar
to that of SV40 virions. The protein-to-DNA ratio ofNP-II calculated from its protein
com-position predicts thatits buoyantdensity should be similar to that of SV40 virions. The lower than expected density of NP-II in a nonionic metrizamide gradient isprobablydue to
exten-sive hydration of the loosely structured NP-II complex. SV40 chromatin in NP-II is as
acces-sible to micrococcal nuclease digestion as the
NP-I complex, suggesting that the capsid
pro-teins inNP-Ilareprobably only loosely attached
to SV40 chromatin. The "porous" structure of
NP-IIallowswatermoleculestofreelypenetrate
inside the complex, whereas the interior of the
maturevirionislessaccessible. Thismayexplain
why NP-II bands at a lower density than the virioneventhough they haveasimilar protein-to-DNA ratios.
The population of NP-II is rather
heteroge-neous. Itprobably contains different
intermedi-atesbetween the active chromatin (NP-I) and theassembled virus. Furtheranalysis of NP-II, using the electronmicroscopetechniquetostudy
the process of virus assembly, will be reported
(manuscriptinpreparation).
ACKNOWLEDGMENTS
We thankJ. E. Darnell for constant encouragementduring thisinvestigation.We aregratefultoJ.Cozzitorto forexcellent
technical assistance andtoJ.Ford,E.Johnson, M. Wilson, and G. Vidali forhelpfulcriticism and discussions.
Thisinvestigationwassupportedby Public Health Service National Cancer Institute grant CA19073 to M. T. Hsu. R. F.-M.was anEMBOFellow. M.C.-P. isapostdoctoralFellow of theUSA-SpainCultureCooperationProgram.
LITERATURE CITED
1. Baltimore, D.,and A. S.Huang.1968.Isopycnic sepa-ration ofsubcellularandcomponents frompolyovirus
infectedand normalHeLacells.Science162:572-574. 2. Bellard, M.,P.Oudet,J. E.Germond,andP. Cham-bon. 1976.Subunitstructureof simian virus 40 mini-chromosome. Eur. J. Biochem.70:543-553.
3. Burgess, R.R., and J. J. Jendrisak.1975. Aprocedure
for the rapidlarge-scale purifiecation of E. coli
DNA-dependentRNApolymerase involving polymin P pre-cipiation and DNA-cellulose chromatography. Bio-chemistry 14:4634-4638.
4. Cremisi, C., P. F. Pignatti, 0. Croissant, and M. Yaniv. 1976. Chromatin-likestructures in polyoma vi-rusand simian virus 40 lytic cycle. J. Virol. 17:204-211. 5. Ford, J. P.,and M.-T. Hsu. 1978. Transcription pattern of invivo-labeled simian virus 40: equimolar transcrip-tion beyond the mRNA 3' terminus. J. Virol. 28: 795-801.
6. Green,M.H.,and T. L. Brooks. 1976. Isolation of two forms ofSV40 nucleoprotein containing RNA polym-erase from infected monkey cells.Virology 72:110-120. 7.Green, M. H., H. I. Miller, and S. Hendler. 1971. Isolation of apolyoma-nucleoprotein complex from in-fected mousecell cultures. Proc. Natl. Acad. Sci. U.S.A. 68:1032-1036.
8. Griffith,J. D. 1975.Chromatinstructure:deduced from aminichromosome. Science 187:1202-1203.
9. Hall, M. R., W. Meinke, and D. A. Goldstein. 1973. Nucleoprotein complexes containing replicating simian virus 40 DNA: comparison withpolyoma nucleoprotein complexes. J. Virol. 12:901-908.
10. Hsu, M.-T.,and P.Berg. 1978. Altering the specificity of restriction endonuclease: effect ofreplacingMg"2with
Mn"2.Biochemistry 17:131-138.
11. Hsu, M.-T., and N. Davidson. 1974. Electron microscope heteroduplex study of the heterogeneity of Mu phage andprophage DNA. Virology 58:229-239.
12. Hsu, M.-T., and J. Ford. 1977. Sequence arrangement of the 5'ends of simian virus 40 16S and 19S mRNAs. Proc. Natl. Acad. Sci. U.S.A. 74:4982-4985.
13. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature(London) 227:680-684.
14.Meinke, W.,M. R. Hall, and D. A. Goldstein. 1975. Proteins in intracellular simian virus 40 nucleoprotein complexes: comparison with simian virus 40 core pro-teins. J. Virol.15:439-448.
15. Penyim, S., and R. Chalkley. 1969. High resolution
polyacrylamide gel electrophoresis ofhistones. Arch. Biochem.Biophys.30:330-337.
16. Ponder,B. A.J.,F.Crew,and L. V. Crawford.1978. Comparison of nucleasedigestion of polyoma virus nu-cleoprotein complex and mouse chromatin. J. Virol. 25: 175-186.
17. Shure, M.,and J.Vinograd. 1976.The number of su-perhelical turns in native virion SV40 DNA and minicol DNAdetermined by the band counting method. Cell 8: 215-226.
18. Solari, A. J. 1974. The molecular organization of the chromatinfiber, p. 493-531. In H. Busch (ed.), Thecell
nucleus. Academic PressInc., New York.
19. Su, R.T.,and M. L.DePamphilis. 1976. In vitro repli-cation of simian virus40DNA in anucleoprotein
com-plex.Proc.Natl. Acad. Sci. U.S.A. 73:3466-3470. 20. Varshavsky,A.J.,V. V.Bakayev,and G. P.
Geor-giev. 1976. Heterogeneity of chromatin subunits in vitro and location of histoneHi.Nucleic Acids Res.3: 477-492.
21. Varshavsky, A. J., S. A. Neodospasov, V. V. Schmatchenko,V. V.Bakayev,P.M.Chumackov, andG.P.Georgiev. 1977. Compact formofSV40 viral minichromosome isresistant tonuclease: possible im-plicationsfor chromatin structure. Nucleic Acids Res. 4:3303-3325.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. 3.sucrosewithdescribedshownwere(b)labeling [:IH]lysine Profile of SV40 complexes in a 5 to 40%O gradient labeled with (a) [3H]thymidine and from 24 to 48 h postinfection and (c)12P from 36 to 48 hpostinfection](https://thumb-us.123doks.com/thumbv2/123dok_us/1518516.104451/5.505.275.466.484.669/sucrosewithdescribedshownwere-labeling-profile-complexes-gradient-thymidine-postinfection-hpostinfection.webp)

![FIG. 12.pulse-labeled[3H]thymidine.midinedium)extractcontainingcontaminatingtron Pulse-chaseof SV40complexeswith A total of 1O SV40-infected cells were with [3H]thymidine (1 mCi/5 ml of me- for 10 min (0) and chased with 100 mM thy- for 1 h (0)](https://thumb-us.123doks.com/thumbv2/123dok_us/1518516.104451/11.505.273.463.79.521/labeled-thymidine-midinedium-extractcontainingcontaminatingtron-chaseof-complexeswith-infected-thymidine.webp)
