JOURNAL OFVIROLOGY, Aug. 1991, p. 4408-4413 0022-538X/91/084408-06$02.00/0
CopyrightC 1991, American SocietyforMicrobiology
Functional
Analysis of Human T-Cell Leukemia Virus Type I
rex-Response Element: Direct
RNA
Binding of Rex
Protein
Correlates
with In
Vivo Activity
CLAUDIA BALLAUN,1G. KING FARRINGTON,2 MARIKADOBROVNIK,1JIMRUSCHE,2 JOACHIMHAUBER,1 AND ERNSTBOHNLEIN1*
SANDOZ Research Institute, Brunnerstrasse59, A-1235 Vienna, Austria,' andRepligen Corporation, One Kendall
Square, Cambridge,
Massachusetts 021392Received 15February1991/Accepted 9May1991
The human T-cell leukemia virus type I rex gene product plays a critical role in the expression ofthe retroviral structuralproteins Gagand Envfromincompletely splicedmRNAs. Rexproteinactsthroughacis
element(rex-response element [RxRE]) whichislocated in theU3/R regionof the3'longterminal repeatand ispresentonallhumanT-cell leukemia virustypeI-specificmRNAs. Two domainsofthepredictedsecondary
structureofthe RxREarecruciallyimportantforRexactioninvivoasmeasuredbytwoassaysystems.Invitro studies using highly purified recombinant Rex protein revealed a specific and direct interaction with
radiolabeled RxREsequences.Thecorrelation betweenourin vivo results and thedirectbindingofRexprotein
to mutantandwild-typeRxREsequencessupportsboth theexistence of the predicted secondarystructureand theimportanceof this direct interaction with thecis-actingRNA sequenceforRexfunctioninvivo.
Human T-cell leukemia virus type I (HTLV-I), the etio-logic agentof adultT-cell leukemia andHTLV-I-associated myelopathy (8, 18-20, 22, 27), is provided with a complex genomeorganization. Inadditiontothe retroviral structural genes gag, pol, and env, HTLV-I encodes at least two
regulatory proteins (Tax and Rex) in the 3' region of the genome (7, 26). Rexactsposttranscriptionally, allowing the cytoplasmic accumulationofincompletely spliced retroviral mRNAsencodingthe structuralgenes gag,pol, and env(9, 11, 12). Function of the rex gene product depends on a cis-actingsequence(rex-response element [RxRE]) which is locatedinthe U3/R region of the 3' long terminalrepeat (1, 9, 23, 25) andseparates thepolyadenylation signal from the site of polyadenylation (1). The in vivo formation of a
predicted, stable, complex RNA secondarystructure, which wassupported by in vitro solution mapping analysis(25),not
only would allowaproperspacing of these signalsinvolved
in polyadenylation but also could benecessaryforrexgene
function.
In this report, we describe the in vivo mapping of cis-actingsequences,within theRxRE, whichare necessaryfor wild-type Rex-mediatedtransactivation.We have generated
aseriesof5', 3', 5'/3', and internalRxREdeletionmutants.
By employing two in vivo assay systems, we defined two
critically important substructures within the RxRE. The establishment of a Rex-specific RNA gel mobility shift assay, using highly purified recombinant wild-typeHTLV-I protein, enabledustodirectlyassaytheseRxREmutantsin vitro. The observed correlation between Rex activity and specificdirect binding ofRex proteintoradiolabeled RxRE RNA in vitro supports the importance of the predicted secondary structure of this cis element. Furthermore, our
results support the notion that a direct interaction of Rex
protein with RxRE sequences is required for HTLV-I rex
function invivo.
*Corresponding author.
MATERIALS AND METHODS
Plasmid construction. The wild-type RxREwas amplified by polymerase chain reaction by using the followingprimers: 5'-aaaaaagatctGGCTCAATAAACTAGCAGGAGTCTAT AAAAGCGT-3' and 5'-aaaaaaagcttGAGCAAGCAGGGT CAGGCAAAGCGTGGAGAGCCG-3'. The BglII-HindIII polymerase chain reaction fragment was ligated into the BamHI-HindIII sites of pKSII(-) (Stratagene) to generate pKS-RxRE. The RxRE polymerase chain reaction
frag-mentwasalsoinserted into the BglII-HindIII-treatedpgTat
vector (15), replacing the rev-response element (RRE) se-quences and generating pgTat-RxRE (2). A SalI-BamHI restriction fragment derived from pgTat-RxRE encompass-ing the RxRE sequence was subsequently cloned into a human immunodeficiency virus type 1 (HIV-1) HXB-2-derived infectious proviral DNA. The 5',3', and5'/3' RxRE deletion mutants were cloned by using the appropriate primers. The various mutants are listed in Table 1 and depicted in Fig. 1B. The internal deletion mutants were generated by oligonucleotide-directed mutagenesis (24) by
using the bacteriophage M13 mutagenesis system (Amer-sham). All mutantswere characterized by restriction analy-sis and dideoxy DNA sequencing (Sequenase 2.0; USB). Themutantfragments were ligatedasBamHI-HindIII
frag-ments into pKSII and pgTat and subsequently into the HXB-2-derived HIV-1 proviralplasmid.
Transfection and in vivoassays. ThepgTAT-RxRE DNAs (wild type and mutants) were cotransfected with a rex expression plasmid (2) into COS cells as described previ-ously (3). The immunoprecipitations and gel analyses were performed by the methods of Hauber et al. (10). HIV-1 proviral DNAs (1
jig)
were cotransfected with an HTLV-Irexexpression plasmid (0.5 ,ug) (pLHRex; 2) into 5 x 106 COS cells. Supernatants were harvested 70 h posttransfec-tion and analyzed for HIV-1 p24 Gag expression by an enzyme-linkedimmunosorbentassay (ELISA) (Coulter).
Protein purification and in vitro assays. Expression and purification of Rex protein will be described in detail in futurepublications(5a, 6). Briefly,theHTLV-Irexgene was
4408
Vol.65,No. 8
on November 10, 2019 by guest
http://jvi.asm.org/
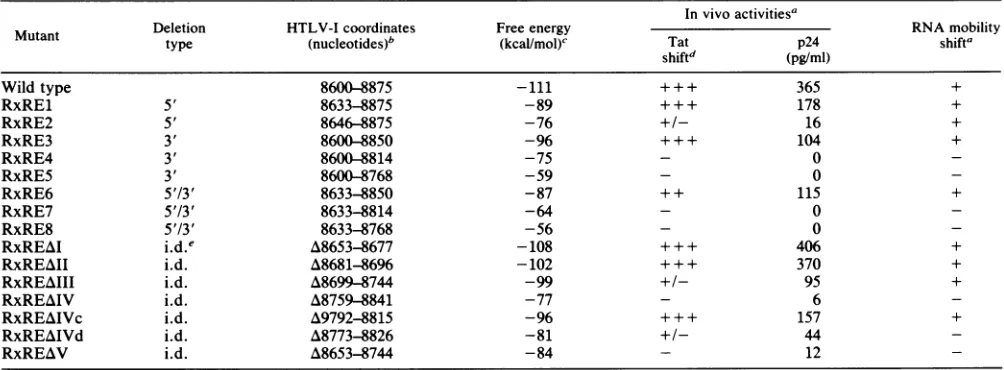
HTLV-I Rex/RxRE RNA BINDING IN VITRO 4409 TABLE 1. Descriptionof mutants
Invivoactivitiesa
Deletion HTLV-Icoordinates Free energy RNAmobility
Mutant type
(nucleotides)b
(kcal/mol)' Tat p24 shiftashiftd (pg/ml)
Wild type 8600-8875 -111 +++ 365 +
RxRE1 5' 8633-8875 -89 +++ 178 +
RxRE2 5' 8646-8875 -76 +/- 16 +
RxRE3 3' 8600-8850 -96 +++ 104 +
RxRE4 3' 8600-8814 -75 - 0
-RxRE5 3' 8600-8768 -59 - 0
-RxRE6 5'/3' 8633-8850 -87 + + 115 +
RxRE7 5'/3' 8633-8814 -64 - 0
-RxRE8 5'/3' 8633-8768 -56 - 0
-RxREAI i.d.e A8653-8677 -108 +++ 406 +
RxREAII i.d. A8681-8696 -102 +++ 370 +
RxREAIII i.d. A8699-8744 -99 +/- 95 +
RxREAIV i.d. A8759-8841 -77 - 6
-RxREAIVc i.d. A9792-8815 -96 + + + 157 +
RxREAIVd i.d. A8773-8826 -81 +/- 44
-RxREAV i.d. A8653-8744 -84 - 12
-aThein vivoTatshift andp24(ELISA)activities and in vitro bindingresultsare
described.
bTheHTLV-Icoordinates (13) arelistedfor the end-pointdeletions.TheHTLV-1coordinates of thedeleted sequencesare also indicatedfortheinternal
deletionmutants.
Thepredicted stability ofthe RxRE sequences is shown.
dActivity ranges from wildtype (+++) to undetectable (-).
ei.d.,internaldeletion.
expressed in Escherichia coli by translational coupling to the E. coli ,B-glucuronidase gene on a high-copy-number plas-mid, resulting in recombinant protein with the authentic HTLV-I amino acid sequence. The bacterial cells were harvested, lysed, and centrifuged, and the pellet was ex-tracted with 8 M urea in 50 mM MES [2-(N-morpholino) ethanesulfonicacid] (pH 6.5). The solubilized proteins were then subjected to S-Sepharose chromatography and gel filtration. Less-pure fractions were further purified by re-verse-phase chromatography on a C-18 column. Protein
purity was analyzed by sodium dodecyl
sulfate-polyacryla-midegel electrophoresis.Radiolabeled RNA (1.5 x 104 cpm) wasincubated with 10 U of RNasin, 150 gigof yeast tRNA
(Boehringer Mannheim), and 1% glycerol in
phosphate-buffered saline for10 minat30°C.The Rexproteinwasthen
added to the binding reaction mixture and incubated for
another 10 min (final volume, 10 ,ul) at 30°C. The reaction
mixtures were loaded onto 5% native polyacrylamide gels (29:1, wt/wt) andelectrophoresed at5 mA, driedon What-man 3MM paper, andexposedto Kodak XAR X-ray films. For the competition experiments, radiolabeled in vitro
tran-scripts and a molar excess ofnonradiolabeled RNA were added tothebinding reaction mixture and incubated for 10 minpriortotheadditionofRexprotein.
RESULTS
In vivo mapping of functional domains within the RxRE sequence. Todefine functional domains in theHTLV-IRxRE
element, wemolecularlycloned andmutagenizedthe RxRE sequence.Various5', 3', 5'/3', and internal deletionmutants werecreated(detaileddescriptioninFig. 1and Table 1) and inserted intoanRRE(5, 16)-deficientgenomicTatindicator
plasmid (15) to generate pgTat-RxRE plasmids (wild type and mutants). In the in vivo assay utilized, a functional RxRE sequence in the presenceofacotransfectedwild-type
rexgeneresults in the formationofthe72-amino-acid(72-aa)
form of the HIV-1 Tat protein (9, 15) as detected by
immunoprecipitation(Fig. 2, lane 2). The 5'deletion mutant RxRE1 (Fig. 2, lane 3), the 3' deletionmutant RxRE3 (Fig.
2, lane 5), the 5'/3' mutant RxRE6 (Fig. 2, lane 8), and the
internal deletionmutants RxREAI, -All, and -AIVc (Fig. 2, lanes 11, 12, and 15) displayed a phenotype comparable to thatofwild-typeactivity (Fig.2, lane2) in this assay system. Transfection of pgTat-RxRE2 and the internal deletion mu-tants pgTat-RxREAIII and pgTat-RxREAIVd only poorly supportedRexfunction(Fig.2, lanes 4, 13, and 16). All other mutants failed to yield detectable levels of the indicator 72-aa Tatprotein(Fig.2 andTable 1). Toanalyze the various RxRE mutants more quantitatively, we constructed
infec-tious HIV-1 proviral clones in which the HIV-1 RRE was
replacedby the variousRxREsequences. With these
provi-ral constructs, thesynthesis ofthe HIV-1p24Gag polypep-tiderequiredthe presenceoffunctionalHTLV-IRexprotein (data not shown) and was quantitated by using an HIV-1
Gag-specific ELISA (Coulter). The results of this assay
(Table 1) supported the mapping data obtained in the Tat shift assay (Fig. 2).
PurifiedRexproteinspecifically binds RxRERNA in vitro. The interaction of HIV-1 Rev protein with the RRE
sug-gested a similar direct interaction of Rex protein with its
HTLV-I cis-acting element which, like the RRE, has the
potentialtoformastablesecondarystructure(Fig. 1A) (1, 9,
25, 28). To test this hypothesis directly, we expressed
HTLV-I Rexprotein in E. coli(6). Thepurity ofthe
protein
was analyzed in denaturing polyacrylamide gels (data not
shown) andWestern blots(immunoblots) (Fig. 3B, lane 1).
Incubation ofRexprotein withradiolabeled HTLV-I RxRE
transcripts resulted in a clearly retarded mobility of the
complex(Fig. 3A,lane2)ascomparedtofree RNA
(Fig. 3A,
lane 1). Immunodepletion of the protein
preparation
with aRex-specific rabbit antiserum (2)resulted in the loss of the
ability to form complexes with radiolabeled RxRE RNA
(Fig. 3A, lane 3). The addition of
protein
A agarose alone(Affi-Gel; Bio-Rad) had noeffecton the complex formation
VOL.65, 1991
on November 10, 2019 by guest
http://jvi.asm.org/
4410 BALLAUN ET AL.
A
1I4tli..4114I
S"14.t11 144441|}11 4
f
t.
,
A
S1i.
x
-t (
('1\11II i BB
'%It-11114,4 )1
/ \
%f.is \ FA I
ts-0
{,-\
I-- e.
1,-,
\_
w...t_ \ t
{,l
\
{,_
_
t, 91tsill \ t,=
4.-I
t
s.s.5
!e
\
1l\
Xs
I
\-l
I
l,<|
4
\I
t|
%#.|q: %,-:
5'- RxRE deletion mutants
Mlutant RxRE 1
SL II ,!
Mutant RxRE 2
SL II
SL IV
SL I
3'-. RxRE deletion mutants
Mutant RxRE 3
SL I I s
Mutant RxRE 4 SL II
SL III
SL IV
SL I
Mutant RxRE 5 SL III
5'/3'- RxRE deletion mutants
Mutant RxRE 6 Mutant RxRE 7
SL II
SL III SL III
SLI
IV
SLMutant RXRE 8
SL II
°
ISL
IIISLL I
B
J. VIROL.
I
on November 10, 2019 by guest
http://jvi.asm.org/
HTLV-I Rex/RxRE RNA BINDING IN VITRO 4411 FIG. 1. (A) Schematic outline of the predicted secondary structure ofHTLV-I RxRE. The depictedHTLV-I sequence extends from nucleotide 8600 to nucleotide 8879 (14). The cap and splice donor sites are indicated, as are the various predicted substructures (28) (stems AandB; stem-loops I through IV). The internal deletion mutants(RxREAI, RxREAII, RxREAIII, RxREAIV, RxREAIVc,RxREAIVd,and RxREAV) are indicated by different shades of gray. (B) Schematic outline of the RxRE mutants. The5',3', and5'/3'RxRE deletion mutants areoutlined. Thedeleted sequenceshave been omitted from the wild-type structure (see panel A). The displayed structures of the RxRE mutantsdo not correspond with the most stable predicted secondary configurations (28) but were depicted in this manner to highlight the substructures whichareaffected by the specific deletions.
(Fig. 3A, lane 4). The untreated protein preparation con-tained a Rex-immunoreactive species in Western blot anal-yses (Fig. 3B, lane 1). However, the immunodepleted prep-aration which failed to complex radiolabeled RxRE RNA contained no detectable amounts of HTLV-I Rex (Fig. 3B, lane 2), while the pellet of the immunoprecipitation and the
Affi-Gel-treatedpreparation clearly yielded an immunoreac-tive protein of the correct molecular weight (Fig. 3B, lanes 3 and4).
The observed Rex-RxRE complex formation was depen-dent on the protein concentration (Fig. 4, lanes 2 to 5). In contrast, purified HIV-1 Rev protein, previously shown to specifically interact with HIV-1 RRE sequences (5, 17), did not bind to the heterologous HTLV-I RxRE sequences under identical experimental conditions (Fig. 4, lanes 7 to 10). Furthermore, increasing amounts of nonradiolabeled RxRE in vitro transcripts specifically competed with the
complex formation (Fig. 5, lanes 3 to 5). Additionof equal amounts of nonspecific HIV-1 RNA, which could not sup-port Rexfunctioninvivo (datanotshown),didnotaffect the RxRE RNA-protein complex (Fig. 5, lanes 6 to 8).
Direct binding of Rex protein to mutant RxRE in vitro transcripts. The Rex-specific RxRE-RNA gelmobility shift
assay permitted us to analyze our series of RxRE mutants
for direct binding in vitro. The synthesized RNAof RxRE mutants 1, 2, 3, and 6 formed complexes with the purified
Rex protein comparable to those of the wild-type RxRE
(Table 1). However, the RxRE mutants 4, 5, 7, and 8failed
to yielddetectable RNA-protein complexes under the same
experimental conditions (Table 1), which correlateswith our invivoresults. Invitro binding studiesof the RxRE internal deletion mutants also correlated with the observed activity in vivo (Table 1). Mutants which had failed to form com-plexes with purified Rex protein also did not compete for the binding of Rex protein to wild-type RxRE RNA (data not shown).
DISCUSSION
Although the trans regulatory proteins of HTLV-I and HIV-1 share no obvious amino acid homology, therexgene productcanfunctionally substitute for Rev action (21). This
observation suggests thatbothtrans-acting proteins sharea common functional mechanism mediated by cis-acting se-quences(4);the HTLV-I Rexactivity dependsonthe RxRE sequence and the HIV-1 Rev functiondepends on the RRE sequence (9, 16). We generated a series of site-directed RxRE mutants todefine importantdomains in the predicted secondary structure of the HTLV-I RNA element. Our in vivo experiments defined two independent domains within the RxRE. Our data (Fig. 1A) and the results recently
reported by Toyoshima et al. (25) map one essential cis elementto a structureonstem-loop IV, whichalso coincides with the sequence requirements determined by Ahmed and
colleagues (1). An additional domain is located on the
cloverleaf structure comprising stem-loops I, II, and III (mutant RxREAV), which is in agreement with a previous
report(25). Intriguingly, deletion oftheindividual
substruc-- 43
1 2 3 4 5 6 7 8 9 10 1 l 12 13
___R__S.
wFw
=
*
Xa
...
r.. ,_wt_ _S? _ _ _
.
E __
-L _t
tffi_w_+__56w7<;
. ._ .7_- - sZ s^
86aa Tat
72aa
- 29
- 18,4
- W
4_
-_b
- m - 14,3-P -P
:k, ;k'
;k~
!k
I~--k
-k
"
k
k~
Op1k
k
K=K
I(C)
K;
K
.,K K K K KK K
K
KK K K K
C $+,7 V
+Rex
FIG. 2. In vivo analysis ofthe RxRE mutants by immunoprecipitation. Thevarious RxRE constructs (indicated atthe bottom) were
cotransfected withachloramphenicol acetyltransferase expression plasmidorpLHRex(2). Thepositions ofthe86-and 72-aaformsof the HIV-1 Tatprotein and molecularmassstandards(in kilodaltons) areindicated.
lbs. wli.ld&.-
I--W-wr ijlllllllll. .411M
VOL. 65,1991
V1-14 1 5 1 6 17 ,*.,A
1%
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.104.494.476.697.2]4412 BALLAUN ET AL.
1 2 3
4
56
7 82 3 4
46-30 4ow
__
_10 Rex
21-
14-FIG. 3. (A) HTLV-I Rex protein specifically interacts with RxRE RNA. Rex protein (1 ,uM; 10-,u reactionvolume) causes a
distinct mobility shift (lane 2) compared to free RNA (lane 1) in nondenaturing 5% polyacrylamide gels. Rex-depleted protein
prep-arations failtoform specific complexes(lane 3). Pretreatment with Affi-Gel has noeffect onthe complex formation (lane 4). Arrows
indicate the positions of free and complexed RNA. (B) Western blot analysis of the Rex proteinpreparation. Rex-immunoreactive poly-peptidescanbedetected in lanes 1 (untreated Rex protein), 3 (pellet of the Rex-depleted preparation), and 4 (Affi-Gel-treated protein preparation). Noimmunoreactive proteincanbedetected in lane 2
(anti-Reximmunoprecipitatedprotein preparation). Molecularmass
standards (inkilodaltons) areindicatedontheleft.
ture stem-loop IorII displayed wild-type activity in both in
vivo testsandthe RNAgelmobility shift assay. Compared
to these mutants, RxREAIII yielded little 72-aaTatprotein andsignificantly reduced p24 Gag values, suggesting that the functional domainmustpartly overlap with stem-loop III. Of
note, mutation of the conserved bulge sequence (CUCAG GUCGAG; nucleotides 8783to8793; seeFig. 1), whichwas crucially important for HTLV-I Rex function (1), also abro-gated HTLV-II Rex responsiveness in vivo (13). Intrigu-ingly, stem-loop III contains an identical bulge sequence (GUCG; nucleotides 8715 to 8718; Fig. 1), which might
represent a functional site of interaction in this region. Experimental results obtained with mutant RxRE2 further indicate that thepresentation of the bulgesequencemight be important. Although all necessary nucleotides are present
(Fig.1andTable1), themoststableconfiguration of RxRE2 isnotpredictedtoform the cloverleafstructure(28;datanot
1 2 3 4 5 6 7 8 910
FIG. 4. Protein titration of the Rex-RxRE complex formation.
Increasing amountsofrecombinant Rexprotein (0.3, 0.6, 1.2,and
1.8 p.M; 10-p.I reaction volume) result in Rex-RxRE complexes
(lanes 2 to5). Addition of identical amounts of recombinant Rev
protein (5) failed to yield any detectable complexes with
radiola-beled RxRE RNA(lanes7to10). Lanes 1 and 6 show themigration
of RxRE RNA in the absence of protein. The positions of the
complexesandfree RNA areindicatedbyarrows.
I
RXRE
unrelated
RNA
FIG. 5. RNAgel mobility shiftcompetition experiment. Nonra-diolabeled RxRE RNA(lanes3to5)andanunrelatedHIV-1 RNA (HIV-1HXB-3 nucleotides 6619to7202; 5) (lanes6to8)wereadded
tothebindingreaction mixtureattheindicated molarexcess.The positionsof freeRxRE RNA(lane 1)andRex-complexedRxRE RNA (lane2) in the native 5% polyacrylamide gelareindicated byarrows.
shown), which could explain the failure of this RxREmutant tosupportRex actionin vivo. The structural stability of the various RxREmutantsmight also influence theirphenotype
in vivo. However, the observed difference in biological
activity between RxREAIII andmutantRxREAV isunlikely
tobetotally accounted forby the loss of stability (Table 1).
Other RxREmutants(e.g., RxRE1 and RxRE6), which form the cloverleaf structure and have predicted free-energy
valuesverysimilartothat ofRxREAV,supportRex-mediated
transactivationsignificantly better than RxREAV (Table 1).
Toassessthecapability of the various RxREsequencesto interact withRex protein directly, weestablished an RNA gel mobility shift assay using purified recombinant Rex protein(6). Several lines of evidence supportthespecificity of the observed Rex-RxRE complex formation. (i) Rex-specific depletion of the protein preparation abrogated the Rex-RxRE complex formation (Fig. 3A) which was inter-nally controlled by Western blot analysis (Fig. 3B). (ii) Rex-RxRE complex formationoccurredinadose-dependent manner, whereas purified recombinant HIV-1 Rev protein (5) failedtoform specific complexes under identical experi-mental conditions(Fig. 4). (iii) Nonradiolabeled RxRE RNA specifically reduced theRex-RxREcomplexes(Fig. 5), while nonrelevant HIV-1 RNA did not abrogate the Rex-RxRE complex formation (Fig. 5). Of note, all Rex function-supportingRxRE mutantsspecifically interacted with wild-type Rex protein in vitro (Table 1). Although it is still possible that Rexproteinisinvolved inRNA3'processing,our experiments clearly indicate that binding of Rex protein to RxRE RNAoccurs intheabsenceofpolyadenylation signals. The correlation betweenourin vivoexperimentsandour in vitrobindingstudiesindicates thatthe interactionofRex protein with RxRERNA isaprerequisitefor Rex-mediated
trans activation in vivo. Itfurthersuggeststhat the putative
A B
1 2 3 4
.I.
zk.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.76.305.58.173.2] [image:5.612.361.529.75.301.2] [image:5.612.100.276.516.645.2]HTLV-I Rex/RxRE RNA BINDING IN VITRO 4413 cellular factors(s) involved in theRev- and Rex-dependent
transport of
incompletely
spliced RNA to the cytoplasmmight
interact with the RNA-protein complex. Only theresultsobtained withmutantRxRE2werenotconsistent. As
already discussed,
thismutantisunlikelytoformthe clover-leafstructure(28; datanotshown)andisrelativelyunstable(free
energy,-76kcal/mol;
Table1). However,itispossiblethat our in vitro reaction conditions partly support the cloverleaf
configuration
while the probability thatthissec-ondary
structure is formed in vivo is less likely.Alterna-tively,
asecondary
eventfollowing
Rex-RxREcomplexationis
required
to support Rexaction in vivo.In
conclusion,
ourin vivo studies definedtwofunctionallyimportant
substructures in the RxREcis-actingelement. Theprecise
localization and the sequencerequirements
of these domainsarecurrently
underinvestigation.
Furthermore, we haveestablishedaspecific
Rex-RxREinvitrobindingassay. For the firsttime, purified
recombinant HTLV-I Rexproteinwas shown to
specifically
binddirectly
to RxRE RNA in vitro. The observed correlation between the in vivo and in vitroexperiments
supports the notion that binding of Rexprotein
to the RxRE RNA is necessaryfor Rex action. Inaddition,
themapping
offunctionally important
domains tosubstructures in the RxRE
(RxREAIV
and RxREAV)indi-catesthe
importance
ofthepredicted secondary
structureinvivo.
Hence, overexpression
ofoptimized
RxRE RNA or chimeric RxREtranscripts
could be used in somatic genetherapy
approaches
directedagainst
HTLV-I.ACKNOWLEDGMENTS
Wethank Jan-MarcusSeifertand FranzAmbergerfor oligonucle-otidesynthesis,BryanCullenandMichaelMalimfortherevgene, and WarnerC. Greene fortherexgene. Wealso thankourcolleague
Helmut Bachmayer for critical comments and Barbara Falle and Martina Ruhlfor excellent secretarial assistance.
REFERENCES
1. Ahmed, Y.F., S. M. Hanly,M. H. Malim,B. R. Cullen, and W.C. Greene.1990.Structure-functionanalysesof the HTLV-I Rex and HIV-1 Rev RNA responseelements: insightsinto the mechanism ofRex and Rev action. GenesDev.4:1014-1022. 2. Bohnlein, S., F. P. Pirker, L. Hofer, K. Zimmermann, H.
Bachmayer, E. Bohnlein, and J.Hauber. 1991. Transdominant repressors for human T-cell leukemia virus type I Rex and human immunodeficiency virus type 1 Rev function. J. Virol. 65:81-88.
3. Cullen, B. R. 1986. Trans-activation of human
immunodefi-ciencyvirus occursviaabimodalmechanism. Cell46:973-982. 4. Cullen, B. R., and W. C. Greene. 1989. Regulatory pathways
governingHIV-1replication. Cell 58:423-426.
5. Daly, T. J., K. S. Cook, G. S. Gray, T. E. Maione,and J. R. Rusche. 1989. HIV-1recombinantRevproteinbindsspecificallyto
the Rev responseelementin vitro.Nature(London)342:816-819.
Sa.Farrington,G.K.,R.Doten,C.
Bailaum,
E.Bohnlein,andJ.R. Rusche. Unpublisheddata.6. Farrington, G.K.,P. Lynch,A.Jensen,E.Bohnlein,R.Doten,
T.Maione,T.Daly,andJ.R. Rusche. 1991. InImmunobiology
of
proteins
andpeptides VI,inpress.7. Gallo,R.C.,F.Wong-Staal,L.Montagnier,and M. Yoshida.1988.
HIVMHTLV-I
genenomenclature. Nature(London)333:504. 8. Gessain, A., F. Barin,J. C. Vernant, 0. Gout, L. Maurs, A.Calender,andG. deThe.1985.Antibodiestohuman T-lympho-tropicvirustype-1in patients withtropical spastical parapare-sis. Lancet ii:407-410.
9. Hanly,S.M.,L. T.Rimsky,M. H.Malim,J.H.Kim,J.Hauber,
M. DucDodon,S.-Y.Le,J.V. Maizel,B. R.Cullen,and W.C. Greene. 1989. Comparative analysis of the HTLV-I Rex and HIV-1 Rev trans-regulatory proteinsand their RNA response elements. GenesDev. 3:1534-1544.
10. Hauber, J., A. Perkins, E. P. Heimer, and B. R. Cullen. 1987. Trans-activation ofhuman immunodeficiencyvirusgene expres-sion ismediatedbynuclearevents.Proc.Natl.Acad.Sci. USA 84:6364-6368.
11. Hidaka, M., J. Inoue, M. Yoshida, and M. Seiki. 1988. Post-transcriptionalregulator (rex)of HTLV-I initiates expressionof viral structural proteins but suppresses expression of regulatory proteins. EMBO J. 7:519-523.
12. Inoue, J.-I., M. Yoshida, and M. Seiki. 1987. Transcriptional (p4Ox) and post-transcriptional (p27x-III) regulators are re-quired for the genes.Proc. Natl. Acad.Sci. USA 84:3653-3657. 13. Kim, J. H., P. A. Kaufman, S. M. Hanly, L. T. Rimsky, and W. C. Greene. 1991. Rex transregulation of human T-cell leukemia virus type II geneexpression. J. Virol. 65:405-414. 14. Malik, K. T. A., J. Even, and A. Karpas. 1988. Molecular
cloning and complete nucleotide sequence of an adult T cell leukaemiavirus/human t cell leukaemia virus type I (ATLV/ HTLV-I) isolate of Caribbean origin: relationship to other members of the ATLV/HTLV-I subgroup. J. Gen. Virol. 69: 1695-1710.
15. Malim, M. H., J. Hauber, R. Fenrick, and B. R. Cullen. 1988. Immunodeficiencyvirus rev trans-activator modulates the activity of the viralregulatory genes. Nature (London)335:181-183. 16. Malim, M. H., J. Hauber, S.-Y. Le, J. V. Maizel, and B. R.
Cullen. 1989. The HIV-1 rev trans-activator acts through a structured target sequence to activate nuclear export of
un-splicedviral mRNA. Nature(London) 338:254-257.
17. Malim, M. H., L. S. Tiley, D. F. McCarn, F. R. Rusche, J. Hauber, and B. R. Cullen.1990. HIV-1 structural gene expres-sion requires binding of the Rev trans-activator to its RNA target sequence.Cell 60:675-683.
18. Osame, M., K. Usuku, N. Uichi, H. Amitani, A. Igata, M. Matsumoto, and H. Tara.1986. HTLV-I associated myelopathy, a newclinicalentity. Lancet i:1031-1032.
19. Poiesz, B. J., F. W. Ruscetti, A. F. Gazdar, P. A. Bunn, J. D. Mihna, and R. C. Gallo. 1980. Detection and isolation of type C retrovirusparticlesfrom fresh cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 77:7415-7419.
20. Poiesz, B. J., F.W. Ruscetti, M. S. Reitz, V. A. Kalyanamaran, and R. C. Gallo. 1981. Isolation of a new type C retrovirus (HTLV) in primary uncultured cells of apatient with Sezary T-cell leukemia. Nature(London) 294:268-271.
21. Rimsky, L. T., J. Hauber, M. Dukovich, M. H. Malim, A. Langlois, B. R. Cullen, and W. C. Greene. 1988. Functional replacement of the HIV-1 Rev protein by the HTLV-I Rex protein. Nature(London)335:738-740.
22. Seiki, M., S. Hattori, Y. Hirayama, and M. Yoshida. 1983. Human adult T-cell leukemia virus: complete nucleotide se-quenceof theprovirusgenomeintegratedin leukemia cell DNA. Proc. Natl. Acad. Sci. USA80:3618-3622.
23. Seiki, M., J.-I. Inoue, M.Hidaka, and M. Yoshida. 1988. Two cis-acting sequencesresponsible for post-transcriptional
trans-regulation ofgene expression ofhumanT-cell leukemia virus type I. Proc. Natl. Acad. Sci. USA85:7124-7128.
24. Taylor, J. W., J.Ott,and F. Eckstein. 1985. Therapid genera-tion of oligonucleotide-directed mutations at high frequency using phosphorothioate-modified DNA. Nucleic Acids Res. 13:8765-8785.
25. Toyoshima, H., M.Itoh, J.-I. Inoue, M. Seiki,F. Takaku, and M. Yoshida. 1990. Secondary structure of the human T-cell leukemia virustype1rex-responsiveelementis essential forrex
regulationof RNAprocessingand transportofunsplicedRNAs. J. Virol. 64:2825-2832.
26. Wong-Staal, F.,and R. C. Gallo. 1985. HumanT-lymphotropic retroviruses. Nature(London)317:395-403.
27. Yoshida, M., I. Miyoshi, and Y. Hanuma. 1982. Isolation and characterization of retrovirus from cell lines of human adult T-cell leukemiaandits implication in the disease. Proc. Natl. Acad. Sci. USA 79:2031-2035.
28. Zuker,M., and P. Stiegler. 1981. Optimal computerfoldingof largeRNA sequencesusing thermodynamicandauxiliary infor-mation. Nucleic Acids Res. 9:133-148.
VOL. 65,1991