Copyright © 1992,AmericanSociety forMicrobiology
Identification and
Characterization of
a
Neutralization Site within
the Second
Variable
Region
of Human
Immunodeficiency
Virus
Type 1
gpl20
MICHAEL S. C. FUNG,1* CECILY R. Y.
SUN,'
WAYNE L.GORDON,'
RUEY-SHYANLIOU,1
TSE WEN
CHANG,'
WILLIAM N. C.SUN,'
ERIC S. DAAR,2AND DAVID D. HO3 TanoxBiosystems, Inc., 10301 Stella Link, Houston, Texas770251;
Division ofInfectious Diseases,Cedars-Sinai Medical Center, University of California at LosAngeles, Schoolof Medicine, LosAngeles, California 900482; and Aaron Diamond AIDS Research Center,
New York University School of Medicine, New York, New York
100163
Received 13 August 1991/Accepted 31 October 1991
Twomonoclonal antibodiesdesignated BAT085 and G3-136 were raised by immunizing BALB/c mice with gpl20purified from humanimmunodeficiencyvirustype1(HIV-1)IIIB-infected H9 cell extracts.Among three HIV-1 laboratory isolates (IIIB, MN, and RF), BAT085 neutralized only IIIB infection ofCEM-SS cells, whereasG3-136 neutralized both IIIB and RF. These antibodies also neutralized a few primaryHIV-1isolates inthe infection ofactivated human peripheral blood mononuclear cells.Inindirect immunofluorescenceassays, BAT085 bound to H9 cells infected with IIIB or MN, while G3-136 bound to H9 cells infected with IIIB orRF, but not MN.Usingsequence-overlapping synthetic peptidesof HIV-1 IIIBgpl20, thebinding site of BAT085 and G3-136 wasmapped to apeptidicsegment in the V2 region(amino acid residues 169 to 183). The binding of these antibodies to immobilized gpl20 was not inhibited by the antibodies directed to the principal neutralization determinant in the V3 region or to the CD4-binding domain of gpl20. In a competition enzyme-linkedimmunosorbent assay, soluble CD4 inhibited G3-136 but not BAT085 from binding togpl20. Deglycosylation of gpl20 by
endo-o-N-acetylglucosaminidase
H or reduction of gpl20 by dithiothreitol diminished its reactivity with G3-136 but not with BAT085. These results indicate that the V2 region of gpl20 contains multiple neutralization determinants recognized by antibodies in both a conformation-dependent and -inconformation-dependent manner.Intensive efforts in AIDS research have been focused on the development ofan effective vaccine for protection of individualsatrisk for humanimmunodeficiencyvirus type 1 (HIV-1)infection(4, 45). It has becomeevidentthathumoral responses play crucial rolesin protective immunity against infection byHIV-1 and itsprogression to disease states. In thisrespect, Wendleretal. (49) showedthathigher neutral-izingantibody titers correlated with better prognosisand low or decreasing neutralizing antibody titers correlated with diseaseprogression. Jacksonetal.(19) and Karpas etal. (22, 23) demonstrated that AIDS-related complex and AIDS patients treated with hyperimmune plasma from asympto-matic HIV-1-seropositive individuals showed clinical im-provement, which correlated with reduction in HIV-1 vire-mia. More recently, preliminary studies by Emini et al. (7) showed that chimpanzees injected with a mouse-human chimeric HIV-1-neutralizing monoclonal antibody (MAb) were protected from infection by the virus.
The envelope proteins of HIV-1, gp120 and gp4l, are knowntobe the major targets forHIV-1-neutralizingactivity inthe seraofHIV-1-infectedindividuals (13, 18, 33, 48) and of animals immunized with the viral proteins (3, 28, 40). Several neutralization sites have been identified in
gpi20,
including an immunodominant epitope in the V3 region (amino acid residues 308 to 322), commonly known as the
principal
neutralization determinant(PND) (9, 21, 24, 27, 32, 43, 44). Antibodies raised against this continuous epitope*Correspondingauthor.
usually exhibit
type-specific
neutralizing activityagainst
the homologous viral isolates (11, 12, 33, 40, 43). However, recentstudies byJavaherianet al. (20) andWhite-Scharfet al. (50)showed thatantibodies raised inanimals againstthe relatively conservedportion
ofthe PND displayed broad-ened reactivity with multiple HIV-1 isolates. Anti-PND antibodies do not block virusbinding
to CD4 but prevent virus internalization subsequent to CD4 binding (31, 44). Anothercontinuousneutralization siteongpl20is located in the C4 region (amino acid residues 413 to 447), which is found to be involved in CD4-gpl20 interaction (29, 47). Antibodies tothis region exhibit broad neutralizing activity against divergentHIV-1 isolates, but this neutralization site appears to be ratherimmunosilentinHIV-1-infected individ-uals(47).Sera from most HIV-1-infected individuals usually have low levelsof broad neutralizingactivityagainst HIV-1 (2, 13, 48), which were found to be associated with antibodies directed to conformational or discontinuous epitopes on gpl20 (13). Consistentwiththis finding, we havereporteda human MAb
(I5e)
which recognizes adiscontinuousepitope ongpl20 involved in CD4binding andin antibody neutrali-zationofmultiple HIV-1 isolates (16). We also generated a HIV-1-neutralizing murine MAb(G3-4),
which identified anotherdistinctconformational epitopeon gpl20involved in theinteraction with CD4(15).Inthisreport, wedescribe thegeneration and characteri-zationoftwomurine MAbsdesignatedBAT085andG3-136 which defineaunique neutralization domain in the V2 region ofgpl20.
848
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALSAND METHODS
MAb reagents and polyclonal antibodies. Murine HIV-neutralizing MAb BAT123 immunoglobulinGl(K) [IgGl(K)] bindstothe PND inthe V3 region ofIIIB
gpl20
(amino acid residues 308 to 322) (32). Murine MAb G3-519[IgGl(K)]
recognizesa component of the CD4-binding domain in the C4
region
ofgpl20 (amino acid residues 423 to 437) (47). Murine MAb G3-4 [IgG2a(K)] recognizes a conformational neutralizing epitope on HIV-1gp120(15).I5e
[IgGl(K)] is aHIV-1-neutralizing
human MAb (a gift from J.Robinson,
Louisiana State University School of Medicine, New
Or-leans)
which also recognizes a distinct conformationalepitope
on gpl20 (16). The murine MAbs were produced fromascitic fluid of BALB/c mice(Harlan Sprague-Dawley, Inc., Houston, Tex.) injected with the hybridomas andaffinity
purified by using recombinant protein A-Sepharose(Repligen,
Boston, Mass.). I5e was produced from culture supernatant of the hybridoma and purified by protein Aaffinity chromatography.
Polyclonal anti-HIV-1gpl20
anti-bodies (D6205) (International Enzymes, Inc., Fallbrook,Calif.)
weregenerated by immunizing
sheep withasyntheticpeptide encompassing
ahighly
conservedpeptidic segment(APTKAKRRVVQREKR)
in the carboxy terminus of HIV-1(BH-10strain)
gpl20
(amino acid residues 497to511)(35-37).
The antibodieswereisolated from the sheep hyper-immune seraby affinity
chromatography usingthe specifiedsynthetic peptide coupled
toSepharose.
Cells and HIV-1 isolates. CEM-SS
cells,
asyncytium-sensitive clone of CEM cells (a
gift
from P. Nara, National CancerInstitute),
were maintained in RPMI 1640 medium(GIBCO Laboratories,
GrandIsland,
N.Y.)supplemented
with
10%
heat-inactivated fetal bovine serum(GIBCO), 100 Uofpenicillin
perml,
and100p.g
ofstreptomycin
per ml.H9 cellschronically
infected withHIV-1IIIB, MN,orRF,were maintained inRPMI 1640medium with15% heat-inactivated fetal bovine serum.These infected cells were derivedfromsingle-cell
clones which were tested to be almost 100%expressing
gpl20
on the cell surfaceby
immunofluorocyto-metric methods. The culturesupernatants fromthese HIV-1-infected cells
containing high
reversetranscriptase
activi-ties werekept
at -160°C
and used forHIV neutralization assays.Peripheral
blood mononuclear cells(PBMCs)
were isolated fromHIV-1-seronegative
donorsby
Ficoll-Paque
density
gradient centrifugation,
and were stimulated for 3days
with 2jig
ofphytohemagglutinin (PHA) (Sigma
Chem-ical
Co.,
St.Louis,
Mo.)
per ml in RPMI 1640 mediumsupplemented
with15% heat-inactivated fetal bovineserum, 10% human interleukin-2(Cellular Products, Inc.,
Buffalo,
N.Y.),
100 Uofpenicillin
perml,
and100 ,ugofstreptomycin
perml. After
stimulation,
PBMCs were washed and main-tained in thecomplete
growth
mediumforinfectivity
assays.
Primary
HIV-1isolates(LS,
RP,
CC,
LL,
TB,
andAC)
wereobtained from
plasma
specimens
fromHIV-1-seropositive
individuals,
aspreviously
described(5, 16,
17).
TheJR-CSF isolate waskindly
provided by
I. Chen(University
of California at LosAngeles,
School ofMedicine,
LosAnge-les).
Generation and
screening
of MAbsagainst
HIV-1gpl20.
Affinity-purified
HIV-1 IIIBgpl20
was used to immunize maleBALB/c mice. Our standardprocedures
forpurification
of
gpl20
from infected H9 celllysates,
immunization ofmice,
andgeneration
ofhybridomas
werepreviously
de-scribed in detail(10, 47).
Briefly,
3days
after the finalimmunization,
the mice weresacrificed,
andspleen
cells were isolated and fused withSp2/0 myeloma
cells. Afterselection using hypoxanthine-aminopterin-thymidine (HAT) medium, culture supernatantsfrom the wells of microdilu-tionplates with hybridomas were tested for reactivity with gpl20as coating antigen by enzyme-linked immunosorbent assays (ELISAs). Culture supernatants showing strong gpl20 reactivity were tested for staining live HIV-1 IIIB-infected cells by an indirect immunofluorescence method (47). Hybrids fromselected fusion wells were expanded and cloned. MAbsfromhybridomas selected for further charac-terizationwere produced in mouse ascitic fluid and purified by proteinA affinitychromatography (47).
gp120 ELISA for screening of hybridomas. The procedures were similar to those described earlier (10, 47). Wells of Immulon 2 microdilution plates (Dynatech, Chantilly, Va.) were coated overnight at room temperature with 100 ,ul of purified gpl20 (0.1
p.g/ml)
in phosphate-buffered saline (PBS). The wells were then treated with 5% BLOTTO in PBS for 1 h at room temperature and washed with PBS containing 0.05% Tween 20 (PBST). Next, 100 ,ul oftest culturesupernatant wasaddedtoeach well for1 h at room temperature.The wellswerethenwashed andincubated for 1h at room temperaturewithhorseradish peroxidase-conju-gatedgoatanti-mouse IgG (Jackson ImmunoResearch Lab-oratories, Inc., West Grove, Pa.) diluted in PBST containing 1% bovine serumalbumin(PBSTB). After anotherwashing step, boundantibodiesweredetected by reaction with0.1% tetramethyl benzidine and 0.0003% hydrogen peroxide as substrates. The optical density at 450 nm (OD450) of the reaction solutionwas read with anELISA reader.Neutralization assays of HIV-1 laboratory isolates using CEM-SS cells. The
syncytium-forming
microassays using CEM-SScells wereperformed
aspreviously
described (10, 39).Briefly,
50 ,ul of diluted MAb was mixed with50 ,ulof viral culture supernatantcontaining200 syncytium-forming units(SFU) ofHIV-1 IIIB, MN,orRFand incubated for1 h at room temperature. The mixtures were added into microculture wellscontaining
5 x 104 DEAE-dextran-treated CEM-SScells,
andthecellculturesweremaintained in 5%CO2
at37°C for 3 to 4days. Thesyncytia
were then enumerated underaninvertedmicroscope.
Theneutralizing
activity
wasexpressed
asID50,
definedasthe concentrationrequired
to achieve 50% inhibition of the infection(i.e.,
Vn/Vo =
50%),
where Vn is the numberof SFU inthetest wells andVo isthe number of SFU in the control without testantibodies. In thestudy
oftheepitope
specificity using
peptide
V1SP, theneutralizing
activity
ofBAT085 at 12.5,ug/ml
(ID50) was tested for inhibitionby
incubating
theantibody
with various concentrations of the peptidefor 20 min at room temperature,prior
to the addition of a viral inoculumof100SFUs ofHIV-1IIIB.Theirrelevantpeptide
a-endorphin (Peninsula
Laboratories, Belmont, Calif.)
was usedforacontrol.Neutralizationassays ofprimaryHIV-1isolates
using
PHA-activated PBMCs. The titration ofprimary
HIV-1 isolates and their neutralization assays wereperformed by
ourstandard
procedures (5, 17).
An inoculum of 50 50% tissue culture infective doses wasincubated with variousconcen-trationsof MAbs
(0.01
to25,ug)
in 1 ml of final culture for 30 minat37°C.
ThecontrolcontainednoMAb. Onday
4to7,
depending
onthereplication
kinetics of theprimary
HIV-1 isolates,the HIV-1p24
antigen
levels of theantibody-treated
and untreated cell cultures were measured with Abbott HIV-1
Antigen
Kits(Abbott
Laboratories,
NorthChicago,
Ill.).The
neutralizing
activity
oftheantibody
isexpressed
as thepercent reduction ofp24
antigen
levels in theantibody-treated wells
compared
with that of the control.on November 10, 2019 by guest
http://jvi.asm.org/
Flow
cytometry.
Thebinding
ofBAT085
and G3-136toH9 cellsinfected withHIV-1IIIB, MN,
orRFwasanalyzed
by
flow
cytometric
methodsusing
aCoulter EPIC cellanalyzer
(Coulter
Electronics, Hialeah,
Fla.)
aspreviously
described(25).
The bound MAbs were detectedby
fluorescein-conju-gated
goat
anti-mouseIgG (Jackson
ImmunoResearchLab-oratories, Inc.).
Generation of HIV-1
gpl20
peptides.
Allpeptides
used inepitope mapping
weresynthesized by
using
the RaMPspeptide
synthesis
system
(Du
PontCo.,
Wilmington,
Del.)
by
the9-fluorenylmethoxycarbonyl synthesis
protocol
de-scribed in the manual(6a).
Thepurity
of theRaMPS-synthesized
peptides
was characterizedby
high-perfor-manceliquid
chromatography,
and thepredicted
structurewasassessed
by
fast-atom bombardmentmassspectrometry
analysis.
Amino acid sequencesof thesynthetic
peptides
forIIIB
(HXB2
clone),
MN,
and RF isolates ofHIV-1 wereobtainedfrom
Myers
etal.(38).
Epitope mapping
using synthetic
HIV-1gpl20 peptides.
Preliminary epitope mapping
wasperformed by using
nitro-cellulose
strips impregnated
withsequence-overlapping
syn-theticoligopeptides
(9
to15amino acidresidues)
encompass-ing
the entirelength
ofHIV-1IIIB
gpl20
polypeptide
chain(24)
(from
S.Petteway
formerly
ofDuPontCo.).
Theamino acid sequenceswerederived from HIV-1 clone HXB2(38).
The reactivities of anti-HIV-1
gpl20
MAbs with these pep-tidesweredeterminedby
the methodspreviously
described(32).
Briefly,
the nitrocellulosestrips
wereincubated
over-night
atroomtemperature
withtestMAbsat10,ug/ml
in5% BLOTTO. Thestrips
were then washed with PBST. Thebound
antibody
was thendetectedby
incubating
thestrips
with diluted horseradish
peroxidase-conjugated
goat
anti-mouse
IgG
(Jackson
ImmunoResearch Laboratories,Inc.)
for1hatroom
temperature.
Thestrips
werewashed,
andthecolor reaction was
developed
with a substrate solutioncontaining
0.3%4-chloro-1-naphthol
and 0.005%hydrogen
peroxide.
Further fine
epitope mapping
wasperformed by peptide
ELISA. Wells of Immulon 2 microtest
plates
were coatedovernight
at roomtemperature
with 100,ul
of thesynthetic
peptides
(1
,ug/ml)
in PBS. The wells werethentreatedwith5%
BLOTTO
for 1 h atroomtemperature.
After the wells were washed withPBST,
100pAl
ofBAT085
or G3-136(1
jig/ml)
was addedto the wells for reaction for 1 hat roomtemperature.
The wells were thenwashed,
and the boundMAb was then detected
by
horseradishperoxidase-conju-gated
goat
anti-mouseIgG.
The colorreaction wasdevel-oped
by
incubation
with theperoxidase
substrate solutionasdescribed above.
Reactivity
wasregarded
aspositive
if theOD450
wastwofoldhigher
thanthat ofthe wellscoatedwith theirrelevant
peptide
a-endorphin.
Competition
ELISA. The effects ofanti-HIV-1
gpl20
MAbs,
peptide
V15P
and recombinant solubleCD4(sCD4)
on the
binding
ofBAT085
and G3-136to HIV-1 IIIBgpl20
were determined
by
theprocedures
previously
described(35-37).
sCD4wasobtained fromBiogen,
Inc.,
Cambridge,
Mass. Wells of Immulon 2 microtest
plates
were coatedovernight
at roomtemperature
with 50 ,uof a solutioncontaining
5,ug
ofaffinity-purified sheep
anti-HIV-1gpl20
(D6205,
International
Enzymes,
Inc.)
per ml.The wellswerethen treatedwith5%
BLOTTO
for1hatroomtemperature.Fifty
microliters
ofpurified gpl20
(0.5
pug/ml)
wasthenadded toeach wellforincubation
for2hatroomtemperature.
Theplates
werethen washed.Onehundredmicroliters
ofdilutedbiotinylated
MAb was added to the wells with or withoutvarious amounts of
competing
MAbs(including
BAT085,
G3-136,
G3-4, BAT123, G3-519, and ISe), peptide V15P or sCD4. The biotinylated MAbswereused at dilutions which gave half-maximalbindingto thecapturedgpl20.
The incu-bationtimewas 1 h at room temperature. Theplates
were washed and reactedwith 100 Ll of horseradishperoxidase-conjugatedstreptavidin(diluted1:2,000 inPBSTB) (Jackson ImmunoResearch Laboratories, Inc.) for 1 h at room tem-perature.Theplateswerewashed
again
and allowed to react with the peroxidase substrate solution as described above. The degree of inhibition wasexpressed
as thepercent
decrease in
OD450
in the test wellscompared
with the OD of control wells without the inhibitors.RIPA. The radioimmunoprecipitation (RIPA)
procedure
wascarried out as described earlier(34, 47).
Briefly,
HIV-1IIIB-infected
H9 cellsweremetabolicallylabeled for 4 h with[35S]cysteine
and[35S]methionine(100,uCi/ml;
ICNPharma-ceuticals, Inc., Irvine,Calif.)and
suspended
in RIPAlysing
buffer (50 mM Trishydrochloride
[pH
8.0], 150 mMNaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate, 1 mM phenylmethylsulfonylfluoride).
To examine the effect ofantibodybinding
togpl60
andgpl20
after reduction of disulfide linkages, the metabolically la-beled cell lysates was treated with 0.1 M dithiothreitol in PBS at 37°C for 1 h. Dithiothreitol was then removed
by
dialysis.The effect ofdeglycosylationof
gpl60
andgpl20
onantibody binding was also examined by treating the cell lysates with 100 mU of
endo-o-N-acetylglucosaminidase
H for 2.5 h at 37°C. The treated or untreatedlysates
werepreclearedwithprotein A-Sepharosebound to rabbit antise-rum to mouse kappa light chain
(K-PAS)
for 3 h at roomtemperature.RIPA wasperformedby adding3
,ug
ofpurified
MAb (BAT085,
G3-136,
or G3-4) and 0.2 ml of a 10% suspension of K-PAS to 200 ,ulof labeled and clarifiedlysate.
The sampleswere incubatedfor 18 h at4°C,and the beads were washed with RIPA lysing buffer. The pellets were
suspended inelectrophoresis samplebuffer and boiled for 3
min.
Proteins were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis,followedbyautoradiog-raphy.
RESULTS
Generation and characterization of MAbs BAT085 and G3-136. Affinity-purified gpl20 prepared from lysates of HIV-1 IIIB-infected H9 cells was used to immunize mice. The culturesupernatantsofhybridsweretestedfor reactiv-ity with
gpl20
by ELISA. Antibodies exhibiting stronggpl20
reactivity
were tested forstaining live HIV-1 IIIB-infected H9 cellsbyanindirect immunofluorescence method inorder to determine whetherthe antibodies reacted withgpl20oninfected cells. The MAbs from cloned hybrids were tested for neutralizing activity against HIV-1 IIIB using a syncy-tium-forming assay. Two neutralizing MAbs
designated
BAT085 and G3-136 [both IgGl(K)] were identified for fur-thercharacterization.
In the flow
cytometric
assayshown in Fig. 1A,BAT085at 10p.g/ml
stained 87 and 35% of H9cellsinfected withHIV-1 IIIB andMN,
respectively.
It didnotstainH9cellsinfected
with HIV-1 RF. G3-136 stained 73 and 91% of H9 cells infected with HIV-1 IIIB andRF, respectively (Fig. 1B)but did not stain H9 cells infectedwithHIV-1 MN.
In the in vitro neutralizationassaysagainstHIV-1
labora-tory
isolatesusing
CEM-SS cells, BAT085neutralized IIIB withID50
of 12.5p,g/ml
(Fig.
2A). Itdid not neutralize MN and RF. G3-136 neutralized IIIB andRF with ID50 of 16 and2
pg/ml, respectively
(Fig. 3A).
It did not neutralize MN.on November 10, 2019 by guest
http://jvi.asm.org/
1.00
IA:
0*ir 1W 1W 1W
ILO
SICZI7'Nn
FIG. 1. Bindingactivities of BAT085(A)andof G3-136(B)toH9 cells infected with HIV-1 IIIB ( *), MN (....), or RF ( .Uninfected H9 cells( )were used foracontrol. The H9 cells(107cellsperml)were incubated withorwithout the antibodiesfor 30 minonice. After the cellswerewashed,theywere
incubated with goat anti-mouse IgGcoupled withfluorescein iso-thiocyanatefor 30min, washed,andfixed withparaformaldehyde. Fluorescenceintensitywasanalyzedbyflowcytometry.
Neutralizing
activitiesagainst
theinfectivity
ofseveralpri-mary HIV-1 isolates were also tested
by using
PHA-acti-vated PBMCs. BAT085neutralized two(LS
andRP)
of six isolates tested with anID50
of lessthan 1,ug/ml (Fig.
2B);
G3-136neutralizedthree
(AC, LS,
andTB)
ofsevenisolateswith
ID50 ranging
from0.03 to0.4,ug/ml
(Fig. 3B).
Epitope mapping
ofBAT085andG3-136. Todelineate theepitopes
ongpl20
that these twoMAbsrecognize,
we firsttested theeffect of other
previously
characterizedneutraliz-ing
anti-HIV-1gpl20
MAbs on thebinding
ofbiotinylated
BAT085 and G3-136 to
gp120.
These other MAbs includeBAT123,
G3-519, G3-4,
andISe;
the lasttwo MAbs recog-nizedistinct conformationalneutralizing
epitopes
ongp120.
In the
competition
ELISA,
purified
HIV-1 IIIBgpl20
wascaptured
by
affinity-purified
sheep
anti-HIV-1gpl20
anti-bodies which arespecific
for ahighly
conservedpeptidic
segmentin the
carboxy
terminusofgp120.
The immobiliza-tion ofgpl20
didnothinderthebinding
ofBAT123, G3-519,
G3-4, I5e, BAT085,
and G3-136.Figure
4A shows that thebinding
ofbiotinylated
BAT085togpl20
waspartially
inhib-itedby
G3-4(80%)
and G3-136(65%)
at 40,g/ml.
In contrast,BAT123, G3-519,
and ISe hadnoeffect.Unconju-gated
BAT085 at 40,ug/ml
completely
abolishedbinding.
When the
binding
ofbiotinylated
G3-136 togpl20
wastested,
G3-4inhibited thebinding
aseffectively
asG3-136did(Fig. 4B). However,
BAT085 inhibitedthebinding by only
33%evenat40
,ug/ml,
andBAT123,
G3-519,
andISe hadnoeffect. These results were confirmed
by
examining
theef-0
0.10
0.01
c 0
0
0
()
04.
100
80
60
40
20
0
10 BAT085, sg/mI
100
0 0.01 0.10 1.00 10.00 100.00
BAT085,sg/ml
FIG. 2. Neutralization ofHIV-1 by BAT085. (A) Inhibition of infection ofCEM-SScellsby laboratory HIV-1 isolateIIIB (0), MN (0),and RF(A).Aviral inoculumof100SFUwasusedtoinfect the cells in the presence or absence of various concentrations of
BAT085. Vnisthemeannumber of syncytia in duplicatetestwells, and Vo is the mean number of syncytia in the duplicate control wells. (B) Inhibition of infection ofPHA-activated PBMCs from HIV-1-seronegative donorsby primaryHIV-1 isolates LS(0), RP (V), CC(O), JR-CSF(O),LL(A),andTB(A). Aviralinoculum of50 50% tissuecultureinfective doseswasusedtoinfect the cells in the
presence or absence of various concentrations of BAT085. The neutralizing activity of the antibody is expressed as the percent
reductionofp24 antigenlevels in the antibody-treated wells
com-paredwiththatof the control.
fects of BAT085 andG3-136 on the binding of biotinylated
G3-4, BAT123, G3-519, and ISe to gpl20. As expected, BAT085 and G3-136 competed with G3-4 but not with BAT123, G3-519, and ISe (16; also unpublished results).
To define thebinding epitope(s) of BAT085 and G3-136on
gp120, the MAbs were examined by using nitrocellulose
stripsimpregnatedwithsequence-overlapping synthetic
pep-tides encompassing the entire length of HIV-1 IIIB gpl20 polypeptide chain. It was found that BAT085 and G3-136
reactedspecificallywithapeptidicsegment, VQKEYAFFY
KLDIIP (referred to as V15P) in the V2 region ofgp120, amino acid residues 169to183. To confirm thisobservation, overlapping oligopeptides encompassing the peptidic
seg-ment (amino acid residues 159 to 198) inHIV-1 IIIB gp120 (Table 1) were synthesized for ELISAs and liquid-phase
competition assays. When these peptides were usedas the solid-phase antigens at 1 ,ug/ml, BAT085 reacted strongly withV15P(OD450 = 1.4), andweakly with L20T(OD450 =
0.13). It did not reactwith F1SY, V1OK, and AlOP, giving OD450s less than twofold greater than the negative control a-endorphin(OD450= 0.01). G3-136also reacted withV15P
A
.8
--..P--- - -0
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.93.277.66.342.2] [image:4.612.322.562.69.366.2]1.0(
0.14
0.0
100 80
60
40
20
0
) 0
1 ... ~.
1 10 100
G3-136, yg/ml
B
*
a
0 0.01 0.10 1.00 10.00 100.00
G3-136. /g/ml
FIG. 3. Neutralization of HIV-1 by G3-136. (A) Inhibition of
infection of CEM-SS cells by laboratory HIV-1 isolates IIIB (0), MN(0),and RF(A). (B)Inhibition of infection of PHA-activated PBMCs fromHIV-1-seronegativedonorsby primaryHIV-1 isolates AC(0),LS(0),LL(A),TB(A),CC(O),JR-CSF(O),and RF(V). The neutralizing activities of the antibody were determined as
described in thelegendtoFig.2.
(OD450 = 0.44) but did not reactwith F15Y, L20T, V1OK,
and A1OP. In the determination ofreactivity with different HIV-1isolates,BAT085 didnot reactwithpeptides M15S and T15P, which correspond tothesame region of peptide V15P
100
c
.0
-Cc
-0.
0
0
L-80 60 40 20
0 0.1
100
.0
*t_
.0
a-80
60
40
20
0
1.0 10.0
Competing antibodies, ug/ml
0 0.1 1.0 10.0
Competing antibodies, p,g/ml
FIG. 4. Inhibition of thebindingofbiotinylatedBAT085(A)and biotinylatedG3-136(B)tocapturedHIV-1 IIIBgpl20in ELISAby BAT085 (A),G3-136(0),G3-4(0), BAT123(A), G3-519(l),and
15e(0).ThecompetitionELISAprocedureis described inMaterials and Methods.
in the gpl20 of HIV-1 MN and RF, respectively. G3-136 reacted with T15P(OD450 = 0.23), butnotwithM15S.
Inthe liquid-phase competition bindingassay examining
theeffects ofpeptidesonthebinding of biotinylated BAT085
[image:5.612.319.559.63.375.2]tocapturedgpl20, V15P inhibited the binding of BAT085to
TABLE 1. Epitopemapping of BAT085 and G3-136 to theV2region of HIV-1gpl20by peptide ELISA
Viralpeptide Sequence (amino acids
159-198")
Reactivity(OD450)andantigen BAT085 G3-136
HIV HXB2
160 170 180 190
F15Y FNISTSIRGKVQKEY 0.01 0.03
V15P VQKEYAFFYKLDIIP 1.40 0.44
L20T LDIIPIDNDTTSYSLTSCNT 0.13 0.02
V1OK VQKEYAFFYK 0.02 0.01
AlOP AFFYKLDIIP 0.03 0.01
HIV MN
M15S M----LL----VS 0.01 0.02
HIV RF
T15P T--K--L----VV- 0.01 0.23
aDeduced amino acid sequences are obtained from Myers et al. (38). Homologous amino acids are represented by dashes. TheOD450of the blank wasbetween 0.01 and 0.03.
0
c
c 0
c 0
0
C)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.62.305.68.373.2] [image:5.612.59.556.542.702.2]100
c
0
.0 .r_
-c
._-c
0) 0-0)
80 60
40
20
0.01 0.10 1.00 10.00
Competing peptides, tg/ml
FIG. 5. Competitionof thebinding between biotinylated BAT085 andcapturedHIV-1IIIB gpl20 by the epitopepeptide V15P (0) and
theirrelevant peptidea-endorphin(0).Theamino acidsequenceof
V15P is shown in Table 1. The ELISA procedure is described in
Materials and Methods.
gp120inadose-dependentmanner(Fig.5).Thea-endorphin
peptide had no effect, evenataconcentrationof 40 p.g/ml.
Thebinding specificity ofV15PtoBAT085 was also
exam-inedintheneutralizationassayofHIV-1 IIIBusing CEM-SS cells.Figure 6 shows that V15P exhibitedadose-dependent
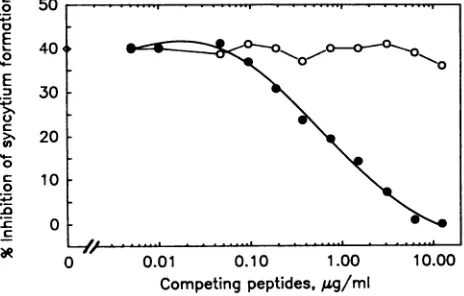
inhibition of the neutralizing activity of BAT085, while a-endorphin hadnoeffect. Either peptide alone didnotaffect theinfectivity of HIV-1 IIIB.
Effectsof sCD4 on thebinding ofBAT085 and G3-136 to
gpl2O. Toexaminewhether the neutralizing epitope mapped to the V2 region was involved in the interaction between
CD4 and gpl20, the effect of sCD4 on the binding of the
biotinylated MAbs to captured gp120 was tested. sCD4
inhibitedthe binding of G3-136, G3-4, and G3-519 togpl20 (Fig. 7), althoughnotcompletelyevenataconcentrationas
high as 40 ,ug/ml. The binding of BAT085 and BAT123 to gpl20 isnotaffected by sCD4. These resultssuggestthatthe
c
0
0
E
(0
-._-c
co
0
c
'
50
40
30
20
10
0
0 0.01 0.10 1.00 10.00
Competing peptides,ug/ml
FIG. 6. Effects ofoligopeptides on the neutralizing activity of
BAT085 against HIV-1 IIIB. Asdescribed in Materials and
Meth-ods, CEM-SS cells(5 x 104)were infectedwith 100 SFU of HIV-1
IIIBin thepresenceofBAT085at12.5,ug/ml equivalenttoID50 (°>)-Various concentrations of the epitope peptide V15P (0) or the
irrelevantpeptideoa-endorphin (0)weremixed with theantibodyas
described in Materials and Methods. The percent inhibition of
syncytium formation is defined as the percent decrease in the
number ofsyncytia in the test wells compared with that of the
control wellswithout theantibody.
70 60
c
50-0
z
40-.c 30-c
20
00
r~10
0
0.01 0.10 1.00 10.00
sCD4, /g/mI
FIG. 7. Inhibitoryactivity of sCD4onthebindingof biotinylated
MAbsto
ca,ptured
HIV-1 IIIBgpl20 in ELISA. Thesebiotinylated antibodies, including BAT085 (D), G3-136 (A), G3-4 (A), G3-519 (O), and BAT123 (0) were used at dilutions which gavehalf-maximal bindingtothe capturedgpl20.Various concentrations of
sCD4 were mixed with the labeled antibodies for binding to the gpl20 for 1 hat room temperature asdescribed in Materials and
Methods.
epitopes recognized by BAT085 and G3-136 may be
dif-ferent, and that ofG3-136 isnear aCD4-binding domain of
gpl20.
Effect of reduction and deglycosylation of gpl20 on its
binding by BAT085 and G3-136. To study whether the binding of BAT085 and G3-136 to gp120 was affected by
conformational changes ofgpl20, the effects of reduction of disulfide linkages anddeglycosylation of gpl20were
exam-ined. As summarized in Table 2, the reactivity of G3-136 withgpl20 inaRIPAwasreduced when the virallysatewas
firstdenaturedbydithiothreitol.Whenaradiolabeled HIV-1
lysate was subjectedto
endo-p-N-acetylglucosaminidase
H digestion, the partially deglycosylated product of 90to 100 kDawasweaklyrecognizedbyG3-136 (Table 2). Inparallelexperiments, the reactivity of BAT085 with reduced or
deglycosylated gp120wasunaltered, whereas the reactivity
ofG3-4 with gp120 was completely abolished under these
conditions.
DISCUSSION
We reporthere thataunique neutralization domain inthe
V2region of HIV-1gp120 has been identified bytwomurine MAbs, BAT085 and G3-136, raised against HIV-1 IIIB gpl20. ThesetwoMAbs canneutralizeinvitro the infectiv-ity of certain laboratory and primary HIV-1 isolates. The immunologic and biologic properties of these MAbs are
summarizedinTable 2.
By usingsequence-overlapping synthetic peptides
encom-passing the entire length of the HIV-1 gpl20 polypeptide chain,thebindingsiteof BAT085 andG3-136wasmappedto thepeptidic segmentV15P(aminoacid residues 169 to183) in HIV-1 IIIB gp120 (Table 1). Confirming our findings,
Stephens et al. (46) recently reported that antisera to the
synthetic peptide definingthe corresponding segmentinthe
V2 region of HIV-1 SF2 gpl20(amino acidresidues 171 to 185) neutralized in vitro the infectivity of the virus. The
specificity ofpeptide V15Pfor BAT085hasbeenconfirmed bythe liquid-phase competitionassays asillustrated in Fig. 5 and6. The absence ofreactivity of these MAbs with the
shorterpeptides
(VlOK
and AlOP) ofthis regionindicates-1'~~~~~~~~~~~,~
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.59.300.68.215.2] [image:6.612.317.553.71.220.2] [image:6.612.64.296.482.630.2]TABLE 2. Properties of neutralizing MAbstothe V2 region of HIV-1gpl20a
Neutralization Staining cells Effectofdegly- Effect of disulfide Bindingtoepitope
MAb of HIV infectedwith HIV sCD4
inhibition
of cosylation on bond reduction on peptide(amino IIIB MN RF IIIB MN RFgpl20
bindinggpl20
bindinggpl20
binding acids 169-183)BAT085[IgGl(K)] + - - ++ + - - None None ++
G3-136 [IgGl(K)] + - ++ + - ++ + Reduced Reduced +
G3-4[IgG2a(K)] + - ++ + - ++ + Abolished Abolished
-aSymbols: +,reactive; -,unreactive; ++,stronglyreactive.
that the actual binding site may be composed of several amino acid residues interspersedwithin peptide V15P. The identities of the crucial amino acid residues remain to be elucidated. Despite the fact that both BAT085 and G3-136
can bind to peptide V15P, only G3-136 reacts with peptide T15P of HIV-1 RF andstrongly stainsH9cellsinfectedwith this viralisolate. It is still unclear which aminoacid residues inthesepeptides actually determine thebinding specificities of different HIV-1 isolates.
Inasmuch as the binding of G3-136 to gpl20 is partially sensitive to deglycosylation and reduction (Table 2), its binding epitope is likely to be conformation dependent. In contrast, thebinding of BAT085togpl20isnotsensitiveto conformational changes of gpl20. This subtle difference in the binding characteristics between G3-136 and BAT085 is consistent with the inability of BAT085 to compete effec-tively with the binding of G3-136 to gpl20 (Fig. 4B). We recently reported that a murine HIV-1-neutralizing MAb,
G3-4, recognized a conformational epitope in HIV-1 IIIB
[image:7.612.68.563.83.154.2]gpl20 (Table 2) (15). Although G3-4 did not react with peptide V15P, it effectively inhibited the binding of G3-136 but not BAT085 to gpl20, suggesting that the binding epitopes ofG3-4 and G3-136 areoverlapping (Table 2). The
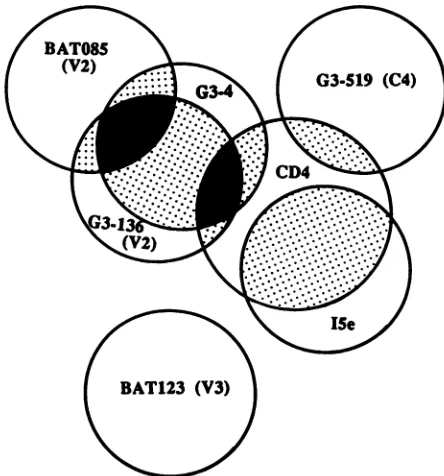
FIG. 8. Schematic representation of the epitopes for BAT085, G3-136, G3-4, G3-519, BAT123, andISe ongpl20and their spatial
relationship with one another and with the CD4-binding site as
determined by competitionELISA.
binding of G3-4 togpl20isstrictly conformation dependent, because its binding is completely abolished when gpl20is deglycosylated or reduced (15). On the basis of these find-ings, we conclude that the V2 region ofgpl20may contain multiple neutralization determinants constituted by either continuous or discontinuous structures.
Results from this study also indicate that the neutraliza-tion determinants in the V2 region recognized by BAT085 and G3-136 are likely to be topographically removed from the PND in the V3region and theCD4-bindingdomain inthe C4region, since thebinding ofBAT085 orG3-136 to gpl20 cannot be inhibited by BAT123 and G3-519 in ELISA. In addition, BAT085 and G3-136 do not inhibit the gpl20 binding of ISe, which recognizes a distinct conformational epitope yet to be defined (16). This observation provides credence to the possibility ofcombining HIV-1 MAbs and variousneutralizationepitopes in passiveimmunotherapyto conferimproved efficacy and broadened viralreactivity. In vitro experiments are in progress in our laboratories to address the functional interactions between these neutrali-zation sites.
Itisinteresting to notethatG3-136 neutralizes a heterol-ogous HIV-1 isolate (RF)moreefficientlythan the homolo-gous isolate(IIIB) (Fig. 3A),althoughits immunoreacitivity is stronger with peptide V15P of IIIB than with the corre-sponding peptideT15PofRF. Apossibleexplanation forthis observation is that the binding site on native HIV-1 IIIB gpl20 is not asaccessible as that ofHIV-1 RF. The acces-sibility oftheneutralization sitesongpl20canbeaffected by theintricate folding of gpl20,as well as the variable extentof glycosylation of gpl20 (8, 14, 30). The juxtaposition of discontinuous regions of gpl20 has been implicated in the association between gpl20and gp4l (26)and in the escape of HIV-1 variantsfromneutralizationbyantibodies(41,51). In linewiththis,recentstudiesby Davis et al. (6) indicated that glycosylation of gpl20 could affect the binding of anti-peptide antibodies to gpl20. Another possible explanation for the higher neutralizing activityof G3-136 against HIV-1 RFis thatsincethe binding ofG3-136 togpl20 ispartially conformation dependent (Table2),G3-136may fit the three-dimensionalconfigurationofthe binding site on thegpl20of the RFisolate better.
In order tounderstand the possible mechanisms of neu-tralization mediated by BAT085 and G3-136, we examined whethertheir binding to
gpl20
can affectCD4-gpl20
inter-action. Indeed, likeG3-519, thebinding of G3-136 andG3-4 togpl20 can be inhibited by sCD4. In contrast, the binding ofBAT085togpl20is not affected in the presence of sCD4. Thesedatasuggests that the neutralizing activities ofG3-136 andG3-4maybemediated by interfering with the interaction between CD4 and gpl20, whereas BAT085 may have a distinct and as yet undefined mechanism. The inability of sCD4tocompletely abolish the binding ofG3-136,G3-4,
or G3-519 to gpl20 can be explained by the fact that theon November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.70.292.439.677.2]structural determinants recognized by these antibodies are either only sterically proximal to the actual CD4 binding sites or are forming parts of the multiple discontinuous regions responsible for CD4 binding (1, 42). The spatial relationship of the newly identified V2 neutralization epitopes relative to the otherpreviously defined neutraliza-tion sites (V3, C4, and the I5e conformational determinant) and totheCD4-binding site isschematically showninFig.8. Onthe basisof thefindingsinthisstudy,weconcludethat the V2 region of gpl20 bears multiple and
overlapping
neutralization determinants. To better design active and passive immunization
strategies,
it isimportant
tostudy
whetherneutralizing antibodies tothisregion play anactive role in the protective immunity against HIV-1 infection in humans.
ACKNOWLEDGMENTS
We thankN. Chang foradvice, J.Robinson forantibody15e, S. Petteway forgp120 peptide-coatednitrocellulosestrips,N. Jonesfor technical assistance in peptide synthesis, and J. Wrightfor prepa-ration of the manuscript.
This study was supported in part by grants from the National Institutes of Health(AI25541,A128747, A127742,andA124030),the ErnstJungFoundation, and theAaron Diamond Foundation.
REFERENCES
1. Ardman, B., M. Kowalski, J. Bristol, W. Haseltine, and J. Sodroski. 1990. Effects on CD4 bindingofanti-peptideserato
the fourth and fifth conserved domains of HIV-1 gpl20. J. AcquiredImmune Defic. Syndr. 3:206-214.
2. Berkower, I., G. E. Smith, G. Chad, and D. Murphy. 1989. Human immunodeficiency virus 1: predominance ofa group-specific neutralizing epitope thatpersists despitegenetic varia-tion. J. Exp. Med. 170:1681-1695.
3. Berman, P. W.,J. E. Groopman,T. Gregory,P. R. Clapham, R.A. Weiss, R. Ferriani, L. Riddle, C. Shimasaki, C. Lucas, L. A. Lasky, and J. W. Eichberg. 1988. Human immunodefi-ciency virus type 1 challenge ofchimpanzeesimmunized with recombinant envelope glycoprotein gpl20. Proc. Natl. Acad. Sci. USA85:5200-5204.
4. Bolognesi, D. P. 1989. Prospects for prevention of and
early
interventionagainstHIV. JAMA 261:3007-3013.
5. Daar, E. S., X. L. Li, T. Moudgil, and D. D. Ho. 1990.
High
concentrations of recombinant soluble CD4 are
required
toneutralize primary human immunodeficiency virus type 1 iso-lates. Proc. Natl. Acad. Sci. USA 87:6574-6578.
6. Davis, D., D. M. Stephens, C. Willers, and P. J. Lachmann. 1990.Glycosylation governs thebindingof
antipeptide
antibod-ies to regions of hypervariable amino acid sequence within recombinantgpl20of humanimmunodeficiency
virustype1.J. Gen. Virol. 71:2889-2898.6a.Du Pont Company. 1987. Du Pontcatalog. Du Pont
Company,
Wilmington,Del.
7. Emini, E.A., W. A.Schleif, K. Murthy,Y. Eda,S.
Tokiyoshi,
S.D.Putney,S.Matsushita,J.H.Nunberg,andJ.W.Eichberg.
1991. Passive immunization with a monoclonal antibody di-rectedtothe HIV-1 gpl20principalneutralization determinant confersprotection against HIV-1
challenge
inchimpanzees, p.72. Th.A.64. Abstr. VII Int. Conf. AIDS, vol. 2. Istituto Superiore Di Sanita,Rome.
8. Fenouillet, E.,J.C. Gluckman,and E. Bahraoui. 1990. Roleof N-linked glycans of envelope
glycoproteins
ininfectivity
of human immunodeficiencyvirustype 1.J.Virol.64:2841-2848. 9. Fung,M. S. C., C. Sun, N. C. Sun, N. T.Chang,
and T. W.Chang. 1987. Monoclonal antibodies that neutralize HIV-1 virions and inhibit syncytiumformation
by
infected cells. Bio/ Technology 54:940-946.10. Fung,M. S.C., C. R. Y. Sun, R. S. Liou, W. Gordon, N.T. Chang, T. W. Chang, and N. C. Sun. 1990. Monoclonal anti-idiotypicantibody
mimicking
theprincipal
neutralization site inHIV-1
gp120
induces HIV-1neutralizing
antibodies in rabbits.J. Immunol. 145:2199-2206.11. Goudsmit, J., C.Debouck,R.H. Meloen,L.
Smit,
M. Bakker,D. M.Asher,A. V.Wolff,C.J.Gibbs, Jr.,and D. C.Gajdusek.
1988. Human
immunodeficiency
virus type 1 neutralizationepitope
with conserved architecture elicitsearly
type-specific
antibodies in
experimentally
infectedchimpanzees.
Proc.Natl. Acad. Sci. USA85:4478-4482.
12. Goudsmit, J.,C.Thiriant,L. Smit,C.Bruck,andC.J.Gibbs. 1988.
Temporal development
of cross-neutralization between HTLV-IIIB andHTLV-IIIRFinexperimentally
infected chim-panzees.Vaccine 6:229-232.13.
Haigwood,
N. L., C. B. Barker, K. W.Higgins,
P. V. Skiles,G. K.Moore,K. A.Mann,D. R.Lee,J.W.
Eichberg,
and K.S. Steimer. 1990. Evidence forneutralizing
antibodies directedagainst
conformationalepitopes
of HIV-1gp120,
p. 313-320.In R. A.Lerner(ed.),Vaccine 90. ColdSpring
HarborLaboratory,
Cold
Spring Spring
Harbor, N.Y.14. Hansen,J.-E.S.,H.Clausen,C.Nielsen,L.S.
Teglbjaerg,
L. L.Hansen, C. M. Nielsen, E. Dabelsteen, L. Mathiesen, S.-I. Hakomori,andJ.0. Nielsen. 1990.Inhibition of human
immu-nodeficiency
virus(HIV)infection in vitroby anticarbohydrate
monoclonal antibodies:
peripheral
glycosylation
of HIVenve-lope glycoprotein
gp120
maybeatargetfor virusneutralization.J. Virol. 64:2833-2840.
15. Ho, D. D., M. S. C.
Fung,
Y. Cao, X. L. Li, C. Sun, T. W.Chang,
and N.C. Sun. 1991. Anotherdiscontinuousepitope
ongpl20
thatisimportant
in humanimmunodeficiency
virustype1 neutralization is identifiedby
a monoclonalantibody.
Proc.Natl. Acad. Sci. USA88:8949-8952.
16. Ho,D. D.,J. A.
McKeating,
X. L.Li,T.Moudgil,
E. S.Daar,N. C.Sun,andJ.E.Robinson.1991.Conformational
epitope
ongpl20
important
in CD4binding
andhumanimmunodeficiency
virus type 1 neutralization identified
by
ahuman monoclonalantibody.
J. Virol. 65:489-493.17. Ho, D. D., T.
Moudgil,
and M. Alam. 1989.Quantitation
of humanimmunodeficiency
virustype 1in the blood of infected persons. N.Engl.
J. Med. 321:1621-1625.18. Ho, D. D.,M. G.
Sarngadharan,
M. S.Hirsh, R. T.Schooley,
T.R.Rota,R. C.Kennedy,
T. C.Chanh,and V. L. Sato.1987. Humanimmunodeficiency
virusneutralizing
antibodies recog-nizeseveralconserved domainsontheenvelope
glycoproteins.
J. Virol.61:2024-2028.
19. Jackson, G. G., J. T. Perkins, M. Rubenis, D. A. Paul, M.
Knigge,
J.C.Despotes,
and P.Spencer.
1988.Passive immuno-neutralization of humanimmunodeficiency
virus inpatients
with advancedAIDS. Lancetii:647-651.20. Javaherian, K.,A.J.
Langlois,
G.J.LaRosa,A. T.Profy,
D.P.Bolognesi,
W. C.Herlihy,
S. D.Putney,
and T. J. Matthews. 1990.Broadly
neutralizing
antibodies elicitedby
the hypervari-ableneutralizing
determinant of HIV-1. Science250:1590-1593. 21. Javaherian, K., A. J.Langlois,
C. McDanal, K. L. Ross,L. I.Eckler,C.L.Jellis,A.T.
Profy,
J.R.Rusche,D. P.Bolognesi,
S. D.
Putney,
and T. J. Matthews. 1989.Principal neutralizing
domain of the human
immunodeficiency
virustype 1envelope
protein.
Proc. Natl. Acad. Sci. USA86:6768-6772.22.
Karpas,
A.,I.K.Hewlett,F.Hill,J.Gray,
N.Byron,
D.Gilgen,
V.Bally,
J. K. Oates, B. Gazzard, and J. E.Epstein.
1990.Polymerase
chain reaction evidence for humanimmunodefi-ciency
virus 1 neutralizationby
passive
immunization in pa-tients withAIDS andAIDS-relatedcomplex.
Proc.Natl.
Acad. Sci. USA 87:7613-7617.23.
Karpas,
A.,F.Hill,M.Youle,V.Cullen,J.Gray,
N.Byron,
F.Hayhoe,
M.Tenant-Flowers,L.Howard, D.Gilgen,
J.K.Oates,D. Hawkins, and B.Gazzard. 1988. Effects of
passive
immuni-zation inpatients
with theacquired immunodeficiency
syn-drome-related
complex
andacquired immunodeficiency
syn-drome. Proc. Natl. Acad. Sci. USA85:9234-9237.
24. Kenealy,W.R.,T.J.Matthews,M.C.
Ganfield,
A.J.Langlois,
D.M.
Waselefsky,
and S. R.Petteway,Jr.1989.Antibodiesfrom humanimmunodeficiency
virus-infected individuals bind to ashortamino acidsequencethatelicits
neutralizing
antibodiesin animals. AIDSRes. HumanRetroviruses 5:173-182.on November 10, 2019 by guest
http://jvi.asm.org/
25. Kim, Y. W.,M. S. C. Fung, N. C. Sun,C. R. Y. Sun,N. T. Chang,and T. W.Chang. 1990. Immunoconjugates that neutral-ize HIV virions kill T cells infected with diverse strains of HIV-1. J.Immunol. 144:1257-1262.
26. Kowalski, M., J. Potz,L.Basiripour,T.Dorfman,W.C.Goh,E. Terwilliger, A. Dayton, C. Rosen,W.Haseltine,andJ. Sodroski. 1987.Functionalregions of the envelopeglycoproteinof human immunodeficiency virus type1.Science 237:1351-1355. 27. LaRosa, G. J., J. P. Davide, K. Weinhold, J. A. Waterbury,
A. T.Profy, J.A.Lewis,A.J.Langlois, G.R.Dreesman,R. N. Boswell, P.Shadduck, L.H.Holley,M.Karplus,D.P.Bolognesi, T.J. Matthews,E. A.Emini,andS.D.Putney. 1990.Conserved sequenceandstructural elements in HIV-1 principal neutraliz-ingdeterminant. Science 249:932-935.
28. Lasky, L. A., J. E.Groopman, C.W.Fennie,P. M.Benz,D.J. Capon,D.J. Dowbenko, G.R.Nakamura,W. M.Nunes, M. E. Renz, and P. W. Berman. 1986. Neutralization of the AIDS retrovirus by antibodies to a recombinant envelope protein. Science 233:209-212.
29. Lasky, L. A., G. M. Nakamura, D. H. Smith, C. Fennie, C. Shimasaki,E.Patzer,P.Berman, T. Gregory,and D.J. Capon. 1987. Delineation of aregion of the humanimmunodeficiency virus type Igpl20glycoprotein critical for interaction with the CD4 receptor. Cell 50:975-985.
30. Leonard, C. K.,M. W.Spellman,L.Riddle,R.J.Harris, J.N. Thomas, and T. J. Gregory. 1990. Assignment of intrachain disulfide bonds and characterization ofpotential glycosylation sitesof the type 1 recombinant humanimmunodeficiency virus envelope glycoprotein (gp120) expressed in Chinese hamster ovarycells. J. Biol. Chem. 265:10373-10382.
31. Linsley, P. S., J. A.Ledbetter, E. Kinney-Thomas, and S.-L. Hu. 1988. Effects of anti-gp120 monoclonal antibodies on CD4 receptorbinding by the env protein of humanimmunodeficiency virus type 1. J. Virol. 62:3695-3702.
32. Liou,R.S.,E. M.Rosen,M.S. C. Fung, W.N.C. Sun, C. Sun, W.Gordon, N. T. Chang,andT. W.Chang. 1989. Achimeric mouse-human antibody that retains specificity for HIV gpl20 and mediates the lysis of HIV-infected cells. J. Immunol. 143:3967-3975.
33. Matthews, T. J., A. J. Langlois, W. G. Robey, N. T. Chang, R. C. Galo, P. J. Fischinger, and D. P. Bolognesi. 1986. Re-stricted neutralization of divergent human T-lymphotropic virus III isolates by antibodies to the major envelope glycoprotein. Proc. Natl. Acad. Sci. USA 83:9709-9713.
34. McDougal, J. S., J. K. A.Nicholson, G. D. Croso, S.P. Cort, M. S. Kennedy,and A. C. Mawle. 1986. Binding of the human retrovirusHTLV-III/LAV/ARV/HIVto theCD4 (T4) molecule: conformation dependence, epitope mapping, antibody inhibi-tion, and potential for idiotypic mimicry. J. Immunol. 137:2937-2944.
35. Moore, J. P. 1990. Simple methods formonitoring HIV-1 and HIV-2gpl20binding to sCD4 by ELISA: HIV-2 has a 25-fold loweraffinity than HIV-1 for sCD4. AIDS 3:297-305.
36. Moore, J.P.,J. A.McKeating, I.M. Jones, P. E.Stephens, G. Clements, S. Thomson, and R. A. Weiss. 1990. Characterization of recombinantgpl20andgpl60from HIV-1: binding to mono-clonalantibodies and sCD4. AIDS 4:307-315.
37. Moore, J. P., L. A. Wallace, E. A. C. Follett, and J. A. McKeating. 1989. An enzyme-linked immunosorbent assay for antibodies to the envelope glycoproteins of divergent strains of HIV-1. AIDS3:155-163.
38. Myers, G., S. F. Josephs, A. B. Rabson, T. F. Smith, and F. Wong-Staal (ed.). 1989. Human retroviruses and AIDS. Los AlamosNationalLaboratories, LosAlamos, N.Mex.
39. Nara,P. L.,W.C.Hatch,N. M. Dunlop, W.G.Robey,L.0. Arthur,M. A.Gonda,and P.J.Fischinger. 1987.Simple,rapid, quantitative, syncytium-formingmicroassay for the detection of human immunodeficiency virus neutralizing antibody. AIDS Res. HumanRetroviruses 3:283-302.
40. Nara, P. L., W. G. Robey, S. W. Pyle, W. C. Hatch, N. W. Dunlop, J. W. Bess, J. C. Kelliher, L. 0. Arthur, and P. J. Fischinger. 1988. Purified envelope glycoproteinsfrom human immunodeficiency virus type1variants induceindividual, type-specificneutralizing antibodies. J. Virol. 62:2622-2628. 41. Nara, P. L., L. Smit, N. Dunlop, W. Hatch, M. Merges, D.
Waters, J. Kelliher, R. C. Gallo, P. J. Fischinger, and J. Goudsmit. 1990. Emergence of viruses resistantto neutraliza-tion byV3-specific antibodies inexperimentalhuman immuno-deficiency virus type1 IIIBinfection ofchimpanzees. J.Virol. 64:3779-3791.
42. Olshevsky, U., E.Helseth,C. Furman,J. Li,W.Haseltine,and J. Sodroski. 1990. Identification of individual human immuno-deficiency virus type 1 gpl20amino acidsimportant for CD4 receptorbinding. J. Virol. 64:5701-5707.
43. Palker, T.J.,M.E.Clark,A.J.Langlois,T.J. Matthews,K.J. Weinhold, R. R. Randall, D. P. Bolognesi, and B. F. Haynes. 1988. Type-specific neutralization of the human immunodefi-ciencyvirus withantibodiestoenv-encodedsyntheticpeptides. Proc.Natl. Acad. Sci. USA 85:1932-1936.
44. Skinner,M.A.,A.J.Langlois,C. B.McDanal,J. S. McDougal, D.P.Bolognesi,and T.J.Matthews.1988.Neutralizing antibod-iesto an immunodominant envelope sequence donot prevent gpl20bindingtoCD4. J. Virol. 62:4195-4200.
45. Sonigo, P.,M.Girard,and D.Dormond.1990. Design and trials of AIDS vaccines. Immunol.Today 11:465-471.
46. Stephens, D. M., D. Davis, and P. J. Lachmann. 1991. The second variableregion of HIV-1external envelope glycoprotein containsaneutralizing epitope, p. 72, TH.A.66. Abstr.VII Int. Conf. AIDS,vol. 2. IstitutoSuperiore DiSanita,Rome. 47. Sun, N.-C., D. D. Ho, C.R. Y.Sun, R.-S. Liou, W. Gordon,
M.S. C.Fung, X.-L.Li, R. C. Ting, T.-H. Lee, N. T. Chang, and T.-W. Chang. 1989. Generation and characterization of monoclonal antibodies to the putative CD4-binding domain of humanimmunodeficiency virus type1gpl20.J.Virol. 63:3579-3585.
48. Weiss,R.A., P. R.Clapham, J.N. Weber, A.G.Dalgleish, L. A. Lasky,andP. W.Berman. 1986.Variable and conserved neu-tralization antigens of human immunodeficiency virus. Nature (London) 324:572-575.
49. Wendler, I., V. Bienzle, and G. Hunsman. 1987. Neutralizing antibodies and the course of HIV-induced disease. AIDS Res. HumanRetroviruses 3:157-163.
50. White-Scharf, M.E.,A. Conley, B. J. Potts, E. A. Emini,J.R. Rusche,and S.Silver. 1991.Monoclonal antibodies directed to the HIV-1 gpl20 V3 loop exhibit patterns of broad HIV-1 neutralization in vitro, p. 176, W.A.1341. Abstr. VII Int. Conf. AIDS. vol. 2. Istituto Superiore Di Sanita, Rome.
51. Willey,R.L., E. K. Ross, A. J. Buckler-White, T. S. Theodore, and M. A.Martin.1989. Functionalinteraction of constant and variable domains of human immunodeficiency virus type 1 gpl20.J. Virol. 63:3595-3600.