Investigation of Host Range of and Host Defense against a
Mitochondrially Replicating Mitovirus
Sabitree Shahi,aAna Eusebio-Cope,a*Hideki Kondo,aBradley I. Hillman,b Nobuhiro Suzukia
aInstitute of Plant Science and Resources, Okayama University, Kurashiki, Okayama, Japan bPlant Biology and Pathology, Rutgers University, New Brunswick, New Jersey, USA
ABSTRACT Mitoviruses (genus Mitovirus, family Narnaviridae) are mitochondrially replicating viruses that have the simplest positive-sense RNA genomes of 2.2 to 4.4 kb with a single open reading frame (ORF) encoding an RNA-dependent RNA polymerase. Cryphonectria parasitica mitovirus 1 (CpMV1) from U.S. strain NB631 of
the chestnut blight fungus,Cryphonectria parasitica, was the first virus identified as a
mitochondrially replicating virus. Despite subsequent discovery of many other mito-viruses from diverse fungi, no great advances in understanding mitovirus biology have emerged, partly because of the lack of inoculation methods. Here we devel-oped a protoplast fusion-based protocol for horizontal transmission of CpMV1 that entailed fusion of recipient and donor protoplasts, hyphal anastomosis, and single-conidium isolation. This method allowed expansion of the host range to many other
C. parasitica strains. Species within and outside the family Cryphonectriaceae, Cry-phonectria radicalisandValsa ceratosperma, also supported the replication of CpMV1 at a level comparable to that in the natural host. No stable maintenance of CpMV1
was observed inHelminthosporium victoriae. PCR-based haplotyping of virus-infected
fungal strains confirmed the recipient mitochondrial genetic background. Phenotypic comparison between CpMV1-free and -infected isogenic strains revealed no overt
ef-fects of the virus. Taking advantage of the infectivity to the standard strain C.
para-sitica EP155, accumulation levels were compared among antiviral RNA silencing-proficient and -deficient strains in the EP155 background. Comparable accumulation levels were observed among these strains, suggesting the avoidance of antiviral RNA silencing by CpMV1, which is consistent with its mitochondrial replication. Collec-tively, the results of study provide a foundation to further explore the biology of mi-toviruses.
IMPORTANCE Capsidless mitoviruses, which are ubiquitously detected in filamen-tous fungi, have the simplest RNA genomes of 2.2 to 4.4 kb, encoding only RNA-dependent RNA polymerase. Despite their simple genomes, detailed biological char-acterization of mitoviruses has been hampered by their mitochondrial location within the cell, posing challenges to their experimental introduction and study. Here we developed a protoplast fusion-based protocol for horizontal transfer of the pro-totype mitovirus, Cryphonectria parasitica mitovirus 1 (CpMV1), which was isolated
from strain NB631 of the chestnut blight fungus (Cryphonectria parasitica), a model
filamentous fungus for studying virus-host interactions. The host range of CpMV1
has been expanded to many different strains of C. parasitica and different fungal
species within and outside the Cryphonectriaceae. Comparison of CpMV1 accumula-tion among various RNA silencing-deficient and -competent strains showed clearly that the virus was unaffected by RNA silencing. This study provides a solid founda-tion for further explorafounda-tion of mitovirus-host interacfounda-tions.
KEYWORDS Cryphonectria parasitica, RNA silencing, chestnut blight, mitochondria, mitovirus, mycovirus
CitationShahi S, Eusebio-Cope A, Kondo H, Hillman BI, Suzuki N. 2019. Investigation of host range of and host defense against a mitochondrially replicating mitovirus. J Virol
93:e01503-18.https://doi.org/10.1128/JVI
.01503-18.
EditorAnne E. Simon, University of Maryland, College Park
Copyright© 2019 American Society for
Microbiology.All Rights Reserved.
Address correspondence to Nobuhiro Suzuki, [email protected].
*Present address: Ana Eusebio-Cope,
Livelihood, Gender and Nutrition Cluster, Sustainable Impact Platform, International Rice Research Institute, Manila, Philippines. S.S. and A.E.-C. contributed equally to this work.
Received29 August 2018
Accepted18 December 2018
Accepted manuscript posted online9 January 2019
Published
crossm
5 March 2019
on November 6, 2019 by guest
http://jvi.asm.org/
M
itoviruses (genusMitovirus, familyNarnaviridae) comprise a unique group of RNA viruses with small and simple positive-sense RNA genomes that are 2.2 to 4.4 kb with a single open reading frame (ORF) encoding a viral RNA-dependent RNA poly-merase (RdRp) (1). Mitoviruses are the only viruses that can infect eukaryotic mitochon-dria, and they are characterized by their capsidless nature and possible completion of their replication cycle within host mitochondria. Most utilize the codon UGA to code for tryptophan rather than using it as a stop signal, consistent with mitochondrial codon usage in fungi. Mitoviruses are omnipresent fungal viruses (see, e.g., references 2 to 6), but they were detected in plants only recently as likely infectious entities (7). Mitovirus genome sequences are also detected in host mitochondrial and nuclear genomes in plants and fungi (6, 8–11), like other nonretrovirus RNA viruses in plants and animals (12–14). It is noteworthy that many mitovirus strains belonging to over 20 species areisolated even from one necrotrophic ascomycete fungal species,Sclerotinia sclerotiorum
(15). Different mitovirus strains often coinfect single fungal strains, as exemplified by
those infectingOphiostoma novo-ulmi(Dutch elm disease fungus) orS. sclerotiorum(5,
16, 17). Phylogenetically related narnaviruses (genusNarnavirus, familyNarnaviridae),
ourmiaviruses (genusOurmiavirus), and leviviruses (familyLeviviridae) have been shown
to replicate in the cytoplasm of yeast, plant, and bacterial cells, respectively (1, 18). A new group of ourmia-like viruses also have been reported from filamentous fungi (see, e.g., reference 19), but little information about them is available.
Despite an increasing number of mitoviruses detected, only a few mitoviruses have been biologically investigated. For example, some mitoviruses, such as Botrytis mito-virus 1 (BcMV1) (formerly Botrytis cinerea debilitation-related mito-virus) and Sclerotinia sclerotiorum mitovirus 1, cause severe symptoms, including hypovirulence in the host fungi (9, 20, 21), while asymptomatic infections were suggested for others (3). BcMV1 induces malformation of infected mitochondria and reduces virulence of the host fungus (21). There are still many unanswered fundamental questions in general about their replication cycles, including (i) how broadly a specific mitovirus can infect and (ii) the kinds of antimitovirus host defense and counterdefense that possibly occur in mitovirus-infected cells. Initial evidence to answer the first question came when a
mitovirus from the fungusSclerotinia homeocarpa(dollar-spot fungus) was found to be
conspecific with a previously characterized element, Ophiostoma novo-ulmi mitovirus
3a-Ld, from the distantly related ascomyceteO. novo-ulmi(22). Similarly, the mitovirus
Ophiostoma novo-ulmi mitovirus 3b was identified in the two taxonomically different
fungiO. novo-ulmiandBotrytis cinerea(gray mold fungus) (21). Thus, it was clear not
only that such mitochondrial elements could in some cases enter, replicate, and be maintained in heterologous hosts but also that they did so with relatively little change to their genomes. Another interesting insight into mitovirus host range was provided through a bioinformatic analysis. Nibert (23) recently showed a direct relationship between the frequency of UGA codon utilization by mitoviruses and their host mito-chondria. That study implied a possibility that an absence or scarcity of mitochondrial tRNA decoding UAG for Trp restricts infection of mitovirus with internal UGA codons. Addressing the second question, of host defense, many fungal viruses replicating in the cytosol are known to be targeted by RNA silencing, as in other eukaryotic viruses (24–27). In this regard, mitovirus-derived small RNAs were shown to be produced for a few mitoviruses, including Heterobasidion mitovirus 1 and BcMV1 (28–30). However, the important question of whether mitochondrial mitoviruses are interfered with by cytoplasmic RNA silencing has yet to be addressed. One of the constraints hampering the advancement of mitovirus research is the lack of artificial introduction methods for mitochondrial viruses, i.e., inoculation methods. This contrasts with the fact that virion or RNA transfection methods or infectious cDNA clones have increasingly been estab-lished for fungal RNA viruses (26, 27, 31–36).
Cryphonectria parasitica mitovirus 1 (CpMV1) (type member of the genusMitovirus)
was the first mitovirus characterized; it was isolated from strain NB631 of the chestnut
blight fungus,Cryphonectria parasitica(order Diaporthales) (2).C. parasiticais important
not only as a plant pathogen causing one of the three most destructive tree diseases
Shahi et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
in the world but also as a virus host having biological tractability (37). CpMV1 was
transmitted vertically to ascospores at a rate of ⬃50% in genetic crosses between
NB631 and anotherC. parasiticastrain, EP155, when infected strain NB631 was used as
a female parent in the crosses (38). This is consistent with maternal inheritance of infected mitochondria but also suggests that not all mitochondria in a thallus are infected. In the current study, we developed a protocol for protoplast-based introduc-tion of the capsidless mitovirus CpMV1 and extended its host spectrum to different
fungal strains of C. parasitica and to different species within and outside the
Cry-phonectriaceae. The horizontal transfer of CpMV1 to theC. parasiticastandard strain
EP155, for which many mutant strains with knockouts of genes involved in the posttranscriptional gene silencing (PTGS) pathway are available, allowed us to suggest the avoidance of antiviral RNA silencing by CpMV1.
RESULTS
Development of a protoplast fusion-based method for CpMV1 lateral trans-mission.Because of the capsidless nature of CpMV1, potential cell-free introduction methods are limited to nonvirion transfection methods. Given the difficulty developing infectious cDNA or synthetic transcripts for mitoviruses (17), we tested a protoplast fusion protocol for CpMV1 that was reported previously for another capsidless virus,
Cryphonectria hypovirus 1 (CHV1) (family Hypoviridae) (39). The protocol entailed
labeling of recipient fungal strains with a hygromycin B resistance (Hygr) gene,
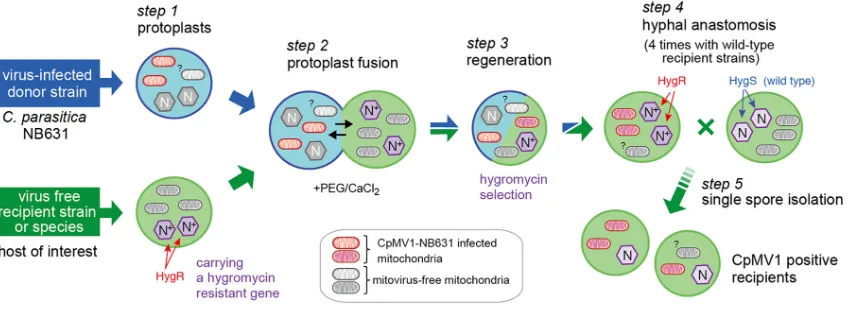
proto-plast fusion, selection of the recipient genetic backgrounds, and repeated pairing with recipient strains followed by single conidial isolation (Fig. 1). We first examined this
method for theC. parasiticastandard strain, EP155, which is vegetatively incompatible
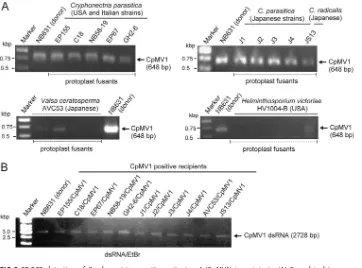
with NB631, the original host of CpMV1. EP155 fungal cultures were tested for the presence of CpMV1 at each step of the procedure shown in Fig. 1. Five fungal cultures obtained after protoplast fusion step 3 carried the virus when tested by reverse transcription-PCR (RT-PCR) (Fig. 2A). Two of the CpMV1-positive cultures were moni-tored further, and the virus was confirmed to be retained after repeated (4 rounds) anastomosis and single conidial isolation (step 4). Five single-spore isolates derived from the two cultures were subjected to double-stranded RNA (dsRNA) isolation (step
5), and 100% of them tested CpMV1 positive and carried the Hygrgene (Fig. 2B and
data not shown). These results showed that the protocol is useful for lateral transfer of CpMV1 between vegetatively incompatible strains.
This technique was used for 9 other strains ofC. parasiticaand one strain each of
Cryphonectria radicalis, Valsa ceratosperma (the fungus of apple Valsa canker, strain
FIG 1Cartoon showing the experimental protocol used for the transfer of Cryphonectria parasitica mitovirus 1 (CpMV1) from NB631 to different fungal strains through protoplast fusion. Protoplasts were prepared from the naturally CpMV1-infected strain NB631 (donor) and a hygromycin B-resistant (Hygr) recipient strain (Table 1) (step 1) and fused with the aid of PEG (step 2). The resultant fusants were grown on PDA-hygromycin B plates (step 3) and anastomosed with virus-free, hygromycin B-susceptible (Hygs) recipient strains (step 4). After 1 week, mycelial plugs from the recipient were anastomosed again with virus-free recipient strains. This paring was repeated 4 times. The final step was single conidial isolation (step 5). Detailed procedures are described in Materials and Methods.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:3.585.44.473.72.228.2]AVC53), andHelminthosporium victoriae(Victoria blight fungus, strain HV1004-B) (Table
1), all of which belong to the division Ascomycota. All tested strains exceptH. victoriae
were shown to stably support CpMV1 replication. CpMV1 was detectable at each step of the procedure shown in Fig. 1 for most fungi tested (RT-PCR results at step 3 are
[image:4.585.41.403.70.338.2]FIG 2RT-PCR detection of Cryphonectria parasitica mitovirus 1 (CpMV1) in recipients. (A) Fungal isolates were obtained from different batches of protoplast fusion (step 3 in Fig. 1) between the donor strain NB631 and recipients shown above the gels (Table 1). RT-PCR was designed to amplify a genomic region from map position 1290 to 1937 (see Materials and Methods for the primer sequences). (B) Fungal strains obtained from step 5 (Fig. 1) were subjected to dsRNA isolation and agarose gel electrophoresis. The 1-kb ladder DNA (Gene Ruler; Thermo Fisher Scientific, Waltham, MA) was used as a size marker in this and subsequent figures.
TABLE 1Fungal strains tested in this study for the ability to support CpMV1 replication
Family Species Strain
Collection site or source
Strain no. or accession no.
Susceptibility
to CpMV1 vicgenotyped Reference(s)
Cryphonectriaceae Cryphonectria parasitica NB631 New Jersey, USA Yes 1112-11 2 EP155 Connecticut, USA ATCC 38755 Yes 2211-22 Standard
strain Δdcl1mutant EP155 derivative NAc Yes NTe 25
Δdcl2mutant EP155 derivative NA Yes NT 25 Δagl2mutant EP155 derivative NA Yes NT 24 Δrdr1mutant EP155 derivative NA Yes NT 58 EP67 Italy ATCC 38753 Yes 2212-22 62, 63 GH2-6a Michigan, USA AF188515 Yes 1211-11 64
NB58-19a New Jersey, USA ATCC 76221 Yes 2112-11 65
C18-VFa West Virginia, USA AB073281 Yes 2111-11 66
1-1 (J1) Ibaraki, Japan MAFF 410878 Yes 1211-21 67 J2 Kanagawa, Japan MAFF 305108 Yes 2222-21 NA EP48 (J3) Hokkaido, Japan MAFF 410557 Yes 2211-22 NA A1 (J4) Ibaraki, Japan MAFF 410726 Yes 1221-11 NA
Cryphonectria radicalis JS13-VFa Kyoto, Japan MYA-4104 Yes 52
Valsaceae Valsa ceratosperma AVC53 Aomori, Japan NA Yes 32, 68 Massarinaceae Helminthosporium victoriae Hv1004-Hygrb USA ATCC 42020 No 69 aVirus-free strains were obtained by single-conidium isolation from the originally virus-infected field isolates.
bTransformant of strain B2 with an empty vector, pCB1004, carrying a Hygrgene.
cNA, not available.
dvicgenotypes at 6 diallelic loci were determined as described by Short et al. (70), and their nomenclature is according to Choi et al. (71). eNT, not tested.
Shahi et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:4.585.41.547.497.695.2]shown in Fig. 2A). The presence of CpMV1 for recipient strains (step 5) was confirmed by RT-PCR and dsRNA agarose gel analysis (Fig. 2B and data not shown). However, after
protoplast fusion withH. victoriaestrain HV1000-B, CpMV1 was detected only until step
3 by RT-PCR and was undetectable at later steps. It is also noteworthy that
CpMV1-infected V. ceratosperma strain AVC53 occasionally lost the virus after prolonged
storage (4 to 6 weeks) (data not shown).
PCR-based mitochondrial haplotyping.Mitochondria are believed to be prone to
fusion and rearrangements, and this has been suggested forC. parasitica(38). During
the process of protoplast fusion, heterokaryon formation and hybrid formation are likely to occur, and there are also chances of mixing the mitochondrial materials. Therefore, it is important to determine the mitochondrial genetic background in the CpMV1-positive recipients obtained after protoplast fusion. PCR-based mitochondrial haplotyping (mitohaplotyping) was carried out, in which a polymorphic NADH
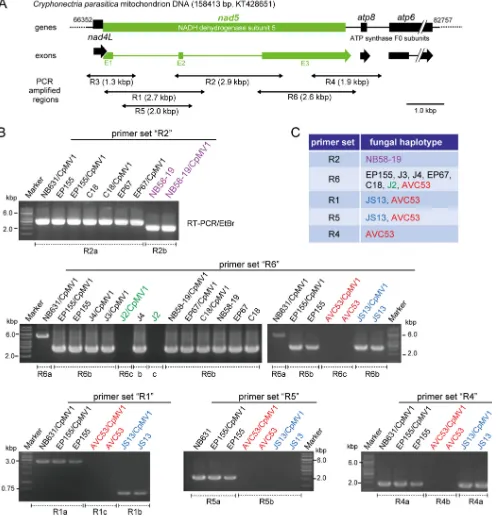
dehy-drogenase subunit 5 gene (nad5) region (Fig. 3A) was targeted. Mitochondrial DNAs
(mtDNAs) extracted from protoplast fusion isolates were PCR amplified using primer sets (R1 to R6) specific for the mitochondrial gene. A combination of primer pairs could distinguish five haplotypes within the order Diaporthales (Fig. 3B). Representative gel
patterns are shown in Fig. 3B. For example,C. radicalisstrain JS13 was differentiated
from other fungal strains by primer set R1, while set R2 could distinguish NB58-19 from
all otherC. parasiticastrains. All primer sets failed to amplify DNA fragments on mtDNA
ofV. ceratosperma. Note that a primer set for the mitochondrial small-subunit ribosomal gene could amplify a fragment of the expected size in all tested fungal strains (data not
shown). Primer set R6 could differentiate the banding pattern ofC. parasiticaNB631
from those of other fungal strains (Fig. 3B). Importantly, all CpMV1-positive recipient strains possessed the same haplotypes as their original recipient strain used for protoplast fusion. These results indicated no sign of recombination within the tested mitochondrial gene region or mixed populations of mitochondria.
CpMV1 accumulation in newly established fungal hosts and their phenotypes.
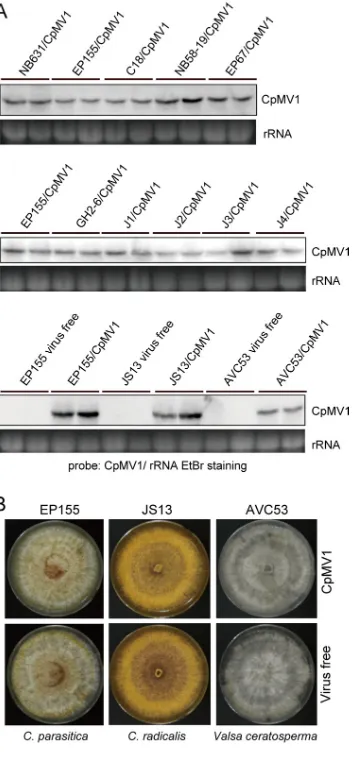
A total of 12 fungal strains, including those belonging to different fungal species, were confirmed to maintain CpMV1. These newly established fungal strains were tested for virus accumulation level by Northern blot analysis. Virus accumulation levels of
Amer-ican, Italian, and JapaneseC. parasiticastrains were similar and were comparable to that
observed in the original host of CpMV1, NB631, or the standard strain EP155. However,
the CpMV1 accumulation level in V. ceratosperma (AVC53) appeared to be
approxi-mately 4-fold lower than that inC. radicalis(JS13) and the EP155 strain (Fig. 4A).
To test possible effects of CpMV1 infection on the host phenotype, we compared the colony morphologies of isogenic pairs of CpMV1-infected and CpMV1-free fungal
strains. Pictures of colonies of representative strains of each of three fungal species (C.
parasiticaEP155,C. radicalisJS13, andV. ceratospermaAVC53) are shown in Fig. 4B. No obvious difference in colony morphology was observed between CpMV1-infected and uninfected fungal strains (Fig. 4B).
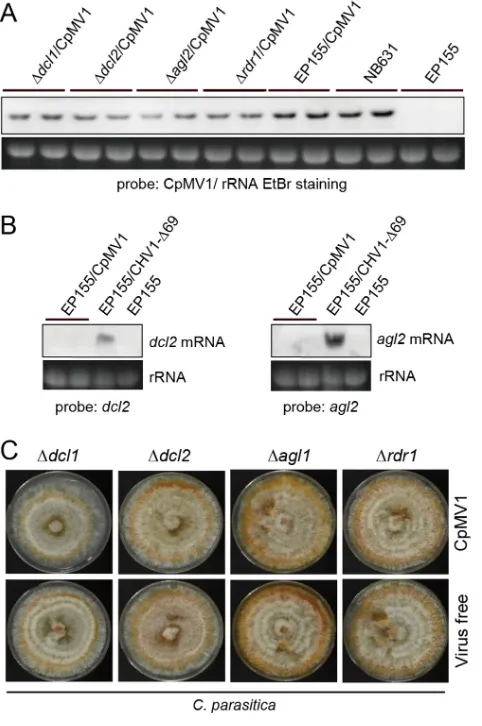
No enhanced accumulation of CpMV1 in fungal mutants lacking RNA silencing-related genes.RNA silencing is a primary antiviral defense mechanism in fungi (24, 25, 40) and is expected to operate in the cytosol of infected cells. Since CpMV1 is a mitochondrial virus, it is important to know whether CpMV1 is targeted by host RNA silencing. To investigate this possibility, CpMV1 was transferred to mutants of EP155
with deletions in the RNA silencing-associated genes (Δdcl2, Δdcl1, Δagl2, and Δrdr1) by
hyphal fusion with CpMV1-infected EP155 as a donor (Fig. 5). Hyphal fusion with each of the RNA silencing mutants was repeated 4 times to ensure homogeneity of the nuclear genetic background (26). Comparative analyses showed no difference in CpMV1 genomic RNA accumulation between RNA silencing-competent (EP155) and mutant strains (Fig. 5A). This result suggested that CpMV1 is not affected by the RNA silencing mechanism, suggesting that CpMV1 may avoid host RNA silencing.
Previous studies have shown that infection of EP155 by CHV1 and several other
viruses highly upregulatesdcl2andagl2transcription (24, 40–43). To investigate this in
the case of CpMV1, accumulations ofdcl2 andagl2 transcripts in EP155 and CpMV1/
on November 6, 2019 by guest
http://jvi.asm.org/
EP155 were assessed by Northern blot analysis (Fig. 5B). The results showed that CpMV1
did not inducedcl2 andagl2 transcripts, whereas transcription levels were remarkably
elevated by CHV1 Δp69, a mutant lacking the RNAi suppressor. This result suggests that CPMV1 escapes host RNA silencing by failing to induce silencing pathway genes and by avoiding their potential activity by remaining sequestered in mitochondria during
FIG 3PCR-based mitochondrial haplotyping of different fungal strains. (A) Physical map of a polymorphic region of theC. parasiticamitochondrial genome containing four genes (nad4L,nad5,atp8, andatp6). The specific primer sets R1 to R6 were adopted from reference 60. (B) Agarose gel profiles of PCR products with specific primer pairs. mtDNA was prepared from each of the CpMV1 recipient fungal strains obtained from step 5 (Fig. 1) and the original virus-free recipient strains and used as templates in PCR. Names of fungal strains are shown above the gel lanes. PCR fragments were amplified from all fungal isolates when a primer set (forward, 5=-GTGCCAGCAGTCGCGGYAANAC-3=; reverse, 5=-GGTGRARTGCTTNCACTTTCATTTATA-3=) for the mitochondrial small-subunit ribosomal gene was used (data not shown). (C) Primer sets used to distinguish mitochondrial haplotypes of CpMV1 recipients.
Shahi et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:6.585.43.541.74.594.2]replication. Moreover, phenotypic characterization of these strains showed no differ-ence between virus-free and CpMV1-infected strains (Fig. 5C).
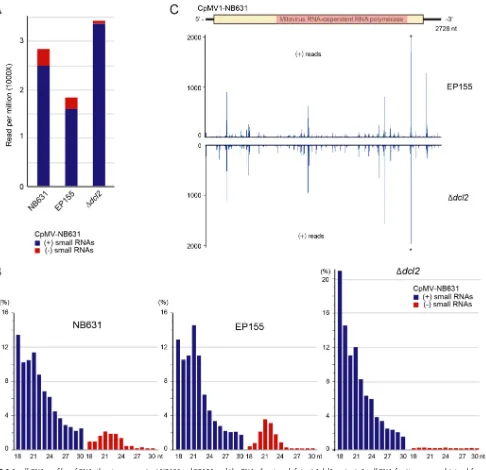
Production of CpMV1-derived small RNAs in infected fungal strains.Small RNAs were prepared from four fungal strains, i.e., virus-free EP155, CpMV1-infected EP155,
CpMV1-infected NB631, and the CpMV1-infected Δdcl2mutant, and sequenced by a
next-generation sequencing (NGS) method. DCL2-dependent virus-derived small RNA profiles are characterized by the production of viral minus-strand small RNAs as well as viral plus-strand small RNAs with a peak of 21 nucleotides (nt) (44, 45).
Comparison of small RNA profiles for the three fungal strains showed accumulation of CpMV1-derived small RNAs in RNA silencing-competent EP155 and NB631 and in the
RNA silencing-deficient Δdcl2mutant (Fig. 6A) but not in virus-free EP155 (data not
shown). The profiles of small RNAs for CpMV1-infected NB631 and EP155 manifested a peak of 21 nt representing viral minus-strand RNA expected from the DCL2-dependent pathway, but few 21-nt viral minus-strand RNAs were detected from the
CpMV1-infected Δdcl2mutant (Fig. 6B). We recently analyzed small RNA profiles for a mutant
(Δp69) of the prototype CHV1 that replicates in the nonmitochondrial cytoplasm
FIG 4Northern blot and phenotypic analyses of Cryphonectria parasitica mitovirus 1 (CpMV1) in different fungal hosts. (A) Northern blotting of CpMV1. Total RNA was prepared from CpMV1-infected fungal strains obtained from step 5 (Fig. 1) and probed with a DIG-labeled dsDNA fragment that corresponds to the CpMV1 genomic region from position 1290 to 1937. Ethidium bromide (EtBr)-stained 28S rRNA (rRNA) served as a loading control for this and subsequent figures. (B) Colony morphologies of isogenic CpMV1-infected and -free fungal strains. Only representative fungal pairs from three different fungal species are shown.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:7.585.117.296.71.450.2](Andika et al., submitted). Similar patterns were observed between CpMV1-infected
EP155 or Δdcl2mutant and for CHV1-infected EP155 or Δdcl2mutant, respectively. A
notable difference was sharper peaks of 21-nt CHV1 Δp69-derived small RNAs in EP155 than of 21-nt CpMV1-derived small RNAs. This difference likely reflects a ratio of CHV1 Δp69-derived small RNAs to total small RNAs that is 4-fold greater than that of CpMV1-derived small RNAs to total small RNAs in EP155.
CpMV1-derived small RNAs detected from the Δdcl2 mutant could be products
digested by DCL2 independently or degradation products during small RNA prepara-tion. Of note is that CpMV1 positive-strand-RNA-derived small RNAs from EP155 and
the Δdcl2mutant had similar peak patterns (Fig. 6C). This observation may suggest that
CpMV1 positive-strand RNA is cleaved specifically, rather than randomly, in a DCL2-independent manner and that adequate precautions are needed to interpret small RNA profiles.
DISCUSSION
Over the past few decades, virion transfection protocols have been developed for diverse mycovirus-host combinations (46, 47). This virion transfection, however, is not
FIG 5Relationship between RNA silencing and CpMV1 infection. (A) Northern blotting of
CpMV1-infected fungal strains disrupted for RNA silencing-associated genes. CpMV1 was transferred from EP155 to mutants with disruptions of thedcl(Δdcl1and Δdcl2),agl(Δagl2), andrdr(Δrdr1) genes, as reported earlier (24, 25, 58). CpMV1-infected fungal strains were then subjected to Northern blotting as described for Fig. 4. CpMV1-infected NB631 and virus-free EP155 were included as references. (B) Effects of CpMV1 infection on dcl2 and agl2 transcript levels. Total RNA was isolated from CpMV1-infected EP155, CHV1-infected EP155, and virus-free EP155.dcl2andagl2mRNA levels were compared among the three strains. (C) Phenotypes of fungal colonies infected and uninfected with CpMV1.
Shahi et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:8.585.85.326.72.435.2]applicable for capsidless viruses, and instead, infectious full-length cDNA or synthetic RNA produced from it has been established for capsidless hypoviruses and
yadokari-viruses (calicivirus᎑like positive-sense single-stranded RNA [ssRNA] viruses, proposed
family Yadokariviridae) (31, 33–35). Capsidless nonsegmented mitoviruses are
phylo-genetically related to capsidless nonsegmented narnaviruses ofSaccharomyces
cerevi-siae(members of the genusNarnavirus) and encapsidated, multisegmented
ourmiavi-ruses of dicot plants (members of the genusOurmiavirus), both of which are replicated
in the nonmitochondrial cytoplasm. Infectious full-length cDNA was developed for
members of the genera NarnavirusandOurmiavirus(48–50). However, no infectious
cDNA clones or synthetic transcripts are available for mitoviruses thus far, although they can be transmitted vertically via spores and horizontally via hyphal fusion only
FIG 6Small RNA profiles of RNA silencing-competent NB631and EP155 and the RNA silencing-deficient Δdcl2mutant. Small RNA fractions were obtained from
CpMV1-infected strains and analyzed (see Materials and Methods). (A) Plus-strand and minus-strand small RNAs (18 to 30 nt) derived from CpMV1 are shown on a per-million-total-small-RNA basis. Blue and red bars denote plus-strand and minus-strand small RNAs. (B) Size distributions of CpMV1 small RNAs. (C) Distributions of CpMV1 small RNAs along the CpMV1 genome. A diagram of the CpMV1 genome is shown at the top. CpMV1-derived plus-strand small RNAs from EP155 and the Δdcl2mutant were mapped to its genome, showing very similar distribution patterns.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:9.585.49.535.69.539.2]between vegetatively compatible strains within a single species (2, 38). Therefore, the protoplast fusion protocol for artificial introduction developed in this study is of great importance for studies of host range and mitovirus-host interactions, because it allows introduction of virus into any host fungus, independent of hyphal incompatibility. Note that protoplast fusion was earlier utilized for lateral transmission of CHV1 and other, unidentified viruses (39, 51, 52).
CpMV1 has been reported only from oneC. parasiticastrain, NB631, and not from
other C. parasitica strains despite extensive surveys for fungal viruses (53, 54). The
current study allowed horizontal transfer of a mitovirus experimentally to a new host
and expanded its host range to an order different from the one to whichC. parasitica
belongs (Table 1). All nine testedC. parasiticaisolates, collected from the United States,
Europe, and Japan were found to be susceptible to CpMV1. Another species within the
genusCryphonectria,C. radicalis, supported the replication of CpMV1, and its
accumu-lation level was comparable to that in C. parasitica. A slightly lower accumulation
pattern was observed forV. ceratosperma, belonging to the family Valsaceae, which is
different from that ofC. parasitica(Cryphonectriaceae). Stable maintenance of CpMV1
was observed in these newly established hosts. This observation contrasts with the situation of cytoplasmically (nonmitochondrially) replicating RNA mycoviruses whose
original hosts are C. parasiticaor Rosellinia necatrix (white root rot fungus) and are
experimentally introduced intoV. ceratosperma. For example, mycoreovirus 1 (a
reovi-rus [encapsidated, multisegmented dsRNA vireovi-rus]), CHV1, and Rosellinia necatrix parti-tivirus 6 (a betapartiparti-tivirus [encapsidated, two-segmented dsRNA virus]) replicate but
are not stably maintained in V. ceratosperma (41). As noted above, examples of
conspecific mitovirus strains found through surveys in taxonomically distinct fungi
belonging to the class Sordariomycetes (Ophiostoma) or Leotiomycetes (Sclerotiniaand
Botrytis) (21, 22) suggested a broader host range for mitoviruses than has been recorded for other mycoviruses. Our current study experimentally demonstrated that one single mitovirus strain, CpMV1 NB631, can infect fungi belonging to different families (Valsaceae and Cryphonectriaceae) within the order Diaporthales. Analysis performed by Nibert (23) implied that infrequency of Trp-encoding UGA codons in mitochondria of some fungi reflects the infrequency of corresponding tRNAs encoded by them and that this in turn restricts the host range of a mitovirus that contains an abundance of Trp-encoding UGA codons. The protoplast fusion technique described in this study will facilitate testing of this hypothesis.
We showed the mitochondrial and nuclear genetic backgrounds of recipients, obtained after protoplast fusion, to be the same as those of the original recipient strains before protoplast fusion, as judged or expected from mitochondrial haplotyping and repeated anastomosis and single-spore isolation (Fig. 1). A total of 5 mitochondrial haplotypes of CpMV1 recipients were distinguishable by PCR products derived from a polymorphic region (Fig. 3C). An interesting finding was that all CpMV1-positive recipients show only one single haplotype identical to that of the original recipient, and neither donor strain haplotypes nor mitochondrial DNA rearrangements, which are often reported for mitochondria resulting from protoplast fusion, were observed for the
polymorphicnad5region examined (Fig. 3B). Loss of mitochondrial genomes from one
partner after protoplast fusion is not uncommon, as exemplified by somatic hybrid
plants of Nicotianaspp. andSolanumspp. (55, 56). Our results suggest that there is
compatibility between genotypes of mitochondria and nuclei, as expected from the fact that many genes whose products are functional in mitochondria are carried by the nuclear genome. Also, the results confirm that mitochondria of the extended hosts discussed above can support replication of CpMV1.
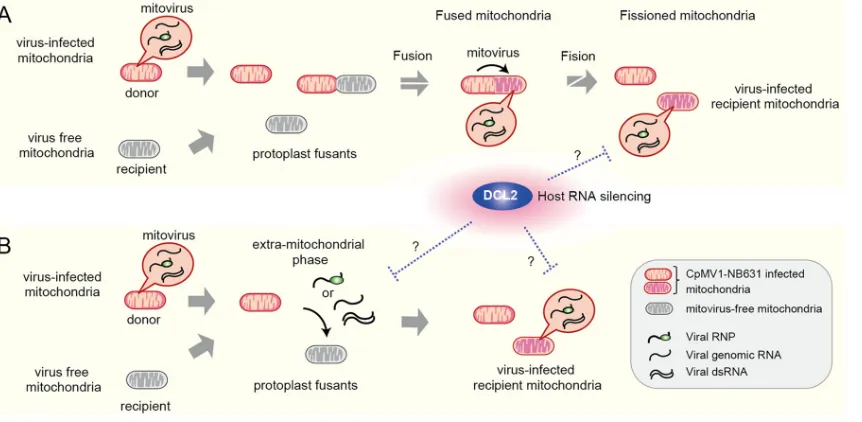
Polashock et al. (38) proposed two possibilities for the CpMV1 transmission mode with the premise that CpMV1 cannot move bidirectionally across the mitochondrial membrane: (i) partial or total replacement of uninfected mitochondria (recipient) with infected ones (donor) or (ii) fusion of infected and noninfected mitochondria, resulting in CpMV1 transmission to the latter. Our results are not consistent with the first scenario but rather favor the second one, that mitochondrial fusion is a mechanism for CpMV1
Shahi et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
transmission to a new host while maintaining the recipient mitochondrial haplotype (Fig. 7A). We cannot completely rule out the possibility that CpMV1 has an extramito-chondrial phase in the cytoplasm and reentry into uninfected recipients’ mitochondria (Fig. 7B). An extramitochondrial phase has been proposed for the presumed mitovirus
M2 fromRhizoctonia solanibased on initial finding of more M2 dsRNA in the
nonmi-tochondrial cytosolic fraction than in the minonmi-tochondrial fraction (11), but follow-up subcellular localization studies have not been published, and details remain unclear. As discussed below, this idea is supported by the observation that DCL2 could produce CpMV1-derived small RNAs and by a recent report that endogenous mitochondrial dsRNA can be exported into the cytosol (57). Either scenario proposed in Fig. 7 includes open questions and needs to be validated in the future.
Investigation of antiviral RNA silencing versus mitoviruses is of great interest from several perspectives. If mitoviruses have a cytosolic phase in their replication cycles, though seemingly unlikely, they would be targeted by RNA silencing. In this context, some recent studies have reported mitovirus-derived small RNAs produced in infected fungi (28, 30), suggesting the accessibility of Dicer to mitovirus RNAs. DCL2-dependent CpMV1-derived small RNAs were also detectable in infected EP155 (Fig. 6). The replicative form (dsRNA) of CpMV1 may be exposed in the cytosol at an unknown step of replication or by accidental release of dsRNAs from mitochondria. In support of this notion, long mitochondrial dsRNAs (endogenous, bidirectional transcription products) were recently shown to be excreted into the cytoplasm and to induce a dsRNA sensor-mediated interferon type I antiviral response in humans (57). The next question is whether CpMV1 is really targeted by RNA silencing. This study allowed us to address this question, because CpMV1 could be moved laterally to the
standard EP155 strain of C. parasitica, for which many mutant strains are available.
Many cytoplasmically (nonmitochondrially) replicating RNA viruses have been shown to be targeted by antiviral RNA silencing and to transcriptionally induce RNA silencing
genes in C. parasitica. Only a few viruses, such as Rosellinia necatrix victorivirus 1
(RnVV1) (a victorivirus [nonsegmented dsRNA virus]), fail to inducedcl2andagl2(26,
40). Like RnVV1, CpMV1 induced neitherdcl2noragl2(Fig. 5B). Figure 5A and C show
comparable accumulation levels of CpMV1 between RNAi-proficient (EP155) and
FIG 7Model of CpMV1 infection of recipient mitochondria. (A) Infection of recipient mitochondria by CpMV1 initiated via fusion with donor mitochondria. CpMV1-infected recipient mitochondria become dominant via fission. (B) Infection of recipient mitochondria by CpMV1 mediated by its extramitochondrial entities, genomic RNA-associated or nonassociated RdRp molecules, or dsRNA. The extram-itochondrial dsRNA of CpMV1 could be diced by DCL2. Considering the production of CpMV1-derived small RNAs (Fig. 6), there should be extramitochondrial dsRNA of CpMV1, which is accessed in the cytosol by DCL2, even in model A. The two possibilities (A and B) are not mutually exclusive.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:11.585.44.475.72.283.2]-deficient (Δdcl2and Δagl2mutant) strains, and no phenotypic alteration was observed in these strains. These results contrast with the situation of other cytoplasmically (nonmitochondrially) replicating viruses, including RnVV1, that usually show higher levels of virus accumulation and more severe symptoms in RNAi-deficient strains. Collectively, this study suggests the possible avoidance of host antiviral RNA silencing, consistent with its mitochondrial replication.
MATERIALS AND METHODS
Fungal strains.The CpMV1-infected U.S. strain NB631 ofC. parasiticawas described earlier (2). The fungal strains used in this study (Table 1) included the standardC. parasiticastrain EP155, which is used extensively in many laboratories, EPI55-derived deletion mutants, and field-collected strains of C. parasiticafrom the United States (EP155, C18, GH2, and NB58-19), Italy (EP67), and Japan (J1, J2, J3, J4). Strains of some other ascomycetous fungi, such asC. radicalis(JS13),V. ceratosperma(AVC53), andH. victoriae(HV1004-B), were also used.C. parasiticaandC. radicalisshare the same niche, where both thrive in chestnut trees.V. ceratospermabelongs to the family Valsaceae, whereasCryphonectriaspp. belong to the family Cryphonectriaceae, but they are within the same order Diaporthales.H. victoriaebelongs to the order Pleosporales.
Protoplast fusion.All tested fungal strains (Table 1) were transformed with pCPXHY2 carrying a
hygromycin B phosphotransferase gene (hph) to obtain hygromycin B-resistant (Hygr) strains. An equal number of protoplasts (approximately 2⫻107) from the CpMV1 donor strain NB631 and a virus-free Hygr recipient strain were fused in the presence of polyethylene glycol (PEG)-CaCl2. The protoplast fusants were regenerated on the regeneration medium and were allowed to grow overnight. The next day, top agar containing 40g/ml hygromycin B was overlaid on regeneration medium for screening the recipient (host of interest). Protoplast regenerants were then allowed to grow over top agar for several days. Hygrmycelia growing over the top agar were transferred into new potato dextrose agar (PDA) plates containing 80g/ml hygromycin B and incubated for 3 to 5 days before detection of CpMV1. After confirming the presence of CpMV1, CpMV1-positive colonies were anastomosed with the original nontransformed recipients that were hygromycin B susceptible (Hygs) and CpMV1 free. Mycelial plugs taken from the recipient’s side were fused again with the original CpMV1-free recipient. Karyons are known to be transferred between donors and recipients (26). A total of four rounds of this hyphal fusion procedure were carried out for each of the fungal strains to avoid carryover of karyons from donors. Finally, single conidial isolates from each of 11 CpMV1-infected fungal hosts were prepared to obtain monokaryons and were used in subsequent characterization (Fig. 1). Steps 1 and 2 shown in Fig. 1 are commonly used for protoplast fusion experiments. However, the objective of this procedure is to follow the mitochondrial lineage from CpMV1-infected monokaryotic recipients, not from CpMV1-infected heterokaryotic fusants.
RT-PCR detection of CpMV1.cDNA was synthesized using Moloney murine leukemia virus reverse
transcriptase (Thermo Fisher Scientific, Invitrogen, Waltham, MA) and a primer, 5=-GGAATTACCTCCAAA C-3=, that corresponds to the RdRp-coding region (positions 1951 to 1966) of CpMV1 NB631 (GenBank accession no. NC_00404). PCR amplification was carried out using QuickTaq(Toyobo, Osaka, Japan) along with primer set 5=-GCAACGGATAGACTTC-3=(map positions 1290 to 1305) and 5=-AAATAGCCTG ATGCCA-3=(map positions 1922 to 1937).
Northern blotting.Northern blotting was performed as described by Eusebio-Cope and Suzuki
(57). The heat-denatured ssRNA fraction (2g) was electrophoresed in a 1% agarose gel under denaturing conditions. The separated RNAs were capillary transferred onto a Hybond-N⫹nylon membrane (GE Healthcare Life Sciences, Pittsburgh, PA) and fixed by a UV cross-linker, CL-1000 (UVP). The membrane was then treated with digoxigenin (DIG)-labeled DNA probes amplified from cDNA by PCR (PCR DIG labeling mix; Roche, Risch-Rotkreuz, Switzerland). The primer set is described above for CpMV1 and was described earlier for thedcl2andagl2mutants (40). Prehybridization and hybridization steps were performed based on the information provided by the supplier (Roche, Risch-Rotkreuz, Switzerland). Two low- and high-stringency washes were carried out in 2⫻SSC (1⫻ SSC is 0.15 M NaCl plus 0.015 M sodium citrate) and 0.1⫻SSC, respectively. Hybridized nucleic acids were detected with ready-to-use CDP-Star (Roche) through digital imaging in the ImageQuant LAS 4000 system (GE Healthcare Life Sciences).
Mitochondrial haplotyping.Mitochondrial DNA extraction was performed as described by Yamada
et al. (59) with slight modification. Fungal mycelia grown on liquid medium (potato dextrose broth [PDB]) were harvested after 3 days of inoculation on the benchtop. Mycelia were ground in the presence of liquid nitrogen with the help of pestle and mortar and were homogenized with extraction buffer (100 mM Tris HCl [pH 8.0], 1% SDS, 2% Triton X-100, 10 mM EDTA, 100 mM NaCl), followed by phenol-isoamyl alcohol and chloroform extraction. The supernatant was then precipitated by adding an equal amount of isopropanol and incubating the sample at room temperature for 1 h. The pellet was then washed twice with 70% ethanol and vacuum dried. Finally, the pellet was dissolved in MilliQ water and stored at⫺20°C until use.
Based on the paper published by Gobbi et al. (60), a highly polymorphic region, i.e., region 2 from mtDNA of EP155, was utilized to make different sets of primers within it (Fig. 3A) to distinguish recipient mitochondria from donor mitochondria. Various amounts of mtDNA (0.5 ng, 1 ng, 5 ng, and 10 ng) were used as templates, and conventional PCR was performed.
Analysis of small RNA sequences.Total RNA samples from threeC. parasiticastrains (NB631, EP155, and the Δdcl2mutant) infected with CpMV1 NB631 were subjected to small RNA sequencing. Small RNA
Shahi et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
cDNA library preparation and subsequent deep sequencing with the Illumina platform (HiSeq 2500; 50-bp single-end reads) were done by Macrogen Inc. (Tokyo, Japan). After trimming adapters and low-quality bases and size filtering (18 to 30 nt in length), the retained read sequences in each library were mapped to the CpMV1 NB631 genome (GenBank accession no. L31849) using CLC Genomics Workbench (version 11; CLC Bio-Qiagen). The virus-derived small RNA reads were used for in-depth analysis with the program MISIS-2 (61).
ACKNOWLEDGMENTS
This study was supported in part by Yomogi Inc., the Uehara Memorial Foundation, by Grants-in-Aid for Scientific Research on Innovative Areas from the Japanese Minis-try of Education, Culture, Sports, Science and Technology (KAKENHI 25252011, 15K14663, 16H06436, 16H06429, and 16K21723 to H.K. and N.S.), and by the New Jersey Agricultural Experiment Station and USDA-NIFA (to B.I.H.).
We are grateful to Donald L. Nuss, Said A. Ghabrial, and Satoko Kanematsu for
generous gifts of C. parasitica strains EP155 and the Δdcl2 and Δagl2 mutants, H.
victoriaestrain Hv1004-Hygr, andV. ceratospermastrain AVC53 and to Sotaro Chiba and
Ida Bagus Andika for fruitful discussions.
We declare that we have no conflicts of interest.
REFERENCES
1. Hillman BI, Cai G. 2013. The familyNarnaviridae: simplest of RNA viruses. Adv Virus Res 86:149 –176.https://doi.org/10.1016/B978-0-12-394315-6 .00006-4.
2. Polashock JJ, Hillman BI. 1994. A small mitochondrial double-stranded (ds) RNA element associated with a hypovirulent strain of the chestnut blight fungus and ancestrally related to yeast cytoplasmic T and W dsRNAs. Proc Natl Acad Sci U S A 91:8680 – 8684.https://doi.org/10.1073/ pnas.91.18.8680.
3. Wu M, Deng Y, Zhou Z, He G, Chen W, Li G. 2016. Characterization of three mycoviruses co-infecting the plant pathogenic fungusSclerotinia nivalis. Virus Res 223:28 –38. https://doi.org/10.1016/j.virusres.2016.06 .009.
4. Osaki H, Nakamura H, Nomura K, Matsumoto N, Yoshida K. 2005. Nucle-otide sequence of a mitochondrial RNA virus from the plant pathogenic fungus,Helicobasidium mompaTanaka. Virus Res 107:39 – 46.https://doi .org/10.1016/j.virusres.2004.06.010.
5. Hong YG, Dover SL, Cole TE, Brasier CM, Buck KW. 1999. Multiple mitochondrial viruses in an isolate of the Dutch elm disease fungus Ophiostoma novo-ulmi. Virology 258:118 –127.https://doi.org/10.1006/ viro.1999.9691.
6. Turina M, Ghignone S, Astolfi N, Silvestri A, Bonfante P, Lanfranco L. 2018. The virome of the arbuscular mycorrhizal fungusGigaspora mar-garitareveals the first report of DNA fragments corresponding to repli-cating non-retroviral RNA viruses in fungi. Environ Microbiolhttps://doi .org/10.1111/1462-2920.14060.
7. Nibert ML, Vong M, Fugate KK, Debat HJ. 2018. Evidence for contempo-rary plant mitoviruses. Virology 518:14 –24. https://doi.org/10.1016/j .virol.2018.02.005.
8. Bruenn JA, Warner BE, Yerramsetty P. 2015. Widespread narnavirus se-quences in plant genomes. Peer J 3:e876.https://doi.org/10.7717/peerj.876. 9. Xu Z, Wu S, Liu L, Cheng J, Fu Y, Jiang D, Xie J. 2015. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopatho-genic fungusSclerotinia sclerotiorum. Virus Res 197:127–136.https://doi .org/10.1016/j.virusres.2014.12.023.
10. Hong YG, Cole TE, Brasier CM, Buck KW. 1998. Evolutionary relationships among putative RNA-dependent RNA polymerases encoded by a mito-chondrial virus-like RNA in the Dutch elm disease fungus,Ophiostoma novo-ulmi, by other viruses and virus-like RNAs and by the Arabidopsis mitochondrial genome. Virology 246:158 –169.https://doi.org/10.1006/ viro.1998.9178.
11. Lakshman DK, Jian JH, Tavantzis SM. 1998. A double-stranded RNA element from a hypovirulent strain ofRhizoctonia solanioccurs in DNA form and is genetically related to the pentafunctional AROM protein of the shikimate pathway. Proc Natl Acad Sci U S A 95:6425– 6429.https:// doi.org/10.1073/pnas.95.11.6425.
12. Chiba S, Kondo H, Tani A, Saisho D, Sakamoto W, Kanematsu S, Suzuki N. 2011. Widespread endogenization of genome sequences of
non-retroviral RNA viruses into plant genomes. PLoS Pathog 7:e1002146. https://doi.org/10.1371/journal.ppat.1002146.
13. Liu H, Fu Y, Jiang D, Li G, Xie J, Cheng J, Peng Y, Ghabrial SA, Yi X. 2010. Widespread horizontal gene transfer from double-stranded RNA viruses to eukaryotic nuclear genomes. J Virol 84:11876 –11887.https://doi.org/ 10.1128/JVI.00955-10.
14. Horie M, Honda T, Suzuki Y, Kobayashi Y, Daito T, Oshida T, Ikuta K, Jern P, Gojobori T, Coffin JM, Tomonaga K. 2010. Endogenous non-retroviral RNA virus elements in mammalian genomes. Nature 463:84 – 87.https:// doi.org/10.1038/nature08695.
15. Mu F, Xie J, Cheng S, You MP, Barbetti MJ, Jia J, Wang Q, Cheng J, Fu Y, Chen T, Jiang D. 2017. Virome characterization of a collection of S. sclerotiorumfrom Australia. Front Microbiol 8:2540.https://doi.org/10 .3389/fmicb.2017.02540.
16. Khalifa ME, Pearson MN. 2013. Molecular characterization of three mi-toviruses co-infecting a hypovirulent isolate ofSclerotinia sclerotiorum fungus. Virology 441:22–30.https://doi.org/10.1016/j.virol.2013.03.002. 17. Hillman BI, Annisa A, Suzuki N. 2018. Viruses of plant-interacting fungi.
Adv Virus Res 100:99 –116.https://doi.org/10.1016/bs.aivir.2017.10.003. 18. Turina M, Hillman BI, Izadpanah K, Rastgou M, Rosa C, ICTV Report
Consortium. 2017. ICTV virus taxonomy profile:Ourmiavirus. J Gen Virol 98:129 –130.https://doi.org/10.1099/jgv.0.000725.
19. Hrabakova L, Koloniuk I, Petrzik K. 2017. Phomopsis longicolla RNA virus 1—novel virus at the edge of myco- and plant viruses. Virology 506: 14 –18.https://doi.org/10.1016/j.virol.2017.03.003.
20. Wu MD, Zhang L, Li GQ, Jiang DH, Hou MS, Huang HC. 2007. Hypoviru-lence and double-stranded RNA in Botrytis cinerea. Phytopathology 97:1590 –1599.https://doi.org/10.1094/PHYTO-97-12-1590.
21. Wu M, Zhang L, Li G, Jiang D, Ghabrial SA. 2010. Genome characteriza-tion of a debilitacharacteriza-tion-associated mitovirus infecting the phytopatho-genic fungusBotrytis cinerea. Virology 406:117–126.https://doi.org/10 .1016/j.virol.2010.07.010.
22. Deng F, Xu R, Boland GJ. 2003. Hypovirulence-associated double-stranded RNA fromSclerotinia homoeocarpaIs conspecific with Ophios-toma novo-ulmi mitovirus 3a-Ld. Phytopathology 93:1407–1414.https:// doi.org/10.1094/PHYTO.2003.93.11.1407.
23. Nibert ML. 2017. Mitovirus UGA(Trp) codon usage parallels that of host mitochondria. Virology 507:96 –100.https://doi.org/10.1016/j.virol.2017 .04.010.
24. Sun Q, Choi GH, Nuss DL. 2009. A single Argonaute gene is required for induction of RNA silencing antiviral defense and promotes viral RNA recombination. Proc Natl Acad Sci U S A 106:17927–17932.https://doi .org/10.1073/pnas.0907552106.
25. Segers GC, Zhang X, Deng F, Sun Q, Nuss DL. 2007. Evidence that RNA silencing functions as an antiviral defense mechanism in fungi. Proc Natl Acad Sci U S A 104:12902–12906.https://doi.org/10.1073/pnas.0702500104. 26. Chiba S, Lin YH, Kondo H, Kanematsu S, Suzuki N. 2013. A novel victorivirus from a phytopathogenic fungus,Rosellinia necatrixis
on November 6, 2019 by guest
http://jvi.asm.org/
tious as particles and targeted by RNA silencing. J Virol 87:6727– 6738. https://doi.org/10.1128/JVI.00557-13.
27. Salaipeth L, Chiba S, Eusebio-Cope A, Kanematsu S, Suzuki N. 2014. Biological properties and expression strategy of Rosellinia necatrix megabirnavirus 1 analyzed in an experimental host,Cryphonectria para-sitica. J Gen Virol 95:740 –750.https://doi.org/10.1099/vir.0.058164-0. 28. Vainio EJ, Jurvansuu J, Streng J, Rajamaki ML, Hantula J, Valkonen JPT.
2015. Diagnosis and discovery of fungal viruses using deep sequencing of small RNAs. J Gen Virol 96:714 –725. https://doi.org/10.1099/jgv.0 .000003.
29. Donaire L, Ayllon MA. 2017. Deep sequencing of mycovirus-derived small RNAs fromBotrytisspecies. Mol Plant Pathol 18:1127–1137.https:// doi.org/10.1111/mpp.12466.
30. Munoz-Adalia EJ, Diez JJ, Fernandez MM, Hantula J, Vainio EJ. 2018. Characterization of small RNAs originating from mitoviruses infecting the conifer pathogenFusarium circinatum. Arch Virol 163:1009 –1018. https://doi.org/10.1007/s00705-018-3712-2.
31. Zhang R, Hisano S, Tani A, Kondo H, Kanematsu S, Suzuki N. 2016. A capsidless ssRNA virus hosted by an unrelated dsRNA virus. Nat Micro-biol 1:15001.https://doi.org/10.1038/nmicrobiol.2015.1.
32. Sasaki A, Onoue M, Kanematsu S, Suzaki K, Miyanishi M, Suzuki N, Nuss DL, Yoshida K. 2002. Extending chestnut blight hypovirus host range within diaporthales by biolistic delivery of viral cDNA. Mol Plant Microbe Interact 15:780 –789.https://doi.org/10.1094/MPMI.2002.15.8.780. 33. Chen B, Choi GH, Nuss DL. 1994. Attenuation of fungal virulence by
synthetic infectious hypovirus transcripts. Science 264:1762–1764. 34. Choi GH, Nuss DL. 1992. Hypovirulence of chestnut blight fungus
con-ferred by an infectious viral cDNA. Science 257:800 – 803.
35. Marzano SYL, Hobbs HA, Nelson BD, Hartman GL, Eastburn DM, McCop-pin NK, Domier LL. 2015. Transfection ofSclerotinia sclerotiorumwith in vitro transcripts of a naturally occurring interspecific recombinant of Sclerotinia sclerotiorumhypovirus 2 significantly reduces virulence of the fungus. J Virol 89:5060 –5071.https://doi.org/10.1128/JVI.03199-14. 36. Hillman BI, Supyani S, Kondo H, Suzuki N. 2004. A reovirus of the fungus
Cryphonectria parasiticathat is infectious as particles and related to the Coltivirusgenus of animal pathogens. J Virol 78:892– 898.
37. Eusebio-Cope A, Sun L, Tanaka T, Chiba S, Kasahara S, Suzuki N. 2015. The chestnut blight fungus for studies on virus/host and virus/virus interactions: from a natural to a model host. Virology 477:164 –175. https://doi.org/10.1016/j.virol.2014.09.024.
38. Polashock JJ, Bedker PJ, Hillman BI. 1997. Movement of a small mito-chondrial double-stranded RNA element of Cryphonectria parasitica: ascospore inheritance and implications for mitochondrial recombina-tion. Mol Gen Genet 256:566 –571.
39. Lee KM, Yu J, Son M, Lee YW, Kim KH. 2011. Transmission of Fusarium boothii mycovirus via protoplast fusion causes hypovirulence in other phytopathogenic fungi. PLoS One 6:e21629. https://doi.org/10.1371/ journal.pone.0021629.
40. Chiba S, Suzuki N. 2015. Highly activated RNA silencing via strong induction of dicer by one virus can interfere with the replication of an unrelated virus. Proc Natl Acad Sci U S A 112:E4911–E4918.https://doi .org/10.1073/pnas.1509151112.
41. Chiba S, Lin YH, Kondo H, Kanematsu S, Suzuki N. 2016. A novel betapartitivirus RnPV6 fromRosellinia necatrixtolerates host RNA silenc-ing but is interfered by its defective RNAs. Virus Res 219:62–72.https:// doi.org/10.1016/j.virusres.2015.10.017.
42. Andika IB, Jamal A, Kondo H, Suzuki N. 2017. SAGA complex mediates the transcriptional up-regulation of antiviral RNA silencing. Proc Natl Acad Sci U S A 114:E3499 –E3506.https://doi.org/10.1073/pnas .1701196114.
43. Zhang X, Shi D, Nuss DL. 2012. Variations in hypovirus interactions with the fungal-host RNA-silencing antiviral-defense response. J Virol 86: 12933–12939.https://doi.org/10.1128/JVI.00961-12.
44. Zhang X, Segers GC, Sun Q, Deng F, Nuss DL. 2008. Characterization of hypovirus-derived small RNAs generated in the chestnut blight fungus by an inducible DCL-2-dependent pathway. J Virol 82:2613–2619. https://doi.org/10.1128/JVI.02324-07.
45. Andika IB, Kondo H, Suzuki N. 23 January 2019. Dicer functions tran-scriptionally and post-trantran-scriptionally in a multilayer antiviral defense. Proc Natl Acad U S Ahttps://doi.org/10.1073/pnas.1812407116. 46. Kondo H, Kanematsu S, Suzuki N. 2013. Viruses of the white root rot
fungus,Rosellinia necatrix. Adv Virus Res 86:177–214.https://doi.org/10 .1016/B978-0-12-394315-6.00007-6.
47. Liu L, Xie J, Cheng J, Fu Y, Li G, Yi X, Jiang D. 2014. Fungal
negative-stranded RNA virus that is related to bornaviruses and nyaviruses. Proc Natl Acad Sci U S A 111:12205–12210. https://doi.org/10.1073/pnas .1401786111.
48. Crivelli G, Ciuffo M, Genre A, Masenga V, Turina M. 2011. Reverse genetic analysis of Ourmiaviruses reveals the nucleolar localization of the coat protein inNicotiana benthamianaand unusual requirements for virion formation. J Virol 85:5091–5104.https://doi.org/10.1128/JVI .02565-10.
49. Esteban R, Fujimura T. 2003. Launching the yeast 23S RNA Narnavirus shows 5’ and 3’ cis-acting signals for replication. Proc Natl Acad Sci U S A 100:2568 –2573.https://doi.org/10.1073/pnas.0530167100.
50. Esteban R, Vega L, Fujimura T. 2005. Launching of the yeast 20 s RNA narnavirus by expressing the genomic or antigenomic viral RNA in vivo. J Biol Chem 280:33725–33734.https://doi.org/10.1074/jbc.M506546200. 51. van Diepeningen AD, Debets AJ, Hoekstra RF. 1998. Intra- and interspe-cies virus transfer inAspergillivia protoplast fusion. Fungal Genet Biol 25:171–180.https://doi.org/10.1006/fgbi.1998.1096.
52. van Diepeningen AD, Debets AJ, Slakhorst SM, Fekete C, Hornok L, Hoekstra RF. 2000. Interspecies virus transfer via protoplast fusions betweenFusarium poaeand black Aspergillus strains. Fungal Genet Newsl 47:99 –100.https://doi.org/10.4148/1941-4765.1216.
53. Liu YC, Dynek JN, Hillman BI, Milgroom MG. 2007. Diversity of viruses in Cryphonectria parasiticaandC. nitschkeiin Japan and China, and partial characterization of a new chrysovirus species. Mycol Res 111:433– 442. https://doi.org/10.1016/j.mycres.2006.12.006.
54. Peever TL, Liu YC, Wang KR, Hillman BI, Foglia R, Milgroom MG. 1998. Incidence and diversity of double-stranded RNAs occurring in the chestnut blight fungus, Cryphonectria parasitica, in China and Japan. Phytopathology 88:811– 817.https://doi.org/10.1094/PHYTO.1998.88 .8.811.
55. Patel D, Power JB, Anthony P, Badakshi F, Pat Heslop-Harrison JS, Davey MR. 2011. Somatic hybrid plants ofNicotianaxsanderae(⫹)N. debneyi with fungal resistance toPeronospora tabacina. Ann Bot 108:809 – 819. https://doi.org/10.1093/aob/mcr197.
56. Iovene M, Savarese S, Cardi T, Frusciante L, Scotti N, Simon PW, Carputo D. 2007. Nuclear and cytoplasmic genome composition of Solanum bulbocastanum(⫹)S. tuberosumsomatic hybrids. Genome 50:443– 450. https://doi.org/10.1139/g07-024.
57. Dhir A, Dhir S, Borowski LS, Jimenez L, Teitell M, Rotig A, Crow YJ, Rice GI, Duffy D, Tamby C, Nojima T, Munnich A, Schiff M, de Almeida CR, Rehwinkel J, Dziembowski A, Szczesny RJ, Proudfoot NJ. 2018. Mito-chondrial double-stranded RNA triggers antiviral signalling in hu-mans. Nature 560:238 –242.https://doi.org/10.1038/s41586-018-0363 -0:10.1038/s41586-018-0363-0.
58. Eusebio-Cope A, Suzuki N. 2015. Mycoreovirus genome rearrangements associated with RNA silencing deficiency. Nucleic Acids Res 43: 3802–3813.https://doi.org/10.1093/nar/gkv239.
59. Yamada Y, Makimura K, Merhendi H, Ueda K, Nishiyama Y, Yamaguchi H, Osumi M. 2002. Comparison of different methods for extraction of mitochondrial DNA from human pathogenic yeasts. Jpn J Infect Dis 55:122–125.
60. Gobbi E, Firrao G, Carpanelli A, Locci R, Van Alfen NK. 2003. Mapping and characterization of polymorphism in mtDNA ofCryphonectria parasitica: evidence of the presence of an optional intron. Fungal Genet Biol 40:215–224.
61. Seguin J, Otten P, Baerlocher L, Farinelli L, Pooggin MM. 2016. MISIS-2: a bioinformatics tool for in-depth analysis of small RNAs and represen-tation of consensus master genome in viral quasispecies. J Virol Methods 233:37– 40.https://doi.org/10.1016/j.jviromet.2016.03.005.
62. Anagnostakis SL. 1984. Nuclear gene-mutations inEndothia( Cryphonec-tria)parasiticathat affect morphology and virulence. Phytopathology 74:561–565.https://doi.org/10.1094/Phyto-74-561.
63. Milgroom MG, Lipari SE, Powell WA. 1992. DNA fingerprinting and nalysis of population structure in the chestnut blight fungus, Cryphonec-tria parasitica. Genetics 131:297–306.
64. Smart CD, Yuan W, Foglia R, Nuss DL, Fulbright DW, Hillman BI. 1999. Cryphonectria hypovirus 3, a virus species in the familyHypoviridaewith a single open reading frame. Virology 265:66 –73. https://doi.org/10 .1006/viro.1999.0039.
65. Hillman BI, Tian Y, Bedker PJ, Brown MP. 1992. A North American hypovirulent isolate of the chestnut blight fungus with European isolate-related dsRNA. J Gen Virol 73:681– 686.https://doi.org/10.1099/0022 -1317-73-3-681.
66. Enebak SA, Hillman BI, Macdonald WL. 1994. A hypovirulent isolate of
Shahi et al. Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
Cryphonectria parasiticawith multiple, genetically unique dsRNA seg-ments. MPMI 7:590 –595.https://doi.org/10.1094/MPMI-7-0590. 67. Eusebio-Cope A, Suzuki N, Sadeghi-Garmaroodi H, Taga M. 2009.
Cyto-logical and electrophoretic karyotyping of the chestnut blight fungus Cryphonectria parasitica. Fungal Genet Biol 46:342–351.https://doi.org/ 10.1016/j.fgb.2009.01.005.
68. Suzaki K, Yoshida K, Ito T. 1997. Pathogenicity to apple branch and phloridzin degrading activity ofValsa ceratospermaisolates from some broad leaf trees including apple tree. Annu Rep Soc Plant Prot N Jpn 48:145–157.
69. Ghabrial SA, Dunn SE, Li H, Xie J, Baker TS. 2013. Viruses of
Helmint-hosporium (Cochlioblus) victoriae. Adv Virus Res 86:289 –325.https://doi .org/10.1016/B978-0-12-394315-6.00011-8.
70. Short DP, Double M, Nuss DL, Stauder CM, MacDonald W, Kasson MT. 2015. Multilocus PCR assays elucidate vegetative incompatibility gene profiles ofCryphonectria parasitica in the United States. Appl Environ Microbiol 81:5736 –5742.https://doi.org/10.1128/AEM.00926-15. 71. Choi GH, Dawe AL, Churbanov A, Smith ML, Milgroom MG, Nuss DL.
2012. Molecular characterization of vegetative incompatibility genes that restrict hypovirus transmission in the chestnut blight fungus Cryphonectria parasitica. Genetics 190:113–127. https://doi.org/10 .1534/genetics.111.133983.