Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Temperature-Sensitive Transformation by an Abelson Virus
Mutant Encoding an Altered SH2 Domain
CELINE A. MAINVILLE,1,2KALINDI PARMAR,1† INDIRA UNNIKRISHNAN,1LI GONG,3 GLEN D. RAFFEL,3,4‡ANDNAOMI ROSENBERG1,2,3,4*
Departments of Pathology1and Molecular Biology and Microbiology,3Medical Scientist Training Program,4and
Graduate Program in Immunology,2Tufts University School of Medicine, Boston, Massachusetts 02111
Received 17 August 2000/Accepted 20 November 2000
Abelson murine leukemia virus (Ab-MLV) encodes the v-Abl protein tyrosine kinase and induces transfor-mation of immortalized fibroblast lines and pre-B cells. Temperature-sensitive mutations affecting the kinase domain of the protein have demonstrated that the kinase activity is absolutely required for transformation. Despite this requirement, mutations affecting other regions of v-Abl modulate transformation activity. The SH2 domain and the highly conserved FLVRES motif within it form a phosphotyrosine-binding pocket that is required for interactions between the kinase and cellular substrates. To understand the impact of SH2 alterations on Ab-MLV-mediated transformation, we studied the Ab-MLV mutant P120/R273K. This mutant encodes a v-Abl protein in which theB5 arginine at the base of the phosphotyrosine-binding pocket has been replaced by a lysine. Unexpectedly, infection of NIH 3T3 or pre-B cells with P120/R273K revealed a temper-ature-dependent transformation phenotype. At 34°C, P120/R273K transformed about 10-fold fewer cells than wild-type virus of equivalent titer; at 39.5°C, 300-fold fewer NIH 3T3 cells were transformed and pre-B cells were refractory to transformation. Temperature-dependent transformation was accompanied by decreased phosphorylation of Shc, a protein that interacts with the v-Abl SH2 and links the protein to Ras, and decreased induction of c-Myc expression. These data suggest that alteration of the FLVRES pocket affects the ability of v-Abl to interact with at least some of its substrates in a temperature-dependent fashion and identify a novel type of temperature-sensitive Abelson virus.
Expression of the protein tyrosine kinase encoded by the v-abloncogene of Abelson murine leukemia virus (Ab-MLV) induces transformation of pre-B cells and immortalized fibro-blast lines in vitro and causes pre-B-cell lymphoma in mice (37). Although the tyrosine kinase activity of the v-Abl protein is absolutely required for transformation, the SH2 domain, a region involved in phosphotyrosine-dependent interactions (11, 48), also plays an important role in the process. This domain contains a phosphotyrosine-binding pocket character-ized by the amino acids FLVRES; replacing ArgB5, located at the base of the pocket, with a Lys or other residues drasti-cally affects the ability of activated c-Abl proteins and Bcr/Abl proteins to transform immortalized fibroblast lines (1, 28). These substitutions drastically reduce binding of tyrosine-phosphorylated peptides to SH2 domains; those affecting other pocket residues have more modest effects on binding and vari-able effects on transformation mediated by Abl or by Src, which has a closely related SH2 domain (3–5, 13, 28, 44, 47). Although the FLVRES motif and ArgB5 play a dominant role in SH2-mediated phosphopeptide binding, they do not fully account for the specificity of SH2 domain substrate inter-action. Residues within the SH2 domain that contact residues C terminal to the phosphotyrosine on the target protein are
one way in which specificity of substrate interaction may be controlled (26, 43). However, analyses of interactions between Src SH2 domain mutants and peptides deviating from its con-sensus pYEEI peptide binding sequence have suggested that specificity reflects more complex interactions (5). Parameters such as intracellular location (which likely affects the proximity of the kinase and particular substrates), the ability to assemble signaling complexes, and interactions with other domains of the kinase are likely to influence substrate selection and play a role in SH2 domain function.
Identifying substrates that interact with SH2 domains and probing the way these interactions contribute to transforma-tion provide one approach to understanding how SH2-medi-ated signaling specificity contributes to host-virus interaction. For the v-Abl protein, the Shc adapter molecule is one such substrate (31, 35). Shc can complex with Grb2 and the G-protein exchange factor, mSos, facilitating the activation of Ras (14, 27). The last event is required for Abl-mediated trans-formation (39). Activation of Ras stimulates multiple down-stream effector proteins, including c-Myc (21–23), another pro-tein that is critical for Abl-mediated transformation (38). Consistent with the idea that signals requiring an intact SH2 domain pass through Ras and lead to c-Myc (52), transforma-tion of RAT-1 cells by Bcr/Abl proteins containing an altered FLVRES motif is complemented by expression of c-Myc (1, 25).
To understand how the v-Abl SH2 domain contributes to transformation and downstream signaling, we examined the biological properties of the P120/R273K mutant. This mutant encodes a v-Abl protein in which a Lys is substituted for Arg
B5. Unexpectedly, P120/R273K retained significant transfor-* Corresponding author. Mailing address: SC315, Tufts University
School of Medicine, 136 Harrison Ave., Boston, MA 02111. Phone: (617) 636-2143. Fax: (617) 636-0337. E-mail: [email protected] .edu.
† Present address: Department of Radiation Oncology, Brigham and Women’s Hospital, Boston, MA 02215.
‡ Present address: Department of Hematology and Oncology, Beth Israel Deaconess Medical Center, Boston, MA 02215.
1816
on November 9, 2019 by guest
http://jvi.asm.org/
mation potential when assayed at 34°C but was compromised for transformation at 39.5°C. These defects were correlated to deficiencies in interactions with Shc and decreased stimulation of c-Myc. These data reveal a novel type of Ab-MLV temper-ature-sensitive (ts) mutant and a previously unappreciated ef-fect of temperature on v-Abl proteins with an altered SH2 domain.
MATERIALS AND METHODS
Cells and viruses.NIH 3T3 cells were grown in Dulbecco’s modified Eagle’s medium supplemented to contain 10% calf serum (Sigma); Ab-MLV-trans-formed NIH 3T3 cells and 293T cells (15) were grown in Dulbecco’s modified Eagle’s medium supplemented to contain 10% fetal calf serum (Intergen). Ab-MLV-transformed pre-B-cell lines were grown in RPMI 1640 medium supple-mented to contain 20% fetal calf serum and 50M 2-mercaptoethanol (Sigma). ThetsAb-MuLV-transformed pre-B-cell line 7C411 was derived by infecting bone marrow cells with thetsAb-MuLV strain P70/H590 (17) and maintained at 34°C, the permissive temperature for thetsAb-MuLV mutants; the nonpermis-sive temperature used was 39.5°C. Retroviral stocks were prepared using tran-sient transfection of 293T cells as described elsewhere (49). The pSR␣-MSVtkneo vector (30) or the pMIG vector (19, 46) was used to express the different Ab-MLV strains. In the pMIG vector, green fluorescent protein (GFP) expres-sion is mediated by an internal ribosome entry site (IRES) sequence andgag /v-ablsequences are cloned upstream of the IRES sequence. Infectivity was eval-uated by Western blot analysis of the levels of v-Abl protein in lysates of NIH 3T3 cells that had been infected for 48 to 72 h with the Ab-MLV stocks prepared with the pSR␣vector (33) or by using fluorescence-activated cell sorting (FACS) to score the frequency of GFP-positive NIH 3T3 cells 24 h after infection with Ab-MLV stocks prepared using the pMIG vector. The titers of virus were also determined using the NIH 3T3 cell transformation assay (40). Bone marrow transformation assays were done as described previously (36). Macroscopic pre-B-cell colonies were counted 10 to 12 days later. For some assays, the infected cells were plated directly into RPMI 1640 medium supplemented to contain 20% fetal calf serum and 50M 2-mercaptoethanol. Cultures were scored as trans-formed when the number of nonadherent pre-B cells exceeded 2⫻106per ml of culture medium (1, 29). To obtain derivatives of 7C411 cells expressing wild-type or Ab-MLV mutants, 106cells were infected in the presence of 4g of Polybrene per ml for 4 h and plated in standard growth medium; 24 h later, 1 mg of G418 per ml was added to the medium to select for cells expressing the superinfecting virus, and the cells were plated in 96-well plates. Individual clones of G418-resistant cells were isolated 7 to 14 days later. Cell cycle parameters were monitored by analyzing propidium iodide-stained cells using a FACScan and Modfit LT software (8).
Construction of viral strains.pSR␣-P120 and the kinase negative mutant P120/D484N have been described elsewhere (35, 49). The P120/R273K mutant was created by PCR using pUC120 as a template (18) and one primer which changed the codon for Arg-273 to a Lys codon. The fragment containing the mutation was subcloned and then shuttled into pSR␣-P120 as aBstEII-DraIII (bases 725 to 2111 of the P120 genome) fragment. Ab-MLV P120 and P120/ R273K were introduced into pMIG by shuttling the viral sequences contained on anEcoRI fragment from the pSR␣vectors into theEcoRI cloning site of pMIG (19, 46). The presence of all mutations and the integrity of all sequences derived by PCR amplification were examined in the DNA Facility, Department of Phys-iology, Tufts University School of Medicine.
Protein analysis.Cells were treated with lysis buffer (10 mM Tris [pH 7.4], 1% sodium dodecyl sulfate [SDS], 1 mM sodium orthovanadate, 1 mM phenylmeth-ylsulfonyl fluoride [PMSF]) as described previously (9, 32). Protein was quanti-tated using a bicinchoninic acid protein assay kit (Pierce), and 50g of each sample was fractionated through SDS-polyacrylamide gels. The proteins were transferred to a polyvinylidene difluoride membrane (U.S. Biochemicals), and the blots were treated according to the Western-Light kit protocol (Tropix) using alkaline phosphatase-conjugated secondary antibodies and the CSPD substrate (Tropix). For immunoprecipitation with anti-p62 Dok antibodies, cells were resuspended in ice-cold TNN lysis buffer (150 mM NaCl, 50 mM Tris [pH 8], 1% NP-40, 10g of leupeptin per ml, 1 mM orthovanadate, and 1 mM PMSF); for immunoprecipitation with anti-Shc antibodies, cells were resuspended in radio-immunoprecipitation assay buffer (10 mM sodium phosphate [pH 7], 150 mM NaCl, 0.1% SDS, 50 mM NaF, 1% NP-40, 0.5% sodium deoxycholate, 2 mM EDTA, 1 mM sodium orthovanadate, 1 mM PMSF, and 10g of leupeptin per ml). The lysates were clarified and equivalent amounts of protein were immu-noprecipitated. The immune complexes were recovered with Sepharose CL-4B
beads and heated at 95°C for 5 min. The proteins were resolved on SDS-polyacrylamide gels and analyzed by immunoblotting. In some experiments, fusion proteins in which SH2 domains fused to glutathioneS-transferase (GST) were used to precipitate proteins. The GST-SH2 proteins were produced by using the pGEX-3X vector (Pharmacia) andEscherichia coliJM109 cells and reacted with cell lysates as described previously (35). Immunofluorescent stain-ing for v-Abl was carried out usstain-ing cells fixed with 1% formaldehyde and the v-Abl monoclonal antibody 24-21 (41). Proteins were analyzed using anti-Gag/v-Abl (H548) (10), anti-Shc (S14630; Transduction Laboratories), anti-Grb2 (G16720; Transduction Laboratories), anti-p62 Dok (MMS239P [Babco] or SC6929 [Santa Cruz Biotechnology]), anti-myc (06-340; Upstate Biotechnology), antiphosphotyrosine (05-321; Upstate Biotechnology), anti-p42/p44 MAPK (9100; New England Biolabs), and anti-GST (SC-138; Santa Cruz Biotechnol-ogy) antibodies. Control antibodies included rabbit gamma globulin, anti--2-6 fructosan (UPC-10) (2), and anti-murine immunoglobulin (chain).
RESULTS
Alteration of the v-Abl SH2 domain reduces NIH 3T3 cell transformation in a temperature-dependent fashion. To de-termine the effects of an altered v-Abl SH2 domain on Ab-MLV transformation, NIH 3T3 cells were infected with the Ab-MLV P120/R273K mutant and monitored for transforma-tion. Ab-MLV P120- or mock-infected cells served as controls. Cells were examined at the standard 37°C used by others for analogous mutants in other forms of Abl and at 34 and 39.5°C, temperatures commonly used with mammalian cells and ts
[image:2.612.339.522.70.297.2]mutants. When cells infected and maintained at 34°C were compared to sister cultures infected at 34°C and then shifted to 39.5°C for 48 h, similar levels of v-Abl proteins were recovered (Fig. 1). All of the extracts contained elevated levels of phos-photyrosine compared to those of mock-infected cells (Fig. 1). FIG. 1. Cells expressing P120/R273K contain low levels of phos-photyrosine at high temperature. NIH 3T3 cells were infected with Ab-MLV P120 or P120/R273K or were mock infected at 34°C (L); some samples were then shifted to 39.5°C (H), and all were lysed 48 h later. Equivalent amounts of total cellular protein were analyzed by using Western blotting with antiphosphotyrosine antibodies or the anti-Gag/v-Abl monoclonal antibody H548 (10). Molecular weight standards (in thousands) are indicated on the left, and arrows highlight bands which display temperature-dependent differences in phosphoty-rosine. The asterisk indicates the v-Abl protein.
on November 9, 2019 by guest
http://jvi.asm.org/
Although these levels were reduced in cells infected with P120/ R273K and maintained at 34°C, the reductions were more obvious when the cells were maintained at 39.5°C. Effects on some proteins (Fig. 1) were more striking, suggesting that temperature may affect the ability of certain substrates to in-teract with P120/R273K. In addition, tyrosine phosphorylation of P120/R273K itself was also reduced.
Although cells infected with P120/R273K displayed reduced levels of cellular phosphotyrosine at 39.5°C, typical rounded transformed cells could be found in the cultures. However, the frequency of these cells appeared to be reduced compared to that in cultures analyzed at 34°C. To examine this issue more fully, the transforming titer of the viruses was measured in a standard NIH 3T3 cell focus assay (40) at all three tempera-tures. As expected (18), the titers of P120 stocks were similar at all temperatures (Table 1). In contrast, the titer of the P120/R273K stock was reduced only 10-fold compared to that of the wild-type virus at 34°C. This difference increased to about 100-fold when assayed at 37°C, and the titer was 300-fold lower at 39.5°C. Several investigators have observed dimin-ished transformation frequencies with other transformingabl
genes containing analogous mutations (1, 28) using transfor-mation assays carried out at 37°C. However, at least for v-Abl, the effects of the R273K substitution are magnified at this temperature. In addition, given the dramatic effects of the R273K substitution on interaction with phosphotyrosine-con-taining peptides (28), the small effect of the substitution on transformation at 34°C is surprising and suggests that interac-tions between isolated SH2 domains and peptides may not fully mimic the situation in vivo.
As noted above, some transformed cells appeared in P120/ R273K-infected cultures maintained at 39.5°C. In addition, fully transformed cultures of NIH 3T3 cells derived with P120/ R273K at 34°C retained a transformed morphology when shifted to 39.5°C and were indistinguishable from cells trans-formed by the wild-type P120 strain (data not shown). These data reveal that thetsphenotype of these cells is considerably weaker than that observed with other ts Ab-MLV mutants which contain substitutions within the kinase domain, in which case almost all the cells revert to a normal morphology at the nonpermissive temperature (18). Alternatively, during the transformation process at 34°C additional genetic changes
which facilitate maintenance of the transformed phenotype at the nonpermissive temperature may accumulate.
Reduced binding of Shc and p62Dok to an SH2 domain with the R273K substitution. Even though substitutions affecting the conserved Arg in the FLVRES motif dramatically reduce the ability of SH2 domains to interact with tyrosine-phospho-rylated peptides (3–5, 28), P120/R273K is only mildly compro-mised for NIH 3T3 cell transformation at 34°C. These data could suggest that SH2 domain interactions in vivo are more complex than can be detected by in vitro binding assays. To explore this hypothesis, the ability of the R273K SH2 domain to bind two v-Abl substrates, Shc and p62Dok, was tested by using GST-SH2 fusion proteins. Shc transmits stimulatory sig-nals via the Grb2-Sos complex to Ras after tyrosine phosphor-ylation by v-Abl (14, 27, 31, 35); tyrosine-phosphorylated p62Dok is believed to stimulate Ras by binding the Ras sup-pressor p120RasGap (7, 16, 51).
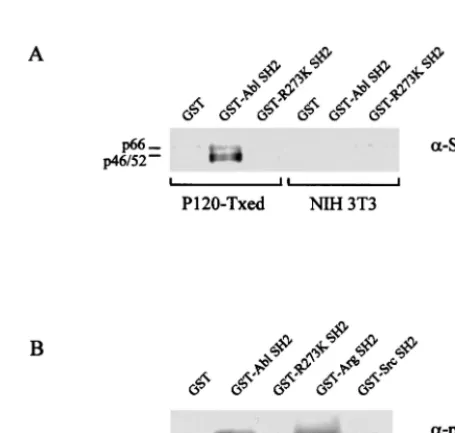
[image:3.612.52.294.92.155.2]As expected (35), Shc recovered from cells transformed with Ab-MLV P120, but not from uninfected NIH 3T3 cells, could be precipitated by the v-Abl SH2 domain (Fig. 2A). In con-trast, Shc was precipitated poorly by the GST-R273K/SH2 protein, with faint interaction being observed on prolonged exposure of the autoradiogram (not shown). Because binding of p62Dok to the v-Abl SH2 domain has not been reported previously, the GST fusion proteins used to study Shc and others expressing the SH2 domains of v-Src and the Abl-re-lated Arg protein (24) were used to precipitate p62Dok. All of these proteins interacted with p62Dok present in lysates of Ab-MLV P120-transformed NIH 3T3 cells or uninfected NIH
[image:3.612.316.544.417.633.2]FIG. 2. Shc and p62Dok bind poorly to an SH2 domain with the R273K substitution. (A) Lysates of uninfected NIH 3T3 cells or cells transformed with Ab-MLV P120 were incubated with GST alone, GST-SH2, or GST-R273K SH2. The proteins were recovered and analyzed by using Western blotting with an anti-Shc antibody. (B) Lysates of NIH 3T3 cells transformed by Ab-MLV P120 were incu-bated with the indicated GST-SH2 domain fusion proteins or GST protein as a control. The recovered proteins were analyzed by using Western blotting with an anti-p62Dok antibody. All samples contained equivalent amounts of GST proteins (data not shown).
TABLE 1. Mutations affecting the FLVRES motif in the SH2 domain reduce NIH 3T3 cell transformation at 39.5°Ca
Virus Focus-forming units (10 5) at:
37°C 34°C 39.5°C
P120 (WTb) 1.0 1.0 1.0
P120/R273K 0.01 0.1 0.003
None ⬍0.00001 ⬍0.00001 ⬍0.00001
aThe NIH 3T3 cell focus assay was used to determine the titers of the viruses (40). The same virus stocks had their titers determined at 34 and 39.5°C; a different set of virus stocks were used for the 37°C experiment. Within each set, the stocks used infected equivalent numbers of NIH 3T3 cells as judged by Western blot analysis similar to that shown in Fig. 1 (37°C) or by FACS analysis of GFP-positive cells (34 and 39.5°C). The data shown are representative of at least three independent experiments. Values with a “⬍” sign indicate that no transformants were observed and reflect the minimum number of foci that could have been detected.
bWT, wild type.
on November 9, 2019 by guest
http://jvi.asm.org/
3T3 cells (Fig. 2B and data not shown). However, binding by the GST fusion expressing the R273K substitution was de-creased approximately 20-fold and was lower than that ob-served for the Src SH2 domain. Thus, SH2 domain interaction with two v-Abl substrates is diminished in the presence of the FLVRES motif substitution, despite the ability of P120/R273K to induce relatively robust transformation at 34°C.
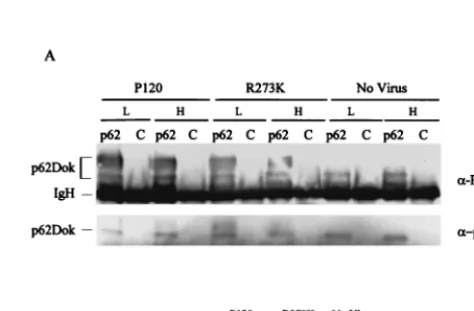
Temperature-dependent transformation correlates to de-creased signals to the Ras pathway intermediates. The dis-crepancy between the in vitro binding data and the transform-ing potential of P120/R273K at 34°C suggests that the bindtransform-ing experiments may not fully reveal the signaling potential of the altered v-Abl protein. To explore this issue in vivo, we exam-ined tyrosine phosphorylation of Shc and p62Dok in NIH 3T3 cells infected with the mutant and maintained at 34 and 39.5°C. Immunoprecipitation and Western analysis revealed that NIH 3T3 cells infected 48 h previously and incubated at 34 and 39.5°C expressed similar levels of Shc, p62Dok, and v-Abl (Fig. 3 and 4). However, tyrosine phosphorylation of these two pro-teins was affected differently in the presence of P120/R273K at the two incubation temperatures. Tyrosine phosphorylation of Shc was decreased about fourfold in cells infected with P120/ R273K at 39.5°C compared to that in cells expressing P120 or P120/R273K at 34°C (Fig. 3); association with Grb2 was more strongly affected and was reduced to levels similar to those seen in mock-infected cells. In contrast, tyrosine phosphoryla-tion of p62Dok was slightly decreased in cells expressing P120/ R273K at both incubation temperatures (Fig. 4). These results suggest that signals for activation of Shc and subsequent asso-ciation of Grb2 are altered by temperature-dependent changes affecting the P120/R273K protein. Because decreases in Shc phosphorylation appear more modest than effects on Grb2
association, assessing tyrosine phosphorylation levels may not fully reflect the ability of Shc to mediate downstream signaling. However, effects on p62Dok were more modest, suggesting that alteration of the FLVRES motif has various effects on different v-Abl substrates and that some are more drastically affected by temperature than others. These data also confirm that in vitro binding does not reveal the full complexity of interactions influenced by the SH2 domain in vivo.
[image:4.612.60.289.72.229.2]Pre-B-cell transformation by P120/R273K is temperature dependent.Some Ab-MLV mutants display different transfor-mation phenotypes in NIH 3T3 cells and pre-B cells (37). To determine if pre-B-cell transformation by the P120/R273K mu-tant is also temperature dependent, primary bone marrow cells were infected, plated in agar or liquid medium (1, 29, 36) at 34, 37, and 39.5°C, and monitored for transformation. Liquid cul-tures were scored as transformed when the cell density ex-ceeded approximately 2⫻106transformed cells per ml, a time at which the cells could be subcultured; for agar assays, the frequency of macroscopic colonies of primary transformants was determined 10 days postinfection. Bone marrow cells in-fected with Ab-MLV P120 became transformed at all temper-atures, and the P120/R273K mutant transformed fewer cells under all conditions (Table 2). Between five- and sevenfold fewer colonies were observed in agar at both 34 and 37°C. However, transformation frequencies were reduced approxi-mately 100-fold at 39.5°C, with one to two very small pre-B-cell colonies being observed; these cell populations could not be expanded. Consistent with the diminished transformation po-tential of the P120/R273K mutant, infected liquid cultures maintained at 34 and 37°C became transformed later than those infected with the wild-type virus. Although some pre-B cells appeared to proliferate in several cultures infected with P120/R273K and maintained at 39.5°C, the population of these
[image:4.612.314.551.461.616.2]FIG. 4. Tyrosine phosphorylation of p62Dok is not temperature dependent. NIH 3T3 cells were infected with Ab-MLV P120 or P120/ R273K or were mock infected and incubated at 34°C (L) or 39.5°C (H), and lysates were prepared 48 h later. (A) A portion of the lysate was immunoprecipitated with anti-p62Dok or the UPC-10 control antibody (2) and analyzed by using antiphosphotyrosine and p62Dok anti-bodies. Lanes labeled “p62” contain samples immunoprecipitated with anti-p62Dok antibody; lanes labeled “C” contain samples immunopre-cipitated with UPC-10 control antibody. The antibodies used to probe the blots are listed to the right. (B) The remaining lysate was analyzed by using Western blotting with the Gag/v-Abl monoclonal anti-body H548 (10).
FIG. 3. Tyrosine phosphorylation of Shc and Shc-Grb2 complexes are reduced in cells expressing P120/R273K at 39.5°C. NIH 3T3 cells were infected with Ab-MLV P120 or P120/R273K or were mock in-fected and incubated at 34°C (L) or 39.5°C (H). (A) The cells were lysed 48 h later, and a portion of the lysate was immunoprecipitated with anti-Shc antibody or rabbit gamma globulin as a control and analyzed by using Western blotting and antiphosphotyrosine, anti-Grb2, and anti-Shc antibodies. Lanes labeled “Shc” contain samples immunoprecipitated with anti-Shc antibody; lanes labeled “C” contain samples immunoprecipitated with rabbit gamma globulin. The anti-bodies used to probe the blots are listed to the right. (B) The remain-ing lysate was analyzed by usremain-ing Western blottremain-ing with the anti-Gag/ v-Abl monoclonal antibody H548 (10).
on November 9, 2019 by guest
http://jvi.asm.org/
cells could not be expanded. These data demonstrate that the temperature-dependent transformation response found in NIH 3T3 cells extends to pre-B cells and represents a general feature of transformation responses to P120/R273K.
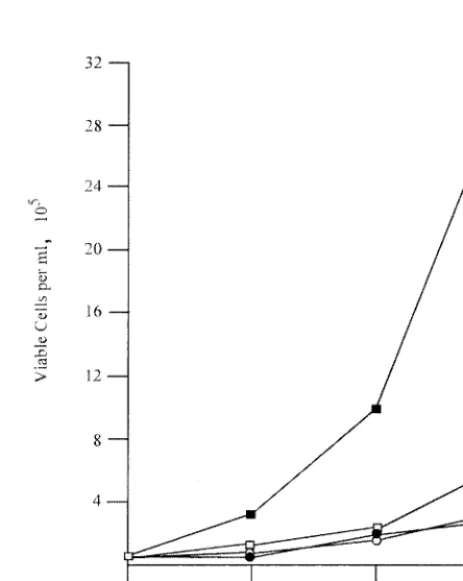
Pre-B cells transformed by P120/R273K grow poorly at el-evated temperature.NIH 3T3 cells transformed at 34°C main-tained a transformed morphology at 39.5°C. To determine if pre-B cells transformed by P120/R273K at 34°C could grow at 39.5°C, these cells and controls transformed by wild-type P120 were shifted to the high temperature, and growth was moni-tored by counting viable cells using phase microscopy. Consis-tent with previous results (8), cells transformed by P120 grew more rapidly at 39.5 than at 34°C, while cells transformed with P120/R273K grew poorly at both temperatures (Fig. 5).
Unlike transformation of NIH 3T3 cells, Ab-MLV transfor-mation of pre-B cells is a multistep process during which cells adapt to culture and often acquire mutations affecting the p53 pathway (34, 45). These changes could affect the ability of cells transformed by P120/R273K to grow at 39.5°C. To test the ability of P120/R273K to stimulate pre-B-cell growth in the absence of this selection, the ability of P120/R273K to rescue the growth of pre-B cells transformed by the P70/H590 ts
mutant was examined (17). This mutant encodes a v-Abl pro-tein containing a point mutation within the kinase domain; pre-B cells transformed by this virus undergo G1 arrest and rapid apoptosis at the nonpermissive temperature. However, when such cells are superinfected with wild-type Ab-MLV, growth is restored (17).
To monitor the effect of P120/R273K in this setting, the 7C411tspre-B-cell transformant was infected with wild-type virus or P120/R273K in vectors that also contain a Neorgene. Clones expressing the superinfecting viruses were selected us-ing G418; immunofluorescent stainus-ing with an antibody that reacts with COOH-terminal determinants present in P120 and P120/R273K, but not in the P70 protein (41), revealed that
⬎90% of the cells in the population expressed the protein encoded by the superinfecting virus. As expected (8), when the cells were shifted to the nonpermissive temperature, the pa-rental cell line underwent G1 arrest and apoptosis, and the cells expressing P120 continued to grow and displayed a cell cycle profile similar to that found when cells were incubated at 34°C (Table 3). Cells expressing P120/R273K displayed an
[image:5.612.53.554.84.169.2]intermediate phenotype characterized by G1arrest and dimin-ished apoptosis. These data demonstrate that P120/R273K fails to transmit adequate signals to sustain growth of pre-B cells at 39.5°C.
TABLE 2. Mutations affecting the SH2 domain of v-Abl reduce pre-B-cell transformationa
Virus Mean no. of colonies⫾SEM on day 10 at b:
No. of transformed dishes/total dishesc
Day 10 at: Day 21 at:
37°C 34°C 39.5°C 37°C 34°C 39.5°C 37°C 34°C 39.5°C
P120 WTd 37⫾2 49⫾6 38⫾5 4/4 4/4 4/4 4/4 4/4 4/4
P120/R273K 5⫾1 10⫾2 0.1⫾0.1 2/4 4/4 0/4 3/4 4/4 0/4
None ⬍0.5⫾0.5 NTe NT 0/4 0/4 0/4 0/4 0/4 0/4
aBone marrow cells were infected with virus stocks of matched titer and monitored for transformation. The data shown are representative of two independent experiments.
bInfected cells were plated in agar immediately after virus adsorption (36); macroscopic colonies of transformed pre-B cells were counted 10 days later. The values given represent colonies per 106nucleated bone marrow cells; values with a “⬍” sign indicate that no colonies were observed and reflect the minimum number that could have been detected.
cInfected cells were plated in liquid in 35-mm tissue culture plates; cultures were assessed by microscopic observation at days 10 and 21 and scored as transformed when the cultures became filled with transformed pre-B cells which could be readily subcultured (1, 29, 36).
dWT, wild type. eNT, not tested.
FIG. 5. Pre-B cells transformed by P120/R273K grow poorly at 39.5°C. Bone marrow cells were transformed by Ab-MLV P120 (squares) or P120/R273K (circles) at 34°C, and primary transformants were expanded and established. Cells were counted and seeded in 35-mm dishes at a density of 105cells per ml and incubated at 34°C (open symbols) or 39.5°C (filled symbols). Duplicate cultures were counted, and viable cells were enumerated using phase microscopy daily for 3 days. The experiments shown are representative of at least three experiments in which two or more cell lines transformed with each virus were analyzed.
on November 9, 2019 by guest
http://jvi.asm.org/
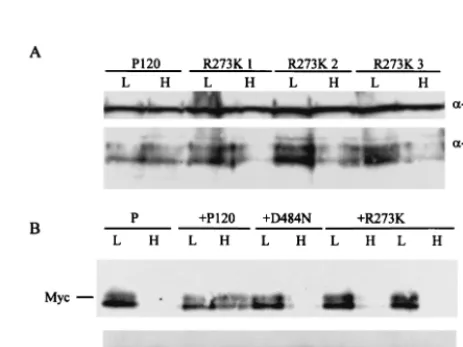
[image:5.612.316.548.345.636.2]Expression of c-Myc is decreased at the nonpermissive tem-perature.Expression of c-Myc is required for Abl-mediated transformation (38). Earlier studies, conducted at 37°C, have documented that the R273K substitution reduces the ability of v-Abl to activate expression from the E2F site in the c-myc
promoter (50, 52). To determine if the temperature-dependent effects of P120/R273K affect the level of c-Myc protein expres-sion, 7C411 cells that had been superinfected with either P120 or P120/R273K were maintained at 34°C or shifted to 39.5°C for 20 h, and lysates were prepared. Cells superinfected with the kinase-negative mutant P120/D484N (35) and parental 7C411 cells were used as controls. In a parallel experiment, pre-B cells transformed by P120 and P120/R273K at 34°C were examined at both temperatures. Western analyses revealed that c-Myc expression was diminished at 39.5°C in all of the cells expressing P120/R273K but not in those expressing P120 (Fig. 6). These data reveal that the R273K substitution affects expression of c-Myc, a critical downstream target of v-Abl (38), in a temperature-dependent fashion.
DISCUSSION
These experiments identified the sequences encoding the v-Abl SH2 domain as a novel and previously unrecognized target for Ab-MLV ts mutations. The P120/R273K mutant displays a temperature-dependent transformation phenotype in both NIH 3T3 and pre-B cells. Earlier work (28) had sug-gested a direct correlation between the poor transformation potential of Abl proteins containing altered SH2 domains and their ability to bind tyrosine-phosphorylated molecules. How-ever, only mild, 5- to 10-fold deficiencies are found when trans-formation assays are carried out at 34°C, a common permissive temperature used for conditional mutants expressed in mam-malian cells. This difference is enhanced somewhat at 37°C and further magnified at 39.5°C, the temperature used for otherts
Ab-MLV mutants (18). At this temperature, the effects of the mutation are particularly prominent in pre-B cells, which are refractory to transformation. These data suggest that the
R273K substitution affects the structure of the phosphoty-rosine-binding pocket and perhaps the entire SH2 domain in a temperature-dependent fashion.
Although temperature-dependent transformation is evident in both NIH 3T3 and pre-B-cell transformation assays, some transformed NIH 3T3 cells can be isolated at the nonpermis-sive temperature, and at least some cells in populations trans-formed at the permissive temperature retain a transtrans-formed morphology when shifted to 39.5°C. In addition, pre-B cells that have been fully transformed with P120/R273K at 34°C can replicate, albeit poorly, at 39.5°C. These data could indicate that genetic or epigenetic changes that occur during the trans-formation process complement transtrans-formation signals from the P120/R273K protein, allowing the emergence of formed cells at the nonpermissive temperature once the trans-formation process is complete. Analyses of the pre-B-cell transformation process have revealed that inactivation of the p53 pathway is one critical step in the process (34, 45), sug-gesting that this event and perhaps other similar events that may occur during NIH 3T3 cell transformation affect the trans-forming potential of P120/R273K. The P120/R273K mutant should allow further exploration of this possibility.
Substitutions affecting ArgB5 have been studied in both Bcr/Abl and an activated, transforming c-ablallele. In both of these instances, transformation of immortalized fibroblasts is reduced by several orders of magnitude at 37°C (1, 28). How-ever, these changes have little effect on the ability of Bcr/Abl to confer factor-independent growth on interleukin-3-dependent hematopoietic cells (12, 20, 42). Host range effects are also found in Rous sarcoma virus mutants encoding SH2 domains containing ArgB5 substitutions or with other changes in the SH2 domain (13, 44, 47). In most of these instances,
[image:6.612.53.293.90.235.2]trans-FIG. 6. Expression of c-Myc is decreased in cells expressing P120/ R273K at 39.5°C. Pre-B-cell lines transformed at 34°C by Ab-MLV P120 or P120/R273K (A) or 7C411 parental cells (P) expressing P120, P120/D484N, or P120/R273K (B) were maintained at 34°C or shifted to 39.5°C for 48 h, and lysates were prepared. The samples were analyzed via Western blotting using anti-c-Myc. The anti-Gag/v-Abl monoclonal antibody H548 (10) was used as a loading control in the top panel; anti-Erk antibody was used as a loading control in the bottom panel. In panel A, three representative pre-B-cell lines trans-formed with P120/R273K are shown; in panel B, two representative clones expressing P120/R273K are shown.
TABLE 3. Expression of P120/R273K does not stimulate G1transita
Cells Temperature(time)
% of viable cells in
phase: Total % viable cells G1 S G2/M
7C411 34°C 46 38 16 ⬎95
39.5°C (17h) 64 19 17 10
39.5°C (47h) ⬎99 ⬍1 ⬍1 ⬍5
7C411/P120 34°C 51 34 15 ⬎95
39.5°C (17h) 55 37 8 ⬎95
39.5°C (47h) 49 45 6 ⬎95
7C411/R273K 34°C 43 41 17 ⬎95
39.5°C (17h) 66 15 19 55
39.5°C (47h) 93 6 1 48
a7C411 cells, transformed with thetsstrain P70/H590 (18), and derivatives expressing P120 or P120/R273K were maintained at 34°C or shifted to 39.5°C for the indicated times. Samples were stained with propidium iodide and analyzed by flow cytometry. Viability was determined by counting trypan blue-stained cells. The results shown are representative of two independent experiments in which three independently derived clones each of 7C411/P120 and 7C411/R273K cells were analyzed.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.316.548.450.623.2]formed chick cells are fusiform, in contrast to the rounded cells induced by the wild-type virus, and immortalized rodent cells are refractory to transformation by the mutants. In two cases, large deletions removing the majority of the v-Src SH2 domain have also been shown to confer atsphenotype (6).
Deficiencies in transformation by P120/R273K are corre-lated to decreased levels of phosphotyrosine on many cellular proteins, as indicated by probing whole-cell lysates with an-tiphosphotyrosine antibodies. Although the identity of many of these proteins is not known, one of those affected is Shc, an intermediate involved in v-Abl-mediated Ras activation (31, 35), an obligate event in the transformation process (39). However, as exemplified by p62Dok, temperature-dependent decreases in tyrosine phosphorylation are not found on all v-Abl-responsive proteins, perhaps reflecting temperature-dependent specificity in SH2-mediated interactions in the presence of the R273K substitution. Alternatively, because phosphorylation was evaluated by Western blotting, tyrosine-phosphorylated p62Dok may be more stable than the modified forms of Shc and other target proteins.
Analyses of multiple SH2 domain substitution mutants sug-gest that the B5 Arg residue plays the dominant role in binding of tyrosine-phosphorylated residues (3–5, 28). Consis-tent with these data, GST-SH2 fusions containing a Lys in place of ArgB5 interact poorly with both p62Dok and Shc in vitro. Correlations of this sort have suggested that decreases in transformation observed for some SH2 domain mutants di-rectly reflect the inability of the transforming protein to inter-act with its substrates in vivo (28). Thus, it is perhaps surprising that P120/R273K retains a strong transforming potential at 34°C. However, similar substitutions affecting the v-Src SH2 domain have a limited effect on transformation, even though these clearly have large effects on peptide binding (3–5, 13, 44, 47). These data indicate that analyses of peptide interactions do not adequately reveal the potential for SH2 domains to participate in signaling when expressed in the context of the rest of the protein in an in vivo setting. Future analyses of P120/R273K may help reveal the mechanisms controlling this difference.
Study of pre-B cells transformed directly by P120/R273K at 34°C and analyses of 7C411 cells expressing P120/R273K both suggest that signals important for stimulation of growth are missing at 39.5°C. Among such signals are those which activate c-Myc, another obligate intermediate in the v-Abl transforma-tion pathway (38). The decrease in c-Myc levels observed at 39.5°C in cells expressing P120/R273K may reflect the inability of P120/R273K to stimulate transcription dependent on the E2F site in the c-Myc promoter (50, 52). Levels of c-mycRNA are decreased in cells incubated at the high temperature (our unpublished data), suggesting that low protein levels reflect changes in expression of the gene. Activation of c-Myc expres-sion requires an active Ras protein (52), and the decreased association of Shc and Grb2 at 39.5°C is likely to impact Ras. The ability of P120/R273K to stimulate c-Myc expression at 34 but not 39.5°C is also consistent with the temperature-depen-dent transformation phenotype. Thus, effects on c-Myc are likely to play a significant role in blunting the transformation response at the high temperature.
In addition to their role as docking sites for tyrosine-phos-phorylated proteins, residues within SH2 domains that contact
amino acids on the substrate molecules influence binding spec-ificity (5, 43). Consistent with this idea, many mutations that map to SH2 domains confer host range effects (13, 44, 47). However, identifying the interactions that mediate these ef-fects has been challenging. Analyses with peptides and SH2 domains with substitutions have failed to fully account for specificity (5), raising the possibility that specificity reflects the assembly of larger, multiprotein complexes which may be af-fected by the subcellular localization of the kinase. The ts
phenotype and the stability of P120/R273K at the high tem-perature suggest that further analysis of interactions between this protein and cellular molecules will help illuminate the ways in which the SH2 domain contributes to Ab-MLV trans-formation.
ACKNOWLEDGMENTS
This work represents an equal contribution by the first two authors. We are grateful to Henry Wortis and Larry Feig for supplying some of the reagents used in this work.
This work was supported by CA 24420 from the National Cancer Institute.
REFERENCES
1.Afar, D. E. H., A. Goga, J. McLaughlin, O. N. Witte, and C. L. Sawyers.1994. Differential complementation of Bcr-Abl point mutants with c-Myc. Science
264:424–426.
2.Auffray, C., J. L. Sikorav, and F. Rougeon.1981. Correlation between D region structure and antigen-binding specificity: evidences for the compari-son of closely related immunoglobulin VH sequences. Ann. Immunol.132D:
77–78.
3.Bibbins, K. B., H. Boeuf, and H. E. Varmus.1993. Binding of the Src SH2 domain to phosphopeptides is determined by residues in both the SH2 domain and the phosphopeptides. Mol. Cell. Biol.13:7278–7287. 4.Bradshaw, J. M., V. Mitaxov, and G. Waksman. 1999. Investigation of
phosphotyrosine recognition by the SH2 domain of the Src kinase. J. Mol. Biol.293:971–985.
5.Bradshaw, J. M., V. Mitaxov, and G. Waksman.2000. Mutational investiga-tion of the specificity determining region of the Src SH2 domain. J. Mol. Biol.299:521–535.
6.Bryant, D., and J. T. Parsons.1982. Site-directed mutagenesis of thesrcgene of Rous sarcoma virus: construction and characterization of a deletion mu-tant temperature sensitive for transformation. J. Virol.44:683–691. 7.Carpino, N., D. Wisniewski, A. Strife, D. Marshak, R. Kobayashi, B.
Still-man, and B. Clarkson.1997. p62(dok): a constitutively tyrosine-phosphory-lated, GAP-associated protein in chronic myelogenous leukemia progenitor cells. Cell88:197–204.
8.Chen, Y. Y., and N. Rosenberg.1992. Lymphoid cells transformed by Abel-son virus require the v-abl protein tyrosine kinase only during early G1. Proc. Natl. Acad. Sci. USA89:6683–6687.
9.Chen, Y.-Y., L. C. Wang, M. S. Huang, and N. Rosenberg.1994. An active v-abl protein tyrosine kinase blocks immunoglobulin light-chain gene rear-rangement. Genes Dev.8:688–697.
10. Chesebro, B., K. Wehrly, M. Cloyd, W. Britt, J. Portis, J. Collins, and J. Nishio.1981. Characterization of mouse monoclonal antibodies specific for Friend murine leukemia virus-induced erythroleukemia cells: Friend-specific and FMR-specific antigens. Virology112:131–144.
11. Cohen, G. B., R. Ren, and D. Baltimore.1995. Modular binding domains in signal transduction proteins. Cell80:237–248.
12. Cortez, D., L. Kadlec, and A. M. Pendergast.1995. Structural and signaling requirements for BCR-ABL-mediated transformation and inhibition of ap-optosis. Mol. Cell. Biol.15:5531–5541.
13. DeClue, J. E., and G. S. Martin.1989. Linker insertion-deletion mutagenesis of the v-srcgene: isolation of host- and temperature-dependent mutants. J. Virol.63:542–554.
14. Downward, J.1994. The GRB2/Sem-5 adaptor protein. FEBS Lett.338:113– 117.
15. DuBridge, R. B., P. Tang, H. C. Hsai, P.-M. Leong, J. H. Miller, and M. P. Calos.1987. Analysis of mutation in human cells by using an Epstein-Barr virus shuttle system. Mol. Cell. Biol.7:379–387.
16. Ellis, C., M. Moran, F. McCormick, and T. Pawson.1990. Phosphorylation of GAP and GAP-associated proteins by transforming and mitogenic ty-rosine kinases. Nature (London)343:377–381.
17. Engelman, A., and N. Rosenberg.1990.bcr/ablandsrcbut notmycandras replace v-ablin lymphoid transformation. Mol. Cell. Biol.10:4365–4369.
on November 9, 2019 by guest
http://jvi.asm.org/
18.Engelman, A., and N. Rosenberg.1990. Temperature-sensitive mutants of Abelson murine leukemia virus deficient in protein tyrosine kinase activity. J. Virol.64:4242–4251.
19. Hawley, R. G., F. H. Lieu, A. Z. Fong, and T. S. Hawley.1994. Versatile retroviral vectors for potential use in gene therapy. Gene Ther.1:136–138. 20. Ilaria, R. L. J., and R. A. Van Etten.1995. The SH2 domain of P210BCR/ABL is not required for the transformation of hematopoietic factor-dependent cells. Blood86:3897–3904.
21.Katz, M. E., and F. McCormick.1997. Signal transduction from multiple Ras effectors. Curr. Opin. Genet. Dev.7:75–79.
22.Kerkhoff, E., and U. R. Rapp.1998. Cell cycle targets of Ras/Raf signaling. Oncogene17:1457–1462.
23.Khosravi-Far, R., S. Campbell, K. L. Rossman, and C. J. Der.1998. Increas-ing complexity of Ras signal transduction: involvement of Rho family pro-teins. Adv. Cancer Res.72:57–107.
24. Kruh, G. D., R. Perego, T. Miki, and S. A. Aaronson.1990. The complete coding sequence of arg defines the Abelson subfamily of cytoplasmic tyrosine kinases. Proc. Natl. Acad. Sci. USA87:5802–5806.
25. Lugo, T., and O. N. Witte.1989. The BCR-ABL oncogene transforms Rat-1 cells and cooperates with v-myc. Mol. Cell. Biol.9:1263–1270.
26. Marengere, L. E. M., Z. Songyang, G. D. Gish, M. D. Schaller, J. T. Parsons, M. J. Stearn, L. C. Cantley, and T. Pawson.1994. SH2 domain specificity and activity modified by a single residue. Nature (London)369:502–505. 27. Marshall, C.1996. Ras effectors. Curr. Opin. Cell Biol.8:197–204. 28. Mayer, B. J., P. K. Jackson, R. A. Van Etten, and D. Baltimore.1992. Point
mutations in theablSH2 domain coordinately impair phosphotyrosine bind-ing in vitro and transformbind-ing activity in vivo. Mol. Cell. Biol.12:609–618. 29.McLaughlin, J., E. Chianese, and O. N. Witte.1989. Alternative forms of the
BCR-ABLoncogene have quantitatively different potencies for stimulation of immature lymphoid cells. Mol. Cell. Biol.9:1866–1874.
30. Muller, A. J., J. C. Young, A.-M. Pendergast, M. Pondel, N. R. Landau, D. R. Littman, and O. N. Witte.1991.BCRfirst exon sequences specifically acti-vate theBCR/ABLtyrosine kinase oncogene of Philadelphia chromosome-positive human leukemias. Mol. Cell. Biol.11:1785–1792.
31. Owen-Lynch, P. J., A. K. Wong, and A. D. Whetton.1995. v-Abl-mediated apoptotic suppression is associated with SHC phosphorylation without con-comitant mitogen-activated protein kinase activation. J. Biol. Chem.270:
5956–5962.
32. Parmar, K., and N. Rosenberg.1996. Ras complements the carboxyl termi-nus of v-Abl protein in lymphoid transformation. J. Virol.70:1009–1015. 33. Pendergast, A. M., M. L. Gishizky, M. H. Havlik, and O. N. Witte.1993. SH1
domain autophosphorylation of P210 BCR/ABL is required for transforma-tion but not growth factor independence. Mol. Cell. Biol.13:1728–1736. 34. Radfar, A., I. Unnikrishnan, H.-W. Lee, R. A. DePinho, and N. Rosenberg.
1998. P19Arfinduces p53-dependent apoptosis during Abelson virus-medi-ated pre-B cell transformation. Proc. Natl. Acad. Sci. USA95:13194–13199. 35.Raffel, G. D., K. Parmar, and N. Rosenberg.1996.In vivoassociation of v-Abl with Shc mediated by a non-phosphotyrosine-dependent SH2 interac-tion. J. Biol. Chem.271:4640–4645.
36.Rosenberg, N., and D. Baltimore.1976. A quantitative assay for
transforma-tion of bone marrow cells by Abelson murine leukemia virus. J. Exp. Med.
143:1453–1463.
37.Rosenberg, N., and O. N. Witte.1988. The viral and cellular forms of the Abelson (abl) oncogene. Adv. Virus Res.35:39–81.
38. Sawyers, C. L., W. Callahan, and O. N. Witte.1992. Dominant negative myc blocks transformation by abl oncogenes. Cell70:901–910.
39. Sawyers, C. L., J. McLaughlin, and O. N. Witte.1995. Genetic requirement for ras in the transformation of fibroblasts and hematopoietic cells by the bcr-abl oncogene. J. Exp. Med.181:307–313.
40. Scher, C. D., and R. Siegler.1975. Direct transformation of 3T3 cells by Abelson murine leukemia virus. Nature (London)253:729–731.
41. Schiff-Maker, L., M. C. Burns, J. B. Konopka, S. Clark, O. N. Witte, and N. Rosenberg.1986. Monoclonal antibodies specific for v-abl- and c-abl -en-coded molecules. J. Virol.57:1182–1186.
42. Skorski, T., A. Bellacosa, M. Nieborowska-Skorska, M. Majewski, R. Mar-tinez, J. K. Choi, R. Trotta, P. Wlodarski, D. Perrotti, T. O. Chan, M. A. Wasik, P. N. Tsichlis, and B. Calabretta.1997. Transformation of hemato-poietic cells by BCR/ABL requires activation of a PI-3K/Akt-dependent pathway. EMBO J.16:6151–6161.
43. Songyang, Z., S. E. Shoelson, M. Chadhuri, G. Gish, T. Pawson, W. G. Haser, F. King, T. Roberts, S. Ratnofsky, R. J. Lechleider, B. G. Neel, R. B. Birge, J. E. Fajardo, M. M. Chou, H. Hanafusa, B. Schaffhausen, and L. C. Cantley.1993. SH2 domains recognize specific phosphopeptide sequences. Cell72:767–778.
44. Tian, M., and G. S. Martin.1996. Reduced phosphotyrosine binding by the v-Src SH2 domain is compatible with wild-type transformation. Oncogene
12:727–734.
45. Unnikrishnan, I., A. Radfar, J. Jenab-Wolcott, and N. Rosenberg.1999. p53 mediates apoptotic crisis in primary Abelson virus-transformed pre-B cells. Mol. Cell. Biol.19:4825–4831.
46. Van Parijs, L., Y. Refaeli, J. D. Lord, B. H. Nelson, A. K. Abbas, and D. Baltimore.1999. Uncoupling IL-2 signals that regulate T cell proliferation, survival, and Fas-mediated activation induced cell death. Immunity11:281– 288.
47. Verderame, M. F., J. M. Kaplan, and H. E. Varmus.1989. A mutation in v-srcthat removes a single conserved residue in the SH-2 domain of pp60v-src restricts transformation in a host-dependent manner. J. Virol.63:338–348. 48.Waksman, G., S. E. Shoelson, N. Pant, D. Cowburn, and J. Kuriyan.1993.
Binding of a high affinity phosphotyrosyl peptide to the Src SH2 domain: crystal structures of the complexed and peptide-free forms. Cell72:779–790. 49. Warren, D., A. J. Heilpern, K. Berg, and N. Rosenberg.2000. The carboxyl terminus of v-Abl protein can augment SH2 domain function. J. Virol.
74:4495–4504.
50. Wong, K. K., J. D. Hardin, S. Boast, C. L. Cooper, K. T. Merrell, T. G. Doyle, S. P. Goff, and K. L. Calame.1995. A role for c-Abl in c-myc regulation. Oncogene10:705–711.
51. Yamanashi, Y., and D. Baltimore.1997. Identification of the Abl- and ras-GAP-associated 62 kDa protein as a docking protein, Dok. Cell88:205–211. 52. Zou, X., S. Rudchenko, K.-K. Wong, and K. Calame.1997. Induction of c-myc transcription by the v-Abl tyrosine kinase requires Ras, Raf1, and cyclin-dependent kinases. Genes Dev.11:654–662.