JOURNAL OFVIROLOGY,
0022-538X/99/$04.00⫹0 Sept. 1999, p. 7497–7504 Vol. 73, No. 9
Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Expression of Noncovalent Hepatitis C Virus Envelope E1-E2
Complexes Is Not Required for the Induction of Antibodies
with Neutralizing Properties following DNA Immunization
A. FOURNILLIER,1E. DEPLA,2P. KARAYIANNIS,3O. VIDALIN,1G. MAERTENS,2C. TRE´PO,1ANDG. INCHAUSPE´1*
INSERM U271, Virus des he´patites, Re´trovirus humains et Pathologies associe´es, 69424 Lyon Ce´dex, France1;
Hepatitis Program, INNOGENETICS, B-9052 Ghent, Belgium2; and Imperial College School of Medicine
at St. Mary’s, Department of Medicine, London W2 INY, United Kingdom3
Received 17 February 1999/Accepted 4 June 1999
Interactive glycoproteins present on the surface of viral particles represent the main target of neutralizing antibodies. The ability of DNA vaccination to induce antibodies directed at such structures was investigated by using eight different expression plasmids engineered either to favor or to prevent interaction between the hepatitis C virus (HCV) envelope glycoproteins E1 and E2. Independently of the injection route (intramuscular or intraepidermal), plasmids expressing antigens capable of forming heterodimers presumed to be the pre-budding form of the HCV envelope protein complex failed to induce any significant, stable antibodies following injection in mice. In sharp contrast, high titers of antibodies directed at both conformational and linear determinants were induced by using plasmids expressing severely truncated antigens that have lost the ability to form native complexes. In addition, only a truncated form of E2 induced antibodies reacting against the hypervariable region 1 of E2 (specifically with the C-terminal part of it) known to contain a neutralization site. When injected intraepidermally into small primates, the truncated E2-encoding plasmid induced antibodies able to neutralize in vitro the binding of a purified E2 protein onto susceptible cells. Because such antibodies have been associated with viral clearance in both humans and chimpanzees, these findings may have important implications for the development of protective immunity against HCV.
Hepatitis C virus (HCV) is the major causative agent of transfusion-associated and community-acquired non-A, non-B hepatitis worldwide (6, 22). More than 70% of HCV infections become chronic, with a significant risk in 5 to 20% of cases of progression to liver cirrhosis (1) and hepatocellular carcinoma (33). Only 20 to 30% of long-term responses occur in patients treated with alpha interferon (IFN-␣), the currently used ther-apy (15). The development of new therapeutic agents as well as a vaccine for prevention or treatment of HCV infections has become a priority. A first step in designing a vaccine is the identification of both host and viral components involved in the development of neutralizing immunity. In the HCV model, such protection may in part be due to neutralizing antibodies targeted at the envelope glycoproteins E1 and E2. Successful in vivo protection of chimpanzees has been achieved following immunization with recombinant E1 and E2 proteins and has been linked to the induction of specific anti-E2 antibodies (5). Such antibodies neutralizing in vitro the binding of purified E2 onto susceptible cells, referred as “neutralizing of binding” (NOB) antibodies (32), have recently been linked to the res-olution of chronic infection in humans (21). Several observa-tions have shown that the hypervariable region 1 (HVR-1) of E2 contains an important neutralization domain. In particular, antibodies present in the sera of infected patients or induced by immunization and targeted at this region can prevent viral infection in cell cultures (37, 44). In contrast to E2 anti-bodies, to date, the participation of anti-E1 antibodies in viral clearance remains undocumented.
Various studies using transient viral and nonviral expression
systems have shown that HCV envelope glycoproteins E1 and E2 interact to form complexes (17, 29). Two forms of E1-E2 complexes are detected: heterogeneous disulfide-linked aggre-gates formed by misfolded proteins and heterodimers stabi-lized by noncovalent interactions composed of native glycop-roteins (8, 10). The latter have been proposed as the prebudding form of the HCV envelope glycoprotein complex. Conformation-sensitive E2-reactive monoclonal antibodies (MAbs [H2 and HMAb 503]) have recently been described which selectively recognize noncovalently associated com-plexes, allowing the distinction to be made between native complexes and misfolded aggregates (8, 18). As described for human immunodeficiency virus envelope proteins (11, 31), in-teractions between HCV glycoproteins could affect epitope presentation and have an important influence not only on the antigenicity of the proteins but also on their immunogenicity. Genetic immunization, which allows the de novo synthesis of the DNA-expressed antigens in the host’s cells (42), has been shown to elicit both protective humoral and cellular immune responses in several animal models of viral infection (2, 30, 39, 40). This vaccination mode, similar to strategies based on the use of attenuated viruses or live expressing vectors, provides the biological context for antigens to be naturally processed with respect to posttranslational modifications, protein folding, and assembly (38). The opportunity for de novo-synthesized proteins to achieve proper maturation is a particularly impor-tant element in the case of proteins that require the help of additional partners to fully mature. An example of such pro-teins are propro-teins constituting viral envelopes. These propro-teins, usually glycoproteins, often display complex interactions be-tween themselves and/or cellular partners for the constitution of functional, native envelope complexes (16, 19). The inter-actions between HCV E1 and E2 proteins thus offer a good * Corresponding author. Mailing address: INSERM U271, 151
Cours Albert Thomas, Lyon 69424, France. Phone: 33.4.72.68.19.88. Fax: 33.4.72.68.19.71. E-mail: [email protected].
7497
on November 9, 2019 by guest
http://jvi.asm.org/
model to study the advantages and limitations of DNA-based immunizations for the induction of antibodies directed at an-tigenic structures existing as complexes and representing crit-ical components of a vaccine (5, 21).
Here, we report on the efficacy of different plasmids engi-neered to favor or limit the formation of E1-E2 complexes at inducing specific antibodies and cytokine release. We showed that expression of presumed native E1-E2 complexes failed to induce any significant humoral responses, whereas optimal re-sponses (including anti-E2 antibodies with neutralizing of binding activity) were obtained in mice and primates with trun-cated forms of the proteins.
MATERIALS AND METHODS
Plasmids and in vitro expression studies.E1 and E2 sequences were amplified from a vector containing the full-length cDNA sequence of the HCV-H strain, 1a and cloned into theXbaI-NotI orSmaI sites of the pCI vector (Promega), resulting in the pCI-based vectors (Fig. 1). Dicistronic expression vectors, allow-ing the coexpression of two distinct genes from two transcripts within the same cell, were generated with the vector pFX (generous gift from M. Nasoff), which contains two cytomegalovirus (CMV) promoters followed by onePacI orNotI
site for the cloning (Fig. 1). All plasmid-cloned fragments were verified by sequencing (34).
Expression of E1 and E2 antigens was examined in cell extracts and superna-tants by using a capture enzyme-linked immunosorbent assay (ELISA) employ-ing highly specific monoclonal antibodies (23a) as well as by immunoprecipita-tion following transient transfecimmunoprecipita-tion of Cos-7 cells. For the capture ELISA, cells from three 35-mm wells were pooled and lysed in a mixture of 10 mM MOPS (morpholine propanesulfonic acid [pH 6.5]), 10 mM NaCl, 1 mM EGTA, and 1% Triton X-100. Cell lysates were then used to coat microtiter plates at a 1⁄2 dilution. After blocking, MAbs directed against either E1 or E2 were incubated. The MAb was detected with a goat anti-mouse peroxidase conjugate. Cotrans-fection of the chloramphenicol acetyltransferase (CAT)-expressing plasmid pcDNA3/CAT (Invitrogen) was employed to monitor transfection efficiency. Transfected cells (Lipofectamine plus; Gibco BRL) were metabolically labeled at 24 h posttransfection with 200Ci of [35S]methionine per ml during 24 h and isolated by using the MACSelect-transfected cell selection kit (Miltenyl Biotec). Cell suspensions were then centrifuged, and the pellets were lysed with 0.5% Nonidet P-40 in a mixture of 10 mM Tris-HCl (pH 7.5), 150 mM NaCl, 2 mM EDTA, 20 mM iodoacetamide, and 10g of aprotinin per ml. HCV-specific proteins were immunoprecipitated from the clarified lysates as described previ-ously (9) by using two anti-E2 MAbs that selectively recognize E1-E2 noncova-lent complexes: a murine antibody, H2 (8), and a human one, 503 (18). Immu-noprecipitates were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis.
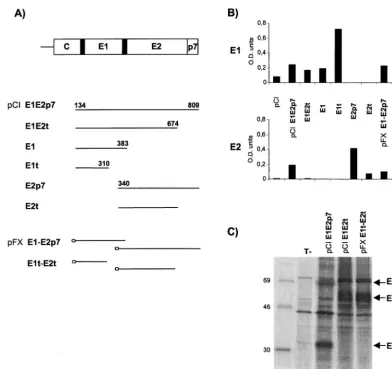
[image:2.612.108.500.69.438.2]DNA-based immunization. (i) Mice.Six- to eight-week-old female BALB/c mice (n⫽5 or 6 per group) were purchased from Charles River. All DNA FIG. 1. Plasmids and in vitro expression studies. (A) Schematic representation of the HCV envelope region. Black boxes correspond to signal peptide sequences. Sequences expressed by the indicated plasmids are shown diagrammatically by bars. Empty boxes represent the two CMV promoters of the pFX vector. (B) Quantitative determination of intracellular expression of E1 and E2 antigens by capture ELISA. CAT determination was realized after cotransfection of pcDNA3/CAT with each plasmid to standardize transfection efficiency. Results are given as OD values. (C) Identification of HCV envelope proteins and complexes. Transfected Cos-7 cells were labeled with [35S]methionine, and cell lysates were immunoprecipitated with either the anti-E2 H2 MAb or the 503 HMAb, and representative results are shown. Transfected plasmids are indicated at the top of each lane, T⫺, pCI. Samples were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (13% polyacrylamide). The positions of the14C-labelled protein markers are shown on the left, and those of HCV-specific proteins are shown on the right.
on November 9, 2019 by guest
http://jvi.asm.org/
preparations were produced with endotoxin-free purification columns (Qiagen). Immunizations were performed with either a gene gun (PowderJect), resulting in the injection of 5g of plasmid DNA into the abdominal skin (referred to as intraepidermal [i.e.] injection) or with a syringe in the anterior tibialis muscle (referred to as intramuscular [i.m.] injection) by using 100g of plasmid DNA per injection at weeks 0, 9, and 21 as previously described (13, 24).
(ii) Tamarins.Six adult female tamarins (Saguinus labiatus) seronegative for HCV were individually housed and cared for according to approved standard operating procedures. Animals were tranquilized for inoculations and blood collections. Four animals were injected with 20g of total DNA per injection per boost in the abdomen with the gene gun, and the two other animals were immunized intramuscularly in the tibialis muscle of one leg with 400g of DNA in saline at weeks 0, 5, 9, and 20.
Antibody titers and isotypes.Induced antibodies were measured with a specific ELISA (INNO-test for anti-E1 and anti-E2 antibodies; Innogenetics) as previ-ously described (23a, 28). Sera from tamarins were analyzed with an antihuman immunoglobulin G (IgG) Fe labeled with horseradish peroxidase (Dako) used as a secondary antibody. Antibody titers were calculated as the serial threefold dilution which gave an optical density (OD) that equaled the cutoff. For mice, the cutoff value was established as the mean OD⫹3SD of 10 sera obtained from control mice, while for tamarins, it was determined for each animal as the equivalent to 3⫻the OD of sera obtained before the primary injection.
Peptide-based epitope mapping.Peptide A1H encompassing the HCV-H hy-pervariable region (HVR-1) of the E2 protein, located between amino acids (aa) 384 and 411 on the polyprotein (41), was used in the ELISA as previously described (13). Sera were tested at a 1:100 dilution, and only those giving absorbance values⬎2.5 times the absorbance value of negative sera were con-sidered positive. Nineteen decamers overlapping by 9 aa and encompassing the A1H peptide were used: N1 to N8 for N-terminally-located peptides and C9 to C19 for C-terminally-located peptides. ELISA plates were first coated with streptavidin (Sigma), and assays were performed as described above by using the 19 peptides, each of which carried a biotinylated spacer peptide at its N terminus (Neosystem).
Neutralization assay.Mice and tamarin sera were tested for their ability to neutralize the binding of the E2 protein onto MOLT-4 cells in the recently developed NOB assay (32). Quantification of NOB antibodies was performed as previously described (21), by incubation of a suboptimal concentration of bio-tinylated recombinant CHO cell E2 protein (1g/ml) with different dilutions of the tested sera. E2 binding to target cells was detected with a streptavidin-phycoerythrin conjugate (2.5g/ml).
Cytokine measurements.In vitro cytokine production was analyzed at 32 weeks post primary injection following antigen-specific in vitro stimulation of splenic cells from immunized mice as previously described (13). Cells were stimulated with the same recombinant E1 and E2 proteins used for antibody titration, added at final concentrations of 2 or 5g/ml. Culture supernatants were harvested at 24 h for interleukin-2 (IL-2) and 48 h for IL-4, IL-5 and IFN-␥ testing, and cytokine levels were measured by quantitative ELISA (Biotrak; Amersham). All cultures were performed in duplicate, and the data indicated represent the ranges of cytokine concentration obtained with the different con-centrations of antigens.
RESULTS
In vitro expression studies: different plasmids can favor or limit the formation of complexed structures.In order to favor or, on the contrary, to prevent E1-E2 interactions, we designed several plasmids expressing either the full-length envelope re-gion of HCV or full-length and/or truncated forms of each envelope protein. These included monocistronic vectors ex-pressing the full-length envelope region (pCI E1E2p7), E1 or E2p7 proteins (pCI E1 and pCI E2p7), or truncated forms of the proteins lacking their transmembrane domains (pCI E1t, pCI E2t, and pCI E1E2t) as well as their counterpart dicis-tronic plasmids (pFX E1-E2p7 and pFX E1t-E2t) (Fig. 1A).
Expression of E1 and E2 was confirmed by immunofluores-cence analyses after transient transfection of Cos-7 cells. Dif-fuse cytoplasmic localization of both antigens was observed independent of the plasmid used (data not shown). Quantita-tive determination of the intracellular expression of E1 and E2 showed differences between the amounts of antigen produced by the different plasmids (Fig. 1B). Expression of the truncated form of E1 led to higher antigen production than that obtained with the full-length form of the antigen. Differences in E2 production between the plasmids used were less pronounced with the exception of one plasmid, pCI E1E2t, which led to a very low production of E2 protein. Antigens were not detected
in the supernatants of transfected cells (i.e., secreted antigens), even when the E1 and E2 proteins were expressed without their hydrophobic anchor domains (from aa 311 for E1 and aa 675 for E2). This suggests that the secretion level of the ex-pressed antigens, if secretion does take place, is very low or that the proteins are unstable. Overall, under the experimental conditions used, the capacity to produce secreted antigens does not appear dramatically different between the plasmids, the E1 and E2 proteins remaining mainly intracellular.
Immunoprecipitation studies revealed that when expressed as full-length proteins either from a monocistronic plasmid, pCI E1E2p7 (Fig. 1C, lane 2), or from a dicistronic plasmid, pFX E1-E2p7 (data not shown), E1 and E2 have the capacity, as expected, to form noncovalent heterodimers coprecipitated selectively by a conformation-sensitive MAb. No E1-E2 non-covalent complexes were detected any longer when the anti-gens were expressed from plasmids encoding C-terminally truncated forms of one or both proteins (pCI E1E2t and pFX E1t-E2t), as shown in the figure (lanes 3 and 4). The lack of immunoprecipitation with the pCI E1E2t plasmid is in accord-ance with previous reports (14, 26, 36) which indicate that the carboxy terminus of E2 is critical for the E1-E2 interaction. The absence of coprecipitation of a truncated form of E1 with a truncated form of E2 indicates that if E1t and E2t form complexes, they are not properly folded, as suggested by Michalak et al. (26), and therefore are not recognized by the conformation-sensitive HMAb 503. Thus, the ability of the different plasmids to express native interactive proteins could be governed by the cloning strategy implemented.
Humoral responses to HCV E1 and E2 antigens in mice: a strict dependence on the form of the DNA-expressed antigens. For all monocistronic pCI constructs, DNA immunizations were performed by using the intraepidermal route, which has been shown to result in higher antibody titers than the i.m. route (13, 28). For the dicistronic pFX constructs, we com-pared three injection modes: i.e., i.m., and a combination of both (i.e.⫹i.m.).
Seroconversion rates and antibody titers are indicated in Fig. 2. Overall, both antibody titers and seroconversion rates were much higher for E2-encoded plasmids than E1-encoded ones. Except for four animals that displayed titers⬎1:2,000, all oth-ers had no detectable or extremely low anti-E1 antibody titoth-ers, whatever the E1-encoding plasmid injected. This observation suggests that E1 is a very poor immunogen and limits the evaluation of the induced anti-E1 responses.
Despite a total of three injections and independent of the injection route, some E2-encoding plasmids never produced any detectable antibodies or produced antibodies only at very low titers: pCI E2p7, pCI E1E2p7, and pFX E1-E2p7 (maxi-mal median titers ranged from 1:40 to 1:8,510). These results were obtained with both mono- and dicistronic plasmids and corresponded to plasmids containing the full-length sequence of E2. Mouse sera were in addition used to perform immuno-precipitation studies with cell extracts expressing E1-E2 com-plexes (obtained by infection with recombinant vaccinia viruses expressing such complexes). No positive signals could be de-tected when sera corresponding to mice injected with plasmids expressing full-length E1 and/or E2 antigens were used (data not shown), arguing for a lack of specific antibodies in these sera. In sharp contrast, plasmids expressing a truncated form of E2 (pCI E2t and pFX E1t-E2t) yielded much higher titers of long-lasting antibodies, with maximal median titers reaching 1:30,000. Such antibodies have been shown in addition to be capable of immunoprecipitating E1-E2 complexes (28). One notable exception was the plasmid pCI E1E2t, expressing a full-length E1 antigen together with a truncated form of E2
VOL. 73, 1999 DNA IMMUNIZATION WITH HCV-SPECIFIC PLASMIDS 7499
on November 9, 2019 by guest
http://jvi.asm.org/
which induces very low anti-E2 antibody titers and very tran-sient anti-E1 antibodies. The use of dicistronic plasmids was overall equivalent to that of the monocistronic ones. Compar-ison of the three different modes of immunization used for injection of the dicistronic constructs indicated that immuni-zation modes which included an i.e. route always resulted in higher antibody titers (Fig. 2B). The latter observation is in agreement with previous observations made by using monocis-tronic E2-expressing plasmids (13, 28). Altogether, the nature of the immunization route could not compensate for the very poor performance of the plasmids expressing full-length E1 and E2 antigens.
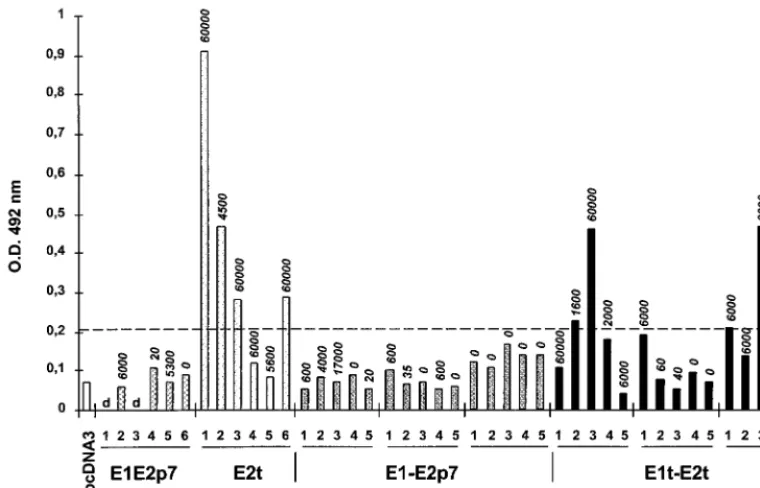
Epitope mapping: peptide epitopes within the HVR-1 are mainly recognized by antibodies induced following injection of truncated DNA-expressed E2 antigen.Because of its associa-tion with the inducassocia-tion of neutralizing antibodies (12, 32, 37, 43), the reactivity of all sera against the hypervariable region located at the N terminus of E2 (HVR-1) (41) was more extensively analyzed (Fig. 3). Essentially two plasmids induced antibodies recognizing a peptide corresponding to the 27 aa of the HVR-1, both of them encoding a truncated form of E2 (pCI E2t and pFX E1t-E2t). However, remarkably, there was no correlation between the overall antibody titers detected in
the ELISA (Fig. 2 and reported in Fig. 3 for each mouse on top of the bars) and the reactivity against the HVR-1 peptide. In the group of mice injected by the i.e.⫹i.m. route with the pFX E1t-E2t plasmid, one serum sample with a titer of 1:60 (mouse 5) displayed a good reactivity, while another with a higher titer of 1:6,000 (mouse 2) had no detectable activity against A1H. These data show that independent of the anti-E2 antibody titers, plasmids encoding a truncated form of E2 remain the best at inducing production of antibodies targeted at this do-main. Whatever the expression context or the injection route, plasmids expressing full-length proteins fail to induce such antibodies.
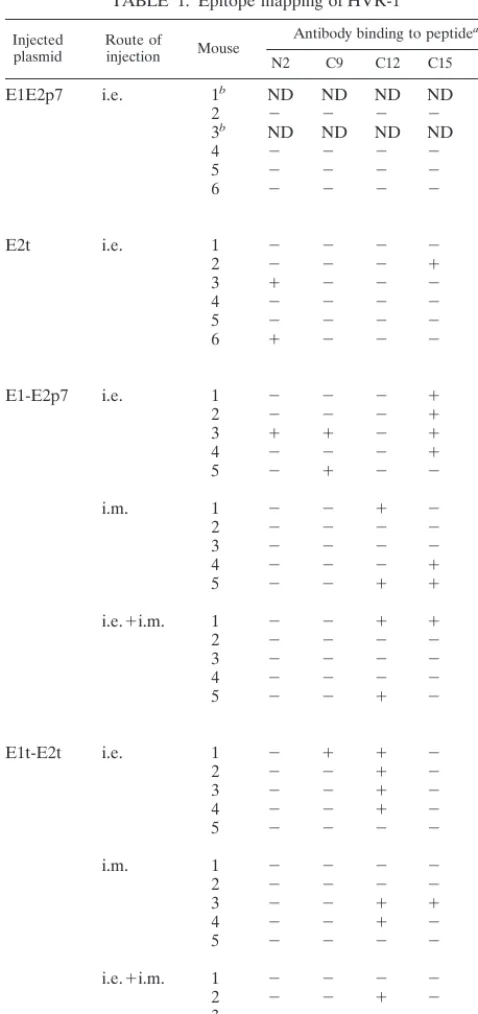
When all 46 mouse serum samples were further analyzed for their reactivity against minimal determinant domains mapping within the HVR-1 (using 19 overlapping peptides termed N1 to N8 and C9 to C19 according to their N- or C-terminal location within the HVR-1), it was observed that in addition to plasmids encoding a truncated form of E2, plasmids encoding full-length antigen displayed a low but detectable reactivity against some peptides (i.e., the pCI E1E2p7 and the pCI E1E2t plasmids reacting with peptide C15) (Table 1). Overall, immune recog-nition was mainly targeted at C-terminally-located peptides independent of the plasmid used. Four of the five reactive FIG. 2. Antibody (Ab) titers and seroconversion. Each plasmid indicated on Fig. 1 was injected into one group of mice either by the i.e. route (monocistronic constructs) or by one of the three injection modes, i.e., i.m., or a combination of both (i.e.⫹i.m.). Median titers, given for seroconverted mice only, are represented. Results are shown as the reciprocal of the serum dilution equivalent to 3⫻the OD of sera from mice injected with the vector pcDNA3. The numbers shown on top of each bar indicate the number of animals that seroconverted per animal group. (A) Responses to proteins expressed by monocistronic pCI-derived constructs. (B) Responses to proteins expressed by dicistronic pFX-derived constructs. p.i., post-primary injection.
on November 9, 2019 by guest
http://jvi.asm.org/
peptides (N2, C9, C12, C15, and C17) were located in this region.
IgG isotypes and in vitro splenic cytokine production: the form of the DNA-expressed antigens has no influence.Analysis of anti-E1 or anti-E2 antibodies (at 21 and 31 weeks post-primary injection) revealed marked differences between the isotypes of E1- and E2-specific antibodies, which were inde-pendent of the injection route and plasmids used. IgG2a or -2b was detected in E2-reactive antibodies, while IgG1 or IgG3 was observed in anti-E1 antibodies (data not shown).
Inasmuch as the IgG isotype profiles of anti-HCV E1 and E2 antibodies suggested the priming of different T helper cell subsets, direct measurements of Th1-specific cytokines (IFN-␥ or IL-2) and Th2-specific cytokines (IL-4 or IL-5) were per-formed. While insignificant levels of IL-4 and IL-5 were de-tected, splenocytes from immunized mice showed production of IFN-␥(ranging from 30 to 132 pg/ml for E1 and 34 to 855 pg/ml for E2) and IL-2 (ranging from 27 to 73 pg/ml for E1 and 32 to 162 pg/ml for E2) following antigen-specific in vitro stimulation. The different levels of cytokines produced could not be correlated to a specific plasmid or an injection route. Overall, the form of the expressed E1 and E2 did not affect the isotype of the induced antibodies or the in vitro splenic cyto-kine profiles.
Humoral immune responses to HCV E2 in tamarins: expres-sion of a truncated form of E2 can induce antibodies with NOB capacity.To evaluate some of the observations made with the murine model, the pCI E2t plasmid was used to immunize small primates. We immunized six tamarins, four by an i.e. injection route (via the gene gun) and two others by the i.m. injection route.
All four of the tamarins immunized intraepidermally devel-oped an E2-specific antibody response, detected either after the first boost in two animals or following the third boost in the two others (Fig. 4). Antibody titers reached 1:6,000 in two
animals (tamarins 1 and 4). The E2-specific antibody titers declined with time to reach low but detectable levels after 3 months of follow-up. Upon i.m. immunization, only one tama-rin of the two injected (tamatama-rin 5) developed antibodies early after the primary injection (3 weeks), but this response was relatively short lived and could not be enhanced by additional booster injections. Anti-E2 antibody isotypes were of the IgG2 type, in agreement with those previously observed in mice (this study and references 13 and 28). Only one animal (tamarin 4) displayed reactivity against the HVR-1 peptide, and the con-tribution of small peptides to the overall responses was basi-cally null in tamarins (no antibodies against any of the 19 decamers were detected).
Surprisingly and in contrast to observations made with mice for which all previously tested plasmids failed to induce anti-bodies with NOB activity (data not shown), three of the four i.e.-injected tamarins developed NOB antibodies (Fig. 4). Sera with NOB activity corresponded to sera displaying the highest anti-E2 antibody titers. This was particularly unexpected in view of the fact that sera from mice injected with the same plasmid induced anti-E2 antibody titers that were always at least 1 log higher than those detected in tamarins (Fig. 2A). This finding suggests that the host specificity in addition to the form of the expressed antigen is a critical component in the induction of antibodies with biological relevance.
DISCUSSION
[image:5.612.111.491.75.319.2]Components of viral envelopes include, apart from cellular proteins, specific viral antigens, mainly glycoproteins, that play a critical role in the induction of neutralizing antibodies. In-teraction between such proteins has been shown to govern the accessibility of epitopes to the immune system and, therefore, the quality of the immune response mounted by an infected or immunized host (11, 31). By using eight different plasmids
FIG. 3. Reactivity of mice sera against the HVR-1. Sera obtained at 25 weeks post-primary injection from each mouse of the different injected groups were tested against the A1H peptide. The different groups of mice are given along the X axis, while the binding results obtained are given as OD values on the Y axis. The cutoff value is indicated by the dotted line.
VOL. 73, 1999 DNA IMMUNIZATION WITH HCV-SPECIFIC PLASMIDS 7501
on November 9, 2019 by guest
http://jvi.asm.org/
engineered to allow or prevent HCV E1 and E2 envelope glycoproteins to form heterodimers proposed as the prebud-ding form of the HCV envelope complex, together with differ-ent immunization routes, we demonstrated here that only plas-mids expressing truncated forms of the antigens that have lost the ability to assemble into native noncovalent complexes are capable of inducing detectable antibodies. For the E2 protein, all plasmids encoding a truncated form of the antigen (with one exception, pCI E1E2t) were associated with a considerable benefit in seroconversion rates and antibody titers induced.
The fact that the pCI E1E2t construct failed to induce a sig-nificant humoral response may be simply related to the very low level of antigen expression observed compared with those of other plasmids (Fig. 1B).
There are different hypotheses that could account for the fact that native complexed antigens fail to induce a significant antibody response. Previous reports have shown that secreted DNA-expressed antigens versus nonsecreted ones seem to in-duce optimum immune responses (3, 20). Indeed, in our study, the secretion of full-length E1 and E2 proteins, complexed or not, may be dramatically impaired or prevented. Thus, the proteins may remain trapped within the endoplasmic reticulum as we observed here. In vitro experiments, with recombinant viruses expressing HCV antigens or a stable cell line expressing the full-length HCV polyprotein (25–27, 36) suggest that these proteins are not secreted. The lack of secretion of the HCV glycoproteins, therefore, may have resulted in the lack of pre-sentation of antigenic epitopes to the immune system. Many reports have nonetheless showed that, even when using recom-binant viruses expressing E1 and/or E2 proteins devoid of their transmembrane domains, such as those used in our study, secretion of antigens is observed at very low levels only (26). Thus, secretion of the HCV E1 and E2 may not play the major role in the induction of the humoral response that we observed here. One other explanation is that the formation of E1-E2 complexes from native full-length proteins would result in the masking of important determinants, as described for the gp120 and gp41 subunits of the human immunodeficiency virus env protein (11), and in particular those associated with NOB activity. Because the NOB determinants remain to be mapped, this hypothesis is presently difficult to evaluate. Finally, when antigens are not or very poorly secreted, induction of antibod-ies may be due to the lysis of expressing cells by the specifically induced cellular immune response via the action of cytotoxic T lymphocytes (7). Although we did not analyze the induction of cytotoxic T lymphocytes in our study, the analysis of in vitro splenic cytokine production showed similar levels of IFN-␥and Il-2 secretion for all plasmids. These data are suggestive of the induction in all cases of a Th1-like response which does not appear to be enhanced by any particular plasmid. Thus, one likely hypothesis would indeed be the masking of important determinants as being the principal limiting factor in the in-duction of antibodies when interactive full-length E1 and E2 proteins are expressed.
[image:6.612.53.292.78.585.2]Our study reveals the very low immunogenic potential of E1 when directly injected as DNA, and this observation is concor-dant with data recently reported by Lee et al. from the rat model (23). In chronically infected chimpanzees, E1 anti-bodies have been efficiently raised by immunization with re-combinant E1 protein and were associated with clearance of viral antigen from the liver (23b). This recent observation indicates, although yet to be confirmed, a possible role of anti-E1 antibodies in the control of liver inflamation and thus accentuates the necessity to optimize E1-expressing plasmids. An important result achieved with constructs expressing a truncated form of E2 is the induction in mice of antibodies displaying reactivity against HVR-1 (Fig. 3), which contains an important neutralization domain (37, 44). Those were exclu-sively associated with the use of an i.e.-based injection route and were independent of the overall anti-E2 antibody titers induced. It has recently been proposed that the avidity of antibodies generated after the direct injection of recombinant plasmids was dependent on the immunization route (4). Al-though further studies concerning the avidity of the anti-HVR-1 antibodies are necessary, our results suggest that the i.e. route could induce antibodies with higher avidity than the TABLE 1. Epitope mapping of HVR-1
Injected
plasmid Route ofinjection Mouse
Antibody binding to peptidea
N2 C9 C12 C15 C17
E1E2p7 i.e. 1b ND ND ND ND ND
2 ⫺ ⫺ ⫺ ⫺ ⫺
3b ND ND ND ND ND
4 ⫺ ⫺ ⫺ ⫺ ⫺
5 ⫺ ⫺ ⫺ ⫺ ⫹
6 ⫺ ⫺ ⫺ ⫺ ⫺
E2t i.e. 1 ⫺ ⫺ ⫺ ⫺ ⫺
2 ⫺ ⫺ ⫺ ⫹ ⫺
3 ⫹ ⫺ ⫺ ⫺ ⫺
4 ⫺ ⫺ ⫺ ⫺ ⫺
5 ⫺ ⫺ ⫺ ⫺ ⫺
6 ⫹ ⫺ ⫺ ⫺ ⫺
E1-E2p7 i.e. 1 ⫺ ⫺ ⫺ ⫹ ⫺
2 ⫺ ⫺ ⫺ ⫹ ⫺
3 ⫹ ⫹ ⫺ ⫹ ⫺
4 ⫺ ⫺ ⫺ ⫹ ⫺
5 ⫺ ⫹ ⫺ ⫺ ⫹
i.m. 1 ⫺ ⫺ ⫹ ⫺ ⫺
2 ⫺ ⫺ ⫺ ⫺ ⫺
3 ⫺ ⫺ ⫺ ⫺ ⫺
4 ⫺ ⫺ ⫺ ⫹ ⫺
5 ⫺ ⫺ ⫹ ⫹ ⫺
i.e.⫹i.m. 1 ⫺ ⫺ ⫹ ⫹ ⫺
2 ⫺ ⫺ ⫺ ⫺ ⫺
3 ⫺ ⫺ ⫺ ⫺ ⫺
4 ⫺ ⫺ ⫺ ⫺ ⫺
5 ⫺ ⫺ ⫹ ⫺ ⫺
E1t-E2t i.e. 1 ⫺ ⫹ ⫹ ⫺ ⫺
2 ⫺ ⫺ ⫹ ⫺ ⫺
3 ⫺ ⫺ ⫹ ⫺ ⫺
4 ⫺ ⫺ ⫹ ⫺ ⫹
5 ⫺ ⫺ ⫺ ⫺ ⫺
i.m. 1 ⫺ ⫺ ⫺ ⫺ ⫹
2 ⫺ ⫺ ⫺ ⫺ ⫺
3 ⫺ ⫺ ⫹ ⫹ ⫺
4 ⫺ ⫺ ⫹ ⫺ ⫺
5 ⫺ ⫺ ⫺ ⫺ ⫺
i.e.⫹i.m. 1 ⫺ ⫺ ⫺ ⫺ ⫺
2 ⫺ ⫺ ⫹ ⫺ ⫺
3 ⫺ ⫺ ⫺ ⫺ ⫺
4 ⫺ ⫺ ⫹ ⫺ ⫺
5 ⫺ ⫺ ⫺ ⫺ ⫺
a⫹, OD values considered positive;⫺, negative values; ND, not determined. bMouse dead.
on November 9, 2019 by guest
http://jvi.asm.org/
i.m. route. Data obtained from a peptide-based scanning anal-ysis indicated that plasmids expressing either truncated or full-length E2 antigens induce antibodies mainly directed at the C terminus of HVR-1. Although some reports suggest that anti-bodies directed at the C terminus alone might not be sufficient for clearance of virus because chronically infected patients contain these antibodies (35, 44), another study shows that serum reacting to the C-terminal 13 aa of HVR-1 (aa positions 398 to 410) prevented isolate-specific infection with HCV in cell culture (37). Further investigation of the significance of anti-HVR-1 antibodies for elimination of HCV is needed to determine which type of antibody would be important to in-duce.
The most dramatic illustration of the benefit achieved by using a truncated HCV E2-expressing plasmid was that it was possible to induce in tamarins NOB antibodies, which have been linked to the prevention of infection in chimpanzees and the control of chronic infection in humans (21, 32). Although the induced NOB titers were low, clinical resolution of hepa-titis C was observed in some infected patients developing sim-ilar titers (21). In the tamarins, induction of such antibodies was dependent on the overall anti-E2 antibody titers, them-selves linked to the injection route (because antibody titers were constantly found to be higher when the i.e. route was used). DNA vaccination has been demonstrated to be effica-cious in a number of preclinical animal models (mice and rabbits), but data from primates have been more limited. The present study underlines the difficulty in extrapolating results observed in one species to another for a given DNA-expressed antigen. It also points perhaps to the most critical role played by the host cell’s machinery or the host-specific immune system compared with that played by the nature of the plasmid itself
or the injection route for the induction of biologically relevant antibodies.
Genetic immunization has been shown to be a very powerful inducer of immune responses when single or noninteractive antigens are expressed. We demonstrate here its limitation when plasmids that express interactive intracellular antigens such as HCV envelope proteins are used. As shown in our study, induction of at least one kind of neutralizing antibody may nonetheless be efficiently generated by using a plasmid engineered to express a nonnative antigen.
ACKNOWLEDGMENTS
We are grateful to PowderJect Vaccines for the lending of the gene gun. We thank D. Rosa and S. Abrigiani for the NOB analysis, J. Dubuisson for provision of antibody H2, and C. Wychowski for critical reading of the manuscript.
This work was supported by the European Commission (through both a BIOMED and BIOTECHNOLOGY grant). A.F. is a recipient of a Poste d’Accueil INSERM.
REFERENCES
1.Alter, H. J.1989. Chronic consequences of non-A, non-B hepatitis, p. 83–97. InL. B. Seef and J. H. Lewis (ed.), Current perspectives in hepatology. Plenum, New York, N.Y.
2.Boyer, J. D., K. E. Ugen, B. Wang, M. Agadjanyan, L. Gilbert, M. L. Bagarazzi, M. Chattergoon, P. Frost, A. Javadian, W. V. Williams, Y. Re-faeli, R. B. Ciccarelli, D. M. McCallus, L. Coney, and D. B. Weiner.1997. Protection of chimpanzees from high-dose heterologous HIV-1 challenge by DNA vaccination. Nat. Med.3:526–532.
3.Boyle, J. S., C. Koniaras, and A. M. Lew.1997. Influence of cellular location of expressed antigen on the efficacy of DNA vaccination: cytotoxic T lym-phocyte and antibody responses are suboptimal when antigen is cytoplasmic after intramuscular DNA immunization. Int. Immunol.9:1897–1906. 4.Boyle, J. S., A. Silva, J. L. Brady, and A. M. Lew.1997. DNA immunization:
induction of higher avidity antibody and effect of route on T cell cytotoxicity. FIG. 4. Profiles of anti-E2 antibodies (Ab) detected in tamarins injected with the pCI E2t construct and NOB titers. Anti-E2 titers are shown as the reciprocal of the serum dilution equivalent to 3⫻the OD of sera from each tamarin obtained before the primary injection. Open symbols correspond to i.e.-immunized animals, and solid symbols represent i.m.-injected animals. Booster injections are indicated by vertical arrows. NOB titers, expressed as the reciprocal of serum dilution which gave 50% neutralization of binding in the NOB assay, are presented in tabular form. p.i., post-primary injection.
VOL. 73, 1999 DNA IMMUNIZATION WITH HCV-SPECIFIC PLASMIDS 7503
on November 9, 2019 by guest
http://jvi.asm.org/
Proc. Natl. Acad. Sci. USA94:14626–14631.
5.Choo, Q.-L., G. Kuo, R. Ralston, A. Weiner, D. Chien, G. Van Nest, J. Han, K. Berger, K. Thudium, C. Kuo, J. Kansopon, J. McFarland, A. Tabrizi, K. Ching, B. Moss, L. B. Cummins, M. Houghton, and E. Muchmore.1994. Vaccination of chimpanzees against infection by the hepatitis C virus. Proc. Natl. Acad. Sci. USA91:1294–1298.
6.Choo, Q.-L., G. Kuo, A. J. Weiner, L. R. Overby, D. W. Bradley, and M. Houghton.1989. Isolation of a cDNA clone derived from a blood-borne non-A non-B viral hepatitis genome. Science244:359–362.
7.Davies, H. L., C. L. Brazollot Millan, and S. C. Watkins.1997. Immune-mediated destruction of transfected muscle fibers after direct gene transfer with antigen-expressing plasmid DNA. Gene Ther.4:181–188.
8.Deleersnyder, V., A. Pillez, C. Wychowski, K. Blight, J. Xu, Y. S. Hahn, C. M. Rice, and J. Dubuisson.1997. Formation of native hepatitis C virus glycop-rotein complexes. J. Virol.71:697–704.
9.Dubuisson, J., H. H. Hsu, R. C. Cheung, H. B. Greenberg, D. G. Russell, and C. M. Rice.1994. Formation and intracellular localization of hepatitis C virus envelope glycoprotein complexes expressed by recombinant vaccinia and Sindbis viruses. J. Virol.68:6147–6160.
10. Dubuisson, J., and C. M. Rice.1996. Hepatitis C virus glycoprotein folding: disulfide bond formation and association with calnexin. J. Virol.70:778–786. 11. Earl, P. L., C. C. Broder, D. Long, S. A. Lee, J. Peterson, S. Chakrabarti, R. W. Doms, and B. Moss.1994. Native oligomeric human immunodeficiency virus type 1 envelope glycoprotein elicits diverse monoclonal antibody reac-tivities. J. Virol.68:3015–3026.
12. Farci, P., A. Shimoda, D. Wong, T. Cabezon, D. De gionnis, A. Srazzera, Y. Shimizu, M. Shapiro, H. J. Alter, and R. H. Purcell.1996. Prevention of hepatitis C virus infection in chimpanzees by hyperimmune serum against the hypervariable region 1 of the envelope 2 protein. Proc. Natl. Acad. Sci. USA93:15394–15399.
13. Fournillier, A., I. Nakano, L. Vitvitski, E. Depla, O. Vidalin, G. Maertens, C. Tre´po, and C. Inchauspe´.1998. Modulation of immune responses to hepa-titis C virus envelope E2 protein following injection of plasmid DNA using single or combined delivery routes. Hepatology28:237–244.
14. Fournillier-Jacob, A., A. Cahour, N. Escriou, M. Girard, and C. Wychowski. 1996. Processing of the E1 glycoprotein of hepatitis C virus expressed in mammalian cells. J. Gen. Virol.77:1055–1064.
15. Fried, M. W., and J. H. Hoofnagle.1995. Therapy of hepatitis C. Semin. Liver Dis.15:82–91.
16. Gething, M.-J., and J. Sambrook.1992. Protein folding in the cell. Nature 355:33–45.
17. Grakoui, A., C. Wychowski, C. Lin, S. M. Feinstone, and C. M. Rice.1993. Expression and identification of hepatitis C virus polyprotein cleavage prod-ucts. J. Virol.67:1385–1395.
18. Habersetzer, F., A. Fournillier, J. Dubuisson, D. Rosa, S. Abrignani, C. Wychowski, I. Nakano, C. Tre´po, C. Desgranges, and G. Inchauspe´.1998. Characterization of human monoclonal antibodies specific to the hepatitis C virus glycoprotein E2 within vitrobinding neutralization properties. Virology 249:32–41.
19. Hartl, F.-U., R. Hlodan, and T. Langer.1994. Molecular chaperones in protein folding: the art of avoiding sticky situations. Trends Biochem. Sci. 19:20–25.
20. Inchauspe´, G., L. Vitvitski, M. E. Major, G. Jung, U. Spengler, M. Maison-nas, and C. Tre´po.1997. Plasmid DNA expressing a secreted or a nonse-creted form of hepatitis C virus nucleocapsid: comparative studies of anti-body and T-helper responses following genetic immunization. DNA Cell Biol.16:185–195.
21. Ishii, K., D. Rosa, Y. Watanabe, T. Katayama, H. Harada, C. Wyatt, K. Kiyosawa, H. Aizaki, Y. Matsuura, M. Houghton, S. Abrignani, and T. Miyamura.1998. High titers of antibodies inhibiting the binding of envelope to human cells correlate with natural resolution of chronic hepatitis C. Hepatology28:1117–1120.
22. Kuo, G., Q.-L. Choo, H. J. Alter, G. L. Gitnick, A. G. Redecker, R. H. Purcell, T. Miyamura, J. L. Dienstag, M. J. Alter, C. E. Stevens, G. E. Tegtmeier, F. Bonino, M. Colombo, W. S. Lee, C. Kuo, K. Berger, J. R. Shuster, L. R. Overby, D. W. Bradley, and M. Houghton.1989. An assay for circulating antibodies to a major etiologic virus of human non-A, non-B hepatitis. Science244:362–364.
23. Lee, S. W., J. H. Cho, and Y. C. Sung.1998. Optimal induction of hepatitis C virus envelope-specific immunity by bicistronic plasmid DNA inoculation with the granulocyte-macrophage colony-stimulating factor gene. J. Virol. 72:8430–8436.
23a.Maertens, G.Unpublished results. 23b.Maertens, G.Personal communication.
24. Major, M. E., L. Vivitski, M. A. Mink, M. Schleef, R. G. Whalen, C. Tre´po, and G. Inchauspe´.1995. DNA-based immunization with chimeric vectors for
the induction of immune responses against the hepatitis C virus nucleocap-sid. J. Virol.69:5798–5809.
25. Matsuura, Y., S. Harada, R. Suzuki, Y. Watanabe, Y. Inoue, I. Saito, and T. Miyamura.1992. Expression of processed envelope protein of hepatitis C virus in mammalian and insect cells. J. Virol.66:1425–1431.
26. Michalak, J.-P., C. Wychowski, A. Choukhi, J.-C. Meunier, S. Ung, C. M. Rice, and J. Dubuisson.1997. Characterization of truncated forms of hep-atitis C virus glycoproteins. J. Gen. Virol.78:2299–2306.
27. Moradpour, D., P. Kary, C. M. Rice, and H. E. Blum.1998. Continuous human cell lines inducibly expressing hepatitis C virus structural and non-structural proteins. Hepatology28:192–201.
28. Nakano, I., G. Maertens, M. E. Major, L. Vivitski, J. Dubuisson, A. Fournillier, G. de Martynoff, C. Trepo, and G. Inchauspe.1997. Immuniza-tion with plasmid DNA encoding hepatitis C virus envelope E2 antigenic domains induces antibodies whose immune reactivity is linked to the injec-tion mode. J. Virol.71:7101–7109.
29. Ralston, R., K. Thudium, K. Berger, C. Kuo, B. Gervase, J. Hall, M. Selby, G. Kuo, M. Houghton, and Q.-L. Choo.1993. Characterization of hepatitis C virus envelope glycoprotein complexes expressed by recombinant vaccinia viruses. J. Virol.67:6753–6761.
30. Raz, E., D. A. Carson, S. E. Parker, T. B. Parr, A. M. Abai, G. Aichinger, S. H. Gromkowski, M. Singh, D. Lew, M. A. Yakauckas, S. M. Bird, and G. H. Rhodes.1994. Intradermal gene immunization: the possible role of DNA uptake in the induction of cellular immunity to viruses. Proc. Natl. Acad. Sci. USA91:9519–9523.
31. Richardson, T. M., Jr., B. L. Stryjewski, C. C. Broder, J. A. Hoxie, J. R. Mascola, P. Earl, and R. W. Doms.1996. Humoral response to oligomeric human immunodeficiency virus type 1 envelope protein. J. Virol.70:753–762. 32. Rosa, D., S. Campagnoli, C. Moretto, E. Guenzi, L. C. Cousens, M. C. Dong, A. J. Weiner, J. Y. N. Lau, Q.-L. Choo, D. Chien, P. Pileri, M. Houghton, and S. Abrignani.1996. A quantitative test to estimate neutralizing antibodies to the hepatitis C virus: cytofluorimetric assessment of envelope glycoprotein 2 binding to target cells. Proc. Natl. Acad. Sci. USA93:1759–1763. 33. Saito, I., T. Miyamura, A. Ohbayashi, H. Harada, T. Katayama, S. Kikuchi,
Y. Watanabe, S. Koi, M. Onji, Y. Ohta, Q.-L. Choo, M. Houghton, and G. Kuo.1990. Hepatitis C virus infection is associated with the development of hepatocellular carcinoma. Proc. Natl. Acad. Sci. USA87:6547–6549. 34. Sanger, F., S. Nicklen, and A. R. Coulson.1977. DNA sequencing with
chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463–5467. 35. Scarselli, E., A. Cerino, G. Esposito, E. Silini, M. U. Mondelli, and C.
Traboni.1995. Occurrence of antibodies reactive with more than one variant of the putative envelope glycoprotein (gp70) hypervariable region 1 in vire-mic hepatitis C virus-infected patients. J. Virol.69:4407–4412.
36. Selby, M. J., E. Glazer, F. Masiartz, and M. Houghton.1994. Complex processing and protein:protein interactions in the E2:NS2 region of HCV. Virology204:114–122.
37. Shimizu, Y. K., H. Igarashi, T. Kiyohara, T. Cbezon, P. Farci, R. H. Purcell, and H. Yoshikura.1996. A hyperimmune serum against a synthetic peptide corresponding to the hypervariable region 1 of hepatitis C virus can prevent viral infection in cell cultures. Virology223:409–412.
38. Tighe, H., M. Corr, M. Roman, and E. Raz.1998. Gene vaccination: plasmid DNA is more than just a blueprint. Immunol. Today19:89–97.
39. Ulmer, J. B., J. J. Donnelly, S. E. Parker, G. H. Rhodes, P. L. Felgner, V. J. Dwarki, R. R. Deck, C. M. DeWitt, A. Friedman, L. A. Hawe, K. R. Leander, D. Martinez, H. C. Perry, J. W. Shiver, D. L. Montgomery, and M. A. Liu. 1993. Heterologous protection against influenza by injection of DNA encod-ing a viral protein. Science259:1745–1749.
40. Wang, B., K. E. Ugen, V. Srkantan, M. G. Agadjanyan, K. Dang, Y. Refaeli, A. I. Sato, J. Boyer, W. V. Williams, and D. B. Weiner.1993. Gene inocu-lation generates immune responses against human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA90:4156–4160.
41. Weiner, A. J., C. Christopherson, J. E. Hall, F. Bonino, G. Saracco, M. R. Brunetto, K. Crawford, C. D. Marion, K. A. Crawford, S. Venkatakrishna, T. Miyamura, J. McHutchinson, T. Cuypers, and M. Houghton.1991. Se-quence variation in hepatitis-C viral isolates. J. Hepatol.13:S6–S14. 42. Wolff, J. A., R. W. Malone, P. Williams, W. Chong, G. Acsadi, A. Jani, and
P. L. Felgner.1990. Direct gene transfer into mouse muscle in vivo. Science 247:1465–1468.
43. Zibert, A., W. Kraas, H. Meisel, G. Jung, and M. Roggendorf.1997. Epitope mapping of antibodies directed against hypervariable region 1 in acute self-limiting and chronic infections due to hepatitis C virus. J. Virol.71:4123– 4127.
44. Zibert, A., E. Schreier, and M. Roggendorf.1995. Antibodies in human sera specific to hypervariable region 1 of hepatitis C virus can block viral attach-ment. Virology208:653–661.