JOURNAL OFVIROLOGY, Dec. 2005, p. 14986–14991 Vol. 79, No. 23 0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.23.14986–14991.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Tat
28-35
SL8-Specific CD8
⫹
T Lymphocytes Are More Effective than
Gag
181-189
CM9-Specific CD8
⫹
T Lymphocytes at Suppressing
Simian Immunodeficiency Virus Replication in a
Functional In Vitro Assay
John T. Loffredo,
1Eva G. Rakasz,
1Juan Pablo Giraldo,
2Sean P. Spencer,
2Kelly K. Grafton,
1Sarah R. Martin,
2Gnankang Napoe
´,
1Levi J. Yant,
2Nancy A. Wilson,
1and David I. Watkins
1,2*
Wisconsin National Primate Research Center1and Department of Pathology and Laboratory Medicine,2University of
Wisconsin, Madison, Wisconsin 53715
Received 14 July 2005/Accepted 3 September 2005
Epitope-specific CD8ⴙT lymphocytes may play an important role in controlling human immunodeficiency
virus (HIV)/simian immunodeficiency virus replication. Unfortunately, standard cellular assays do not
mea-sure the antiviral efficacy (the ability to suppress virus replication) of CD8ⴙT lymphocytes. Certain
epitope-specific CD8ⴙT lymphocytes may be better than others at suppressing viral replication. We compared the
antiviral efficacy of two immunodominant CD8ⴙ T lymphocyte responses—Tat28-35SL8 and Gag
181-189CM9—by using a functional in vitro assay. Viral suppression by Tat-specific CD8ⴙT lymphocytes was
consistently greater than that of Gag-specific CD8ⴙ T lymphocytes. Such differences in antigen-specific
CD8ⴙ-T-lymphocyte efficacy may be important for selecting CD8ⴙT lymphocyte epitopes for inclusion in
future HIV vaccines.
Several lines of evidence suggest that CD8⫹T lymphocytes are important in suppressing human immunodeficiency virus/ simian immunodeficiency virus (HIV/SIV) replication. The ap-pearance of HIV-specific CD8⫹T lymphocytes is correlated temporally with a precipitous reduction in viral load (8, 20) implying that these virus-specific effector cells control viral replication. However, the massive loss of memory CD4⫹ T cells during acute HIV/SIV infection also likely contributes to the initial reduction in viral replication (21, 26). Additional evidence implicating CD8⫹T cells in the control of viral rep-lication comes from the depletion of circulating CD8⫹ lym-phocytes in SIV-infected macaques, which results in an in-crease in plasma viral concentrations (16, 24, 35). In addition, CD8⫹-T-lymphocytes exert selective pressure on viral se-quences in vivo, selecting for immune escape variants in both the acute (3, 9, 30) and the chronic (6, 7, 10, 11, 14, 32) phase of HIV/SIV infection.
Over the past decade, new methodologies have improved our ability to detect CD8⫹-T-lymphocyte responses against HIV/SIV. However, we still do not know which of these HIV-specific CD8⫹T lymphocytes actually contributes to control of viral replication. While neutralization assays distinguish effec-tive antibodies from ineffeceffec-tive ones, most current cellular assays rely on indirect readouts to measure CD8⫹ -T-lympho-cyte efficacy (44). Early studies demonstrated that CD8⫹cells (38) and later virus-specific cytotoxic T lymphocytes (45) in-hibited immunodeficiency virus replication in vitro. Recently, investigators using functional in vitro assays suggested that
CD8⫹-T-lymphocyte clones directed against early-expressed viral proteins, Nef (1, 46) and Rev (39, 40), may be particularly effective in suppressing viral replication. Another study dem-onstrated effective viral suppression using Pol-specific CD8⫹T lymphocytes (37). Furthermore, dendritic cells pulsed with in-activated autologous virus can expand virus-specific CD8⫹T cells, which are then capable of controlling HIV replication (23). While most data suggest that there are differences among the various CD8⫹-T-lymphocyte populations in their antiviral efficacy (the ability to suppress virus replication), current stud-ies are limited by relying on a small number of well-defined clones. Moreover, the attributes of an effective CD8⫹ -T-lym-phocyte response are still undefined.
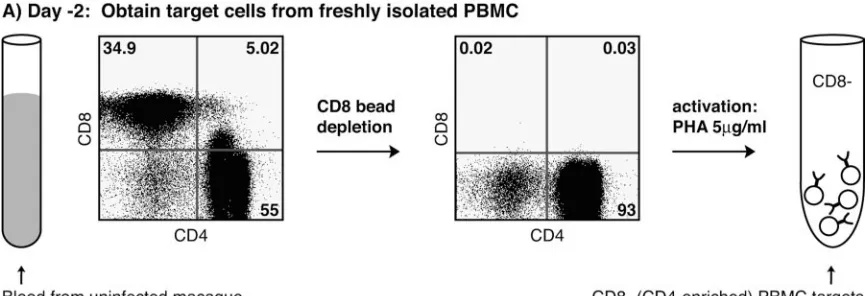
Development of the viral suppression assay (VSA).To
bet-ter address questions of antiviral efficacy, we developed a func-tional in vitro viral suppression assay to assess the ability of CD8⫹T lymphocytes to control SIV replication. We depleted uninfected Indian rhesus macaque (Macaca mulatta) PBMC of CD8⫹cells using CD8 nonhuman primate microbeads on an AutoMACS bead separation unit (Miltenyi, Auburn, CA) ac-cording to the manufacturer’s protocol. Depletions were
⬎99% effective (Fig. 1A). We stimulated the CD8⫺fraction with 5g/ml of phytohemagglutinin (Sigma, St. Louis, MO) for 18 to 24 h. The CD8⫺ targets were then incubated with SIVmac239 (17) at a multiplicity of infection of 5⫻10⫺5for
4 h. This multiplicity of infection reproducibly infected the CD8⫺peripheral blood mononuclear cells (PBMC) target cells and provided exponential SIV replication during the initial days of the assay.
We used in vitro-stimulated epitope-specific CD8⫹-T-cell lines as effector cells. Cell lines were generated from fresh or CD8-enriched PBMC from SIVmac239-infectedMamu-A*01⫹
* Corresponding author. Mailing address: Department of Pathology and Laboratory Medicine, University of Wisconsin-Madison, 585 Sci-ence Dr., Madison, WI 53711. Phone: (608) 265-3380. Fax: (608) 265-8084. E-mail: [email protected].
14986
on November 8, 2019 by guest
http://jvi.asm.org/
macaques in the chronic phase by using previously described methods (42). Intracellular cytokine staining assays verified that the in vitro cultures produced gamma interferon and tu-mor necrosis factor alpha in response to their cognate antigen and were not functionally impaired as has been observed in vivo (5, 13, 41).
We performed Mamu-A*01 tetramer stains (2) to measure epitope specificity. Typically, after 1 to 3 months, CD8⫹-T-cell lines reached a suitable specificity (⬎50%) for cell sorting. We sorted CD3⫹, CD8⫹, and Mamu-A*01 tetramer⫹lymphocytes using a MoFlo cell sorter (DakoCytomation, Fort Collins,
CO). Post-sort analysis was performed to assess purity (typi-cally⬎99%) and viability as assessed by trypan blue exclusion (Gibco, Grand Island, NY) (Fig. 1B). In cases where a cell line was⬎94% tetramer-positive CD8⫹T lymphocytes but in lim-ited quantities, we used unsorted cells after verifying that other immunodominant CD8⫹-T-lymphocyte responses were absent. We added 5.0⫻105infected CD8⫺lymphocytes (targets) to
each well of a 24-well plate (Fig. 1C). Sorted CD8⫹T cells (effectors) were added to wells at effector to target ratios (E:T) of 1:10 and 1:20. We chose these E:Ts because early testing indicated E:Ts above 1:10 would sometimes provide viral sup-FIG. 1. VSA schematic. (A) Target cells are freshly isolated PBMC, depleted of CD8⫹cells, and activated with phytohemagglutinin (PHA). (B) Effector cells are SIV-specific in vitro-stimulated CD8⫹T cells that are sorted to high specificity. (C) After a 4-h incubation of the target cells with SIVmac239, we combined target and effector cells at E:T ratios of 1:10, and 1:20. The cocultures were maintained for 8 to 11 days. 0.5 ml of
supernatant was taken every 2 days for vRNA quantification. MOI, multiplicity of infection.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:2.585.75.509.70.218.2]pression in our major histocompatibility complex (MHC)-mis-matched controls, whereas ratios below 1:20 would not provide consistent levels of viral suppression. The final volume of cell culture medium was 2 ml and contained 50 U of interleukin-2 (NIH AIDS Research and Reference Reagent Program, Ger-mantown, MD)/ml. The cocultures were maintained for 8 to 11
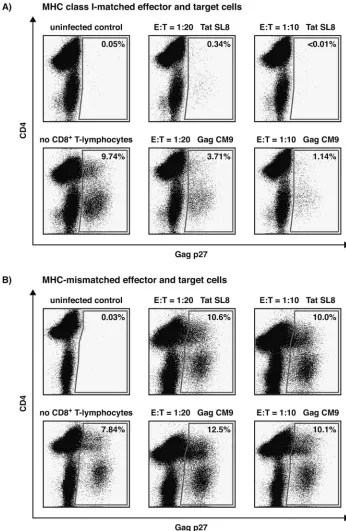
[image:3.585.116.462.69.600.2]days. Every 2 days, 0.5 ml of supernatant was collected and replaced with fresh medium. We purified vRNA from 400l of supernatant using the MagAttract vRNA minikit on a BioRo-bot M48 workstation (QIAGEN, Valencia, CA). Viral loads were determined by quantitative reverse transcription-PCR on a LightCycler 2.0 (Roche Diagnostics, Indianapolis, IN) as FIG. 2. Day 8 intracellular Gag p27 staining of representative VSA results with Tat28-35SL8- and Gag181-189CM9-specific CD8⫹T-lymphocytes
from a single macaque (animal 2125). Results for MHC class I-matched effector and target cells (A) and MHC-mismatched effector ( Mamu-A*01⫹) and target (Mamu-A*01⫺) cells (B) at an E:T of 1:10 are shown. Viral suppression only occurred in an MHC class I-restricted fashion.
14988 NOTES J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
previously described (27). At the end of a VSA, intracellular Gag p27 staining was performed on the coculture to measure SIVmac239 infection. We first stained for the cell surface
mark-ers CD3, CD4, and CD8, followed by intracellular Gag p27 staining using Fix and Perm (CALTAG, Burlingame, CA) ac-cording to the manufacturer’s protocol with fluorescently con-jugated 55-2F12 Gag p27 antibody (NIH AIDS Research and Reference Reagent Program, Germantown, MD) at⬃0.75 mg/ ml.
Differences in the antiviral efficacy of Tat28-35SL8- and
Gag181-189CM9-specific CD8ⴙT lymphocytes.Tat28-35SL8 and
Gag181-189CM9 are Mamu-A*01-restricted immunodominant
epitopes (6, 29, 31) often studied in macaque vaccine experi-ments (4, 6, 7, 15, 36, 43). These two responses account for
⬎50% of the total SIV-specific CD8⫹ responses in Mamu-A*01⫹macaques (29). Despite extensive studies, the antiviral efficacies of these two high frequency CD8⫹-T-lymphocyte re-sponses have not been addressed. We used the VSA to com-pare the ability of Tat28-35SL8- and Gag181-189CM9-specific
CD8⫹T lymphocytes to suppress SIVmac239 replication.
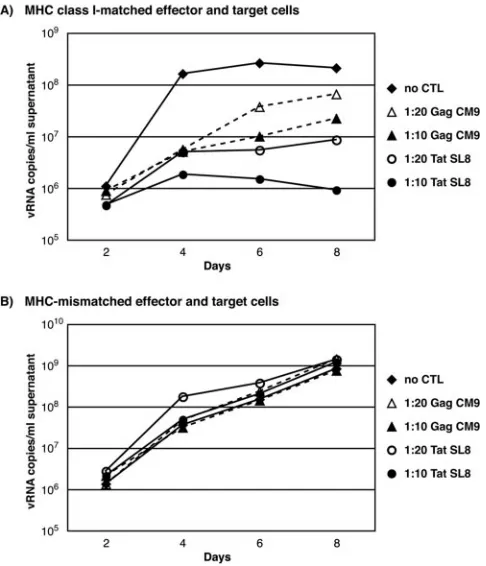
Both cellular responses diminished viral replication at E:Ts of 1:10 and 1:20 as determined by intracellular Gag p27 stain-ing (Fig. 2A) and quantitative reverse transcription-PCR (Fig. 3A) compared to wells containing only target cells. To ensure viral suppression was MHC class I-restricted, we combined
Mamu-A*01⫺target cells with Mamu-A*01-restricted effectors
as controls in each assay. Using MHC-mismatched effector and target cells, we observed negligible reduction in SIV replica-tion compared to wells that contain targets only in the absence of effective CD8⫹T cells (Fig. 2B and 3B).
At E:Ts of 1:10 and 1:20, the Tat28-35SL8-specific CD8⫹T
lymphocytes suppressed viral replication better than CD8⫹T lymphocytes against Gag181-189CM9 did. By day 8, at an E:T of
1:10, the Tat28-35SL8-specific CD8⫹T lymphocytes effectively
suppressed viral replication, reducing the number of Gag p27-positive cells to below background (Fig. 2A). Viral RNA con-centrations in these wells were 240-fold lower than control wells (Fig. 3A). These results were consistent in at least two independent assays with CD8⫹-T-cell lines derived from the same animal (data not shown).
Next, we examined whether suppression was animal- and/or epitope-specific using CD8⫹-T-cell lines generated from sev-eral Mamu-A*01⫹ SIV-infected macaques. In all instances, Tat-specific CD8⫹T lymphocytes suppressed viral replication more than Gag-specific CD8⫹ T lymphocytes did (Fig. 4). Although animal 2095 had the least effective Tat28-35
SL8-spe-cific CD8⫹T lymphocytes, there was still a 10-fold decrease in intracellular Gag p27 staining and a 47-fold decrease in viral FIG. 3. Quantitative PCR of representative VSA results with
Tat28-35SL8- and Gag181-189CM9-specific CD8⫹T lymphocytes from
macaque 2125. Results for MHC class I-matched effector and target cells (A) MHC-mismatched effector (Mamu-A*01⫹) and target (Mamu-A*01⫺) cells (B) at an E:T of 1:10 are shown. Viral
suppres-sion only occurred in an MHC class I-restricted fashion. FIG. 4. Day 8 viral suppression by Tat28-35SL8- and Gag181-189
CM9-specific CD8⫹T lymphocytes derived from several SIV-infected ma-caques (animal identification number shown below bars) at an E:T ratio of 1:10 as measured by fold reduction of intracellular Gag p27 (A) and fold reduction of vRNA copies/ml (B). Bars:■, Tat-specific cells;䊐, Gag-specific cells.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.43.286.73.356.2]RNA concentration on day 8 relative to wells without effector cells (Fig. 4). Our results displayed similar trends over time in multiple independent assays (data not shown).
While Tat-specific CD8⫹ T lymphocytes cells were more effective in their ability to suppress SIV replication, we ob-served some animal-to-animal variability (Fig. 4). This variabil-ity may reflect differences in the immunological and virological status of animals at the time of cell line generation. For in-stance, studies correlate the impairment of CD8⫹-T-cell func-tion and proliferative capacity with the loss of CD4⫹T cells (22, 25, 28). Animal 2095 had low CD4 counts (⬍200 cells/l) when PBMC were obtained to generate the Tat28-35
SL8-spe-cific CD8⫹-T-cell line. Interestingly, this animal produced the least effective Tat-specific cell line (Fig. 4).
While epitope-specific CD8⫹T-lymphocytes can inhibit viral replication by cytolytic and noncytolytic mechanisms, the re-duction in SIV replication by our epitope-specific CD8⫹ T lymphocytes is likely occurring by cytolytic activity in a MHC class I-restricted fashion. Previous findings demonstrated that epitope-specific CD8⫹-T-lymphocytes suppressed viral replica-tion primarily through direct cytolytic activity (12, 45). Al-though it appears that noncytolytic effects may play a role, it is likely minor and contingent on direct cell-to-cell interactions via MHC-TCR recognition. Although these studies demon-strated that soluble factors inhibit HIV/SIV replication in a MHC class I-unrestricted fashion (12, 45), we observed mini-mal, if any, viral suppression when the effector and target cells were MHC mismatched (Fig. 2B and 3B). Nevertheless, we cannot rule out the involvement of noncytolytic activity in our assay.
When performing fluorescence-activated cell sorting analy-sis at the end of the assay, we found that the percentages of effector cells directed against Tat28-35SL8 and Gag181-189CM9
in the cocultures were comparable in most cases. There were some instances in which Gag181-189CM9-specific CD8⫹T
lym-phocytes persisted in two- to fourfold greater numbers than those against Tat28-35SL8. However, even in these cases, the
greater abundance of Gag181-189CM9-specific CD8⫹ T
lym-phocytes did not suppress of SIVmac239 replication as well as
Tat28-35SL8-specific CD8⫹T lymphocytes. This suggests that a
difference in antiviral efficacy is not simply due to lymphocyte longevity.
Previous studies have failed to address the antiviral efficacy of Tat28-35SL8- and Gag181-189CM9-specific CD8⫹
T-lympho-cytes, although they have suggested possible reasons for the observed efficacy of the Tat-specific CD8⫹T cells. Tat-specific mRNA is detected as early as 4 h after HIV infection, whereas Gag-specific mRNA appears approximately 20 h postinfection (18, 19, 33, 34). In addition, Tat28-35SL8-specific cells appear to
have a higher functional avidity (⬃0.18 nM) compared to that of the Gag181-189CM9-specific cells (⬃13.3 nM) (30). These
differences might enable the Tat-specific CD8⫹T cells to rec-ognize cells early in the virus replication cycle and those that present only small amounts of antigen.
Our viral suppression assay measures the ability of antigen-specific CD8⫹T lymphocytes to suppress SIV replication in vitro. We used this assay to show that Tat28-35SL8-specific
CD8⫹T lymphocytes are consistently more effective at sup-pressing SIVmac239 replication than Gag181-189CM9-specific
CD8⫹ T lymphocytes. Using this assay, we hope to define
additional potentially protective CD8⫹-T-lymphocyte re-sponses. It is plausible that findings from our functional in vitro assay can be extrapolated to the ability to suppress HIV/SIV replication in vivo. Therefore, differences in CD8⫹ -T-lympho-cyte suppressive efficacy may be critically important when choosing epitopes for future HIV vaccines.
We thank William Rehrauer, Jess Maxwell, and Tim Jacoby for MHC class I PCR-SSP typing and gratefully acknowledge Laura Val-entine, Alex Ko, and Andrea Weiler for immunological assay assis-tance. David O’Connor and Thomas Friedrich provided helpful dis-cussions. We also thank the Immunology and Virology Core Laboratories at the National Primate Research Center, University of Wisconsin-Madison for technical assistance.
This research was supported by National Institutes of Health grants R01 AI049120-04 to D.I.W. and P51 RR 000167 to the Wisconsin National Primate Research Center (WNPRC). This study was con-ducted in part at a facility constructed with support from Research Facilities Improvement grant numbers RR15459-01 and RR020141-01 (WNPRC).
REFERENCES
1.Ali, A., R. Lubong, H. Ng, D. G. Brooks, J. A. Zack, and O. O. Yang.2004. Impacts of epitope expression kinetics and class I downregulation on the antiviral activity of human immunodeficiency virus type 1-specific cytotoxic T lymphocytes. J. Virol.78:561–567.
2.Allen, T. M., B. R. Mothe, J. Sidney, P. Jing, J. L. Dzuris, M. E. Liebl, T. U. Vogel, D. H. O’Connor, X. Wang, M. C. Wussow, J. A. Thomson, J. D. Altman, D. I. Watkins, and A. Sette.2001. CD8⫹lymphocytes from simian immunodeficiency virus-infected rhesus macaques recognize 14 different epitopes bound by the major histocompatibility complex class I molecule mamu-A*01: implications for vaccine design and testing. J. Virol.75:738–749. 3.Allen, T. M., D. H. O’Connor, P. Jing, J. L. Dzuris, B. R. Mothe, T. U. Vogel,
E. Dunphy, M. E. Liebl, C. Emerson, N. Wilson, K. J. Kunstman, X. Wang, D. B. Allison, A. L. Hughes, R. C. Desrosiers, J. D. Altman, S. M. Wolinsky, A. Sette, and D. I. Watkins.2000. Tat-specific cytotoxic T lymphocytes select for SIV escape variants during resolution of primary viraemia. Nature407:
386–390.
4.Amara, R. R., F. Villinger, J. D. Altman, S. L. Lydy, S. P. O’Neil, S. I. Staprans, D. C. Montefiori, Y. Xu, J. G. Herndon, L. S. Wyatt, M. A. Candido, N. L. Kozyr, P. L. Earl, J. M. Smith, H. L. Ma, B. D. Grimm, M. L. Hulsey, J. Miller, H. M. McClure, J. M. McNicholl, B. Moss, and H. L. Robinson.2001. Control of a mucosal challenge and prevention of AIDS by a multiprotein DNA/MVA vaccine. Science292:69–74.
5.Appay, V., D. F. Nixon, S. M. Donahoe, G. M. Gillespie, T. Dong, A. King, G. S. Ogg, H. M. Spiegel, C. Conlon, C. A. Spina, D. V. Havlir, D. D. Richman, A. Waters, P. Easterbrook, A. J. McMichael, and S. L. Rowland-Jones.2000. HIV-specific CD8⫹T cells produce antiviral cytokines but are impaired in cytolytic function. J. Exp. Med.192:63–75.
6.Barouch, D. H., J. Kunstman, J. Glowczwskie, K. J. Kunstman, M. A. Egan, F. W. Peyerl, S. Santra, M. J. Kuroda, J. E. Schmitz, K. Beaudry, G. R. Krivulka, M. A. Lifton, D. A. Gorgone, S. M. Wolinsky, and N. L. Letvin.
2003. Viral escape from dominant simian immunodeficiency virus epitope-specific cytotoxic T lymphocytes in DNA-vaccinated rhesus monkeys. J. Vi-rol.77:7367–7375.
7.Barouch, D. H., J. Kunstman, M. J. Kuroda, J. E. Schmitz, S. Santra, F. W. Peyerl, G. R. Krivulka, K. Beaudry, M. A. Lifton, D. A. Gorgone, D. C. Montefiori, M. G. Lewis, S. M. Wolinsky, and N. L. Letvin.2002. Eventual AIDS vaccine failure in a rhesus monkey by viral escape from cytotoxic T lymphocytes. Nature415:335–339.
8.Borrow, P., H. Lewicki, B. H. Hahn, G. M. Shaw, and M. B. Oldstone.1994. Virus-specific CD8⫹cytotoxic T-lymphocyte activity associated with control of viremia in primary human immunodeficiency virus type 1 infection. J. Vi-rol.68:6103–6110.
9.Borrow, P., H. Lewicki, X. Wei, M. S. Horwitz, N. Peffer, H. Meyers, J. A. Nelson, J. E. Gairin, B. H. Hahn, M. B. Oldstone, and G. M. Shaw.1997. Antiviral pressure exerted by HIV-1-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid selection of CTL escape virus. Nat. Med.3:205–211.
10.Chen, Z. W., A. Craiu, L. Shen, M. J. Kuroda, U. C. Iroku, D. I. Watkins, G. Voss, and N. L. Letvin.2000. Simian immunodeficiency virus evades a dom-inant epitope-specific cytotoxic T lymphocyte response through a mutation resulting in the accelerated dissociation of viral peptide and MHC class I. J. Immunol.164:6474–6479.
11.Evans, D. T., D. H. O’Connor, P. Jing, J. L. Dzuris, J. Sidney, J. da Silva, T. M. Allen, H. Horton, J. E. Venham, R. A. Rudersdorf, T. Vogel, C. D. Pauza, R. E. Bontrop, R. DeMars, A. Sette, A. L. Hughes, and D. I. Watkins.
1999. Virus-specific cytotoxic T-lymphocyte responses select for amino-acid
14990 NOTES J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
variation in simian immunodeficiency virus Env and Nef. Nat. Med.5:1270– 1276.
12.Gauduin, M. C., R. L. Glickman, R. Means, and R. P. Johnson.1998. Inhibition of simian immunodeficiency virus (SIV) replication by CD8⫹T lymphocytes from macaques immunized with live attenuated SIV. J. Virol.
72:6315–6324.
13.Goepfert, P. A., A. Bansal, B. H. Edwards, G. D. Ritter, Jr., I. Tellez, S. A. McPherson, S. Sabbaj, and M. J. Mulligan.2000. A significant number of human immunodeficiency virus epitope-specific cytotoxic T lymphocytes de-tected by tetramer binding do not produce gamma interferon. J. Virol.
74:10249–10255.
14.Goulder, P. J., R. E. Phillips, R. A. Colbert, S. McAdam, G. Ogg, M. A. Nowak, P. Giangrande, G. Luzzi, B. Morgan, A. Edwards, A. J. McMichael, and S. Rowland-Jones.1997. Late escape from an immunodominant cyto-toxic T-lymphocyte response associated with progression to AIDS. Nat. Med.
3:212–217.
15.Horton, H., T. U. Vogel, D. K. Carter, K. Vielhuber, D. H. Fuller, T. Shipley, J. T. Fuller, K. J. Kunstman, G. Sutter, D. C. Montefiori, V. Erfle, R. C. Desrosiers, N. Wilson, L. J. Picker, S. M. Wolinsky, C. Wang, D. B. Allison, and D. I. Watkins.2002. Immunization of rhesus macaques with a DNA prime/modified vaccinia virus Ankara boost regimen induces broad simian immunodeficiency virus (SIV)-specific T-cell responses and reduces initial viral replication but does not prevent disease progression following challenge with pathogenic SIVmac239. J. Virol.76:7187–7202.
16.Jin, X., D. E. Bauer, S. E. Tuttleton, S. Lewin, A. Gettie, J. Blanchard, C. E. Irwin, J. T. Safrit, J. Mittler, L. Weinberger, L. G. Kostrikis, L. Zhang, A. S. Perelson, and D. D. Ho.1999. Dramatic rise in plasma viremia after CD8⫹ T cell depletion in simian immunodeficiency virus-infected macaques. J. Exp. Med.189:991–998.
17.Kestler, H., T. Kodama, D. Ringler, M. Marthas, N. Pedersen, A. Lackner, D. Regier, P. Sehgal, M. Daniel, N. King, et al.1990. Induction of AIDS in rhesus monkeys by molecularly cloned simian immunodeficiency virus. Sci-ence248:1109–1112.
18.Kim, S. Y., R. Byrn, J. Groopman, and D. Baltimore.1989. Temporal aspects of DNA and RNA synthesis during human immunodeficiency virus infection: evidence for differential gene expression. J. Virol.63:3708–3713. 19.Klotman, M. E., S. Kim, A. Buchbinder, A. DeRossi, D. Baltimore, and F.
Wong-Staal.1991. Kinetics of expression of multiply spliced RNA in early human immunodeficiency virus type 1 infection of lymphocytes and mono-cytes. Proc. Natl. Acad. Sci. USA88:5011–5015.
20.Koup, R. A., J. T. Safrit, Y. Cao, C. A. Andrews, G. McLeod, W. Borkowsky, C. Farthing, and D. D. Ho.1994. Temporal association of cellular immune responses with the initial control of viremia in primary human immunode-ficiency virus type 1 syndrome. J. Virol.68:4650–4655.
21.Li, Q., L. Duan, J. D. Estes, Z. M. Ma, T. Rourke, Y. Wang, C. Reilly, J. Carlis, C. J. Miller, and A. T. Haase.2005. Peak SIV replication in resting memory CD4⫹T cells depletes gut lamina propria CD4⫹T cells. Nature
434:1148–1152.
22.Lichterfeld, M., D. E. Kaufmann, X. G. Yu, S. K. Mui, M. M. Addo, M. N. Johnston, D. Cohen, G. K. Robbins, E. Pae, G. Alter, A. Wurcel, D. Stone, E. S. Rosenberg, B. D. Walker, and M. Altfeld.2004. Loss of HIV-1-specific CD8⫹T-cell proliferation after acute HIV-1 infection and restoration by vaccine-induced HIV-1-specific CD4⫹T cells. J. Exp. Med.200:701–712. 23.Lu, W., and J. M. Andrieu.2001. In vitro human immunodeficiency virus
eradication by autologous CD8⫹T cells expanded with inactivated-virus-pulsed dendritic cells. J. Virol.75:8949–8956.
24.Matano, T., R. Shibata, C. Siemon, M. Connors, H. C. Lane, and M. A. Martin.1998. Administration of an anti-CD8 monoclonal antibody inter-feres with the clearance of chimeric simian/human immunodeficiency virus during primary infections of rhesus macaques. J. Virol.72:164–169. 25.Matloubian, M., R. J. Concepcion, and R. Ahmed.1994. CD4⫹T cells are
required to sustain CD8⫹cytotoxic T-cell responses during chronic viral infection. J. Virol.68:8056–8063.
26.Mattapallil, J. J., D. C. Douek, B. Hill, Y. Nishimura, M. Martin, and M. Roederer.2005. Massive infection and loss of memory CD4⫹T cells in multiple tissues during acute SIV infection. Nature434:1093–1097. 27.McDermott, A. B., J. Mitchen, S. Piaskowski, I. De Souza, L. J. Yant, J.
Stephany, J. Furlott, and D. I. Watkins.2004. Repeated low-dose mucosal simian immunodeficiency virus SIVmac239 challenge results in the same viral and immunological kinetics as high-dose challenge: a model for the evaluation of vaccine efficacy in nonhuman primates. J. Virol.78:3140–3144. 28.Migueles, S. A., A. C. Laborico, W. L. Shupert, M. S. Sabbaghian, R. Rabin, C. W. Hallahan, D. Van Baarle, S. Kostense, F. Miedema, M. McLaughlin, L. Ehler, J. Metcalf, S. Liu, and M. Connors.2002. HIV-specific CD8⫹T cell proliferation is coupled to perforin expression and is maintained in nonpro-gressors. Nat. Immunol.3:1061–1068.
29.Mothe, B. R., H. Horton, D. K. Carter, T. M. Allen, M. E. Liebl, P. Skinner, T. U. Vogel, S. Fuenger, K. Vielhuber, W. Rehrauer, N. Wilson, G. Franchini, J. D. Altman, A. Haase, L. J. Picker, D. B. Allison, and D. I. Watkins.2002. Dominance of CD8 responses specific for epitopes bound by a single major histocompatibility complex class I molecule during the acute phase of viral infection. J. Virol.76:875–884.
30.O’Connor, D. H., T. M. Allen, T. U. Vogel, P. Jing, I. P. DeSouza, E. Dodds, E. J. Dunphy, C. Melsaether, B. Mothe, H. Yamamoto, H. Horton, N. Wilson, A. L. Hughes, and D. I. Watkins.2002. Acute phase cytotoxic T lymphocyte escape is a hallmark of simian immunodeficiency virus infection. Nat. Med.
8:493–499.
31.O’Connor, D. H., B. R. Mothe, J. T. Weinfurter, S. Fuenger, W. M. Rehrauer, P. Jing, R. R. Rudersdorf, M. E. Liebl, K. Krebs, J. Vasquez, E. Dodds, J. Loffredo, S. Martin, A. B. McDermott, T. M. Allen, C. Wang, G. G. Doxiadis, D. C. Montefiori, A. Hughes, D. R. Burton, D. B. Allison, S. M. Wolinsky, R. Bontrop, L. J. Picker, and D. I. Watkins.2003. Major histocompatibility complex class I alleles associated with slow simian immunodeficiency virus disease progression bind epitopes recognized by dominant acute-phase cy-totoxic-T-lymphocyte responses. J. Virol.77:9029–9040.
32.Phillips, R. E., S. Rowland-Jones, D. F. Nixon, F. M. Gotch, J. P. Edwards, A. O. Ogunlesi, J. G. Elvin, J. A. Rothbard, C. R. Bangham, C. R. Rizza, and A. J. McMichael.1991. Human immunodeficiency virus genetic variation that can escape cytotoxic T cell recognition. Nature354:453–459. 33.Pomerantz, R. J., D. Trono, M. B. Feinberg, and D. Baltimore.1990. Cells
nonproductively infected with HIV-1 exhibit an aberrant pattern of viral RNA expression: a molecular model for latency. Cell61:1271–1276. 34.Ranki, A., A. Lagerstedt, V. Ovod, E. Aavik, and K. J. Krohn.1994.
Expres-sion kinetics and subcellular localization of HIV-1 regulatory proteins Nef, Tat, and Rev in acutely and chronically infected lymphoid cell lines. Arch. Virol.139:365–378.
35.Schmitz, J. E., M. J. Kuroda, S. Santra, V. G. Sasseville, M. A. Simon, M. A. Lifton, P. Racz, K. Tenner-Racz, M. Dalesandro, B. J. Scallon, J. Ghrayeb, M. A. Forman, D. C. Montefiori, E. P. Rieber, N. L. Letvin, and K. A. Reimann.1999. Control of viremia in simian immunodeficiency virus infec-tion by CD8⫹lymphocytes. Science283:857–860.
36.Shiver, J. W., T. M. Fu, L. Chen, D. R. Casimiro, M. E. Davies, R. K. Evans, Z. Q. Zhang, A. J. Simon, W. L. Trigona, S. A. Dubey, L. Huang, V. A. Harris, R. S. Long, X. Liang, L. Handt, W. A. Schleif, L. Zhu, D. C. Freed, N. V. Persaud, L. Guan, K. S. Punt, A. Tang, M. Chen, K. A. Wilson, K. B. Collins, G. J. Heidecker, V. R. Fernandez, H. C. Perry, J. G. Joyce, K. M. Grimm, J. C. Cook, P. M. Keller, D. S. Kresock, H. Mach, R. D. Troutman, L. A. Isopi, D. M. Williams, Z. Xu, K. E. Bohannon, D. B. Volkin, D. C. Monte-fiori, A. Miura, G. R. Krivulka, M. A. Lifton, M. J. Kuroda, J. E. Schmitz, N. L. Letvin, M. J. Caulfield, A. J. Bett, R. Youil, D. C. Kaslow, and E. A. Emini.2002. Replication-incompetent adenoviral vaccine vector elicits ef-fective anti-immunodeficiency-virus immunity. Nature415:331–335. 37.Tomiyama, H., M. Fujiwara, S. Oka, and M. Takiguchi.2005. Cutting edge:
epitope-dependent effect of Nef-mediated HLA class I down-regulation on ability of HIV-1-specific CTLs to suppress HIV-1 replication. J. Immunol.
174:36–40.
38.Tsubota, H., C. I. Lord, D. I. Watkins, C. Morimoto, and N. L. Letvin.1989. A cytotoxic T lymphocyte inhibits acquired immunodeficiency syndrome virus replication in peripheral blood lymphocytes. J. Exp. Med.169:1421–1434. 39.van Baalen, C. A., C. Guillon, M. van Baalen, E. J. Verschuren, P. H. Boers, A. D. Osterhaus, and R. A. Gruters.2002. Impact of antigen expression kinetics on the effectiveness of HIV-specific cytotoxic T lymphocytes. Eur. J. Immunol.32:2644–2652.
40.Van Baalen, C. A., M. Schutten, R. C. Huisman, P. H. Boers, R. A. Gruters, and A. D. Osterhaus.1998. Kinetics of antiviral activity by human immuno-deficiency virus type 1-specific cytotoxic T lymphocytes (CTL) and rapid selection of CTL escape virus in vitro. J. Virol.72:6851–6857.
41.Vogel, T. U., T. M. Allen, J. D. Altman, and D. I. Watkins.2001. Functional impairment of simian immunodeficiency virus-specific CD8⫹T cells during the chronic phase of infection. J. Virol.75:2458–2461.
42.Vogel, T. U., T. C. Friedrich, D. H. O’Connor, W. Rehrauer, E. J. Dodds, H. Hickman, W. Hildebrand, J. Sidney, A. Sette, A. Hughes, H. Horton, K. Vielhuber, R. Rudersdorf, I. P. De Souza, M. R. Reynolds, T. M. Allen, N. Wilson, and D. I. Watkins.2002. Escape in one of two cytotoxic T-lympho-cyte epitopes bound by a high-frequency major histocompatibility complex class I molecule, Mamu-A*02: a paradigm for virus evolution and persis-tence? J. Virol.76:11623–11636.
43.Vogel, T. U., M. R. Reynolds, D. H. Fuller, K. Vielhuber, T. Shipley, J. T. Fuller, K. J. Kunstman, G. Sutter, M. L. Marthas, V. Erfle, S. M. Wolinsky, C. Wang, D. B. Allison, E. W. Rud, N. Wilson, D. Montefiori, J. D. Altman, and D. I. Watkins.2003. Multispecific vaccine-induced mucosal cytotoxic T lymphocytes reduce acute-phase viral replication but fail in long-term con-trol of simian immunodeficiency virus SIVmac239. J. Virol.77:13348–13360. 44.Yang, O. O.2003. Will we be able to “spot” an effective HIV-1 vaccine?
Trends Immunol.24:67–72.
45.Yang, O. O., S. A. Kalams, A. Trocha, H. Cao, A. Luster, R. P. Johnson, and B. D. Walker.1997. Suppression of human immunodeficiency virus type 1 replication by CD8⫹cells: evidence for HLA class I-restricted triggering of cytolytic and noncytolytic mechanisms. J. Virol.71:3120–3128.
46.Yang, O. O., P. T. Sarkis, A. Trocha, S. A. Kalams, R. P. Johnson, and B. D. Walker.2003. Impacts of avidity and specificity on the antiviral efficiency of HIV-1-specific CTL. J. Immunol.171:3718–3724.