0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.12.7812–7818.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Plant Virus-Derived Small Interfering RNAs Originate Predominantly
from Highly Structured Single-Stranded Viral RNAs†
Attila Molna
´r,
1‡ Tibor Csorba,
1Lo
´ra
´nt Lakatos,
1E
´ va Va´rallyay,
1Christophe Lacomme,
2and Jo
´zsef Burgya
´n
1*
Agricultural Biotechnology Center, Plant Biology Institute, P. O. Box 411, H-2101 Go¨do¨llo¨, Hungary,1and Scottish Crop
Research Institute, Invergowrie, Dundee DD2 5DA, United Kingdom2
Received 8 November 2004/Accepted 29 January 2005
RNA silencing is conserved in a broad range of eukaryotes and includes the phenomena of RNA interference in animals and posttranscriptional gene silencing (PTGS) in plants. In plants, PTGS acts as an antiviral system; a successful virus infection requires suppression or evasion of the induced silencing response. Small interfering RNAs (siRNAs) accumulate in plants infected with positive-strand RNA viruses and provide specificity to this RNA-mediated defense. We present here the results of a survey of virus-specific siRNAs characterized by a sequence analysis of siRNAs from plants infected with Cymbidium ringspot tombusvirus
(CymRSV). CymRSV siRNA sequences have a nonrandom distribution along the length of the viral genome, suggesting that there are hot spots for virus-derived siRNA generation. CymRSV siRNAs bound to the CymRSV p19 suppressor protein have the same asymmetry in strand polarity as the sequenced siRNAs and are imperfect double-stranded RNA duplexes. Moreover, an analysis of siRNAs derived from two other nonrelated positive-strand RNA viruses showed that they displayed the same asymmetry as CymRSV siRNAs. Finally, we show thatTobacco mosaic virus(TMV) carrying a short inverted repeat of the phytoene desaturase (PDS) gene triggered more accumulation ofPDSsiRNAs than the corresponding antisensePDSsequence. Taken together, these results suggest that virus-derived siRNAs originate predominantly by direct DICER cleavage of imperfect duplexes in the most folded regions of the positive strand of the viral RNA.
Eukaryotic organisms have developed a highly adaptable and specific mechanism to protect their genomes against ab-errant endogenous or exogenous RNA molecules. This phe-nomenon, referred to as RNA silencing, is an ancient defense mechanism induced by double-stranded RNAs (dsRNAs) that leads to homology-dependent degradation of target RNAs. RNA silencing is conserved across kingdoms and is manifested as quelling in fungi, RNA interference (RNAi) in animals, and cosuppression or posttranscriptional gene silencing (PTGS) in plants. The unifying feature of RNA silencing is the presence of 21- to 26-nucleotide (nt) small interfering RNAs (siRNAs) (17, 18, 40). In addition, biochemical and genetic analyses have shown that the core mechanisms of RNA silencing are shared among different eukaryotes (5, 20, 32, 40, 55, 60). RNA silenc-ing is induced by dsRNAs or structured ssilenc-ingle-stranded RNAs (ssRNAs) that are processed into siRNAs by RNase III-like enzymes such as DICER (6, 37). siRNAs guide the sequence-specific degradation of target mRNAs by the RNA-induced silencing complex (RISC) (19). The RISC mediates the cleav-age of a target mRNA when there is perfect or nearly perfect base pairing between the mRNA and a short guide RNA and mediates translation repression when there is partial comple-mentarity (3, 10, 13, 23). siRNAs can also guide another
ef-fector complex, namely, the RNA-induced initiation of tran-scriptional gene silencing (RITS) complex, to direct the chromatin modification of homologous DNA sequences (53). In addition to siRNAs, there are other short regulatory RNAs in the RNA silencing machinery called micro-RNAs (miRNAs), which are the products of endogenous noncoding genes. Mature miRNAs are derived by DICER-mediated cleavage from an inverted-repeat mRNA precursor harboring short dsRNA stem-loops (2, 7, 25, 26, 28, 35, 43). Importantly, miRNAs are encoded by genes distinct from those encoding the mRNAs whose expression they control, while siRNAs are generally derived from the same RNAs that are targeted by RNA silencing. A third class of small RNAs, the so-called
trans-acting siRNAs, are distinct from miRNAs in that they are
likely derived from long dsRNAs produced by an RNA-depen-dent RNA polymerase; they also appear to be different from exogenous siRNAs, as they primarily repress the expression of other genes rather than their own expression (39, 52).
Regardless of their origins, the occurrence of dsRNAs in the cytoplasm of plant cells induces PTGS. It has been demon-strated that RNA-dependent RNA polymerase is also involved in PTGS (11, 36), presumably by converting target ssRNAs into dsRNAs, which are then processed by DICER to generate siRNAs (58). Remarkably, PTGS can generate mobile silenc-ing signals with sequence-specific information that spread from cell to cell through plasmodesmata (short-distance movement) and systemically through the vascular system (phloem-medi-ated long-distance movement) to different organs of the plant (21, 33).
An important characteristic of RNA silencing in plants is the involvement of two functionally distinct siRNA species that * Corresponding author. Mailing address: Agricultural
Biotechnol-ogy Center, Plant BiolBiotechnol-ogy Institute, P. O. Box 411, H-2101 Go¨do¨llo¨, Hungary. Phone: 36-28-526-155. Fax: 36-28-526-145. E-mail: burgyan @abc.hu.
† Supplemental material for this article may be found at http://jvi .asm.org/.
‡ Present address: The Sainsbury Laboratory, John Innes Centre, Colney, Norwich NR4 7UH, United Kingdom.
7812
on November 8, 2019 by guest
http://jvi.asm.org/
likely arise from separate DICER-like activities (17, 48, 50). The 21-nt siRNAs are sufficient for RISC-mediated cleavage of target transcripts and are believed to be involved in short-distance signaling, while the 25-nt siRNAs are associated with DNA methylation and the systemic spread of silencing (17, 21). Plant viruses are known as strong inducers as well as targets of PTGS. During a virus infection, the accumulation of 21-nt double-stranded siRNAs is observed in local and systemic tis-sues (18, 49), indicating the activation of PTGS. Elevated siRNA levels are correlated with a reduction in viral titer, and in some cases, immunity or recovery in upper noninoculated leaves (41, 49). Thus, PTGS acts as an RNA-mediated defense response to protect plants against viral infection (34, 54, 57). Consequently, to counteract RNA silencing, many plant vi-ruses have evolved proteins that suppress various steps of the silencing machinery (29, 47, 48, 56). It is not clear how most viral suppressor proteins counteract silencing at the molecular level. The tombusviral 19-kDa silencing suppressor protein p19 represents an exception. Indeed, recent advances in our un-derstanding of the molecular mechanism underlying p19’s sup-pressive activity revealed that p19 specifically binds 21-nt dou-ble-stranded siRNAs in vitro and in vivo, preventing siRNA incorporation into effector complexes such as the RISC (27, 48). The three-dimensional X-ray crystal structure of a p19-siRNA complex revealed that a p19 dimer acts as a caliper, binding the ends of the siRNA duplex while measuring its length (51, 59).
Cymbidium ringspot virus (CymRSV) is a member of the
Tombusvirusgenus containing a positive single-stranded RNA
genome with five open reading frames (ORFs) (44). It is widely assumed that positive-strand RNA viruses replicate their ge-nomes via dsRNA intermediates that may activate the siRNA-generating machinery (1). However, the existence of long viral dsRNA intermediates and their accessibility for DICER cleav-age have not been proven experimentally. In addition, our recent studies suggested that highly structured viral RNAs might also be processed into siRNAs in virus-infected plants (24, 49).
The work presented here addresses the origins and molec-ular nature of virus-derived siRNAs and reports the develop-ment of cloning and sequencing approaches to characterize viral siRNAs accumulating in infected plants. Based on our findings, we suggest that virus-derived siRNAs are predomi-nantly produced by direct DICER cleavage of imperfect du-plexes originating from highly base-paired structures from the positive-strand viral genomic RNA.
MATERIALS AND METHODS
Plasmid constructs, in vitro RNA transcription, and plant inoculation.The infectious cDNA clones of CymRSV, i.e., Cym19stop, TMV.hpPDS60, TMV
.PDSas110, and PVX, were reported previously (9, 12, 24, 49). A full-length
infectious clone of the tobacco mosaic virus (TMV) legume strain will be de-scribed elsewhere.
In vitro transcription of the previously mentioned viral constructs from linear-ized DNA templates and the inoculation of RNA transcripts ontoNicotiana benthamianaplants were performed as described earlier (49). TMV-derived plasmids were linearized withSmaI, and in vitro RNA transcripts were capped with a cap analogue (New England Biolabs, Hitchin, United Kingdom) as pre-viously reported (24).
RNA extraction and analysis.The total RNA was extracted from 100 mg of leaf tissue (12). Briefly, the homogenized plant materials were resuspended in 600l of extraction buffer (0.1 M glycine-NaOH, pH 9.0, 100 mM NaCl, 10 mM
EDTA, 2% sodium dodecyl sulfate, and 1% sodium lauroylsarcosine) and mixed with an equal volume of phenol. The aqueous phase was treated with equal volumes of phenol and chloroform, precipitated with ethanol, and resuspended in sterile water. RNA gel blot analysis of high-molecular-weight RNAs was performed as described previously (12).
Detection, isolation, labeling, and sequencing of virus-derived siRNAs.RNA gel blot analysis of siRNAs was performed as described previously (49). For preparations of labeled viral siRNAs, 15 to 20g of total RNA from virus-infected plants was subjected to electrophoresis through a 12% denaturing poly-acrylamide gel, followed by staining with a 1⫻Tris-borate-EDTA, 0.5g/ml ethidium bromide solution for 20 min. ss- and dsRNAs were visualized with UV light and excised from the gel. The gel slices were crushed, covered with 2 volumes of elution buffer (80% formamide, 40 mM PIPES, pH 6.4, 1 mM EDTA, and 400 mM NaCl), and incubated overnight. The gel residues were pelleted by centrifugation, and the supernatants were ethanol precipitated. The purified siRNAs (approximately 1g) were dephosphorylated and subsequently labeled in 10-l reaction mixtures in the presence of [␥-32P]ATP and RNasin with 8 U
of T4 polynucleotide kinase. The labeled virus-specific siRNAs were used for hybridization (49).
Purified siRNAs were cloned according to the original siRNA cloning protocol as described previously (14), with some minor modifications. Briefly, denaturing gel-purified viral dephosphorylated small RNAs were ligated to 3⬘-terminal adapter oligonucleotides. The ligation products were 5⬘ end labeled with [␥-32
P]ATP and recovered after migration in a sequencing gel (8%
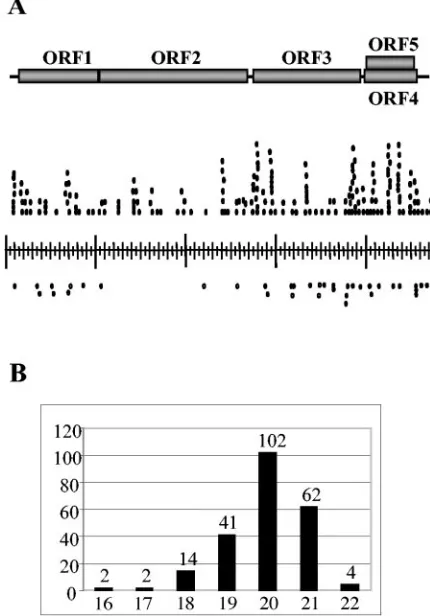
polyacryl-FIG. 1. Origin and characterization of siRNAs derived from CymRSV. (A) Schematic representation of CymRSV genome (top). Each of the shaded boxes represents a viral ORF. The position of each siRNA sequence is represented by a small circle alongside the viral genome. Circles above the genome bar, siRNAs derived from the positive viral strand; circles below the genome bar, siRNAs derived from the negative viral strand. siRNAs with the same sequences are indicated as chains of connected circles. (B) Size distribution of se-quenced siRNAs. Numbers below the columns indicate the sizes of virus-specific siRNAs, and numbers above the columns indicate the numbers of siRNAs of different sizes.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:2.585.312.527.314.622.2]amide, 8 M urea). 5⬘adapters were then ligated to the phosphorylated interme-diate products. These ligation products were then gel purified, excised, and eluted as previously described (14). After reverse transcription-PCR amplifica-tion, the oligonucleotide-linked cDNAs were cloned as concatemers into the pBluescript SK(⫹) vector. The recombinant clones were randomly selected and sequenced.
IP.For immunoprecipitation (IP), 2 g of CymRSV- or Cym19stop-infectedN. benthamianaleaves showing systemic symptoms were collected at 6 days posti-noculation and used to prepare extracts in IP buffer (30 mM HEPES-KOH, pH 7.5, 100 mM NaCl, 66 mM KCl, 1 mM MgCl2, 1 mM dithiothreitol, 0.2 mM
phenylmethylsulfonyl fluoride), and the extracts were then centrifuged at 15,000
⫻gfor 10 min. IPs were performed at 41°C for 3 to 5 h. Beads were centrifuged and washed in 1⫻IP buffer two times. Input extracts and IP eluates were used for Western blotting analysis and RNA isolation. RNA molecules were separated in a sequencing gel (12% polyacrylamide, 8 M urea), blotted, and hybridized with an appropriate probe.
RESULTS AND DISCUSSION
The majority of viral siRNAs are derived from viral positive-strand RNA. To evaluate the polarity of siRNAs in virus-infected plants, we extracted total RNAs from systemically infected leaves of CymRSV-infectedN. benthamianaplants at 7 days postinoculation. The extracted RNA samples were sep-arated by 8% denaturing polyacrylamide gel electrophoresis (PAGE), blotted, and hybridized with a coat protein (CP) probe (48) representing the central part of the virus genome. CP-specific siRNAs accumulated to a relatively high level and comigrated exclusively with a 21-mer RNA oligonucleotide (not shown). The purified siRNAs were cloned and sequenced to determine their exact size as well as the origin of CymRSV-specific siRNAs. Random sequencing of a total of 70 clones identified 228 siRNAs homologous to the sequence of the CymRSV genome (see Table S1 in the supplemental material). Eight siRNAs containing nonviral sequences were not ana-lyzed further. The nucleotide sequences of the virus-derived siRNAs were derived from regions along the length (4,733 nt) of the viral genome (Fig. 1A). A size distribution analysis of the CymRSV siRNAs confirmed that these RNAs belonged to the short siRNA fraction, with an average size of 20 to 21 nucleotides (Fig. 1B).
[image:3.585.46.283.67.246.2]The most accepted virus-induced RNA silencing model (1, 54, 57) suggests that a DICER-like enzyme generates siRNAs from double-stranded replicative intermediates of RNA vi-ruses, and thus the expected ratio between positive- and neg-ative-strand-derived siRNAs should be 1:1. However, sequenc-ing of the cloned siRNAs indicated that 80% of viral siRNAs were derived from the positive-strand viral RNA, reinforcing our previous observations of the nature of virus-derived siRNAs (49). In addition, the sequences of 228 siRNAs ho-mologous to the CymRSV genomic RNA revealed that 194 FIG. 2. Analysis of CymRSV siRNA composition. (A) RNA
sam-ples were extracted from CymRSV-infected plants and separated in a 10% native polyacrylamide gel alongside with both single-stranded and double-stranded 21-nt synthetic siRNAs. (B) RNase A digestion assay with purified CymRSV-specific siRNA (CymRSV siRNAnat) under
nondenaturing conditions. Synthetic double-stranded siRNAs (con-trol) and CymRSV siRNAnatwere incubated in high (2⫻SSC)- and
low (0.1⫻ SSC)-salt buffer in the presence of increasing RNase A concentrations. (C) The CymRSV siRNAnat shown in panel A was
labeled and used as a probe in a Northern blot experiment to hybridize both negative- and positive-strand transcripts for the CymRSV coat protein (CP). A32P-labeled, PCR-amplified DNA fragment containing
the CP region was also used as a probe to show the amount of loaded CymRSV CP transcript.
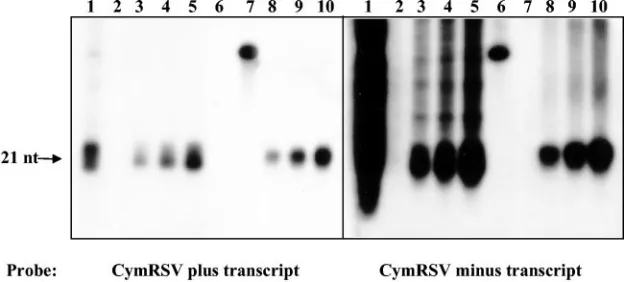
FIG. 3. Analysis of CymRSV p19-bound siRNA polarity. RNA samples were extracted from CymRSV-infected plants or from immunopre-cipitates (anti-p19 IP) by use of a p19 antibody. Lanes 1, input RNA from protein extract used for IP; lanes 2, RNA extract from IP using preimmune antiserum; lanes 3 to 5, 2, 4, and 6l of RNA extract, respectively, from anti-p19 IP; lanes 6, 20 pM 46-mer RNA oligonucleotide containing positive-strand sequence for CymRSV CP; lanes 7, 20 pM 46-mer RNA oligonucleotide containing negative-strand sequence for CymRSV CP; lanes 8 to 10, 2, 4, and 6l of RNA extract, respectively, from CymRSV-infected plants. The membrane was hybridized with CymRSV positive-strand (left) and negative-strand (right)␣-32P-labeled RNA transcripts.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.585.136.449.527.668.2]virus-specific siRNA clones could be assigned to several clus-ters that likely represent hot spots for siRNA production (Fig. 1A). These results suggest that siRNAs in the CymRSV system are predominantly processed by DICER cleavage of imperfect duplexes, which can be formed from the positive-strand viral RNA. This process resembles the production of short-lived, imperfectly pairing, double-stranded intermediates during miRNA generation (4, 8). The preferential incorporation of positive-strand siRNAs into the RISC cannot explain this asymmetric accumulation because the sequence composition of siRNAs does not display a significantly reduced stability profile at the 5⬘end (see Table S1 in the supplemental mate-rial). This observation of asymmetry contrasts with other re-cent interpretations in which siRNAs were thought to arise from dsRNAs that were produced either by viral transcription events or by the action of host RNA-dependent RNA poly-merase (1, 54, 57).
Virus-derived double-stranded siRNAs are imperfect du-plexes.The finding that virus-derived siRNAs are asymmetric raised the question of whether these siRNAs are present in infected cells in a ss- or dsRNA form. To determine the struc-ture of viral siRNAs, we examined the mobilities of these molecules under native nondenaturing conditions. RNA
ex-tracts of CymRSV-infected tissues were separated by 10% native PAGE along with both single-stranded and double-stranded 21-nt synthetic siRNAs. The results presented in Fig. 2A indicate that only one additional low-molecular-weight RNA fraction could be detected in virus-infectedN. benthami-ana plants compared to the noninfected plant extract. This low-molecular-weight RNA fraction comigrated with siRNA duplexes, strongly suggesting that it was composed of a double-stranded siRNA. Northern hybridization of the same samples confirmed that this double-stranded siRNA was derived from CymRSV RNA (27; data not shown).
If viral siRNAs are mostly derived from highly structured regions of the positive-strand viral RNA, then these double-stranded siRNAs (Fig. 2A) should contain non-base-paired nucleotides, which are nuclease sensitive. To confirm this hy-pothesis, we performed RNase A digestion to assess whether these molecules were perfect or imperfect duplexes. Perfect dsRNAs are resistant to RNase A in a high salt buffer (2⫻SSC [1⫻SSC is 0.15 M NaCl plus 0.015 M sodium citrate]), while imperfect dsRNAs with mismatches are rapidly degraded un-der the same conditions (45). The double-stranded siRNA fraction purified from the native gel (CymRSV siRNAnat) was
[image:4.585.127.464.69.364.2]sensitive to RNase A, indicating the presence of mismatches FIG. 4. Identification of polarity of 21-nt siRNAs accumulating in PVX- and TMV-infected plants. (A) Accumulation of virus-specific high-molecular-weight genomic RNAs and siRNAs. The left (PVX) and right (TMV) panels show the viral RNAs (top) and siRNAs (bottom) in RNA extracts of virus-infected tissues. High-molecular-weight RNA extracts were separated in ethidium bromide-stained agarose gels, and virus-specific siRNAs were separated by 10% denaturing PAGE and hybridized with appropriate virus-specific probes. RNA oligonucleotide size markers were run along with the siRNAs. m, mock-infected control plants. (B) Known amounts of positive and negative strands of PVX and TMV RNAs were in vitro transcribed, subjected to Northern blotting, and then hybridized with 5⬘-end-labeled virus-specific siRNAs (panel A). As a loading control for in vitro-generated transcripts, the same membrane was hybridized with a32P-labeled PCR-amplified DNA fragment derived
from PVX or TMV as indicated. Numbers above the lanes indicate the quantities of loaded RNA in ng. “⫹” and “⫺” indicate the polarities of the loaded viral RNA transcripts. M, control RNA sample extracted from a healthy plant; G, CymRSV genomic transcript.
on November 8, 2019 by guest
http://jvi.asm.org/
between the two viral siRNA strands (Fig. 2B, top panel). In contrast, control synthetic perfectly double-stranded siRNAs were completely resistant to RNase A, even at a high nuclease concentration and in high-salt buffer (Fig. 2B, bottom panel). In addition, labeled CymRSV-derived small RNAs isolated by native PAGE (CymRSV siRNAnat) hybridized more
inten-sively to the negative-strand coat protein (CP) transcript than to the corresponding positive-strand transcript (Fig. 2C). The finding that virus-derived siRNAs are imperfect dsRNAs is consistent with the model that viral siRNAs are processed from highly base-paired structures. These observations also corroborate our previous results that artificially extended base-paired structures imbedded in viral ssRNAs give rise to corre-spondingly high levels of siRNAs. Together, these observations suggest that highly structured regions within viral ssRNAs are able to trigger RNA silencing (49).
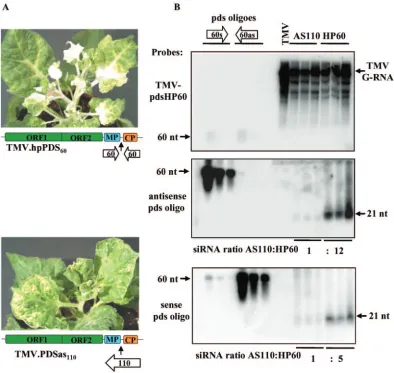
[image:5.585.96.490.68.441.2]p19-bound siRNAs have predominantly positive-strand po-larity.We recently reported that the majority of virus-specific siRNAs are bound by p19 in tombusvirus-infected plants. These p19-bound siRNAs are double-stranded and can effi-ciently trigger the sequence-specific degradation of CymRSV viral RNA in aDrosophila melanogaster embryo extract (27). To test whether p19-bound double-stranded viral siRNAs also have an asymmetric composition with respect to strand speci-ficity, we analyzed the ratio of positive to negative strands in p19-bound siRNAs. RNAs were extracted from anti-p19-im-munoprecipitated p19-siRNA complexes derived from CymRSV-infected plants (27) and subsequently analyzed by Northern hybridization using strand-specific probes. As ex-pected from the sequence analysis of siRNAs, the majority of p19-bound siRNAs originated from the positive strand of the viral RNA (Fig. 3), further supporting the model that FIG. 5. Accumulation ofPDS-specific siRNAs in TMV vector-infected plants carrying hairpin (TMV.hpPDS60) and antisense (TMV.PDSas110) PDSsequences, as previously described (24). (A) Virus-infected plants developing photobleached white leaves typical ofPDS-silenced plants at 3 weeks postinoculation. The genome organization of TMV viral vectors is represented schematically, with colored boxes representing the different viral ORFs. Open arrows show the positions and orientations of inserted sequences in the TMV genome. (B) RNA gel blot analysis of viral RNAs extracted at 2 weeks postinoculation from plants infected with the TMV.hpPDS60hairpin or TMV.PDSas110antisense construct. RNAs were
separated in 1.2% denaturing agarose gels, blotted, and hybridized with a TMV.hpPDS60probe.PDS-specific siRNAs were separated by 10%
denaturing PAGE and hybridized with a labeled 60-mer oligonucleotide corresponding to the sense and antisense counterparts of thePDScDNA from the TMV.hpPDS60construct. TMV, wild-type virus-infected RNA extract. One, 5, and 25 pM 60-mer sense (s) and antisense (as)PDS
oligonucleotides, indicated by open arrows, were used to control the specificity of the probes.
on November 8, 2019 by guest
http://jvi.asm.org/
CymRSV siRNAs originate from highly structured viral RNA regions. An alternative explanation for the specific accumula-tion of positive-strand siRNAs could be that the positive strand of a perfect double-stranded siRNA is preferentially incorpo-rated into the RISC or other effector complexes instead of the negative strand, with the latter being rapidly eliminated by cellular nucleases. However, our previous observation that p19 sequesters siRNAs before their incorporation into the RISC, when they are still double-stranded siRNAs (27), strongly ar-gues against this alternative model.
The preferential accumulation of positive-strand siRNAs is a general characteristic of positive-strand RNA viruses. To investigate whether the characteristics of CymRSV siRNAs reflect the general rule for virus-derived siRNA generation in virus-infected plants, we also included two other nonrelated positive-strand RNA viruses, Potato virus X (PVX) (9) and TMV (46), in our analysis. As with CymRSV, infections ofN.
benthamiana plants with PVX and TMV resulted in the
ap-pearance of single siRNA fractions of 21 to 22 nt (Fig. 4A). To analyze whether PVX- and TMV-derived siRNAs are also asymmetric, we induced the transcription of the CP-encoding regions of the two viruses with both positive and negative polarities. These transcripts were then subjected to Northern hybridization and probed with 5⬘-labeled siRNAs isolated from either PVX- or TMV-infected plants. In both cases, the ma-jority of the labeled virus-specific siRNAs hybridized more strongly to the negative transcript, indicating that similar to the case with CymRSV, PVX and TMV siRNAs are derived mainly from viral RNAs with a positive polarity (Fig. 4B).
It was recently reported that the expression of a host gene fragment as a short inverted repeat from TMV leads to more efficient silencing of the cognate mRNA than the expression of a similar sequence in just the sense or antisense orientation (24). To better understand this observation at the molecular level, we compared the siRNA accumulation in plants infected with a viral vector carrying a hairpin structure to that in plants infected with an antisense sequence. To this end, viral tran-scripts carrying 60-bp inverted repeats (TMV.hpPDS60) or the
corresponding antisense sequence (TMV.PDSas110)
homolo-gous to the endogenous phytoene desaturase (PDS) gene (24) were used to inoculate N. benthamiana plants. RNAs were extracted from TMV-infected leaves showing the white colora-tion characteristic of the photobleaching phenotype observed for PDS-silenced plants and then analyzed for both viral RNA accumulation and virus-derived siRNA composition (Fig. 5). The photobleaching phenotype of PDS-silenced plants was more pronounced with the TMV.hpPDS60hairpin construct
than in plants infected with the TMV.PDSas110antisense
con-struct (Fig. 5A), in spite of similar viral RNA accumulation in virus-infected plants (Fig. 5B, top panel). More importantly, the siRNA accumulation originating from the hairpin se-quence was significantly higher for both the plus (12 times) and minus (5 times) polarities than that from the antisense se-quence containing the 60-nt sese-quence of the corresponding hairpin construct (Fig. 5B, bottom panels).
Taken together, these results strongly suggest that one or more of the plant-encoded DICER-like enzymes are able to recognize highly structured regions within viral ssRNAs and process them into siRNAs. This property resembles the DICER-like activity involved in the generation of miRNAs
from short-hairpin precursors (16, 22, 30, 38, 42, 58), although recent data indicate that siRNAs and miRNAs require differ-ent DICER-like activities in plants (15, 50, 58). The discrete size (21 nt) of virus-derived siRNAs, however, suggests that one of the still unidentified plant DICERs has evolved specif-ically to process highly structured viral RNAs and is likely a key player in the PTGS-mediated antiviral defense. A recent re-port demonstrated that anArabidopsis dcl2mutant transiently accumulated lower levels ofTurnip crinkle virus-derived siR-NAs than did wild-type plants (58). However, there is likely to be some degree of redundancy because dcl1, dcl2, anddcl3
mutants were not defective in siRNA production when they were infected with other viruses.
The fact that plant DICER-like proteins generate predom-inantly positive-strand siRNAs makes the siRNA-based anti-viral surveillance system more effective since the siRNAs po-tentially target the negative viral strand for RISC-mediated cleavage, which is required for replication and is much less abundant than the positive-strand viral RNA. In further sup-port of this model, it was resup-ported that transgenic plants ex-pressing a positive-strand viral sequence specifically target the negative-strand RNA of TMV. This suggests that the trans-genically expressed sense viral sequence can be processed into positive-strand siRNAs, which in turn specifically target the negative-strand viral RNA replicative intermediate for degra-dation (31). Since the sense siRNAs derived from folded RNA are partially complementary to the positive-strand viral RNA, positive siRNAs could also guide the translational repression of viral RNA in a manner analogous to miRNA-mediated translational control, which relies on only partial complemen-tarity between the miRNA and the target. This type of gene silencing is common in animal systems (2, 5) but also seems to operate in plants (3, 10). Further experiments are required to address whether this additional level of control occurs.
ACKNOWLEDGMENTS
We thank Daniel Silhavy, Gyo¨rgy Szittya, Ga´bor Giczey, and Alan Herr for critical readings of and helpful comments on the manuscript. L.L. is a recipient of a Bolyai Ja´nos Fellowship. This research was supported by grants from the Hungarian Scientific Research Fund (OTKA; T046728), the “RIBOREG” EU project (LSHG-CT-2003503022), and the Scientia Amabilis Foundation.
REFERENCES
1.Ahlquist, P.2002. RNA-dependent RNA polymerases, viruses, and RNA silencing. Science296:1270–1273.
2.Ambros, V., R. C. Lee, A. Lavanway, P. T. Williams, and D. Jewell.2003. MicroRNAs and other tiny endogenous RNAs in C. elegans. Curr. Biol.
13:807–818.
3.Aukerman, M. J., and H. Sakai.2003. Regulation of flowering time and floral organ identity by a MicroRNA and its APETALA2-like target genes. Plant Cell10:10.
4.Bartel, B., and D. P. Bartel.2003. MicroRNAs: at the root of plant devel-opment? Plant Physiol.132:709–717.
5.Baulcombe, D.2004. RNA silencing in plants. Nature431:356–363. 6.Bernstein, E., A. A. Caudy, S. M. Hammond, and G. J. Hannon.2001. Role
for a bidentate ribonuclease in the initiation step of RNA interference. Nature409:363–366.
7.Carrington, J. C., and V. Ambros.2003. Role of microRNAs in plant and animal development. Science301:336–338.
8.Chapman, E. J., A. I. Prokhnevsky, K. Gopinath, V. V. Dolja, and J. C. Carrington.2004. Viral RNA silencing suppressors inhibit the microRNA pathway at an intermediate step. Genes Dev.18:1179–1186.
9.Chapman, S., T. Kavanagh, and D. Baulcombe.1992. Potato virus X as a vector for gene expression in plants. Plant J.2:549–557.
10.Chen, X.2003. A MicroRNA as a translational repressor of APETALA2 in Arabidopsis flower development. Science11:11.
on November 8, 2019 by guest
http://jvi.asm.org/
11.Dalmay, T., A. Hamilton, S. Rudd, S. Angell, and D. C. Baulcombe.2000. An RNA-dependent RNA polymerase gene in Arabidopsis is required for post-transcriptional gene silencing mediated by a transgene but not by a virus. Cell101:543–553.
12.Dalmay, T., L. Rubino, J. Burgyan, A. Kollar, and M. Russo.1993. Func-tional analysis of cymbidium ringspot virus genome. Virology194:697–704. 13.Doench, J. G., C. P. Petersen, and P. A. Sharp.2003. siRNAs can function
as miRNAs. Genes Dev.17:438–442.
14.Elbashir, S. M., W. Lendeckel, and T. Tuschl.2001. RNA interference is mediated by 21- and 22-nucleotide RNAs. Genes Dev.15:188–200. 15.Finnegan, E. J., R. Margis, and P. M. Waterhouse.2003. Posttranscriptional
gene silencing is not compromised in the Arabidopsis CARPEL FACTORY (DICER-LIKE1) mutant, a homolog of Dicer-1 from Drosophila. Curr. Biol.
13:236–240.
16.Grishok, A., A. E. Pasquinelli, D. Conte, N. Li, S. Parrish, I. Ha, D. L. Baillie, A. Fire, G. Ruvkun, and C. C. Mello.2001. Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell106:23–34.
17.Hamilton, A., O. Voinnet, L. Chappell, and D. Baulcombe.2002. Two classes of short interfering RNA in RNA silencing. EMBO J.21:4671–4679. 18.Hamilton, A. J., and D. C. Baulcombe.1999. A species of small antisense
RNA in posttranscriptional gene silencing in plants. Science286:950–952. 19.Hammond, S. M., E. Bernstein, D. Beach, and G. J. Hannon.2000. An
RNA-directed nuclease mediates post-transcriptional gene silencing in Dro-sophila cells. Nature404:293–296.
20.Hannon, G. J., and D. S. Conklin.2004. RNA interference by short hairpin RNAs expressed in vertebrate cells. Methods Mol. Biol.257:255–266. 21.Himber, C., P. Dunoyer, G. Moissiard, C. Ritzenthaler, and O. Voinnet.
2003. Transitivity-dependent and -independent cell-to-cell movement of RNA silencing. EMBO J.22:4523–4533.
22.Hutvagner, G., J. McLachlan, A. E. Pasquinelli, E. Balint, T. Tuschl, and P. D. Zamore.2001. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science293:834– 838.
23.Hutvagner, G., and P. D. Zamore.2002. RNAi: nature abhors a double-strand. Curr. Opin. Genet. Dev.12:225–232.
24.Lacomme, C., K. Hrubikova, and I. Hein.2003. Enhancement of virus-induced gene silencing through viral-based production of inverted-repeats. Plant J.34:543–553.
25.Lagos-Quintana, M., R. Rauhut, W. Lendeckel, and T. Tuschl.2001. Iden-tification of novel genes coding for small expressed RNAs. Science294:853– 858.
26.Lagos-Quintana, M., R. Rauhut, A. Yalcin, J. Meyer, W. Lendeckel, and T. Tuschl.2002. Identification of tissue-specific microRNAs from mouse. Curr. Biol.12:735–739.
27.Lakatos, L., G. Szittya, D. Silhavy, and J. Burgyan.2004. Molecular mech-anism of RNA silencing suppression mediated by p19 protein of tombusvi-ruses. EMBO J.23:876–884.
28.Lee, R. C., and V. Ambros.2001. An extensive class of small RNAs in Caenorhabditis elegans. Science294:862–864.
29.Li, W. X., and S. W. Ding.2001. Viral suppressors of RNA silencing. Curr. Opin. Biotechnol.12:150–154.
30.Llave, C., K. D. Kasschau, M. A. Rector, and J. C. Carrington.2002. En-dogenous and silencing-associated small RNAs in plants. Plant Cell14:1605– 1619.
31.Marano, M. R., and D. Baulcombe.1998. Pathogen derived resistance tar-geted against the negative strand RNA of tobacco mosaic virus: RNA strand specific gene silencing? Plant J.13:537–546.
32.Meister, G., and T. Tuschl.2004. Mechanisms of gene silencing by double-stranded RNA. Nature431:343–349.
33.Mlotshwa, S., O. Voinnet, M. F. Mette, M. Matzke, H. Vaucheret, S. W. Ding, G. Pruss, and V. B. Vance.2002. RNA silencing and the mobile silencing signal. Plant Cell14(Suppl.):S289–S301.
34.Moissiard, G., and O. Voinnet.2004. Viral suppression of RNA silencing in plants. Mol. Plant Pathol.5:71–82.
35.Mourelatos, Z., J. Dostie, S. Paushkin, A. Sharma, B. Charroux, L. Abel, J. Rappsilber, M. Mann, and G. Dreyfuss.2002. miRNPs: a novel class of ribonucleoproteins containing numerous microRNAs. Genes Dev.16:720– 728.
36.Mourrain, P., C. Beclin, T. Elmayan, F. Feuerbach, C. Godon, J. B. Morel,
D. Jouette, A. M. Lacombe, S. Nikic, N. Picault, K. Remoue, M. Sanial, T. A. Vo, and H. Vaucheret.2000. Arabidopsis SGS2 and SGS3 genes are required for posttranscriptional gene silencing and natural virus resistance. Cell101:
533–542.
37.Nykanen, A., B. Haley, and P. D. Zamore.2001. ATP requirements and small interfering RNA structure in the RNA interference pathway. Cell107:309– 321.
38.Pasquinelli, A. E., and G. Ruvkun.2002. Control of developmental timing by microRNAs and their targets. Annu. Rev. Cell Dev. Biol.18:495–513. 39.Peragine, A., M. Yoshikawa, G. Wu, H. L. Albrecht, and R. S. Poethig.2004.
SGS3 and SGS2/SDE1/RDR6 are required for juvenile development and the production oftrans-acting siRNAs in Arabidopsis. Genes Dev.18:2368– 2379.
40.Plasterk, R. H.2002. RNA silencing: the genome’s immune system. Science
296:1263–1265.
41.Ratcliff, F., B. D. Harrison, and D. C. Baulcombe.1997. A similarity between viral defense and gene silencing in plants. Science276:1558–1560. 42.Reinhart, B. J., and D. P. Bartel.2002. Small RNAs correspond to
centro-mere heterochromatic repeats. Science297:1831.
43.Reinhart, B. J., F. J. Slack, M. Basson, A. E. Pasquinelli, J. C. Bettinger, A. E. Rougvie, H. R. Horvitz, and G. Ruvkun.2000. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature
403:901–906.
44.Russo, M., J. Burgyan, and G. P. Martelli.1994. Molecular biology of tombusviridae. Adv. Virus Res.44:381–428.
45.Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
46.Shivprasad, S., G. P. Pogue, D. J. Lewandowski, J. Hidalgo, J. Donson, L. K. Grill, and W. O. Dawson.1999. Heterologous sequences greatly affect for-eign gene expression in tobacco mosaic virus-based vectors. Virology255:
312–323.
47.Silhavy, D., and J. Burgyan.2004. Effects and side-effects of viral RNA silencing suppressors on short RNAs. Trends Plant Sci.9:76–83. 48.Silhavy, D., A. Molnar, A. Lucioli, G. Szittya, C. Hornyik, M. Tavazza, and
J. Burgyan.2002. A viral protein suppresses RNA silencing and binds si-lencing-generated, 21- to 25-nucleotide double-stranded RNAs. EMBO J.
21:3070–3080.
49.Szittya, G., A. Molnar, D. Silhavy, C. Hornyik, and J. Burgyan.2002. Short defective interfering RNAs of tombusviruses are not targeted but trigger post-transcriptional gene silencing against their helper virus. Plant Cell14:
359–372.
50.Tang, G., B. J. Reinhart, D. P. Bartel, and P. D. Zamore.2003. A biochem-ical framework for RNA silencing in plants. Genes Dev.17:49–63. 51.Vargason, J., G. Szittya, J. Burgyan, and T. M. Hall.2003. Size selective
recognition of siRNA by an RNA silencing suppressor. Cell115:799–811. 52.Vazquez, F., H. Vaucheret, R. Rajagopalan, C. Lepers, V. Gasciolli, A. C.
Mallory, J. L. Hilbert, D. P. Bartel, and P. Crete.2004. Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs. Mol. Cell
16:69–79.
53.Verdel, A., S. Jia, S. Gerber, T. Sugiyama, S. Gygi, S. I. Grewal, and D. Moazed.2004. RNAi-mediated targeting of heterochromatin by the RITS complex. Science303:672–676.
54.Voinnet, O.2001. RNA silencing as a plant immune system against viruses. Trends Genet.17:449–459.
55.Voinnet, O.2002. RNA silencing: small RNAs as ubiquitous regulators of gene expression. Curr. Opin. Plant Biol.5:444.
56.Voinnet, O., Y. M. Pinto, and D. C. Baulcombe.1999. Suppression of gene silencing: a general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA96:14147–14152.
57.Waterhouse, P. M., M. B. Wang, and T. Lough.2001. Gene silencing as an adaptive defence against viruses. Nature411:834–842.
58.Xie, Z., L. K. Johansen, A. M. Gustafson, K. D. Kasschau, A. D. Lellis, D. Zilberman, S. E. Jacobsen, and J. C. Carrington.2004. Genetic and func-tional diversification of small RNA pathways in plants. PLoS Biol.2:E104. 59.Ye, K., L. Malinina, and D. J. Patel.2003. Recognition of small interfering
RNA by a viral suppressor of RNA silencing. Nature3:3.
60.Zamore, P. D.2002. Ancient pathways programmed by small RNAs. Science
296:1265–1269.