0022-538X/91/052467-09$02.00/0
Copyright C) 1991, American Society for
Microbiology
Both Nonstructural Proteins
NS2B and NS3 Are Required for the
Proteolytic Processing of
Dengue
Virus Nonstructural Proteins
BARRY FALGOUT, MICHtLE PETHEL, YI-MING ZHANG,t ANDCHING-JUH LAI*
MolecularViralBiology Section, Laboratory of Infectious Diseases,NationalInstituteof Allergy and
Infectious
Diseases,
Bethesda,
Maryland
20892 Received 9 August1990/Accepted4February 1991The cleavages at the junctions of the flavivirus nonstructural (NS) proteins NS2A/NS2B, NS2B/NS3, NS3/NS4A, and NS4B/NS5 share an amino acid sequence motif and are presumably catalyzed by a
virus-encoded protease. We constructed recombinant vaccinia viruses expressing various portions of the NS region of the dengue virus type 4 polyprotein. By analyzing immune precipitates of35S-labeled lysates of recombinant virus-infectedcells, wecouldmonitor theNS2A/NS2B, NS2B/NS3,andNS3/NS4Acleavages. A
polyprotein composed of NS2A, NS2B, and the N-terminal 184 amino acids of NS3 was cleaved at the NS2A/NS2B and NS2B/NS3 junctions,whereasasimilar polyprotein containing only the first 77 amino acids
of NS3wasnotcleaved.Thisfindingis consistent with the proposal that theN-terminal180aminoacids of NS3 constituteaproteasedomain. PolyproteinscontainingNS2A and NS3 with large in-frame deletions of NS2B
werenotcleavedattheNS2A/NS2BorNS2B/NS3 junctions.Coinfection witharecombinant expressingNS2B
complemented these NS2B deletions forNS2B/NS3cleavage and probably alsoforNS2A/NS2Bcleavage. Thus, NS2B is also required for theNS2A/NS2B andNS2B/NS3cleavages andcanactintrans. Other experiments showedthatNS2Bwasneeded, apparently in cis,forNS3/NS4Acleavageandforaseries of internal cleavages
in NS3. Indirect evidence that NS3 can also act in trans was obtained. Models are discussed for a
two-component proteaseactivity requiringbothNS2B and NS3. The four serotypes of dengue virus (DEN) constitute a
subgroup of the Flaviviridae, afamily of about 70
serologi-cally related viruses (15, 55). Many members of this family
aretheetiologicagentsofavariety of human diseases, such as dengue fever, yellow fever, Japanese encephalitis, and
tick-borne encephalitis. Most flaviviruses, including the medically important ones, are transmitted to humans by mosquitoortickbite.
Likeotherflaviviruses, DEN iscomposed of the envelope (E) and membrane (M) proteins, embedded inahost-derived
membrane, and the capsid (C) protein, which is complexed with thesingle-strandedgenomic RNA of approximately 11 kb (35). The genomic RNA is of thepositive sense and is
capped but notpolyadenylated (12, 54). Subgenomic RNA
was not detected in flavivirus-infected cells (5, 36). The
completesequencesof several flavivirusgenomeshave been determined(6, 7, 13, 14, 16, 17, 24, 25, 27, 29, 30-32, 38-40, 43, 50, 53, 57). Analysis of thesesequencesrevealed that the
flavivirusgenomecontainsoneopenreading framespanning
morethan10kb. Individual flavivirusproteinsareevidently
expressed by translation of the genomic RNA into a
poly-protein followed by proteolytic cleavages. For theprototype yellow fever virus, the order of the flavivirus polyprotein
was established to be NH2-C-prM-E-NS1-NS2A-NS2B-NS3-NS4A-NS4B-NS5-COOH, whereprM is theprecursor
toMand NSare nonstructuralproteins (2, 43, 44). Several
cleavage sites have been reassignedastheN-terminal amino
acid sequences for almost all Kunjin virus proteins have become available (13, 47, 49). The N-terminal amino acid
sequence data obtained for other flavivirus systems are in
agreement with the cleavage sites predicted by sequence
*Correspondingauthor.
tPresent address: Laboratory of Molecular Retrovirology, Georgetown University, Washington,DC20007.
homologytoKunjin virus (3, 4, 6, 7, 9, 10, 22, 29, 34, 38, 50, 53, 56),exceptfortworeportsofanN-terminally shortened
form of C (7, 38). The C-terminalamino acid sequences of
most of the flavivirus proteins have also been determined (37, 48, 56). These results show that for all proteins except C, the C terminus exactly abuts the N terminus of the downstream protein, such that no aminoacids are excised
during proteolyticprocessing.
Analysis of the amino acid sequences near the cleavage sites of the flavivirus polyprotein suggests that there are
several distinct classesofcleavage mechanisms.The
cleav-ages atthe C/prM, prM/E, E/NS1, and NS4A/NS4B junc-tions are thought to be mediated by signalase (43, 47). Support for this for all but the NS4A/NS4B cleavage has been obtained frominvitro translationandprocessing stud-ies(33, 37, 45, 51). Theproteasethat cleaves theNS1/NS2A junction hasnotbeenidentified, although it has beenshown thatthe 8 C-terminal amino acids of NS1 and theN-terminal 70% of NS2Aarerequired for this cleavage (19, 20, 26).The prM/M cleavage apparently represents another processing mechanism, in which cleavage occursin anacidified
vesic-ular compartmentatalate stepduringvirion maturation(42). The remaining cleavages at the NS2A/NS2B, NS2B/NS3, NS3/NS4A, andNS4B/NS5 junctionsshareashortsequence
motifnearthecleavage sites (43, 47). These cleavagesfollow
apair of basicaminoacids(RRorKRorRK),or
exception-ally QR at the NS2B/NS3 junction in DEN, and precede either G, S, or A. The cleavage site for processing the C terminus ofanchored C also shares this motif(37, 48).It has
beenspeculatedthatthis class ofprocessingeventsoccursin
thecytoplasmand ismediatedbyavirus-encodedproteinase (43, 47). Recently, two laboratories have proposed that
flavivirus NS3 is a protease on the basis of a limited sequencehomology toserine proteases (1, 21). Four sepa-rateregions of homologywereidentified within the
N-termi-nal 180 amino acids ofNS3;threeencompassedthecatalytic
2467
on November 10, 2019 by guest
http://jvi.asm.org/
triad D, H,S, and the fourth was designated as a substrate-binding domain.
We have initiated studies to analyze the DEN type 4 (DEN4) polyprotein cleavages and to identifyviralfunctions that are required for these proteolytic processing events. Our approach has beeninvivo expressionof DNAfragments that code for regions of the polyprotein of interest, using vaccinia virus as a vector. We previously used this approach toshow that NS2A is required for NS1INS2A cleavage (19). In this report, we describe evidence that NS2B and the proposed protease domain of NS3 are both required for the NS2A/NS2B and NS2B/NS3 cleavages. We show thatNS2B iscapable of acting in trans to effect cleavage at these sites. Indirect evidence is also provided that NS3 can act in trans. In addition, we demonstrate that NS2B is necessary for NS3/NS4Acleavage, as well asforseveral apparently spe-cificcleavages within NS3.
MATERIALS AND METHODS
Cells and viruses. CV-1 cells andTK-141cellsweregrown as monolayers in Eagle minimal essential medium
supple-mented with 10% fetal bovine serum. The vaccinia virus recombinant vSC8 has been described previously (8). Recombinant vaccinia viruses were constructed from
vac-cinia virus WR and plasmid pSC11 derivatives containing DEN4cDNA sequences to be expressed asdescribed
pre-viously (8, 19). The nomenclature of these vaccinia virus recombinants followed previous practice: for example, a
virus derived from pSC11/NS1-NS2A was named vNS1-NS2A, where NS1-NS2A is the portion ofthe DEN4 poly-protein encoded in theconstruct.
Plasmid construction. Standard recombinant DNA tech-niques were used for the construction and propagation of plasmids. The vaccinia virus intermediate transfer vector
pSC11, the derivatives pSC11[BglII] andpSC11/NS1-NS2A, and the sources of the DEN4 cDNA have been described
previously (8, 19, 57). The structures of the DEN4 cDNA
segments cloned into pSC11, encoding various portions of the DEN4polyprotein between NS2A and NS4B, are dia-grammed in Fig. 1. The structures of these plasmids were
verified inallcasesby appropriate restrictionenzyme
diges-tions. Oligodeoxyribonucleotides wereusedtointroducean
initiation codon andatermination codon for NS2A, NS2B, orNS3 andatermination codonatthe EcoRVsite (nucleo-tides [nt] 4748to 4753) orat the BstBI site (nt 5068-5073), creating NS3 C-terminally truncated after 77 amino acids (12%) or 184 amino acids (30%), respectively.
Dideoxynu-cleotidesequencing (46)wasusedtoverifythesequencesin the region of the oligodeoxyribonucleotides. More details about thenucleotide and amino acidsequencesatthe termini of these clones are givenin Table 1. DEN4 nucleotide and
amino acid numbersarefrom Mackowetal. (30) and Zhaoet
al. (57).
Thetwointernal in-frame deletions of NS2Bwerecreated
by polymerase chain reaction. These deletions, ANS2B(10) and ANS2B(20), encode only the N-terminal SerofNS2B fusedtothe last 10orlast20residues ofNS2B, respectively.
Thepolymerase chain reactionproducts (about 600 bp)were
subcloned, and the sequence of the entire amplified region
(approximately nt 4460 to 5080) was determined for both
deletionmutantsandforawild-type clone that hadnotbeen amplified by polymerase chain reaction. The deletion mu-tants and the wild type were found to differ from the published sequencein this regionat four locations: all had
silent basechangesat nt4721 (GtoA)andnt4925(AtoT)
NS2A NS2B NS3 NS4A NS4B
vNS2A-NS2B
vNS2A-NS2B-1 2%NS3
vNS2A-NS2B-30%NS3
vNS2A-ANS2B(1 0)-30%NS3
vNS2A-ANS2B(20)-30%NS3
vNS2B
vNS2B-NS3
m
I lI I
1E
I I I
I
E11
C1)
IEI
Z
I
vNS2B-NS3-NS4A-84%NS4B
v30%NS3
vNS3*
vNS3
vNS3-NS4A-84%NS4B
FIG. 1. Diagrams of DEN4 proteins encoded by recombinant vaccinia viruses.Thediagramonthe topdepicts the linear map of
DEN4nonstructuralproteins NS2A, NS2B,NS3, NS4A, and NS4B (N-terminaltoC-terminal) in the form ofapolyprotein. The cleav-age site betweenneighboring proteins is shown byavertical line. Theshadedarea attheNS4A/NS4Bjunction representsasequence of19hydrophobic amino acids which is thoughttodirect signalase-mediated cleavage between NS4A and NS4B (43, 47). The other diagramsdepictthe DEN4proteins encoded by the vacciniavirus
recombinantslistedattheleft(see also Table 1). Thetwodeletions inNS2Bareindicatedby linesconnecting discontinuousregionsof thepolyprotein. The NS3* protein hasasingle-base deletion result-ing inaframeshift afterapproximately 90% of NS3,resultingina
17-amino-acid C-terminaltailtranslated from thenewframe, repre-sentedbythe shorthorizontal line.
and nonsilentchangesat nt4827(AtoG; resultinginArgto Gly) and nt 4860 (A to G; Ser to Gly). In addition, ANS2B(20)hadabasechangeat nt5050(AtoT; GlutoVal), whichseems tohave beenintroduced by polymerase chain reaction. Notethat the NS3residue changedliesoutside of sequences inNS3 homologous to serine proteases(1, 21).
Analysis of DEN-specific proteins. The procedures for infection of CV-1 cells with recombinant vaccinia virus, preparationof
[35S]methionine-labeled
lysates, immunopre-cipitation with DEN4 hyperimmune mouse ascitic fluid, sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), andfluorographyhave been described previ-ously (19). 35S-labeled lysate of DEN4-infectedLLC-MK2 cellswaspreparedasdescribedpreviously (18)andwasthe giftof MichaelBray.I I I I
I
I I
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.329.559.68.394.2]TABLE 1. Terminal nucleotide sequences ofrecombinant DNA constructs and the predicted N andC termini of expressed polyproteins
Protein Nucleotidesequencea Structure ofterminusb
5'end Nterminus
NS2A . . .
agatotccaccATGGCCGGA,3479.
. . MAG1126..-NS2B . . .agatcccatATGGGATCT4133. . .
MGS1344.
NS3 . . .agatccATGTCA4523. MS1474..
-3'end C terminus
NS2B . . .4515CAAAGAtgaggatct. . . . 1472QR
12%NS3 . . .4746ATGATCtagctaactagggatct. . . . .1549mi
30%NS3 . . 5067TTTCGCtagctagctagcgaa. . . .1656FR
NS3*c .
.6180GGCA11T-TT.
..TTTtag6240.
. . . ..2027GIFTKIGNGASQGKEITKF
NS3 . .6369AGGAAGtaaggatct. . . 209ORK
84%NS4B . . .7437AACCCGGGAATTCTGtga. . . . 2446NPGIL
a Lowercase letters are noncoding sequences, either 5' of the initiating ATG or 3' of the last codon. Capital letters are coding sequences. Underlined coding sequences are the 5'- or3'-mostcodons of the relevant DEN4 protein. Stop codons aredouble underlined. DEN4 nucleotide numbers are those of Mackow et al. (30) and Zhao et al.(57).
b Underlinedamino acids are derived from translation of vector or oligodeoxyribonucleotide sequences orout-of-frametranslation of DEN4 sequences. Amino acid numbersare those of Mackow et al. (30) and Zhao et al. (57).
cThe C terminus of NS3* results from a frameshift in the original NS3 clone. DEN4 nt 6187 is deleted, indicated by -. The resulting protein is colinear with NS3 through amino acid 2028 (555 residues, or90%oofNS3), at which pointtranslationin anew readingframeadds 17 residues unrelated toNS3 to the C terminus before a termination codon is encountered.
RESULTS
DEN4 NS proteins predicted from
alignment
withKuDjin
NSproteins.Processing ofmost DENNS proteins fromthepolyprotein
has beenlittlestudied. The cleavages that define theDEN4NSproteins have been predicted (30, 40) on the basis of alignment of the deduced DEN4 polyprotein se-quence(30)withthatoftheflavivirusKunjin (13), for which the N-terminal amino acid sequences ofmostNS proteins have beendetermined(13,47, 49). This allows prediction of the sizesof the DEN4 NSproteins. These dataare summa-rized in Table2.Identification of NS2B and NS3. Tostudy the proteolytic processing of DEN4 NS proteins, we expressed various segmentsofthe DEN4polyprotein in vivo from clonedDNA sequences, using vaccinia virus as a vector. The DEN4
TABLE 2. PredictedNSproteinsofDEN4 Amino Molecular N-terminal Protein Nucleotidesa acidSa b cleavagesite'
acldsa (kDa)b cevg le
NS1 2421-3476 774-1125 39.6 FTVQA/DMGCV NS2A 3477-4130 1126-1343 24.1 SQVTA/GQGTS NS2B 4131-4520 1344-1473 14.0 GASRR/SWPLN NS3 4521-6374 1474-2091 69.5 VKTQR/SGALW NS4A 6375-6824 2092-2241 16.4 ASGRK/SITLD NS4B 6825-7560 2242-2486 26.5 GLIAA/NEMGL NS5 7561-10262 2487-3386 103 QTPRR/GTGTT
a Nucleotide numbers and amino acid numbersarefromMackow etal.(30) andZhaoetal.(57).
bPredictedmolecular massesarefor theproteinbackboneonly; contribu-tionsfrommodifications (such as glycosylation of NS1) have not been taken intoaccount.
cArbitrarily,five amino acids on either side of eachcleavage site have been listed. Cleavageoccursbetweentheresiduesseparatedbyaslash.The amino acidfollowing the slash is the N-terminal residue of the protein listed on that line.Thefirstamino acidto theleft of each slashonanygivenline is the C terminus ofthe preceding protein, whichis foundonthe line above.The
FTVQAsequenceontheNS1 line is the C terminus ofE.TheC terminus of NS5 is definedbyastop codon.
cDNA sequences coding for the predicted amino acid se-quences of NS2B and NS3 were used to construct the recombinant vaccinia viruses vNS2B and vNS3 (Fig. 1; Table 1). Cellswereinfected withoneof these recombinants orwith
vSC8,
anegative
controlvaccinia virus recombinant thatdoes not contain DEN4 sequences. TheDEN-specific
proteinsexpressed by these cells are shown in Fig. 2. The vNS2B lane containedafaintdoublet band withamolecular massof about12kDa andafewbackground bands thatwere alsoseenin the vSC8 lane. Thelarger memberof the doublet comigrated with aband seen in theDEN4 lane. (Note that the vNS2B lane is overexposed compared with the DEN4 lane.) This 12-kDa band was identified as NS2B, even though the molecularmass of NS2Bwaspredictedtobe 14 kDa (Table 2). Itis noteworthy thatSpeight
etal. (47,48)
found thatKunjin
virus NS2B had an observed molecular mass of10 kDadespite
apredicted
molecularmass of 14 kDa.Theseinvestigators
concludedthat smallhydrophobic
proteins such as NS2B can have anomalous mobilities in SDS-PAGE. Theorigin
ofthesmaller memberof the doublet is not known, but it could be due to internal initiation of translation at the ninthamino acid ofNS2B,
which would reduce thepredicted size of NS2B by1 kDa. NS3ofDEN4 waspredictedtohavemolecularmassof 69.5kDa(Table 2). Aproteinwasobserved in theDEN4lane withanapparent molecularmass of 73 kDa.The recombinantvNS3(lane 5)
expressed aprotein
thatcomigrated
with this 73-kDa pro-tein, and thiswasassignedasNS3. Similarly,Speight
etal. found thatKunjin
virus NS3migrated
slightly
slower than expected (47). Recombinant vNS3 alsoexpressed
asecond proteinslightly smallerthanNS3.This smallerprotein
could have arisenfrominternalinitiation oftranslation, perhaps
atamino acid26 or 42ofNS3, which wouldreduce thesizeof NS3 by 3 or 4.5
kDa,
respectively.
The sequence around amino acid42isparticularly
favorable fortranslation initia-tion.Toobtainfurtherevidence that the 73-kDa bandwas
NS3,
weanalyzed the
proteins produced
by
vNS3*andv30%oNS3.
The C-terminally truncated NS3
products
ofthese viruseson November 10, 2019 by guest
http://jvi.asm.org/
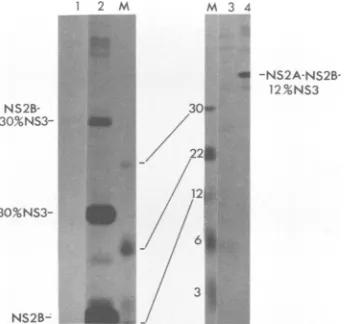
[image:3.612.55.563.98.255.2]1 2 3 4 5 6 DM
E
30%NS3
-prM
18
-NS2B
FIG. 2. Expression of DEN4 NS2B and NS3. Aliquots of
[Simethionine-labeledlysates ofvirus-infected cells were
immu-noprecipitated with DEN4-specific hyperimmune mouse ascitic
fluid, and the precipitates were resolved by SDS-PAGE and
de-tected by fluorography. The positions of DEN4-specific protein
bands are indicated, as are the sizes (in kilodaltons) of the
14C-labeled marker proteins in lane M In lanes 1 to 6, recombinant
vaccinia viruseswere usedtoinfect CV-1 cells 1 vSC8(negative control); 2, vNS2B; 3, v30%NS3;4, vNS3*; 5, vNS3; 6,
vNS2B-NS3. Lane D contains DEN4-infectedLLC-MK2celllysate.Lanes
1to3wereexposedtofilmfor 9days;the other laneswereexposed
for 1 day.
wereexpectedto havemolecular massesof 64 and 20kDa,
respectively.
ThevNS3* lane containedtwoclosely spaced bands, atestimated molecularmassesof 71 and 68 kDa. Thev30%NS3lanecontained faint bandsat23 and 19 kDa(note that this lane is greatly overexposed compared with the vNS3* and vNS3 lanes). Neither vNS3* nor v30%NS3
produced the 73-kDa band thatwas seenin the vNS3 lane. Ourinterpretation of these datawasthat the largerband in eachcasewasthecorrectlyinitiated N53product, while the smallerband.was aberrant andwas perhaps due tointernal initiation of translation. In all cases, the mobilities of the
full-length and truncated N53 species were slightly lower thanexpected.Takentogether,theseresultsprovide genetic
evidence that the 73-kDa band made by DEN4 and vNS3
was NS3.
The viral protease activity lies within NS2B-NS3. Having
identified the DEN4proteinsNS2B andNS3, wewishedto
characterize the viral sequences requiredfor the cleavages
that definethese proteins. Inparticular, we wantedto map
the location ofthe hypothetical viral proteinase. As a first
step toward thisgoal, the products ofrecombinant
vNS2B-Nt
3 (Fig. 1; Table 1) were analyzed. vNS2B-NS3 madeNS2B and
Ni3
(Fig. 2, lane 6). This result mapped thehypothetical virally encoded proteinase responsible for the
NS2B/NS3 cleavage to the NS2B-NS3 region. Such a pro-teinase is presumably also responsible for other cleavages such as NS2AINS2B, N53/NS4A, and NS4BINS5, which
containthe same cleavage sitemotifas
NS2BlNS3.
Inaddition toNS2BandNS3,several other specificbands
were seen in the vNS2B-NS3 lane. The largest of these
bands,
with an apparent molecular mass of86 kDa, had amobilityconsistentwiththatexpectedforuncleaved
NS2B-NS3. Recombinant vNS2B-NS3* expressed a protein which ran slightly faster than this 86-kDa species, as would be expected for an uncleaved NS2B-90%NS3 product (data not shown). This supports the identification of the 86-kDa spe-cies as uncleaved NS2B-NS3. The other specific bands, marked on the side of the figure with asterisks, were smaller than NS3 but larger than
NS2B.
These products must have been derived frominternalcleavages within NS3. A compar-ison of the products made by vNS2B-NS3 infected cells with those made by cells coinfected withvNS2B
plus vNS3 indicated that the internal cleavages within NS3 are greatly facilitated when NS2B is in cis (not shown). Since many of these bands were not present in DEN4-infected cells, they may represent adventitious cleavages within NS3 in the vaccinia virus recombinant expression system. However, the two prominent bands near 50 kDa in lane 6 comigrated with two faint bands in the DEN4 lane, and these species mnay represent authentic products not previously predicted from the N-terminal amino acid sequence data. These 50-kDa bands were faintly visible in the vNS3 and vNS3* lanes upon long exposure, suggesting that they were composed entirely of NS3 sequences. We speculate that theinternal
cleavages in NS3 also occur at sites which share the se-quence motif found at the previously predicted cleavage sites. Six such sites are found in DEN4NS3,
after amino acids 1488, 1501, 1616, 1850, 1931, and 2012. Partial cleav-ages at these sites could generate many possible products.NS3 is required for the NS2A/NS2B and
NS2B/NS3
cleav-ages. The finding that a hypothetical viral protease maps to NS2B-NS3 is consistent with the notion thatNS3
is a protease (1, 21). Since the postulated protease domain includes approximately the N-terminal 180 amino acids ofNS3,
we investigated the processing of vNS2A-NS2B-30%NS3 (Fig. 1; Table 1), which terminates translation after 184 amino acids ofNS3. This recombinant expressedNS2B
and 30%NS3 (Fig. 3, lane 2). Note that vNS2A-NS2B-30%NS3 produced much more30%NS3 and
NS2B
than the corresponding products expressed byv30%NS3
andvNS2B;
the reason for this is not known. A third band with an apparent molecular mass of 38 kDa was also seen in lane 2. This size was consistent with either uncleaved NS2A-NS2B or uncleaved NS2B-30%NS3. However, the recombinant vNS2A-NS2B made a product with an apparent molecular mass of 32 kDa (see Fig. 5), which is presumably uncleaved NS2A-NS2B. The 38-kDa species was thus identified as uncleaved NS2B-30%NS3. Since vNS2A-NS2B-30%NS3 made30%NS3,theNS2B/NS3cleavage occurred. Similarly, the production ofNS2B by this recombinant indicated that cleavage between NS2A and NS2B occurred. This implied that NS2A ought to have been made, although it was not detected. These results are consistent with the hypothesis that the N-terminal 180 residues ofNS3 contains a protease domain.
Todetermine whether the putative NS3 protease domain isnecessary for theNS2A/NS2B andNS2B/NS3 cleavages, we constructed and analyzed recombinant
vNS2A-NS2B-12%NS3
(Fig. 1; Table 1). This virus contains a DEN cDNA segment that encodes an NS3 product truncated in the middle of the protease domain, removing two of the pro-posed regions of homology toserine proteases, including the domain containing the proposed catalytic serine residue; therefore, vNS2A-NS2B-12%NS3 should lack NS3 protease activity. vNS2A-NS2B-12%NS3 made neitherNS2B
nor12%NS3,
but did make a protein with an apparent molecular mass of 45 kDa, which was assigned as uncleaved NS2A-NS2B-12%NS3 (Fig. 3, lane 4). Recombinant v12%NS3,on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.104.262.71.276.2]1 2 M M3 4 1 2 3 D
-0
-NS2A-NS2B-... 12%NS3
NS2B- 30Ow
30%NS3-m.i. 22/
1i../ 22S
30%NS3- 5
* / 3
NS2B- _
FIG. 3. The putative protease domain of NS3 is required for the NS2A/NS2B and NS2B/NS3 cleavages. Samples were treated as described in the legend to Fig. 2. The positions of DEN4-specific bandsareindicated, as are the sizes of the marker proteins in lanes
M. Inlanes1to 4, recombinant vaccinia viruses were used to infect CV-1 cells: 1,vSC8; 2,
vNS2A-NS2B-30%oNS3;
3, vSC8; 4, vNS2A-NS2B-12%NS3. The gelontheleft was12% acrylamide; the one on theright was 17% acrylamide.encoding only the truncated NS3 protein, made no detect-ableproduct(data not shown). Thefailuretodetect12%NS3 could have been due to the lack of specific antibodies or instability ofthe protein. Although the failure to detect a 12%NS3 product from vNS2A-NS2B-12%NS3 was not in-formative, the finding that this virus failed to make NS2B suggested that both
NS2AINS2B
andNS2BINS3
junctions were not cleaved. Furthermore, the uncleaved products NS2A-NS2B and NS2B-12%NS3 were not detected, sug-gestingthat nocleavage had takenplace. These resultsare consistent with thehypothesis thatthe protease domainof NS3 is required for both the NS2A/NS2B and NS2B/NS3 cleavages.NS2B is required for NS3/NS4A cleavage. Evidence that NS2Bis alsorequired fortheproteolytic activity ispresent inFig.4,which showsanalysis oftherecombinants vNS2B-NS3-NS4A-84%NS4B and vNS3-NS4A-84%NS4B (Fig. 1; Table 1). RecombinantvNS2B-NS3-NS4A-84%NS4B made both NS2B and NS3, indicating that the NS2B/NS3 and NS3/NS4A cleavages occurred. In addition, there was a series of other bands in this lane. Many of these bands (asterisks) were smaller than NS3, and their identities are discussed above(Fig. 2). Threebands, designatedc,d,and e,werelargerthanNS3. Bande waspreviously assignedas uncleaved NS2B-NS3. The sizes of bands c and d were consistent with uncleavedNS2B-NS3-NS4A-84%NS4B and NS2B-NS3-NS4A, respectively. The existence ofthe latter species would imply that NS4A/NS4B cleavage had oc-curred. In contrast, the vNS3-NS4A-84%NS4B lane con-tained neithermature-sized NS3noranyof the other bands seenin thevNS2B-NS3-NS4A-84%NS4Blane. These obser-vations indicate that NS2B has a role in the NS3/NS4A cleavage and suggestsaroleforNS2B in theNS2A/NS2B,
NS2B/NS3,
andNS4BINS5
cleavages, sincethese all share thesameaminoacid sequencemotifatthecleavagesite. The vNS3-NS4A-84%NS4B lane had doublet bands at 85 kDa (labeled b) and at 105 kDa (labeled a). These sizes were consistent with uncleaved NS3-NS4A andNS3-NS4A-84%NS4B,
respectively. This assignmentwould suggest thata-
b--c
-d
-e
-NS3 -E
-NS1
-prM
_-NS2B
FIG. 4. NS2B isrequired for NS3/NS4A cleavage. Sampleswere
treatedasdescribed in thelegendtoFig. 2. The positions ofsome
DEN4-specific bandsareindicated. The asterisks and lettered bands indicate otherspecific products discussed in thetext. Inlanes1 to3, recombinant viruses were used to infect CV-1 cells: 1, vSC8; 2, vNS3-NS4A-84%NS4B; 3, vNS2B-NS3-NS4A-84%NS4B. Lane D
contains DEN4-infectedLLC-MK2cellslysate.
NS4A/NS4B cleavage did notrequire NS2B, which agrees with
the
proposal that this cleavage is mediated by signal peptidase(43,47).Deletions of NS2B are defective for cleavage attheNS2A/ NS2B and NS2B/NS3 junctions. To test whether NS2B is required fortheNS2A/NS2BandNS2B/NS3 cleavages,we constructed the recombinants
vNS2A-ANS2B(10)-30%NS3
and vNS2A-ANS2B(20)-30%NS3 (Fig. -1; Table 1). These twoviruseswere similar tovNS2A-NS2B-30%NS3, except that eachcontainsalarge in-frame deletioninNS2B, retain-ing onlytheN-terminal serine residueand theC-terminal10 or 20 amino acids of NS2B. Both vNS2A-ANS2B(10)-30%NS3 (Fig. 5, lane 1) andvNS2A-ANS2B(20)-30%oNS3
(Fig. 5, lanes 2, left and right) made no 30%NS3 but expressedproductswithmolecularmassesofapproximately
45kDa,aspredicted foruncleavedNS2A-ANS2B-30%NS3. Consistent withthis
assignment
is the observation thatthe product ofvNS2A-ANS2B(20)-30%oNS3
wasslightly larger
than that of
vNS2A-ANS2B(10)-30oNS3.
In lane 2(left
side), the band at approximately 60 kDa(unlabeled
tick mark) might notbe DENspecific,
since this band was not seeninotherexperiments inwhich thesamevirus stockwas used (see, for example, lane 2,right side).
The failure toobserve products at or near the position of 30%NS3 indi-cated that neither NS2B/NS3 nor NS2A/NS2B
cleavage
occurred with these NS2B deletionmutants.NS2B exhibitsatrans-actingfunction.TheNS2Bdeletions described abovecontainedthe last 10or20residuesattheC terminus ofNS2B, whose lengths most
probably kept
the NS2B/NS3cleavagerecognition
sequenceintact.Thefailure of these NS2B deletionstocleaveNS2B/NS3
wastherefore probably due to the loss ofaspecific
NS2B function. The samemightnotbetruefortheNS2A/NS2Bcleavage,
sinceon November 10, 2019 by guest
http://jvi.asm.org/
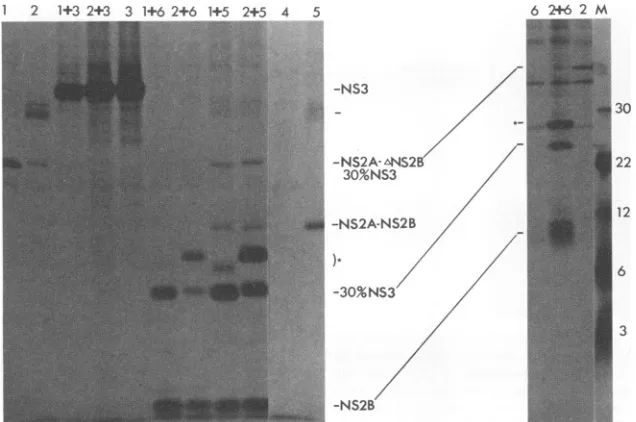
[image:5.612.93.265.74.236.2] [image:5.612.391.480.75.315.2]1 2 1+3 2+3 3 1+62+6 1+5 2+5 4 5
-NS3
-NS2A-ANS2B 30%NS3
-NS2A-NS2B
-30%NS3
-NS2B
30
22
12
[image:6.612.155.473.76.287.2]6
FIG. 5. Demonstration that NS2B acts in trans. Samples were treated asdescribed in the legend to Fig. 2. The positions of some
DEN4-specific bands areindicated, as are the sizes ofthemarkerproteins in laneM.The unlabeledtick mark and the asterisk indicatebands discussed further in the text. In lanes 1 to 6, recombinant viruses were used to infect CV-1 cells: 1,
vNS2A-ANS2B(10)-30%NS3;
2, vNS2A-ANS2B(20)-30%NS3;3,vNS3; 4, vSC8; 5,vNS2A-NS2B; 6, vNS2B.Laneswithtwonumberswerecoinfections with both indicated viruses; for example, lane 1+3 was infectedwithvNS2A-ANS2B(10)-30%NS3 plus vNS3. Thegelontheleftwas 12%acrylamide;thegelontheright was17% acrylamide.
onlythefirstresidueof NS2Bwasretained.Failuretocleave atNS2A/NS2B could thereforehave been due todestruction of the cleavage site; however, see below.
Tofurtherinvestigatethe roleofNS2B in thesecleavages, we performedcomplementationexperiments bycoinfection of cells withtworecombinantviruses.Vaccinia virus recom-binant vNS2A-ANS2B(10)-30%NS3 or vNS2A-ANS2B(20)-30%NS3wasusedasthesourceof uncleaved precursor, and vNS2B, vNS3, or vNS2A-NS2B was used as a possible source of complementing activity. Both deletion mutants were partiallycomplemented forNS2B/NS3 cleavageupon coinfectionwith vNS2B(Fig.5, lane 1+6 and lanes 2+6, left andright) orvNS2A-NS2B (lanes 1+5 and 2+5), as shown by the appearance of 30%NS3 and the reduction in the amount of uncleaved precursor. No such complementing activity was seen upon coinfection with vNS3 (lanes 1+3 and 2+3). Thus, NS2B can act in trans to complement the defect inNS2B/NS3 cleavage caused by deletionof NS2B. These results showed that the target site for NS2B/NS3 cleavage does not require more than the 10 C-terminal residues of NS2B. Furthermore, this demonstrated that providingtheputativeNS3protease intrans (assumingthat the NS3 made by vNS3 is an active protease) failed to correct the cleavage-defective phenotype of these NS2B deletionmutants.Since a multiplicity of infection of up to 10 PFU of eachinfecting virus per cell was used, it was highly unlikelythat the observed incomplete complementation was due toasignificant fraction of cells that were not coinfected. Rather,theincompletecomplementation was perhaps due to failure of the DEN protein products to reach the same intracellularsitequantitatively. The 30%NS3 bands in lanes 2+5and 2+6 migrated slightly more slowly than the corre-sponding bands in lanes 1+5 and 1+6. The difference in apparentmolecular mass was only 0.25 kDa. We attributed this smalldifference totheGlu-to-Valchangeatamino acid 1650 in the 30%NS3 derived from
vNS2A-ANS2B(20)-30%NS3. However, we cannot rule out other possible ex-planations, such as slightly imprecise NS2B/NS3 cleavage. Thereappeared to be more 30%NS3 product upon comple-mentation than uncleaved precursor in the absence of com-plementation. We assume that this was due to greater stability or more efficient immuneprecipitation of the prod-uct than the precursor. Also note that more NS2B was presentin the coinfected lanes than with vNS2B alone (lane 6). We have notinvestigated this phenomenon further.
In addition to the 30%NS3 band seen when each of the NS2B deletion mutants was complemented by vNS2B or vNS2A-NS2B, another band slightly larger than 30%NS3 was seenineachof these lanes(asterisk). The second band in the vNS2A-ANS2B(20)-30%NS3 coinfections was larger than that in the vNS2A-ANS2B(10)-30%NS3 coinfections. Thissuggested that the second bandswereeither uncleaved ANS2B-30%NS3orNS2A-ANS2B. Theuseof monospecific seraforeither NS2A or NS3 should resolve theidentity of these second bands.Althoughwecouldnotbe certain of the identity of these species, the fact that we have never detected NS2Asuggested that thesewereprobably /NS2B-30%NS3. If this is the case, itimplies that theNS2A/NS2B cleavage also was complemented in trans by NS2B in the coinfections and that the NS2A/NS2B cleavage site se-quence does not extend beyond the N-terminal serine of NS2B.
NS3 mayactin trans tocleaveNS2A/NS2B.The results in Fig. 5 alsoprovidedindirectevidencethatNS3 couldactin trans. A band was seen in the vNS2A-NS2B lane with a molecularmassof 32kDa,whichwaspresumably uncleaved NS2A-NS2B. This precursorwasexpectedtohavea molec-ularmassof 38kDa,butasdiscussedabove,themobilityof NS2Bwas knownto beaberrant. Note also that noNS2B was seen in this lane. However, upon coinfection with vNS2A-ANS2B(10)-30%NS3 (lane 1+5) orvNS2A-ANS2B (20)-30%NS3 (lane 2+5), a strong NS2B signal was
on November 10, 2019 by guest
http://jvi.asm.org/
duced. ThisNS2Bmust have been derived from cleavage of the NS2A-NS2B product ofvNS2A-NS2B, since the other viruses in the coinfections did not encode NS2B. This provides evidence that the 30%NS3 encoded by vNS2A-ANS2B(10)-30%NS3 and
vNS2A-ANS2B(20)-30oNS3
was abletocleave theNS2A/NS2Bjunction in trans. The appar-ent excess production of NS2B product compared with NS2A-NS2Bprecursor was presumed to be due to differen-tialstability orimmunoprecipitability. Incontrast, attempts to complement vNS2A-NS2B with vNS3 orv30%NS3 re-sulted inlittleor noNS2B production (data not shown). One possible explanation forthelackofcomplementation is that the products of vNS2A-NS2B and of vNS3 or v30%NS3 failedtoreach the samespecificintracellular site.Finally,it was noteworthy that uncleaved NS2A-NS2B was not ex-pressedbyvNS2A-NS2B-30%NS3(Fig. 3), although a band correspondingtoNS2B-30%NS3was seen. This observation suggests that NS2A/NS2B cleavage preceded NS2B/NS3 cleavage. Whether this implies ahighly regulated cleavage mechanismorisan adventitious result remains to be deter-mined.DISCUSSION
We have constructed recombinant vaccinia viruses ex-pressing various portions of the DEN4 genome to study proteolytic processing ofthe NS region of the DEN4 poly-protein. Our results indicate that the N-terminal 30% of NS3 is required for the NS2A/NS2B and NS2B/NS3 cleavages. Indirect evidencethatNS3 can actintrans atthese cleavage sites was
obtained
from the results ofthe coinfections of vNS2A-NS2BplusvNS2A-ANS2B(10)-30%NS3 or vNS2A-ANS2B(20)-30%NS3 (Fig. 5), where NS2B was produced. However, attemptstodirectly demonstratetransactivityof NS3 by coinfections with vNS2A-NS2B or vNS2A-NS2B-12%NS3 andvNS3 orv30%NS3 have failed. Nevertheless, thesefindings are consistent with the proposed modelthat NS3hasproteaseactivity(1, 21). Two groupshaverecently described results of in vitro translation of transcripts of clonedDNAthatsuggest that theputativeprotease domain ofNS3 isrequired for both the NS2A/NS2BandNS2B/NS3 cleavages (11, 41).Inaddition, ourresultsdemonstrate that NS2B is alsorequired forthe NS2A/NS2B and NS2B/NS3 cleavages. Complementation experiments show that NS2B can act in trans to facilitate NS2B/NS3 cleavage and also probably NS2A/NS2B cleavage. Thepossibilitythat homol-ogous recombination efficiently generated NS2B in cis in theseexperiments is unlikely because ofthe shortoverlap of 30 or 60nucleotidesbetweenNS2Band the NS2Bdeletions and the short duration of the experiments. Furthermore, other studiesin ourlaboratoryhave shown that both NS2B andNS3 can actintrans tofacilitateNS4B/NS5 cleavage,in anexperiment in which homologous recombinationwasnot a possibility (5a). Also, Chambers et al. have recently reported observations that NS2B isrequired for NS4B/NS5cleavage
in vitro (11). Thus, both NS2B and NS3 together appear to be required for a protease activity that cleaves NS2A/NS2B, NS2B/NS3, andNS4BINS5.
The flavivirus protease activity is presumably also responsible for the NS3/NS4A cleavage,andperhapstheC-terminalprocessing ofC(43,47). Wehavenodataregardingtherequirementfor NS3 for either ofthese processing events, but our results show that NS2B is needed for theNS3/NS4Acleavage.This activity of NS2B is readily demonstrated when NS2B is present in cis, but this cleavage activity is absent or very inefficientwhenNS2B issuppliedin trans(datanotshown).NS4A/NS4B cleavage seems to occur in the absence of NS2B, consistent with the suggestion that this cleavage is mediated by signalase (43, 47). Finally, the recombinant vNS2B-30%NS3 made a product of a size consistent withan uncleaved NS2B-30oNS3 precursor, but did not produce uncleaved NS2A-NS2B. This suggests that NS2A/NS2B cleavage preceded NS2B/NS3cleavage.
It is interesting that NS3 alone appears to exhibit some limitedproteolytic activity, as vNS3 makes minor amounts of some of theNS3 internal cleavage products. Coupled with the findingthatboth NS2B and NS3 are requiredand each canapparentlyactintrans tomediatetheNS2A/NS2B and NS2B/NS3cleavages, this suggests that NS2B interacts with NS3 in some fashion to promote the protease activity inherent in NS3. Such a two-component proteinase is appar-ently required for at least one of theprocessingevents in the comovirus M segment polyprotein (28). One possibility is that these twoproteins associate as a complex to form the activeproteinase. The observation that both NS2B and NS3 together seem to immunoprecipitate better than when they arepresentalone isconsistent with the presence of suchan NS2B/NS3 complex. A related possibility is that NS2B is needed to target NS3 to the correct cellular location for maximal proteolyticactivity. NS3is known to be membrane associated (23, 52), despitelackingany obvious membrane-spanning hydrophobic segment. It is conceivable that the correctmembraneassociation of NS3 depends on an inter-action between NS3 and the highly hydrophobic NS2B. Consistent with this idea are thepreliminary results ofcell fractionation studies which show that NS3 made by vNS2B-NS3 is entirely membrane associated, while about one-quarter of the NS3 madeby vNS3 is soluble (17a). It is also possible that NS2B acts in trans to modify NS3 in some mannertoincrease theproteolytic activityinherent in NS3. An alternate explanation forthe observed trans activity is thatNS2B is aprotease. We consider this unlikelyfor the followingreasons.First,wedetectednocleavage activityin recombinants, such as vNS2A-NS2B and vNS2A-NS2B-12%NS3,thatcontain intact NS2B in theabsence of the NS3 protease domain. Second, it would seem genetically uneco-nomicalfor DEN to code forfunctionallyredundant prote-ases. Third, sequence homology toproteases has not been found in NS2B. Finally, it is possible that the role of NS2B is to maintain the polyprotein precursor in a conformation cleavablebyNS3. One would expect NS2Btobecis-acting in thiscase, which is consistent with the data forNS3/NS4A cleavage. However, trans activity is possible in such a model. For example, the NS2A-ANS2B-NS3 precursor could be defective for cleavage as a result of misfolding causedby deletion ofNS2B, and
providing
NS2B in trans might allow cleavageto occurifNS2B caninteractwith the incorrectly folded molecule and restore it to a cleavable conformation. Althoughwecannot ruleoutthispossibility, it seems lesslikelythanthesimplertwo-component protease model, whereby NS2B functions to activate the proteaseactivity
of NS3. Experiments are in progress to further characterize the role of NS2B in the flavivirus protease activity.ACKNOWLEDGMENTS
WethankMyronHill and PeterCollins forthe synthesisofthe oligodeoxyribonucleotides,RobertChanockforacriticalreadingof
themanuscript,and ToddHeishmanand SandraChangfor editorial assistance.
on November 10, 2019 by guest
http://jvi.asm.org/
REFERENCES
1. Bazan, J. F., and R. J. Fletterick. 1989. Detection of a trypsin-like serine protease domain in flaviviruses and pestiviruses. Virology 171:637-639.
2. Bell, J. R., R. W. Kinney, D. W. Trent, E. M. Lenches, L. Dalgarno,and J. H. Strauss. 1985. Amino-terminal amino acid sequences ofstructural proteins of threeflaviviruses. Virology 143:224-229.
3. Biedrzycka,A.,M. R. Cauchi, A. Bartholomeusz, J. J. Gorman,
andP. J. Wright. 1987. Characterization of protease cleavage sites involved in the formation of the envelope glycoprotein and
three non-structural proteins of dengue virus type 2, New GuineaC strain.J. Gen. Virol. 68:1317-1326.
4. Boege, U., F. X. Heinz, G. Wengler, and C. Kunz. 1983. Amino acid compositions and amino-terminal sequences of the
struc-tural proteinsof aflavivirus, European tick-borne encephalitis virus. Virology 126:651-657.
5. Boulton, R. W., and E. G. Westaway. 1977. Togavirus RNA: reversible effect of ureaon genomes and absence of subgenomic
viralRNA inKunjin virus-infectedcells. Arch. Virol. 55:201-208. 5a.Cahour, A., B. Falgout, and C.-J. Lai. Unpublished
observa-tions.
6. Castle, E., U.Leidner, T. Nowak, G. Wengler, and G. Wengler.
1986. Primary structure of the West Nile flavivirus genome regioncodingfor all nonstructural proteins. Virology 149:10-26.
7. Castle, E., T.Nowak,U.Leidner, G. Wengler, and G. Wengler.
1985. Sequence analysis of the viral core protein and the membrane-associated proteins Vi and NV2 of the flavivirus
WestNile virusandofthe genome sequence for these proteins. Virology145:227-236.
8. Chakrabarti,S., K.Brechling, and B. Moss. 1985. Vaccinia virus expression vector: coexpression of P-galactosidase provides visualscreeningofrecombinant virus plaques. Mol. Cell. Biol. 5:3403-3409.
9. Chambers, T. J., D. W.McCourt, and C. M. Rice. 1989. Yellow fevervirusproteins NS2A, NS2B, and NS4B: identification and partialN-terminal amino acid sequence analysis. Virology 169: 100-109.
10. Chambers, T. J., D. W. McCourt, and C. M. Rice. 1990. Production of yellow fever virus proteins in infected cells: identification of discrete polyprotein species and analysis of cleavage kinetics using region-specific polyclonal antisera. Vi-rology 177:159-174.
11. Chambers,T.J., R. C. Weir, A.Grakoui, D. W. McCourt, J. F.
Bazan, R. J.Fletterick, and C. M. Rice. 1990. Evidence that the N-terminal domain ofnonstructural protein NS3 from yellow fever virus is a serine protease responsible for site-specific cleavagesinthe viralpolyprotein. Proc. Natl. Acad. Sci. USA 87:8898-8902.
12. Cleaves, G. R., and D. T. Dubin. 1979. Methylation status of intracellular dengue type 240S RNA. Virology 96:159-165.
13. Coia, G., M. D. Parker, G. Speight, M. E. Byrne, and E. G. Westaway. 1988. Nucleotide and complete amino acid se-quencesofKunjin virus: definitive gene order and characteris-ticsof thevirus-specified proteins. J. Gen. Virol. 69:1-21.
14. Dalgarno, L., D. W.Trent, J. H. Strauss, and C. M. Rice. 1986. Partial nucleotide sequence ofthe Murray Valley encephalitis
virus genome: comparison ofthe encoded polypeptides with yellow fever virus structural and non-structural proteins. J.
Mol. Biol. 187:309-323.
15. DeMadrid, A. T., and J. S. Porterfield. 1974. The Flaviviruses
(group B Arboviruses): a cross-neutralizing study. J. Gen.
Virol.23:91-96.
16. Deubel, V., R. M. Kinney, and D. W. Trent. 1986. Nucleotide
sequence and deduced amino acid sequence of the structural proteins ofdengue type 2 virus, Jamaica genotype. Virology 155:365-377.
17. Deubel, V., R. M.Kinney,and D. W. Trent. 1988. Nucleotide sequenceanddeducedaminoacid sequence of the nonstructural proteinsofdenguetype 2virus, Jamaica genotype: comparative analysisof thefull-length genome. Virology 165:234-244. 17a.Falgout,B. Unpublished observations.
18. Falgout, B., M. Bray, J. J. Schlesinger, and C.-J. Lai. 1990.
Immunizationof micewith recombinantvaccinia virus express-ing authentic dengue virus nonstructural protein NS1 protects against lethal dengue virusencephalitis. J. Virol. 64:4356-4363. 19. Falgout,B., R. Chanock, and C.-J. Lai. 1989. Properprocessing of dengue virus nonstructural glycoprotein NS1 requires the N-terminal hydrophobic signal sequence and the downstream nonstructural protein NS2a. J. Virol. 63:1852-1860.
20.
Falgout,
B., and C.-J.Lai.
1990. Synthesis of dengue virus nonstructural protein NS1 requires the N-terminal signalse-quence and the downstream nonstructural protein NS2A, p. 192-195.In M. A. Brinton and F. X. Heinz (ed.), New aspects of positive-strand RNAviruses. AmericanSociety for Microbi-ology, Washington, D.C.
21. Gorbalenya, A. E., A. P.Donchenko, E. V. Koonin, and V. M.
Blinov.
1989.N-terminal domains ofputative helicases of Flavi and Pestiviruses may be serine proteases. Nucleic Acids Res.17:3889-3897.
22. Gruenberg, A., W. S. Woo, A. Biedrzycka, and P. J. Wright.
1988. Partial nucleotide sequence and deduced amino acid sequence of the structural proteins of dengue virus type 2,New
Guinea C and PUO-218 strains. J. Gen. Virol. 69:1391-1398. 23. Grun, J. B., and M. A. Brinton. 1987. Dissociation of
NS5
fromcell fractions containing West Nile virus-specific polymerase activity. J. Virol. 61:3641-3644.
24. Hahn, Y. S., R. Galler, T.Hunkapiller, J. M.
Dalrymple,
J. H.Strauss, and E. G. Strauss. 1988. Nucleotide sequence of dengue 2 RNA and comparison of the encoded proteins with thoseof otherflaviviruses. Virology 162:167-180.
25. Hashimoto, H., A. Nomoto, K. Watanabe, T. Mori, T. Takezawa, C. Aizawa, T. Takegami, and K. Hiramatsu. 1988.
Molecular cloning and complete nucleotide sequence of the genome of Japanese encephalitis virus
Beijing-1
strain. Virus Genes 1:305-317.26. Hori, H., and C.-J. Lai. 1990. Cleavage of dengue virus NS1-NS2A requires an octapeptide sequence at the C terminus of NS1. J. Virol. 64:4573-4577.
27. Irie, K., Y. Sasaguri, R. Putnak, and R. Padmanabhan. 1989.
Sequence analysis of cloned dengue virus type 2 genome (New Guinea-C strain). Gene 75:197-211.
28. Krausslich, H.-G., and E.
Wimmer.
1988. Viral proteinases. Annu. Rev. Biochem. 57:701-754.29. Lee, E., C. Fernon, R. Simpson, R. C. Weir, C. M. Rice, and L.
Dalgarno. 1990. Sequence of the 3' half of the Murray Valley encephalitis virus genome and mapping of the nonstructural
proteins NS1,NS3, andNS5. Virus Genes 4:197-213.
30. Mackow, E., Y. Makino, B. Zhao, Y.-M. Zhang, L. Markoff, A. Buckler-White, M. Guiler, R. Chanock, and C.-J. Lai. 1987. The nucleotide sequence of dengue type 4 virus: analysis of genes coding for nonstructural proteins. Virology 159:217-228. 31. Mandl,C. W., F. X. Heinz, and C. Kunz. 1988. Sequence of the
structural proteins of
tick-borne
encephalitis virus(Western
subtype) and comparative analysis with other Flaviviruses. Virology 166:197-205.32. Mandl, C. W., F. X. Heinz, E.
Stockl,
and C. Kunz. 1989. Genome sequence oftick-borne
encephalitis virus(Western
subtype) and comparative analysis of nonstructural proteins with other Flaviviruses. Virology 173:291-301.33. Markoff, L. 1989. In vitro processing of dengue virus structural proteins: cleavage of the pre-membrane protein. J. Virol. 63: 3345-3352.
34. McAda, P. C., P. W. Mason, C. S.
Schmaljohn,
J. M.Dalrymple,
T. L. Mason, and M. J. Fournier. 1987. Partial nucleotide sequence of the Japanese encephalitis virus genome. Virology 158:348-360.35. Monath, T. P. 1985. Flaviruses, p. 955-1004. In B. N. Fields, D. M. Knipe, R. M. Chanock, J. L. Melnick, B. Roizman, and R. E. Shope (ed.), Virology. Raven Press, New York. 36. Naeve, C. W., and D. W. Trent. 1978. Identification of Saint
Louis encephalitis virus mRNA. J. Virol. 25:535-545. 37. Nowak, T., P. M. Farber, G. Wengler, and G. Wengler. 1989.
Analyses of the terminal sequences of West Nile virus structural proteins and of the in vitro translation of these proteins allow the proposal of a complete scheme of the proteolytic cleavages
on November 10, 2019 by guest
http://jvi.asm.org/
involved in their synthesis. Virology 169:365-376.
38. Osatomi, K., and H. Sumiyoshi. 1990. Complete nucleotide sequenceof dengue type 3 virus genome RNA. Virology 176: 643-647.
39. Pletnev, A. G., V. F. Yamshchikov, and V. M. Blinov. 1986. Tick-borne encephalitis genome: the nucleotide sequence cod-ing for virion structural proteins. FEBS Lett. 200:317-321. 40. Pletnev, A. G., V. F. Yamshchikov, and V. M. Blinov. 1990.
Nucleotide sequence of the genome and complete amino acid sequence of the polyprotein oftick-borne encephalitis virus. Virology174:250-263.
41. Preugschat, F., C.-W. Yao, and J. H. Strauss. 1990. In vitro processing ofdengue virus type 2 nonstructural proteins NS2A, NS2B, and NS3. J. Virol. 64:4364-4374.
42. Randolph, V. B., G. Winkler, and V. Stollar. 1990. Acidotropic amines inhibit processing of flavivirus prM protein. Virology 174:450-458.
43. Rice, C., E. M. Lenches, S. R. Eddy, S. J. Shin, R. L. Sheets, and J. H.Strauss. 1985. Nucleotide sequence of yellow fever virus: implications for flavivirus gene expression and evolution.
Sci-ence229:726-733.
44. Rice, C. M., R. Aebersold, D. B. Teplow, J. Pata, J. R. Bell, A.V. Vorndam, D. W. Trent, M. W. Brandriss, J. J. Schles-inger, and J.H. Strauss. 1986. Partial N-terminal amino acid sequencesof three nonstructuralproteins of two flaviviruses. Virology 151:1-9.
45. Ruiz-Linares, A., A. Cahour, P. Despres, M. Girard, andM.
Bouloy. 1989.Processing of theyellowfevervirus polyprotein: role of cellular proteases in the maturation of the structural proteins. J. Virol. 63:4199-4209.
46. Sanger, F., S. Nicklen,and A. R.Coulson. 1977. DNA sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
47. Speight, G.,G.Coia,M. D.Parker,and E.G.Westaway. 1988. Gene mapping andpositive identification ofnon-structural pro-teinsNS2a, NS2b, NS3, NS4b, and NS5 of the flavivirus Kunjin and theircleavage sites.J. Gen. Virol. 69:23-34.
48. Speight, G., and E. G. Westaway. 1989. Carboxy-terminal analysis of nine proteins specified by the flavivirus Kunjin: evidence thatonly the intracellularcoreprotein is truncated.J.
Gen. Virol. 70:2209-2214.
49. Speight, G.,and E.G.Westaway. 1989. Positive identification of NS4A, the last of the hypothetical nonstructural proteins of Flaviviruses. Virology 170:299-301.
50. Sumiyoshi, H., C. Mori, I. Fuke, K. Morita, S. Kuhara, J. Kondou, Y. Kikuchi, H. Nagamatu, and A. Igarashi. 1987. Complete nucleotide sequence of the Japanese encephalitis virusgenome RNA.Virology 161:497-510.
51. Svitkin, Y. V., T. Y. Ugarova, T. V. Chernovskaya, V. N.
Lyapustin,V. A. Lashkevich,and V. I.Agol. 1981. Translation of tick-borne encephalitis virus (Flavivirus) genome in vitro: synthesis oftwostructuralpolypeptides. Virology 110:26-34. 52. Takegami,T.,andS. Hotta. 1989. In vitro synthesis ofJapanese
encephalitis virus(JEV) RNA: membrane and nuclear fractions ofJEV-infected cells possess high levels of virus-specific RNA polymerase activity. Virus Res. 13:337-350.
53. Wengler, G., E. Castle,U.Leidner, T.Nowak, and G.Wengler. 1985. Sequence analysis of the membrane protein V3 of the flavivirus West Nile virus and of its gene.Virology 147:264-274. 54. Wengler, G., G.Wengler, and H. J. Gross. 1978. Studiesonthe virus-specific nucleic acids synthesized in vertebrate and
mos-quito cells infected withflaviviruses. Virology 89:423-437. 55. Westaway, E. G., M. A. Brinton, S. Y. Gaidamovich, M. C.
Horzinek, A.Igarashi, L. Kaariainen, D. K. Lvov, J. S. Porter-field, P. K. Russell, and D. W. Trent. 1985. Flaviviridae. Intervirology 24:183-192.
56. Wright,P.J.,M.R.Cauchi,andM. L.Ng. 1989. Definition of the carboxy termini of the three glycoproteins specified by dengue virus type 2.Virology 171:61-67.
57. Zhao, B., E. Mackow, A. Buckler-White, L. Markoff, R. M. Chanock, C.-J. Lai, and Y. Makino. 1986. Cloning full-length dengue type 4 viral DNA sequences: analysis of genes coding
for structural proteins. Virology 155:77-88.