0022-538X/87/061765-07$02.00/0
CopyrightC 1987, AmericanSocietyforMicrobiology
DNA-Dependent
RNA
Polymerase Subunits
Encoded within
the
Vaccinia Virus Genome
ELAINE V. JONES,t CARMIE PUCKETT,t AND BERNARDMOSS*
Laboratory of ViralDiseases, NationalInstituteof Allergy and Infectious Diseases, Bethesda, Maryland 20892
Received 24 December 1986/Accepted 22 February 1987
Antiserumtoamultisubunit DNA-dependent RNA polymerase fromvacciniavirionswaspreparedtocarry
outgenetic studies. This antiserum selectivelyinhibited theactivity of the viral polymerase but hadnoeffecton
calfthymus RNA polymeraseII. The specificity oftheantiserum wasfurther demonstrated by
immunopre-cipitation ofRNApolymerase subunits from dissociatedvirusparticles. Thepresencein vaccinia virus-infected
cells ofmRNA thatencodesthepolymerasesubunitswasdetermined by in vitro translation.
Immunoprecipi-tablepolypeptides withMrs ofabout 135,000, 128,000, 36,000, 34,000, 31,000, 23,000, 21,000, 20,000, and
17,000weremade when early mRNAwasaddedtoreticulocyteextracts.The subunitswereencoded within the
vaccinia virusgenome, asdemonstratedbytranslationof early mRNA that hybridizedtovaccinia virus DNA.
The locationsof the subunitgenes weredetermined initially by hybridization of RNAtoaseriesof overlapping
40-kilobase-pair DNA fragments that were cloned in a cosmidvector. Further mapping was achieved with clonedHindIIIrestriction fragments.Resultsof these studiesindicated that RNA polymerase subunitgenes are
transcribedearlyininfectionandaredistributed within thehighly conserved central portion of the poxvirus genomeinHindIIl fragmentsE,J, H, D, and A.
Eucaryotic RNApolymerasesarecomplex enzymes
con-taining multiplesubunits (6, 34). Class II RNA polymerases, which are responsible for the synthesis ofmRNA and are
isolated from diverse groups ofeucaryotes including
mam-mals(18, 22, 36), amphibians (33), insects (12), higher plants (15),andyeast(9), display strikinglysimilar subunit
compo-sitions. Typically, there are two large subunits that are greater than 100,000 daltons and eight or more small ones ranginginsizefrom 10,000to40,000 daltons. Immunological cross-reactivity between subunits of different species (13, 14, 19, 20, 37, 39)suggeststheevolutionaryconservation of structureand function.
AlthoughDNAvirusesgenerallyusehostRNA polymer-asefortranscription, poxvirusesmaybe notableexceptions.
A DNA-dependent RNA polymerase ispresent in vaccinia
virus particles and the cytoplasm of infected cells (17, 27).
The purified enzymeis similar in overall size to eucaryotic
RNA polymerase and also contains numerous subunits (2, 38). Two large polypeptides of 130,000 to 140,000 daltons andatleastsevensmalleronesranginginsizefrom13,000to 36,000daltons have been described. An additional
polypep-tide of77,000 daltons was associated with the RNA poly-merasethatwasisolatedfrominfectedcells(28). Functional as well as structural similarities exist between vaccinia
virus-associated and eucaryotic class IIRNApolymerases.
Neither purified enzyme can transcribe double-stranded DNAefficiently,and bothrequire single-strandedDNA and
Mn2+ for optimal activity (2, 28, 38). In each case, the correct initiation oftranscription occurs only in crude sys-temsorwhen thepolymerase issupplemented with protein
factors(10, 11, 31, 35).
*Correspondingauthor.
t Present address: Smith Kline & French Laboratories, Swedeland, PA 19151.
tPresent address: BiologyDivision, 147-75CaliforniaInstitute of Technology, Pasedena, CA91125.
Acentral question with regardtothe biology of poxviruses concernsthenatureof the RNApolymerasethat ispackaged within the virus particle. The virus-associated polymerase
differs from thatof the hostinchromatographic properties, electrophoretic mobilities of the component polypeptides, and resistance to a-amanitin (2, 28, 38). These differences
could result from the virus-induced modification of the cellular polymerase that are analogous to those that occur
after infection with bacteriophage T4 (5). Alternatively,
some or all of the subunits could be virus-specific. In this
study we undertooktodetermine which ofthe polypeptides
associated withpurifiedvacciniavirus RNApolymerase are
virus-encoded. In thisreportwedemonstrate thatgenesfor at leasteight subunits arelocated within thecentral, highly
conservedregionofthepoxvirusgenome.Elsewhere(4),we have reported that the largest RNA polymerase subunit of
vaccinia virushasconsiderablesequencehomologywith the
corresponding RNA polymerase subunits of Escherichia
coli, Saccharomyces cerevisiae, and Drosophila melano-gaster.
MATERIALS ANDMETHODS
Preparation of antiserum. RNA polymerase was purified
fromvaccinia virus particles asdescribedpreviously (2) by
using successive columns ofDEAE-cellulose, DEAE-Bio-Gel (Bio-Rad Laboratories, Richmond, Calif.), phospho-cellulose, and aminopentyl agarose. Approximately 100 ,ug
of purified enzyme was emulsified with complete Freund adjuvant and injected into a rabbit. Thirty days later, the animal received the first ofthreeweeklyboosterinoculations of 50 ,ugofpurifiedenzymeinincomplete Freundadjuvant. Bleedingswerecarriedout7daysaftereach booster inocu-lation.
Preparation of [35S]methionine-labeled RNA polymerase. Approximately 2.5 x 109 HeLa cells were infected with 30 PFU of vaccinia virus per cell in 500 ml ofEagle medium
containing 5% horseserum. After1 h the cellswerediluted 10-foldintomethionine-freemediumcontaining5%dialyzed
1765
on November 10, 2019 by guest
http://jvi.asm.org/
1766 JONES ET AL.
horse serum and 5 mCi of [35S]methionine (600 Ci/mmol;
Amersham-Searle).The cellswereincubated forafurther23
h at 37°C and then collected by centrifugation. Virus was
purified by
sedimentationthrougha sucrosecushion and twosuccessive sucrose
gradients
(26). A soluble deoxycholateextract was
prepared,
and RNA polymerase waspurifiedasdescribed
previously (2),
except that the enzyme was stepelutedwith 0.75 M NaClfrom the
phosphocellulose
column,the
aminopentyl
agarose column wasomitted,
and glycerolgradient centrifugation
was added.Constructionof cosmids. Acosmidlibraryof vaccinia virus
DNA was
prepared
inthevectorpHC79
(Boehringer
Mann-heim
Biochemicals, Indianapolis, Ind.).
Vacciniavirus DNAwas
partially
digested with Sau3A and fractionated by su-crosegradient
sedimentation. Each fractionwasassayed by agarosegel
electrophoresis,
andfragments
ofapproximately40kilobase
pairs
wereligated
toBamHI-digested andalka-line
phosphatase-treated
pHC79,asdescribedby Maniatisetal.
(23).
Colonies were transferred to nitrocellulose, and cosmidscontaining
viralDNA wereidentifiedby hybridiza-tionto32P-labeled
vaccinia virusDNA. Initialcharacteriza-tion of
positive
colonies wasperformed
by agarose gelelectrophoresis
ofHindIll- andSall-digested cosmid DNA.Subsequently,
thecosmidDNAwaslabeledin vitro with32pand
hybridized
tostrips
of nitrocellulosecontaining
electro-phoretically
resolved HindIll orSallfragments ofvacciniavirusDNA. Inthismanner, the segmentofthevacciniavirus
genomewithin each cosmid was determined. A
representa-tive set of cosmids that
spanned
thegenome was used forthis
study.
Hybrid
selection and cell-free translation of RNA. HeLa cells were treated withcycloheximide
(100 ,ug/ml) to blockprotein synthesis
andafter10minwereinfected with30 PFUof
purified
vaccinia viruspercell.Cycloheximide
wasmain-tainedin the
medium,
andthecellswereharvestedafter 4 hat
37°C. Cytoplasmic
RNAwaspurified by
CsClcentrifuga-tion
(8).
PlasmidDNA(25,ug)
wastransferredtonitrocellu-lose membranes
(Schleicher
&Schuell,
Inc., Keene, N.H.)(32) and hybridized
toRNAasdescribedbyJones and Moss(16).
After the filters were washed, RNA was eluted and translated in a micrococcal nuclease-treated reticulocytelysate (7, 30). Following
translation, sampleswereincubatedwith antiserum as described below and prepared for
poly-acrylamide
gel electrophoresis.
Immunoprecipitationandpolyacrylamide gel
electrophore-sis. Solubilized virus
proteins
or translationmixtures wereincubated withpreimmunerabbit serum at 4°C for 4 h, and
antigen-antibody
complexes were boundto Staphylococcus aureusprotein
A(10%
solution; Pansorbin;Calbiochem-Behring,
LaJolla,
Calif.; or protein A Sepharose CL-4B;Sigma
ChemicalCo.,
St. Louis, Mo., or Pharmacia FineChemicals,
Piscataway,
N.J.). The supernatant that re-mainedafter
sedimentation in a microcentrifugewastrans-ferred to a fresh tube andincubated overnight at 4°C with
antiserum.
Staphylococcal
proteinAwasaddedto bind theimmunecomplexes.Thebeads werewashed twice with 0.05
M Tris
hydrochloride
(pH7.5)-0.15
MNaCl-0.1%
sodiumdeoxycholate
and twice with 2 Murea-0.4
MLiCl-0.01
M Trishydrochloride
(pH 8.0). The beads were boiled insample
bufferjust beforeelectrophoresis.
Protein samplesalsQ
wereroutinely
analyzedwithout immunoprecipitation.Inthatcase,thetranslationmixture, soluble virus extract, or
purified
RNApolymerasewasdiluted with several volumes ofsample
bufferandboiled for5 min. Samples were applied to 10%polyacrylamide
gels containing sodium dodecyl [image:2.612.318.556.86.217.2]sul-fate,
andelectrophoresis
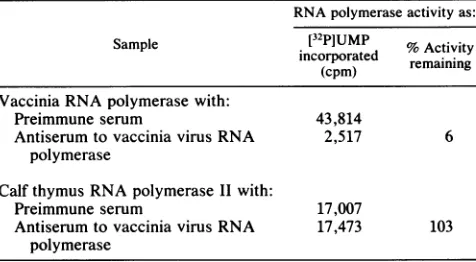
was continued until the hemeTABLE 1. Inhibition ofvacciniavirus RNApolymeraseactivity RNApolymeraseactivityas:
Sample [32P]UMP %Activity
incorporated remaining (cpm)
Vaccinia RNApolymerase with:
Preimmuneserum 43,814
Antiserumtovaccinia virus RNA 2,517 6 polymerase
CalfthymusRNA polymerase I1 with:
Preimmuneserum 17,007
AntiserumtovacciniavirusRNA 17,473 103 polymerase
reached the bottomof thegel. Separation ofthetwo
largest
subunits was achieved by electrophoresis in 5%
polyacryl-amide gels. The gels were then fixed, soaked in En3Hance (New England NuclearCorp., Boston, Mass.), dried under vacuum, and exposed to X-Omat XAR-5 film (Eastman
Kodak Co., Rochester, N.Y.).
RNA polymerase assay. RNA polymerase activity was measured by using a single-stranded DNA template as
described by Baroudy and Moss(2), except that[ot-32P]UTP
wasused. Samples were then monitored by Cerenkov count-ing. Calf thymus RNA polymerase was assayed under the sameconditions.
RESULTS
Inhibition of vaccinia virus RNA polymerase activity with antiserum. DNA-dependent RNA polymerase that was pu-rified from vacciniavirions waspreviously shown to havea
molecular weight of approximately 500,000and to be
dissoci-able into polypeptides with molecular weights of
approxi-mately 140,000, 137,000, 37,000, 35,000, 31,000, 22,000,and 17,000(2, 38). Antiserum, which was preparedfrom arabbit
thatreceivedmultipleinoculationsof purifiedvaccinia virus
RNA polymerase, inhibited the activity of the RNA
poly-merase by more than90%, whereas preimmune serumhad
little or no effect (Table 1). The antiserum was notdirected
toward conserved epitopes in viral and cellular RNA
poly-merases because calf thymus RNA polymerase II activity was not inhibited (Table 1). This antiserum also inhibited transcription of early vaccinia virus genes that were added to extracts ofvaccinia virus-infected cells (31).
Immunoprecipitation of vaccinia virus RNA polymerase subunits. The specificity of the antiserum was tested by immunoprecipitation of vaccinia virus proteins. [35S]methi-onine-labeled polypeptides, which were present in a soluble extractof virus particles or purified RNA polymerase, were resolved by sodium dodecylsulfate-polyacrylamide gel elec-trophoresis (Fig. 1, lanes 1 and 4). Under these electropho-retic conditions, the two large polymerase subunits were not resolved, and they migrated with a molecular weight of approximately 135,000. Prominent 36,000-Mr (36K), 34K, 23K, 20K, and 17K polypeptides and faint 31K and 21K polypeptides in the RNA polymerase preparation were sim-ilar in size to those described previously (2). (The 17K
polypeptidemigrated off the bottom of the gel and is not seen
inFig. 1.) The prominent 70K polypeptide and minor 55K to 200Kpolypeptides in the RNA polymerase preparation (Fig. 1,lane 4) were reduced in amount when the purification was
performedasoriginally described (2), which was the
proce-dure used to obtain RNA polymerase for immunization. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
VACCINIA VIRUS RNA POLYMERASE SUBUNITS 1767
When the partially purified polymerase was incubated withimmune serum, at least eight polypeptides were specif-ically precipitated (Fig. 1, lanes 5 and 6). Because the polypeptides were physically associated in a complex, this type of analysis did not reveal whether the antiserum was directed against eachindividual polypeptide or against only a few. The absence from the immunoprecipitate of the prominent 70K polypeptide as well as the minor ones greater than50,000daltons, however, indicates that they are neither recognized by the antiserum nor tightly associated with the RNA polymerase in the triple detergent buffer used for antibody binding. Some additional polypeptides were
pre-cipitated from the crude virus extract (Fig. 1, lane 3);
however, this was largely nonspecific because most were also seen after incubation with preimmune serum (Fig. 1, lane 2).
Immunoprecipitation of vaccinia virus RNA polymerase subunits synthesized in vitro. The following experiments weredesigned to determine whether mRNA for RNA poly-merase subunits was present in the cytoplasm of vaccinia
virus-infectedcells and could be translated in vitro. Results
of preliminary control experiments indicated that specific
immunoprecipitablepolypeptides were not made when RNA
from uninfected cells was added to micrococcal
nuclease-treated reticulocyte lysates. An autoradiograph of the total
[35S]methionine-labeled
translation products produced in such a lysate programmed with RNA from vaccinia virus-infected cells is shown inFig. 1, lane 7. As usual, large-sizedtranslation products are underrepresented. The antiserum
specifically precipitated 36K, 34K, 31K, 23K, and 20K
polypeptides (Fig. 1, lane8). Although notclearly visible in
Fig. 1, immunoprecipitable 135K, 21K, and 17K
polypep-tides also were made and will be shown in subsequent
figures.
The RNA that was used to program thereticulocyte lysate
M 1 2 3 4 56 7 8 9
200.
__135
92.5
-68-4__
43 -e
_
-
~~~~36
2573
2D5.7_.s
_ w23
18.4.
FIG. 1. Immunoprecipitation ofRNApolymerasesubunits syn-thesized in vivo and in vitro. Autoradiographs of radioactively
labeled proteins heated at 100°C with sodium dodecyl sulfate and mercaptoethanol andsubjected topolyacrylamide gel
electro-phoresisareshown. LaneM,"C-labeledstandardproteinmarkers; lanes 1 to3,proteins extractedwithdeoxycholatefrom
[35S]methi-onine-labeledpurified virions;lanes 4to6, [35S]methionine-labeled purifiedvaccinia virusRNApolymerase; lanes 7to9,
[35S]methio-nine-labeledreticulocyte cell-freetranslationproductsoftotal cyto-plasmic RNA from HeLa cells treated with cycloheximide and infectedwithvaccinia virus for4h. Lanes1, 4, and 7containtotal protein; lanes 3, 5,and 8containproteinsthatwere immunoprecipit-ated with antiserumtovacciniaRNApolymerase;lanes 2, 6,and 9 contain proteins that were immunoprecipitated with preimmune
serum.Molecularweights (103)areindicatedtotheleftand
right
ofthegels.
19 3 21 10 A 6 E 200
21 10
925 -.
68
-4:3
25.1
18.4-'_ -31
-34
_ -23
-21 --20
cos 21 cos 3 cos 19
cos 6
coa 1Q
Hindill C NMKF E 01&GLJL H
I I1i I
A B
. I
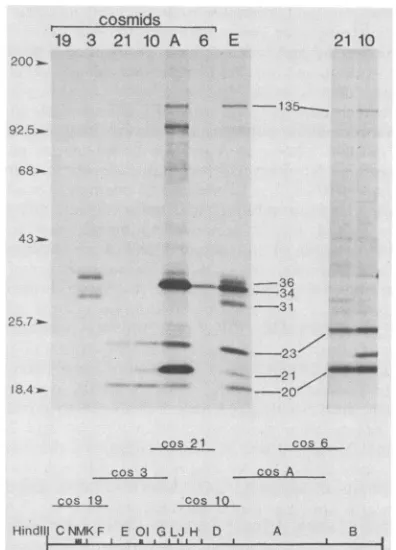
FIG. 2. Mapping ofRNApolymerase subunitgenes by transla-tion ofmRNA selected by hybridization to a cosmid library. A schematic representation of the vaccinia virus DNA contained within6cosmid (cos) clones is shownatthe bottom. The letters, which refertoHindIlI fragments, are shown forreference. Cyto-plasmic RNA, which was obtained from HeLa cells that were treated withcycloheximideandinfected withvaccinia virus for 4 h, washybridizedtocosmids that were immobilizedonnitrocelluose filters. Thespecifically boundRNA waseluted andtranslated ina micrococcal nuclease-treatedreticulocyte cell-free system contain-ing[5S]methionine. Labeledproteinswereincubated withvaccinia virus RNA polymerase antiserum, boundto S. aureus protein A, and dissociated with sodium dodecyl sulfate forpolyacrylamidegel electrophoresis. Autoradiographsareshown. Inlanes19, 3, 21,10, A, and 6, the reticulocyte lysates were programmed with mRNA that wasselectedbyhybridizationtotheindicatedcosmid.The two lanes at the extremeright are longerautoradiographic exposures. Lane E containsimmunoprecipitated translation productsoftotal cytoplasmic early RNA. Molecularweights (103) of marker proteins areindicated.
was obtained from cells that were infected with vaccinia
virus in the presence of cycloheximide. With this protein
synthesisinhibitor, earlyRNAis made inlargeamountsand
DNA replication and the switch to late
transcription
isprevented. When similarexperimentswerecarriedoutwith RNAthatwasobtainedat6hafterinfection in the absence
of inhibitors, much lower amounts ofimmunoprecipitable
polypeptides were obtained (data not shown). All
subse-quentexperimentsthereforewerecarriedoutwith RNA that
was made in the presence ofcycloheximide.
Hybridization ofRNA tovaccinia virus DNA
fragments
ina cosmidlibrary.Todetermine whether the RNApolymerasesubunitsarevirus encoded andtolocate their mappositions, wetranslated mRNA that
hybridized
tovaccinia virus DNA segments. A set of six cosmidscontaining
overlapping40-kilobase-pair vaccinia virus genome fragments (Fig. 2)
were immobilized on nitrocellulose membrane filters and used to
hybrid
select RNA that waspurified
from the VOL.61, 1987
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.340.538.74.349.2]1768 JONES ET AL.
cytoplasm of cycloheximide-treated, virus-infected cells.
Following translation andimmunoprecipitation,the
[35S]me-thionine-labeled polypeptideswere dissociated with sodium
dodecyl sulfate andanalyzed by polyacrylamide gel
electro-phoresis.
A control lane (Fig. 2, lane E) shows that all
size-immunoprecipitable polymerasesubunits, includingatleast
oneof135,000daltons,weremade with the unselected RNA
preparations. No immunoprecipitable products were
ob-tained when RNA that hybridized to cosmid 19 was used,
suggesting that no subunits map near the left end of the
genome(Fig. 2).Twoimmunoprecipitable polypeptides with
molecular weights of 36,000 and31,000 were obtainedwith
RNAthathybridized to cosmid3. Because neither of these
polypeptides was made with RNA that
hybridized
tocosmids19or 21,theymustmap within the centralregionof
cosmid 3. The 135K, 23K, and 20K polypeptides were
mapped within cosmid 21.' The longer
autoradiographic
exposure on the right side ofFig. 2 moreclearly shows the
135K polypeptide. The 135K, 23K, and 20K
polypeptides
also were made with RNA that hybridized to cosmid 10,
which has a large region of overlap with cosmid 21. An
additional21Kband was resolved, suggestingthat the gene
forthissubunitmay belocatedneartherightendof cosmid
10. Subunits of about 135,000, 34,000, 23,000, and 20,000
daltons alsoappearedto be encoded within cosmid A. The
formation ofa23Kpolypeptide withmRNAsthat
hybridized
tocosmids 21,10,andAsuggest that there is more than one
polymerase subunit of this size or that there are
repetitive
DNAsequences. Additionalpolypeptidessuchasthe
prom-inent one of 95,000 daltons and minor ones below it
might
result from premature termination or other artifacts of in
vitro translation (see below). The synthesis ofa 34K
poly-peptidewith RNAthathybridizedtocosmid6suggests that
the gene is located within the
overlapping right
half ofcosmidA.
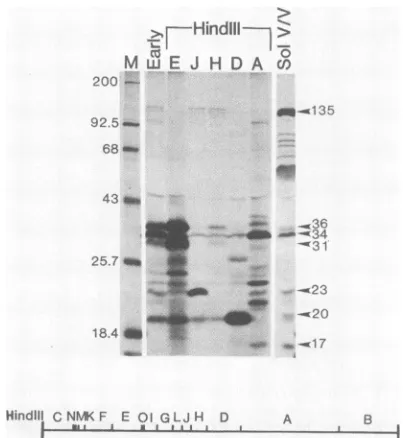
Furthermapping.CosmidAcontainsthe
largest
ofthe 15fragmentsthat were obtainedby digestion of vaccinia virus
DNA with HindIII. Except for the two terminal HindIll
fragments,
the rest havebeen cloned in pBR322previously (3). The 13 availableHindIIIfragmentswereusedforhybrid
selection, and the mRNAs were translated as described
above. Only fragments E, J, H, D, and A hybrid selected
mRNA foroneor more polypeptides that wererecognized
by antiserumto RNApolymerase (Fig. 3). HindIII-E, which
is entirely contained within cosmid 3, encoded prominent
polypeptides of 36,000 and 31,000 daltons. Because of the
overexposure of the autoradiograph, additional smaller
polypeptides, some of which may represent premature
ter-mination products of the larger ones, were detected (see
below). Twopolypeptides that comigratedwith RNA
poly-merasesubunitsof135,000and23,000daltonswereencoded
withinthe
HindlIl
J fragment. Because the largerpolypep-tidealso was synthesized from RNA that hybridized to the
adjacent HindIII H fragment, this gene evidently spans the
HindIII-J and -Hjunction. Previous studies showingthat a
long transcript crosses that HindIII site (1, 21, 40) and
nucleotide sequence data (4) support this interpretation. mRNA for the 20K polypeptide was selected most
effi-ciently by HindIII-D. We are not sure whether minor 20K
bands synthesized with mRNA that hybridized to DNA
fragments other than
HindIII-D
represent additional RNApolymerase subunits or are simply artifacts of in vitro
translation. The failure ofHindIII-D to hybrid select mRNA
for alarge subunit is significantbecause it implies that the
large subunits that map to the left and right are encoded by
separate genes. The data
(Fig.
3)
obtainedby hybrid
selec-tion with theHindlIl Afragment
aresimilartothose shown inFig.
2. Because of the shorterelectrophoresis
time,
however,
the 17Kpolypeptide
was retained on thegel
and could bemapped
withinHindIII-A.Resolution ofthetwo
high-molecular-weight
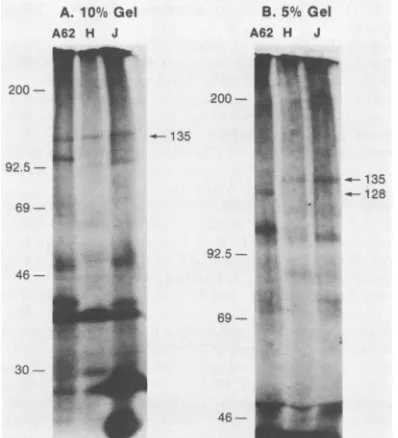
RNA polymer-asesubunits. To better characterize the twolarge
subunits,
efforts were made to
improve
theirelectrophoretic
separa-tion. mRNA was
hybrid
selected toHindIII-J,
HindIII-H,andaBamHIsubclone of cosmid A
(which
has been shownto encode one of the
large subunits; unpublished
data).
Translation and
immunoprecipitation
wereperformed
asdescribed
above,
butequal portions
of theimmunoprecipi-tates were
electrophoresed
on 10 and 5%polyacrylamide
gels.
Under our standardelectrophoresis
conditions(10%
gel),
the cosmid A subclone A62 and theHindIII
J and Hfragments
hybrid
selected mRNAencoding
135Kproteins
thatcomigrated (Fig. 4.)
Inthe 5%gel,
however,
thelarge
polypeptides
fromHindIII-Aand from the J-Hjunction
wereclearly distinguishable (Fig. 4).
The molecularweight
of theformer was estimated to be
128,000
and of the latter was135,000.
Additional translationproducts
from mRNA that washybrid
selectedto thesubclone ofHindIII-A and from HindIII-Jand-Hmayrepresent premature
terminations.Asummary ofthedatais
presented
inFig.
5. Oneof thehigh-molecular-weight polypeptides
(135K)
mapped
at theHindIII-J and -H
junction,
and another(128K)
mapped
withinthe
large
HindlIl Afragment.
An RNApolymerase
subunit of
36,000
daltons and anadditionalpolypeptide
of31,000 daltons
mapped
withinHindIII-E.
The 34K subunitmapped
withinHindIII-A,
although
a faint 34K band alsoHindAll >
Mui E J H D A ch 200
-92.5 _
68 _
43 _Ub
_-135
- s .36
_M o-34 31
V.0 -W20 ...- ; ....1 7
18.4 i
Hindlil CNMKF E 01 GLJH D
I a.l A a I 11 A B
-I
FIG. 3. FurthermappingofRNApolymerase subunits.Hybrid
selectionandcell-free translationwere carriedout asdescribedin thelegendtoFig.2,except thatplasmids containing vaccinia virus DNAHindlll E, J, H,and Dfragmentswereimmobilizedonfilters. TheHindIIIAfragmentwascloned inacosmid. Lane Mcontains
'4C-labeled markerproteins. Other lanes contain immunoprecipi-tated translationproductsoftotalearly cytoplasmicRNA(Early)or RNA that was selected by hybridizationto the indicated HindIII DNAfragment.The lanemarked SolV/V containssolubleproteins that were extracted from [35S]methionine-labeled vaccinia virions withdeoxycholate and immunoprecipitated.Molecularweights(103)
areindicatedtotheleftandrightofthegels.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.336.538.395.614.2]was producedby using RNA that hybridized to HindIII-E. Two 23K polypeptides, one mapping to HindIII-J and the
other to HindIII-A, were found. In addition, a major 20K
polypeptide mapped to HindIII-D, and 21K and 17K
polypeptides mapped to HindIII-A.
DISCUSSION
The subunit structure of the vaccinia virus-associated
DNA-dependent RNA polymerase conforms to that of
eu-caryoticRNApolymerases ingeneral. Results of the
exper-iments reported here indicate, however, that many or
per-haps all ofthe subunits are virus-encoded. Although the
polyvalent antiserumthat we prepared to the viral enzyme
didnotinhibit the activity of calf thymus RNA polymerase
II,this doesnotmean that the viral and cellularpolymerases
areunrelated.One might expect that the mostimmunogenic
regions of the viral polymerase are those that are most
distinct fromcellularpolymerase. Morrison and Moyer (25)
have prepared a monoclonal antibody that reacts with the
large subunitsof cellular RNA polymerase II and rabbitpox
RNA polymerase. In addition, analysis of the predicted
amino acid sequence of the largevacciniavirus RNA
poly-merase subunit revealed extensive homologywith the large
RNApolymerase subunits of E. coli, S. cerevisiae, and D.
melanogaster(4). Other vacciniavirus-encodedRNA
poly-merasesubunitsare also likely toshare some homology with
RNApolymerases from procaryotesandeucaryotesbecause
A.10% Gel A62 H J
200-92.5
69-135
B.5% Gel
A62 H J
200-~4
*4'.7 4 - 135
- 128
92.5- A
69-
46--FIG. 4. Separation of thetwolarge molecularweight subunitsof vacciniaRNApolymerase. Hybrid selectiontoplasmids containing theHindIll J andHfragmentsortoplasmid A62,aBamHI subclone ofHindIll A(A62), was performedas describedin the text. The translationproductswereimmunoprecipitatedwith antiserumtothe
viral RNApolymerase, and equal amountsof the
immunoprecipi-tated material were loaded onto 5 and 10%polyacrylamide gels. Autoradiographs of theimmunoprecipitatesthatelectrophoresedin
a10 and5%gelareshowninpanelsA andB,respectively.Lane M
contains '4C-labeledmarkerproteins.Thelanesdesignated A62, H,
orJshowimmunoprecipitatesof translationproductsof mRNAthat
was selected by hybridization to plasmids containing a BamHI
subclone of HindIII-A or the HindlIl H or HindIII J fragments,
respectively. Molecular weights of marker proteins and
im-munoprecipitableRNApolymerase subunitsare shownontheleft
andrightof thegels,respectively.
Hirdll C ii F I E NIIGILIJIHI D I
36K 135K 20K
31K 23K
Hindil A
128K 34K 23K 21K 17K
FIG. 5. Locations of RNA polymerase subunit genes. Letters indicate theHindIIIfragments.
immunological relatedness has been demonstrated for both
large and smallsubunitsfrom widelydifferentorganisms (13, 14, 19, 20, 37, 39).
Because our antiserum was made against extensively
purified RNApolymerase, it was directed against
polypep-tides thatweretightly associated withtheenzyme. Whether
each of these polypeptides was a functional subunit,
how-ever, cannot be determined at this time. Conversely, the
vigorous purification procedure may have led to the
disso-ciation of importantaccessory polypeptides. The
participa-tion of addiparticipa-tional polypeptides in transcription is likely
becausehighly purifiedenzyme is unable to initiatecorrect
transcription on adouble-stranded viral DNA template (2,
38). These problems of interpretation are not specific for
vacciniavirus,however, but are general ones foreucaryotic
RNApolymerases.
Our basic approach to mapping the RNA polymerase
subunits was to immunoprecipitate reticulocyte cell-free
translationproducts ofmRNAthatwere selected by
hybrid-ization to cloned vaccinia virus genome fragments. The
majortechnical problem that was encountered in this
pro-ject related to the difficulty in translating mRNAs for the
high-molecular-weight
subunits. In some experiments, theautoradiographic bands were too faint to permit
positive
identification. This difficulty may have resulted from both
degradation ofmRNA and premature termination. Indeed,
discreteimmunoprecipitable polypeptide bands below those
of full size were
frequently
noted and caused somediffi-culties in
interpretation.
Similar problems arose when thevaccinia virus DNA polymerase gene was mapped
previ-ously (16).
It is not surprising that all of the RNA polymerase
subunits mapped withinthecentralhighly conserved region
ofthe poxvirus genome and not within the more variable
ends. Nevertheless, the wide dispersion of the subunits
within the central region was unanticipated. Although
fur-ther mapping is required formost ofthe RNApolymerase
subunits,
detailedtranscriptional analysis
and DNAse-quence data have beenreported for severalregions inwhich
polymerase subunits appear to be encoded. Within the
HindIII J fragment, five early mRNAs (590, 1,070, 1,790,
2,380, and 3,840 nucleotides long) were mapped by blot
hybridization
andSi
nucleaseanalysis
(1, 4, 40). The 590-and2,380-nucleotide
species
were identified asmajor
andreadthrough forms ofthethymidine kinasemessage,
respec-tively. The
1,790-nucleotide
RNA is coterminal with thelonger
thymidine
kinase message and encodes a 41Kearly
protein
of unknown function. The1,070-nucleotide RNA,
which also iscoterminal with the
1,790-nucleotide
mRNA,encodes a 21K
polypeptide
thatis almostcertainly
the23KRNA polymerase subunit described here (4, 21,
40).
TheDNA sequence
encoding
along
transcript
of greater than3,800 nucleotides is bisected
by
theHindIII siteseparating
fragments J and H. The translational
product
ofthislong
mRNA was
reported
previously
to be110,000
daltons(3);
however, our
reappraisal
of theoriginal
autoradiograph
indicated that this value was anunderestimate and that the
46-30
--I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.320.559.76.114.2] [image:5.612.85.284.361.580.2]1770 JONES ET AL.
size of the polypeptide predicted by theopen readingframe was147,000 daltons (4). Thus, thetruesize of the 135KRNA
polymerase subunit is probably 147,000 daltons. The large subunit of rabbitpox RNA polymerase complex has also been mappedtothe same region(24, 25).
Although ithasnot yetbeentranscriptionally mapped, the
entire sequence of the HindIll D fragment has been
pub-lished (29). Inspection of this sequence revealed 13 open
reading frames encodingproteins ofgreaterthan 145amino
acids. Based on this analysis, there arefour open reading
frames intheHindIII Dfragmentthatencode proteins with molecularweightsintherangeof the 20K RNA polymerase
subunit. Because temperature-sensitive mutants have been
localized to someof theseopenreading frames (29),
identi-fication ofthe 20K subunitgene is ofimmediate interest.
Neither detailed transcriptional analysis nor extensive
DNA sequencinghasbeenreported foreither the
HindIll
A orE fragments. However,Morrisonetal. (24)mappeda34KRNA polymerase subunit within HindIII-A, which is
con-sistent withourresults.
Attemptsatdetermining molar ratios for thevaccinia virus
RNA polymerase subunits previously ledto the suggestion
of multiple copies of 34K and 23K polypeptides (2). Data
from the results of this studyindicatethat for the 23K, and
possiblythe34K,polypeptidemorethanone geneexiststhat
presumably codes for different polypeptides of similar size.
On the other hand, results of preliminary peptide analyses suggestthat the 36Kand 31Kpolypeptides that map tothe
HindIII Efragmentmay bederivedfromone gene
(unpub-lisheddata). Thelowamountof the 31KpolypeptideinRNA
polymerase raises further doubts about its significance. We
estimate, therefore, that there are at least nine viral genes
that encodepolypeptides thatarephysicallyassociated with
thevacciniavirus RNA polymerase.Associationof the large
subunit ofcellular RNA polymerase with rabbitpox virus
RNA polymerase also has beensuggested (25).
The ability to synthesize all of the polymerase subunits
from cells that are infected with vaccinia virus in the
presence of cycloheximide indicates that these genes are
expressed early in infection. Otherdata, however, suggest
that some subunits may be expressed at late times in the
virus life cycle as well. Mahr and Roberts (21) observed
mRNA speciesthat hybridizedtothe
HindIlI
Jfragment andwhichcould encodethe 23Kand 135Ksubunits in
prepara-tions of lateRNA(8 hafter infection).Results ofS1 nuclease
analysisalso suggestthepossibility ofalate RNA start site
upstream ofthe early one encoding the large subunit (4).
Morrison and co-workers (24, 25) found by immunoblot
analysisof rabbitpox-infectedcells thataccumulation ofthe
large subunit is detected only after viral DNA replication occurs.
Mapping ofthe RNApolymerase subunitsopenstheway
foravarietyofnewexperiments. Nucleotide sequencing of
twoofthe subunitgenesiscompleted, and forone of these
geneshomologywithprocaryoticand eucaryotic RNA poly-merasesubunits has been demonstrated (4). Efforts to fine mapandsequencethe otherRNA polymerasesubunitgenes
are inprogress. This structural information will aid in the preparationofspecific antiserum,either by usingexpression vectorstogeneratelargeamountsof individualpolypeptides or by oligopeptide synthesis. The production of large amountsof individual subunitsby recombinant DNA
meth-ods also should be useful in defining their catalytic roles. Additionally, the map and sequence information will facili-tate thegeneration and analysis ofRNA polymerase
muta-tions.
ACKNOWLEDGMENTS
We thank Bahige Baroudy for purified vaccinia virus RNA polymerase, Rose Mage for preparation of antiserum, Theodore Theodore for the lambda packaging mixture, AlanGershowitz for preparing the cosmid library, Helena Mishoe for a sample of calf thymus RNA polymeraseII, and Norman Cooper for purifying vaccinia virus.
LITERATURE CITED
1. Bajszar, G., R. Wittek, J. P. Weir, and B. Moss. 1983. Vaccinia virus thymidine kinase and neighboring genes: mRNAs and polypeptides of wild-type virus andputative nonsensemutants. J. Virol. 45:62-72.
2. Baroudy, B. M., and B. Moss. 1980. Purification and character-ization of a DNA-dependent RNA polymerase from vaccinia virions. J. Biol. Chem. 255:4372-4380.
3. Belie Isle, H., S. Venketesan, and B. Moss. 1981. Cell-free translation of early and late mRNAs selected by hybridization to cloned DNA fragments derived from the left 14 million to 72 million daltons of the vaccinia virus genome. Virology
112:306-317.
4. Broyles, S. S., and B. Moss. 1986. Homology between RNA polymerases of poxviruses, prokaryotes, and eukaryotes: nu-cleotide sequence and transcriptional analysis of vaccinia virus genes encoding 147-kDa and 22-kDa subunits. Proc.
Natl.
Acad. Sci. USA 83:3141-3145.5. Burgess, R. R., A. A. Travers, J. J. Dunn, and E. K. F. Bautz. 1969. Factor stimulating transcription by RNA polymerase. Nature (London) 221:43-46.
6. Chambon, P. 1975. Eucaryotic nuclear RNA polymerases. Annu. Rev. Biochem. 44:613-638.
7. Cooper, J. A., and B. Moss. 1978. Transcription of vaccinia virus mRNA coupled to translation in vitro. Virology 88: 149-165.
8. Cooper, J. A., and B. Moss. 1979. In vitro translation of immediate early, early and and late classes of RNA from vaccinia virus-infected cells. Virology 96:368-380.
9. Dezelee, S., and A. Sentenac. 1973. Role of DNA-RNA hybrids in eukaryotes: purification and properties of yeast RNA poly-merase B. Eur. J. Biochem. 34:41-52.
10. Fogelsong, D. P. 1985. In vitro transcription of a cloned vaccinia virus gene by a soluble extract prepared from vaccinia virus-infected HeLa cells. J. Virol. 53:822-826.
11. Golini, F., and J. R. Kates. 1985. A soluble transcription system derived from purified vaccinia virions. J. Virol. 53:205-213. 12. Greenleaf, A. L., and E. K. F. Bautz. 1975. RNA polymerase B
from Drosophila melanogaster larvae: purification and partial characterization. Eur. J. Biochem. 60:169-179.
13. Huet, J., A. Sentenac, and P. Fromageot. 1982. Spot-immuno-detection of conserved determinants in eukaryotic RNA poly-merases: study with antibodies to yeast RNA polymerases subunits. J. Biol. Chem. 257:2613-2618.
14. Ingles, C. J. 1973. Antigenic homology of eukaryotic RNA polymerases. Biochem. Biophys. Res. Commun. 55:364-371.
15. Jendrisak, J., and T. J. Guilfoyle. 1978. Eucaryotic RNA polymerases: comparative subunit structures, immunological properties, and alpha-amanitin sensitivity of the class II en-zymes from higher plants. Biochemistry 17:1322-1327. 16. Jones, E. V., and B. Moss. 1984. Mapping of the vaccinia virus
DNA polymerase gene by rescue and cell-free translation of selected mRNA. J. Virol. 49:72-77.
17. Kates, J. R., and B. R. McAuslan. 1967. Poxvirus DNA-dependent RNA polymerase. Proc. Natl. Acad. Sci. USA 58:134-141.
18. Kedinger, C., and P. Chambon. 1972. Animal DNA-dependent RNA polymerases: purification of calf thymus BI and
BII
enzymes. Eur. J. Biochem. 28:283-290.
19. Kedinger, C., F. Gissinger, and P. Chambon. 1974. Animal DNA-dependent RNA polymerases: molecular structures and immunological properties of calf thymus enzymeAland of calf thymus and rat liver enzymes B. Eur. J. Biochem. 44:421-436. 20. Kramer, A., and E. K. F. Bautz. 1981. Immunological related-ness of subunits of RNA polymerase II from insects and J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
mammals. Eur. J. Biochem. 117:449-455.
21. Mahr, A., and B. E. Roberts. 1984. Organization of six early transcripts synthesized from vaccinia virus EcoRI DNA frag-ments.J. Virol.49:497-509.
22. Mandel, J. L., and P. Chambon. 1971. Purification of RNA polymeraseBactivity fromratliver. FEBS Lett. 15:175-180. 23. Maniatis, T., E. F. Fritsch, and J.Sambrook. 1982. Molecular
cloning: alaboratory manual. Cold Spring Harbor Laboratory,
ColdSpringHarbor, N.Y.
24. Morrison, D. K., J. K. Carter,and R. W. Moyer.1985.Isolation andcharacterization ofmonoclonal antibodies directedagainst
twosubunits of rabbitpoxvirus-associated, DNA-directedRNA polymerase.J. Virol.55:670-680.
25. Morrison, D. K.,andR.W. Moyer.1986.Detection ofasubunit
of cellular pol II within highly purified preparations ofRNA polymerase isolated from rabbit poxvirus virions. Cell 44:587-596.
26. Moss, B. 1968. Inhibition ofHeLacellprotein synthesis bythe vacciniavirion. J. Virol. 2:1028-1037.
27. Munyon, W., E. Paoletti, and J. T. Grace, Jr. 1967. RNA polymerase activityinpurified infectious vaccinia virus. Proc. Natl. Acad. Sci. USA58:2280-2287.
28. Nevins, J. R., and W. K.Joklik.1977. Isolationandproperties of the vaccinia virus DNA-dependent RNA polymerase. J. Biol. Chem.252:6930-6938.
29. Niles, E. G., R. C. Condit, P. Caro, K. Davidson, L. Matusick, andJ. Seto. 1986.Nucleotidesequenceandgeneticmapofthe
16-kbvaccinia virusHindlllDfragment. Virology 153:96-112. 30. Pelham, H. R. B., and R. J. Jackson. 1976. An efficient mRNA-dependenttranslationsystemfrom reticulocytelysates. Eur.J. Biochem. 67:247-256.
31. Puckett, C., and B. Moss. 1983. Selective transcription of vaccinia virusgenes in template dependent solubleextractsof
infected cells. Cell 35:441-448.
32. Ricciardi, R. P., J. S. Miller, and B. E. Roberts. 1979. Purifica-tion and mappingof specific mRNAs by hybridization-selection and cell-free translation. Proc. Natl. Acad. Sci. USA 76:4927-4931.
33. Roeder, R. G. 1974. Multiple forms of deoxyribonucleotide acid-dependentribonucleic acid polymeraseinXenopus laevis: isolation and partial characterization. J. Biol. Chem. 249:241-248.
34. Roeder,R.G.1976. RNApolymerase, p.285-329.ColdSpring HarborLaboratory, ColdSpring Harbor,N.Y.
35. Rohrmann, G., and B. Moss. 1985. Transcription of vaccinia virus early genes by a template-dependent soluble extract of purified virions.J. Virol.56:349-355.
36. Schwartz, L. B., and R. G. Roeder. 1975. Purification and subunit structure of deoxyribonucleic acid-dependent ribonu-cleic acidpolymerase II from themouseplasmacytoma, MOPC 315.J. Biol. Chem. 250:3221-3228.
37. Sklar, V. E.F.,L. B.Schwartz,and R.G.Roeder.1975. Distinct molecular structures of nuclear class I, II, and III DNA-dependent RNA polymerases. Proc. Natl. Acad. Sci. USA 72:348-352.
38. Spencer, E.,S.Schuman,andJ.Hurwitz.1980.Purification and properties of vaccinia virus DNA-dependentRNApolymerase. J. Biol.Chem. 255:5388-5395.
39. Weeks, J. R., D. E. Coulter, and A. L. Greenleaf. 1982. Immunological studies ofRNApolymerase IIusing antibodies to subunits of Drosophila and wheat germ enzyme. J. Biol. Chem. 257:5884-5891.
40. Weir, J. P., G. Bajszar, and B. Moss. 1982. Mapping of the vaccinia virusthymidine kinase gene by markerrescueandby cell-free translation of selectedmRNA. Proc. Natl. Acad. Sci. USA 79:1210-1214.