JOURNALOFVIROLOGY,Sept. 1992, p.5542-5552 Vol. 66, No.9 0022-538X/92/095542-11$02.00/0
Copyright ©1992, American SocietyforMicrobiology
Expression of the Influenza A Virus M2 Protein Is Restricted
to
Apical Surfaces of Polarized
Epithelial Cells
PATSY G.
HUGHEY.'
RICHARD W.COMPANS,1*
SUZANNE L. ZEBEDEE,2t ANDROBERT A. LAMB2'3Departmentof
Microbiology, University of
Alabama atBirmingham, Birmingham,
Alabama35294,1
andDepartmentof
Biochemistry,
Molecular and CellBiology,2
and HowardHughes
MedicalInstituteResearchLaboratories,3 Northwestern University, Evanston, Illinois 60208-3500
Received 22April1992/Accepted 10 June 1992
The M2protein of influenza A virus isasmall,
nonglycosylated
transmembraneprotein that isexpressedon surfaces of virus-infected cells. A monoclonal antibody specificfor the M2proteinwasused toinvestigate
its expression in polarized epithelial cells infected with influenza virus or a recombinant vaccinia virus that expressesM2. Theexpression of M2onthe surfaces of influenza virus-infected cellswasfound to be restricted to the apical surface, closely paralleling that of the influenza virushemagglutinin (HA).
Membranedomain-specific immunoprecipitation indicated that the M2 protein was inserted
directly
into the apical membrane with transport kinetics similar to those of HA. Inpolarized
cells infected with a recombinant vaccinia virus that expressesM2,wefound that 86to93% ofsurface M2wasrestrictedtotheapical
domain compared with 88 to 90% ofHAin a similar assay. These results indicate that the M2protein undergoes directional transport in the absence of other influenza virus proteins and that M2 contains the structural featuresrequiredforapical transport inpolarized epithelialcells. The ultrastructural localization of the M2 protein in influenza virus-infected MDCK cells was investigated by immunoelectron microscopy using M2antibody and a gold
conjugate.
In cells in which extensive virusbudding
wasoccurring,
theapical
cell membrane was labeled with gold particles evenly distributed between microvilli and the surroundingmembrane. In addition, a significant fraction of the M2 label was
apparently
associated with virions. A monoclonalantibodyspecificfor HA demonstrated asimilarlabelingpattern.These results indicate that M2 islocalized in closeproximitytobudding and assembled virions.The M2 protein of influenza A virus (for a review, see reference22)consists of 97 amino acids(24)and isoneof the smallest known transmembrane proteins.M2
belongs
tothe type Isignal-anchorprotein class and has been used previ-ously to investigate the structural featuresresponsible
for this uncommon topology (37, 38). The external domain ofM2, which consists of approximately 24 N-terminal amino acids (24), appears to determine the type I signal-anchor topologyof M2and ofcertain chimerictype IIproteinsthat contain these sequences (37, 38). M2 is cotranslationally
inserted into the roughendoplasmic reticulum membrane via a 19-residue hydrophobicsignal-anchor domain(17), and 54
C-terminal amino acids form the cytoplasmic domain (24).
Recent evidence indicates that the M2 protein forms
disul-fide-linkeddimers, tetramers, andpossiblyhigher-order oli-gomers and is modified by the addition of palmitate to a
cytoplasmic cysteine residue in most influenza A virus strains (15, 48, 54), but a potential N-linked glycosylation site in the external domain is unused (24, 59). M2 is trans-ported through the Golgi complex to the surfaces of cells infected with influenza virus or recombinant expression vectors (24, 59), similar to the pathways traversed by the viralglycoproteins (6). Large numbers of M2 molecules are present on infected-cell surfaces, with a ratio of approxi-matelytwo M2molecules per hemagglutinin (HA) trimer in CV-1cells(24), but only a few (-23 to 60) M2 molecules are
estimated to be incorporated into each virion (57). The
*Correspondingauthor.
tPresent address: R. W. Johnson Pharmaceutical Research In-stitute, San Diego,Calif.
contrastbetween the numbers ofmoleculesexpressedonthe cell surface and invirions suggests that M2 may be partially excluded from buddingvirusparticles.
Polarizedepithelialcellsarecharacterizedby the presence of apical and basolateral plasma membrane domains that havedistinctiveprotein andlipidcompositions (6).Polarity isdevelopedby thedirectional transportof cellular proteins and lipids (1, 6, 25, 26, 33, 53) and maintained by tight
junctions that physically separate apical and basolateral domains(52). Polarized epithelialcellsfromdifferent tissues, such as hepatocytes (1), kidney cells (26, 28, 31, 40), and intestinal cells (25, 33), utilize distinct pathways of protein
sortingand may also differ in the relative amount of signal-mediated and bulk flow(default) transport to the cell surface. Although cellular sorting mechanisms are not yet
under-stood, directional transportapparently involves the recogni-tion ofintrinsicstructural features of targeted molecules by cellular sorting receptors (6). Newly synthesized proteins are sorted duringorimmediately after exit from the
trans-Golginetwork(11),when apical and basolateral proteins are
incorporated into distinct populations of transport vesicles that contain sorting and/or targeting receptors (55). Many exogenous proteins, such as viral glycoproteins, are also
restrictedtospecific plasma membrane domains when intro-duced into polarized cells. The influenza virus glycoproteins,
hemagglutinin(HA) and neuraminidase, are restricted to the apical domain of polarized cells infected with influenza virus or virus expression vectors (20, 31, 46). In contrast, the
glycoproteins of vesicular stomatitis virus (40, 46) and ret-roviruses(44, 46) are transported to basolateral
membranes,
where assembly of these viruses occurs (43, 44). Directional transport ofthese glycoproteins in the absence of other viral
5542
on November 9, 2019 by guest
http://jvi.asm.org/
proteins indicates that the proteins contain intrinsic struc-tural features recognized by the cellular sortingmachinery
(20, 46). It is not known whether positive signals are required for both apical and basolateral transport. Proteins that lack sorting signals presumably would be transported to the cell surface by a default pathway, which has not been conclu-sively identified in kidney cells. The identification of specific sorting signals should be greatly simplified by finding small proteins that are transported directionally to either apical or basolateral membranes.
The small size and the unusual topology of M2 make it an attractive probe to analyze plasma membrane biogenesis, and the transport and ultrastructural distribution of the M2 protein have not been determined in polarized epithelial cells. Therefore, we wished to determine whether M2 was transported to the surfaces of polarized epithelialcells via a directional pathway. Previous studies have focused on the polarized transport of classical type I or II transmembrane glycoproteins, and the expression of type I signal-anchor proteins in polarized cells hasnotbeenreported. In addition, an analysis of M2 expression and localization in polarized cells, which restrict influenza virus glycoprotein expression and virus budding to the apical domain (20, 31, 43, 46), would be of interest in order todeterminewhetherthe site of M2 expression is correlated with the site ofvirusbudding. In this study, we used amonoclonal antibody specific for the external domain of M2 (57) in immunofluorescence, immu-noelectron microscopy, surface radioimmunoassay, and plasma membrane domain-specific immunoprecipitation as-says to investigate the transport and distribution of M2 in polarized epithelial cells.
MATERIALS AND METHODS
Cells and viruses. All cells wereobtained from the Amer-ican Type Culture Collection and maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 100 U of penicillin per ml, 100 ,ugofstreptomycin per ml, and 10% newborn calfserum or 9%newborn calfserum plus 1% fetal calf serum. TK- 143 cells were grown in DMEM containing 25,ug of5-bromodeoxyuridine perml. Stocks of influenza virus
A/WSN/33
(HlNl) were prepared in MDBKcells and titered by plaque assay in MDCK cells. Vaccinia virus recombinant stocks were grown in CV-1 cells and titered by plaque assay in CV-1 and Vero C1008 cells. Recombinant vaccinia virus VV-HA, which contains the influenza virus
A/WSN
HA gene, was provided by Y. Matsuoka (32). Recombinant vaccinia virus VVpSC11,whichexpresses the
P-galactosidase
gene,wasprovided byR. Owens. Vaccinia virus was concentrated by
centrifuga-tion through asucrose cushion (35% sucrose in 10 mM Tris [pH9.1])for 80minat13,500 rpm,resuspendedin 1 mM Tris buffer (pH 9.1), and stored at -70°C.
Polarity assays. The polarity of cells grown on
0.45-p,m-pore-size nitrocellulose filters (Milli-HA filters; Millipore
Corp., Bedford, Mass.) was determined by measuring the transepithelial resistance and the ability to exclude
[3H]inu-lin across monolayers (7) andby examining the polarityof influenza virusbuddingbyelectronmicroscopy. Inaddition,
the distribution of a monoclonal antibody designated D6,
which reacts specifically with basolateral membranes of MDCK cells, was assessed by immunofluorescence and
11I-labeled
antibody binding.Antibodies and immunological reagents. Monoclonal anti-bodies specific for the extracellular domain of M2 (57) or
parainfluenza type 3 virus HA-neuraminidase (42) were
described previously. A hybridoma line secreting a
mono-clonal antibody specific for influenza virus AIWSN HA,
designated 21J, was derived from BALB/c mice immunized with purified influenza virus. The antibody specificity was determined by immunofluorescence and immunoprecipita-tion assays using influenza and VV-HA virus-infected MDCKcells. The hybridoma line secretingthe monoclonal antibody D6, which reacts with a MDCK cell basolateral antigen, was derived fromBALB/c miceimmunizedwith a smooth membrane fraction that was obtained from
homog-enized influenza virus-infected MDCK cells and separated by centrifugation on a 25 to 60% sucrose step gradient.
Hybridoma culture supernatants were screened using indi-rect immunofluorescence on uninfected MDCK cells. The basolateral polarity of the D6 antigen wasconfirmedby using
"2I-labeled
antibody binding to filter-grown MDCK cells.Affinity-purified fluorescein- or rhodamine-conjugated sec-ondary antibodies were obtained from Southern
Biotechnol-ogy Associates, Inc. (Birmingham, Ala.). Protein A- and proteinG-Sepharose were obtained from Pierce (Rockford,
Ill.). Goat anti-mouse immunoglobulin (Ig) conjugated to colloidalgold wasobtained fromProbe Tech, Inc. (Perkasie,
Pa.).
Constructionofrecombinantvaccinia virus VV-M2. A TK-recombinant vaccinia virus that expresses M2 under the control of anearly vaccinia virus promoter was constructed and designated VV-M2. Briefly, the influenza A/Udorn/72
virus M2 cDNA (23) was inserted intothe pSC11vaccinia
virusinsertionplasmid (9). TK- 143 cells weremaintainedin DMEM minus 5-bromodeoxyuridine for 18 h and then in-fected with wild-type vaccinia virus(IHD-Jstrain;
multiplic-ity of infection[MOII = 1) for 2 h at 37°C. Tenmicrograms
ofpSC11-M2 plasmid DNA and25 ,g oflipofectin reagent
(BRL Life Technologies, Inc., Gaithersburg, Md.) were combined, incubated for 15 min at20°C, and then added to the infected cells. After 20 h at 37°C, the cells exhibited
extensive cytopathology, and virus washarvestedbythree
freeze-thaw cycles followed by brief sonication. Recombi-nant virus was passaged twice in TK- 143 cells incubated
with 25 ,ugof5-bromodeoxyuridine per ml and isolated by three rounds of plaque purification in TK- 143 cells incu-bated with 300 ,ug of X-Gal
(5-bromo-4-chloro-3-indolyl-3-D-galactopyranoside) per ml and screenedby indirectimmu-nofluorescence and immunoprecipitation assays using M2 antibody.
Immunofluorescence assays. MDCK cells were grown to confluence on 12-mm glass coverslips and infected with influenza virus (MOI = 10) for 1 h at 37°C, and then the inoculum was replaced with DMEMplus 2% newborn calf serum. At intervals postinfection
(p.i.),
the infected cells were reacted with either M2- orHA-specific
antibody
(as-citic fluids diluted 1:1,000 inphosphate-buffered
saline[PBS]-1%bovine serum albumin
[BSA]).
Primary
antibody
wasvisualized usingfluorescein-conjugated
secondary
anti-body diluted to 10 ,ug/ml in PBS-1% BSA. Allantibody
reactions were performed at 4°C for 30 min. Cells were mounted and photographed with a Nikon
Optiphot
micro-scope.
"51-labeled
antibody binding assays of influenza virus-infected MDCK cells. MDCK cellswereseededon 0.45-,m-pore-size nitrocellulosefilters at adensity
of -5 x104
cells per 12-mm filter and infected with influenza virus(MOI
=10)
2 days after
reaching
confluence. At intervals from 2 to 10 h p.i., the cellswerefixed with 1%paraformaldehyde
in PBS containing0.1 mMCa2"
and 1.0 mMMg2"
(PBS-CM)
and thenquenchedwith50mM
NH4Cl
in PBS.Nonspecific
on November 9, 2019 by guest
http://jvi.asm.org/
5544 HUGHEY ET AL.
backgroundwas reduced by adding3% BSAto wash
solu-tions. Apical orbasolateral cell surfaceswere reactedwith
M2 or HAantibodies (diluted 1:1,500 in PBS-3% BSA) for
30minat4°Cand thenwashedfivetimes(10mineach)with
PBS-3% BSA. Apical or basolateral membranes were
re-acted with 251I-labeledrabbit anti-mouseIgG (2.5 ,ugofIgG
perml; -150,000cpm per filter)for30minat4°C,whilethe
opposite surface ofthe filterwas incubated with 2.5 p,gof
unlabeled rabbit anti-mouse IgG per ml. The filters were
washed five times, allowed to dry, excised from the filter
holder, immersed in scintillation fluid, and counted in a
Beckman liquid scintillation counter. Nonspecific back-groundwas determinedby reacting the infected cellswith
25I-labeled antibodyin the absence ofprimary antibodyand
was subtracted from theexperimental samples.
Membrane domain-specific cell surface
immunoprecipita-tionassays. MDCK cells were seededon 0.45-,tm-pore-size filters at adensityof0.5 x 106 to 1.0 x 106 cellsper30-mm
filterand infected with influenzavirus(MOI=10)2to 3days
after reaching confluence. Cells were incubated in DMEM
minus cysteine from 5 to 6 hp.i., pulse-labeled for 15 min
with
[35S]cysteine
(200 ,uCi) applied to the basolateralsur-face, chased incompleteDMEM for0 to 120
min,
andthenwashed twice with PBS-CM and once with PBS-CM-3%
BSA at4°C. Apicalorbasolateralsurfaceswerereactedwith
M2 or HA antibody (diluted 1:1,000 in PBS-CM-3% BSA)
for 60 min at 4°C and then washed extensively with
PBS-CM. To block unreacted antibody binding sites, the cells
were solubilized with lysis buffer (150 mM NaCl, 10 mM
Tris,5 mMEDTA,1% TritonX-100, 1%deoxycholate,0.1%
sodium dodecylsulfate [SDS]; pH 7.8) containingan
equiv-alentnumber ofunlabeledinfluenzavirus-infectedcells,and
then thelysateswerecentrifugedto removeinsoluble
mate-rial. The immune complexes were reacted with protein A-Sepharose (precoated with rabbit anti-mouse IgG and
preadsorbedwithunlabeledvirus-infectedcelllysates)for60
minat4°C,washedfourtimes withlysisbuffer,solubilized in
reducing buffer, and analyzed on SDS-12.5%
polyacryl-amidegels. Cellsurfaceimmunoprecipitationwasalso used
to detect M2 and HA on the surfaces of polarized Vero
C1008 cells infected with recombinant vaccinia viruses.
VeroC1008 cells inplasticdisheswereinfectedwithpurified
VV-M2 or VV-HA virus (MOI = 10) and radiolabeled
continuouslyfrom5 to8hp.i.with[355]cysteineor
[3S]me-thionine-cysteine, respectively. At 8 h p.i., the cells were
washedwithPBS, reacted with M2orHAantibodiesfor30
minat 4°C, and solubilized in lysis buffer containing
unla-beled recombinant virus-infected cell lysates. Nonspecific background was determined by reacting the surfaces of
VVpSC11virus-infectedcellswithM2orHAantibody. The
immunecomplexes wererecovered using protein
G-Sepha-rose (preincubated with unlabeled recombinant
virus-in-fected cell lysates) and analyzed on SDS-12.5%
polyacryl-amidegels.
125I-labeled
antibody binding assays of VV-M2virus-in-fectedVeroC1008 cells. Avariationofthe antibody binding assay discussed abovewasusedto quantitate M2onapical
and basolateral surfaces of Vero C1008 cells infectedwith
VV-M2.Vero C1008 cells were infected 5 to 7 days after seeding at a density of -5 x 104 cells per 12-mm filter.
Recombinant vaccinia virus (MOI = 5) was allowed to
adsorb for 2 h, and then the inoculum was replaced with
DMEM-2% calf serum. At intervalsfrom 4 to12 hp.i., the samples wereremoved, washed twicewith PBS, andfixed
with 1% paraformaldehyde in PBS. The antibody labeling
was performed as described above. For comparison, Vero
C1008 cellswereinfected withVV-HAandassayed with HA antibody. Nonspecific background was determined by react-ing recombinant virus
VVpSC11-infected
cells withprimary
and secondary antibodies and was subtracted from the
experimental samples.
Immunoelectron microscopy. MDCK cells weregrown in 24-wellplastic plates and infected with influenza virus
(MOI
= 15) for 1 h at 37°C. The cellsweremaintained in DMEM-2% calfserumuntil 7 hp.i., rinsed three times with PBS at 4°C, and reacted on apical surfaces with M2orHA
antibody
(ascitic fluids diluted 1:1,000 in PBS-0.1% BSA) for 1 h at
4°C.After extensivewashings, the cells were reacted witha secondary antibody conjugated to 10-nm-diameter colloidal
gold (diluted1:50 inPBS-0.1%BSA) for 1 h at 4°C. The cells werefixed with 1%glutaraldehydein PBS andpostfixed with 1% osmium tetroxide in PBS for 1 h each at4°C.Specimens wereembedded in epoxy resin for electron microscopy and examined witha Philips EM301microscope.
Quantitation of immunolabeling by electron microscopy.
The distribution of colloidal gold particles on apical cell membranes and virionswas quantitatedbydetermining the
apparentassociation ofgold particles with cellular and virion membranes inphotomicrographsfrom four separate experi-ments. Influenza virus-infected cells reacted with monoclo-nal antibody specific for the HA-neuraminidase protein of
parainfluenza type 3 virus (42) and the gold probe and uninfected MDCK cells reacted with M2orHAantibodyand thegold probewereexaminedasnegative controls toensure the specificity of the immunolabelingreagents. No
signifi-cantbindingof influenzavirus-specificprimary antibodyto uninfected cellsorofheterologousIgstoinfected cellswere observed. To compare the specific label density on apical
cell membranes and virions, a Bioquant II quantitative
morphometry
program was used to measurethe lengths of cellular and viral membranes in individual electronmicro-graphs
of infected MDCK cells. The apparentdistribution ofgold particles on apical and virion membranes was then
determined,
and the label density (in gold particles permicrometer)oncellular and viral membranes was calculated. RESULTS
Expression
of M2 ininfluenza virus-infected MDCK cells. Thesynthesis
of the M2 and HAproteinswascompared in influenza virus-infected MDCK cells thatwerepulse-labeled for 30 minathourly
intervals from 3 to 7 h p.i. (Fig. 1). Maximum HAsynthesis
occurredearly
in the infection, around 4to5 hp.i.,
whereas M2protein synthesisoccurred athighest
levels at approximately6 h p.i. This result is inagreementwithrecentwork
indicating
thatthe ratio of M2 to Ml mRNAs in MDCK cells increases duringthe course of influenza virusinfection,
with an abrupt increase in thesplicing efficiency
to form M2 mRNA predicted to occur around 5 hp.i. (51).
When the cells were radiolabeledcontinuously
from 8 to 10 h p.i., we observed two bandscorresponding
toHAmoleculesthat differ in oligosaccharidecomposition
(12). However,
no change in the migration of M2wasobservedduring
the 2-hlabelinginterval, consistent withprevious
reports(54).The timecourseof appearance andthe distribution of the M2
protein
on the surfaces of influenza virus-infected MDCK cellswereinvestigated
by indirectimmunofluores-cenceand
compared
with cellsurface HA. Polarized MDCK cellswereinfectedwithinfluenzavirus(MOT
= 10), reacted with monoclonalantibody
specific for the extracellular do-main of M2orHAatintervalsfrom 3to 8h p.i., and stained J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
Time (hpi)
A
3 4 5 6 7
8-10
HA
_ _ HAo
B
Jzs
_
M2
FIG. 1. Timecourseof M2 and HA proteinsynthesisininfluenza
virus-infected MDCK cells. MDCK cells were grown in plastic
dishes, infectedwith influenza virus (MOI = 10),andpulse-labeled
for 30 min at hourly intervals from 3 to 7 h p.i. or labeled
continuously from8to10h p.i. HA (A) and M2 (B)were
immuno-precipitated using monoclonal antibodies and protein G-Sepharose
and analyzedon 7.5 and 12.5% polyacrylamide gels, respectively. M2 doublets in reducing SDS-polyacrylamide gels have been
re-ported previously (24,54).
withafluorescein-conjugated secondary antibody. Cell
sur-face expression of M2 was detected by 4.5 h p.i. (Fig. 2),
while surface HAwasdetectedas early as3 hp.i. (datanot
shown). The apical membranes of virus-infected MDCK cellsexhibitedapunctatefluorescence pattern after labeling withM2 and HA antibodies. However, the M2fluorescence
pattern appeared to be slightly more diffuse than that
ob-served for HA and extended to the cell periphery, giving cells a polygonal appearance, whereas lower levels of HA
fluorescencewereobserved nearregions of cell-cellcontact.
M2 and HA were also colocalized intracellularly (not
shown), although HAwasdetected earlier thanM2,
consis-tentwith thetime courseof protein synthesis.
Itwasreported previouslythat influenza virus buddingas
well asexpression of the viral glycoproteins, HA and
neur-aminidase, are restricted to apical membranes of polarized epithelial cells (20, 31, 43, 46). Therefore, we wished to
examine the kinetics of M2 accumulation on apical and
basolateral membranes of influenza virus-infected MDCK cells that weregrown on permeable nitrocellulose filters to allow antibody access to either membrane surface. For comparison,we quantitatedthe binding of the D6
monoclo-nal antibody, which recognizes anantigen present on
baso-lateral membranesof MDCKcells, toapical and basolateral surfaces and found that 94% of the total D6 binding was
restrictedtothebasolateral domain of MDCKcells. Surface expression of M2andHAonapical andbasolateral surfaces
of influenza virus-infected MDCK cells was assayed at intervals from 2 to 10 h p.i. using M2 and HA primary antibodies and 1251I-labeled secondary antibody. We found thattheexpressionof HAonapicalsurfaces precededthat of
M2 by -2 h (Fig. 3). M2 and HA cell surface expression increased at approximatelysimilar ratesfrom 6to 10 h p.i., while the ratio of M2 to HA antibodies bound to apical membranes increasedfrom -1:6 at 6 h p.i. to -1:2 at 10 h p.i. M2was detected on the apical surfaces of MDCKcells
grownonglasscoverslipssomewhatearlier, around4to5 h p.i., buttherateofsurfaceaccumulationwasnotfoundtobe appreciably different from that observed with the
filter-grown cells (data not shown). Both M2 and HA were
predominantly expressed on apical membranesof influenza
virus-infected MDCK cells throughout the infection time course, and approximately 84 to 86% of the total M2
antibody and90 to 93% of the HA antibody were bound to apical surfaces at 8 to10 hp.i.Thus, the M2 andHAproteins
exhibit similar patterns of polarized expression on apical membranes.
Different transport pathways havebeen reported for
tar-geting of proteins to apical plasma membranes (6). Apical proteins in hepatocytes are transported exclusively via an indirectpathway, appearingfirst onbasolateral membranes
before being endocytosed and redirected to the apical sur-face (1). In contrast, intestinal cells appear to useboth direct andindirect pathways to the apicalsurface,dependingupon the protein in question (25, 33). The majority of apical membraneproteins inkidney cellsappear to be transported
exclusively by a direct pathway that does not involve the basolateral surface (26-28, 31), but certain membrane pro-teins, including the polymeric Ig receptor, are transported
to apical surfaces by an indirect pathway when expressed
in MDCK cells (3, 35). Therefore, to examine the kinetics
of transport and the initial site of M2 plasma membrane
insertion in influenzavirus-infected MDCK cells, we used pulse-chase metabolic labeling in conjunction with plasma
membrane domain-specific immunoprecipitation and
SDS-polyacrylamide gel electrophoresis. For comparison, the transport of theHA protein toapical orbasolateral surfaces
ofinfected cells was determined. M2 was first detected on the apical surfaces ofinfluenza virus-infected MDCK cells after a 15-minpulse and60-minchase (Fig. 4). The level of apical M2increasedafter 2 h, while levels of M2proteinon
basolateral membranes were very low at both time points,
consistent with the low levels of basolateral M2 detectedby
1251I-labeled
antibodybinding. Afraction oftheHAproteinwas cleaved to HAl and HA2 during transport, and
signifi-cantlevelsof bothcleavedand uncleaved HA were detected on apical surfaces after 1-h chase. Similar results were
obtained using domain-specific biotinylation and
streptavi-din precipitation (data not shown). These results strongly
suggest that M2 is targeted directly to the apical domain without appearing transiently on the basolateral surface,
similartothe directapical transport of HA.The M2 protein
appears on the plasma membrane at a slightly slower rate than HA, which could be due to a difference in the oligo-merization rate of M2(15, 48) and HA(12) orin the rateof theirtransport to the plasma membrane.
Polarized expression of M2 from a recombinant vaccinia virus. Theobservation that M2expression is predominantly restrictedto apical surfaces of influenza virus-infected cells raised the question whether M2 is directionally
transported
inthe absenceof other viral proteins, ashas been shown for the influenza virus glycoproteins, HA and neuraminidase (20, 46). Alternatively, the polarized transport of M2
might
bedue to an interaction between M2 and other viral
proteins,
such as the glycoproteins. To address this
question,
we constructed VV-M2, a recombinant vaccinia virus that ex-presses the M2 protein. Thepolarized
monkey
kidney
cell line Vero C1008 (45) was usedto analyzeM2transport in the absence of other influenza virusproteins,
as we obtained higher levels of M2expressionwith therecombinant virus in these cells than in MDCK cells. The M2protein
was detected on the surfaces of VV-M2 virus-infected Vero C1008 cells by indirect immunofluorescence(data
not shown) and by cell surfaceimmunoprecipitation (Fig. 5)
using M2 antibody.
Similarly,
HAexpressed
from recombi-nant vaccinia virus VV-HAwasdetected onthe cell surface using surfaceimmunoprecipitation
(Fig.
5).
The site of M2on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.83.278.75.214.2]S!::t 8 t ~~~~~~~~~~~~~~~~~~~O
h a 5 | r B§_~~~~~~~~4
^ A 8 .*, ^> ~.4 S j
Zv~~~~~~rs.
Ai,,'0
_
#z~~~~~~~~~~
FI.elsrfceexresin f n nfuez viu-nfce MDCK cells MDKcel'wr ,inetdwhiflnzAWSvru
(MI=1)adrece
ihmnolnlatbdyseii
o M At_
)o
H_
Dt;
)a 45(
7''.
n60(
n)
5(CadF
h~~~F
piPrmranioywsvsaieusnafloeci-ojugat6ed
seodrniod.Teepsr
tims
ofpoorps
Ieeajse
torema~inM.c.nstn
thoghu
th expermen.5546
on November 9, 2019 by guest
http://jvi.asm.org/
anti-HA anti-M2
V H S M S
HA NP HAl
... .;
0 2 4 6 8 10
[image:6.612.359.522.73.244.2]Time (hpl)
FIG. 3. M2 accumulation on apical and basolateral surfaces of
influenza virus-infected MDCKcells. Filter-grown MDCKcellswere
infected withinfluenzaA/WSNvirus and assayed forM2 (squares)or
HA(triangles)onapical (closed symbols)orbasolateral (open
sym-bols) surfaces using monoclonal antibodiesand 125I-labeled
second-ary antibody. Values represent the 125I-labeled antibody (Ab) (in kilocountsperminute[kcpm])boundtoinfectedcells after subtract-ingnonspecific background andwerederivedfrom themeanvalues of
quadruplicate samples (standarderrorof themean, <15%).Similar
resultswereobtained intwoindependent experiments.
cell surfaceexpressionwasinvestigated in filter-grown
mon-key kidney cells infectedwith the VV-M2 virus using
125I-labeledantibody binding (Fig. 6). The expression of HAon
apical and basolateral membraneswascompared using Vero
C1008 cells infected withVV-HA,arecombinantvirus that
A B
0 1 2
A B A B A B
0 1 2
A B AB A B
M2
FIG. 5. M2 is expressed on the surfaces of Vero C1008 cells
infected with W-M2 recombinant virus. PolarizedVero C1008 cells
were grown in plastic dishes, infected with recombinant virus
W-M2 (M), W-HA (H), or WpSC11 (S), and metabolically
labeled from5to8h p.i. At 8 h p.i., the plasmamembraneswere
reacted with M2orHA antibody for 30min at 4°C.After excess
antibodieswereremoved the cellswere solubilized in lysis buffer
containingunlabeled infected celllysates. The immune complexes
wererecovered usingprotein G-Sepharose and analyzedon12.5%
polyacrylamide gels. Contaminating vaccinia virus bands that
mi-grateconsiderablyfaster than HAwerealso precipitated fromcells
reacted with HAantibody. Lane V, which contains marker proteins frompurified influenza virions,wasoverexposedtovisualize M2.
expressesHA(32). The M2 proteinwasdetected on apical
membranes ofthe recombinantvirus-infected cells 2 h later inthe infectiontimecoursethan HA, eventhough
immuno-fluorescence assays indicated that a similar percentage of cellswasinfected with either virus (datanotshown). Signif-icant levels of HAwere detectedonthe apical surface by 6
hp.i., while M2wasexpressed atlow levelsat 8 hp.i. and HA I
HAl I
E
0
10
C
0
.0 .0
4
HA2
M2
FIG. 4. Kinetics of M2 cell surface appearance in influenza
virus-infectedMDCK cells.Influenza virus-infected MDCK cellson
nitrocellulosefilterswerepulse-labeledfor15minat -6hp.i. and
chased for0 to 2h, as indicated above the lanes. Apical (A) or
basolateral (B)cell surfaceswerereacted at4°CwithM2(A)orHA
(B) antibody,the cellswerelysed,and the immunecomplexeswere
recoveredandanalyzedon12.5%polyacrylamide gels.
4 6 8 10 12
Time (hpi)
FIG. 6. M2 isexpressedonapicalmembranesofpolarizedVero
C1008 cells infected with VV-M2 virus. Filter-grownVero C1008
cellswereinfectedwith VV-M2(squares)orVV-HA(triangles)and
assayed for M2or HA on apical (closed symbols) orbasolateral
(open symbols) surfaces using monoclonal antibodies and 1251_
labeledsecondaryantibody.Valuesrepresent the125I-labeled
anti-body (Ab) (inkilocountsperminute[kcpm])bound to infected cells
aftersubtractingnonspecific backgroundandwerederivedfrom the
mean values of triplicate samples (standard error of the mean,
<15%). Similar results wereobtained inthreeindependent
experi-ments.
20
15
0-E
Q
0 0
c .0
.4
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.74.291.73.225.2] [image:6.612.67.303.409.667.2] [image:6.612.334.552.483.631.2]5548 HUGHEY ET AL.
accumulated at higher levels between 8 and 12 h
p.i.
Both M2 and HA were predominantly restricted to theapical
surfaces ofpolarized monkey kidney cells infected withthe respective viruses. Approximately 88 to 90% of the total
surface HAwasexpressed on the
apical
domainbetween 8and 12 h p.i. compared with 86 to 93% of the surface M2 expressed at 10 to 12 hp.i. Therefore, the
degree
ofapical
polarity was very similar for M2 and HA
expressed
using
vacciniavirus recombinants. The basolateral expression of
HIV gp160 has been demonstrated previously by a similar assay in polarized epithelial cells infected with a
recombi-nant vaccinia virus(36). These results demonstrate that the localization of M2 and HA is similar in polarized epithelial
cells infected with influenza virus or the recombinant
vac-cinia viruses and indicate that the M2 protein contains the
information necessary for directional transport to apical
membranes in the absence ofotherinfluenzavirus proteins.
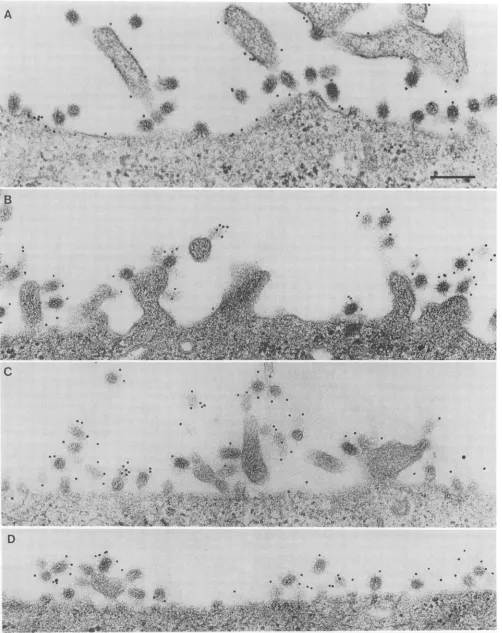
UltrastructurallocalizationofM2 ininfluenza virus-infected cells. The localization ofthe M2 protein in influenza
virus-infected MDCK cells was examined at the ultrastructural level using immunoelectron microscopy. At 7 h p.i., atime
when extensivevirusbuddingwas occurring, the cellswere
labeled at 4°CwithM2-specific monoclonal antibody and a
gold probe (Fig. 7). For comparison, virus-infected cells
were also labeled with monoclonal antibodies specific for
HAor anunrelated protein and thegold probe. M2-specific
labeling was observed on apical membranes, including
mi-crovilliofinfluenzavirus-infected MDCKcells. In addition, gold particles were associated with fully assembled and
releasedvirions andwereclose toboth cell membranes and budding virions. Infected cells labeled with HA-specific antibody exhibited similar distributions of gold particles (Fig. 7).Nolabelingwasobservedonbasolateral surfaces of
filter-grown MDCK cells reacted with M2 antibody and secondary antibodyconjugated with goldorferritin(datanot
shown).The specificity of the immunolabeling reagentswas
demonstrated by the lack ofsignificant binding of M2 anti-body to uninfected-cell surfaces or of the gold probe to
infected cells in the absence of influenza virus-specific antibodyorin thepresenceofanunrelated antibodyspecific
forthe HA-neuraminidase protein (42) of parainfluenzatype
3virus (data not shown).
The distribution of M2 labelwas determined by
quantitat-ing the number of gold particles associated with either cell membranes or budding influenza virus particles on MDCK
cells labeled with M2 antibody at 7 h p.i. (Table 1). A
significant proportion of the M2 label was found to be
associatedwith partially and completely assembled virions. Electronmicrographs of virus-infected MDCK cells labeled
withM2 antibodyand the gold probewere analyzed further
by quantitative morphometry to estimate the relative
con-centrationsofM2 labelonvirion and apicalcell membranes.
These calculations indicate that the density of M2-specific
label associated with viral or cellular membranes was
roughly equivalent, suggesting that M2 was neither
specifi-cally enriched in virion membranes compared with cell
membranes nor excluded from budding virus particles.
DISCUSSION
We observed that the M2 protein accumulates
predomi-nantly on apical surfaces of influenza virus-infected
polar-izedepithelialcells withkinetics similartothose of HA. The
delayinM2surfaceexpressioncompared with that ofHAis
apparentlydue tothe time courseof M2 protein synthesis,
which isdelayedinpartbecause of the inefficientsplicingof
segment 7mRNA(51). The finding that M2surface expres-sion was restricted totheapical domainofpolarized
epithe-lial
cells infected with recombinant virus VV-M2 suggests that M2contains intrinsic structural features thatare recog-nized by the cellular transportmachinery,despite
the small size andunusual topology of themolecule. Furtheranalysis
of signals for M2 transport using site-directed
mutagenesis
will be of interest for the identification of the structural
requirements for apical transport. The determinants of
sig-nal-mediated and defaulttransportpathways tothe surfaces
of kidney cells remain unresolved, and it is uncertain
whether specific sortingsignals arerequired fortransportto apical orbasolateral membranes orboth membranes. Non-directional default transport would indicate that proteins lacking sorting information are incorporated nonspecifically into transport vesicles departing from the trans-Golgi net-work. Conversely, directional transport by a default mech-anism would suggest that proteins that lack sorting signals are excluded from the vesicles involved in signal-mediated transport. Therefore, if M2 transport occurs by default, our findings would support the idea that the default pathway for membrane protein transport is restricted to apical surfaces of kidney cells.
Specific structural features which correlate with direc-tional transport have been recently identified in otherprotein molecules. A subset of membrane proteins that contain a covalently attached glycophosphatidylinositol residue is re-stricted to the apical domain of polarized epithelial cells (28). A signal directing basolateral transport in MDCK cells is formed by a 14-residue, arginine-rich sequence located ad-jacent to the membrane in the polymeric Ig receptor cyto-plasmic domain (8). A mutated protein lacking this sequence was transported directly to the apical surface, while a chimeric protein containing the signal was redirected from the apical to basolateral surface. A different signal for basolateral transport was identified in a mutated influenza virus HA which was targeted directly to the basolateral domain of MDCK cells after a single cytoplasmic cysteine residue was changed to tyrosine (4). Specific signals in the cytoplasmic domains of several other proteins, including the Fc receptor, lysosomal glycoprotein
lgpl20,
and low-density lipoprotein receptor, are required for basolateral expression in MDCK cells (18, 19). The finding that mutated proteins lacking putative basolateral targeting signals are predomi-nantly expressed on apical surfaces of MDCK cells suggests that basolateral signals are dominant over apical signals in kidney cells or that proteins which lack basolateral targeting signals are transported to apical surfaces by default (4, 8, 16, 18, 19). The available evidence suggests a correlation be-tween basolateral polarity and endocytosis of certain pro-teins (4, 8, 16, 18, 19, 35) that contain a critical tyrosine residue in cytoplasmic sequences that function as internal-ization signals (10, 21). The mutation or deletion of specific cytoplasmic tyrosine residues profoundly alters the polarity and endocytosis of certain proteins (4, 18, 19, 27, 35). It is now evident that cytoplasmic tyrosine residues are not sufficient for basolateral targeting, since we found that the M2 protein was predominantly restricted to the apical do-main of polarized cells despite the presence of two or three cytoplasmic tyrosine residues (23, 29) in sequences that are similar to proposedinternalization
signals (10, 21). Both the position of tyrosine in the cytoplasmic domain and the specific amino acids thatsurround
tyrosine residues appar-ently influence the ability of these sequences to serve as targeting signals for endocytosis or basolateral transport (16, 18, 19, 21, 27). Therefore, it is likely that the cytoplasmic J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
A
.*Jt
',-
4k,".. .V,
. C.
0
N*
1o74
4V~~~~4
*t#4SS fir S~~~~~~~~~~~~~~~~~~~~~~~A
*
%Arr~~~~~~~~~~~~~~1
B
..;
6.
.0.
t4s
-
.:*
4.
11
11a .. 94 --1W
.Af
1.
C . ...
a S *
*
, &
aFa
I.
.:
-;0 0'
FIG. 7. Distribution ofM2 and HA on the surfaces of influenza virus-infected MDCKcells.MDCKcellswere grown on plastic dishes, infected with influenzaA/WSNvirus(MOI = 15) andlabeled withmonoclonal antibodyspecific forM2 (A and B) or HA (C and D) at 7 h p.i. Primaryantibody was visualized using a secondary antibody bound to 10-nm-diameter goldparticles. Bar,0.2 um.
5549
..*
..
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.60.562.73.706.2]5550 HUGHEY ET AL.
TABLE 1. Distribution of M2 label on apicalmembranes of influenzavirus-infected MDCK cells and virions
Locationof Membrane No.of % of total
No.
of M2 label length(,um) particles particlespartocles/,um
Cell 49.1 107 22 2.2
Virus 55.7 196 40 3.5
Uncertainb 104.8 187 38 1.8
Total 104.8 490 100 4.7
a The label concentrationwasdeterminedbyquantitatingthecumulative lengths(inmicrometers)of viral andapicalcell membranes and thenumber anddistribution ofgold particles.Thedensityofparticlesin the uncertain
category isrepresentedasthe number of suchparticlespertotallengthofviral
and cellular membranes.
b Someparticles could notbeclearlylocalizedtoeitherapicalcell
mem-branes or virionsand werescoredasuncertain.
tyrosineresidues in M2arenotin the properenvironmentto function asbasolateral
targeting
signals.
We have observedthat M2 is not excluded from coated
pits
when influenza virus-infected cells areincubated at37°Cwith M2antibody
(16a), but it has not been determined whether M2 isinter-nalized to a significant extent in the absence of
antibody.
Furtheranalysisof the polarityand endocytosis of M2and
other proteins containing
cytoplasmic tyrosines
will be re-quiredto clarifyrelationshipsbetween theseevents.The punctate immunofluorescence pattern observed for M2 on apical surfaces of influenza virus-infected MDCK
cells was similar to that observed for HA. However, the
slightly more diffuse fluorescence pattern observed for M2
appeared to reflect subtle differences in the ultrastructural localization of the two proteins with respect to
budding
virions, presumably duetodifferencesin M2 and HA incor-poration intovirions(57).
Immunoelectronmicroscopy
fur-ther localized M2 to microvilli and thesurrounding
apicalcell membrane ofvirus-infected MDCK cells and to com-pletelyassembled andbuddingvirions,which didnotexhibit
obviousdifferencesin thefrequencyordensityof M2label.
Itwassurprisingthatalargepercentage of the M2labelwas foundtobevirion-associated andthat the apparent density
of M2 label on viral and cellular membranes was roughly equivalent, considering the low level of M2 incorporation into virions (57) comparedwith the large number of mole-culesontheinfected-cell membrane (24).The restriction of M2totheapicaldomainofpolarizedcells, which is the site ofinfluenza virus budding, and the association of M2 with both cellular membranesandnascentvirionsareconsistent with apossible role for M2 in virus assembly. However, the
mechanism responsible for excluding the majority of M2
moleculesfromvirions such that only a few molecules are
incorporated duringvirus assembly is not known. As treat-ment of virions with the drug amantadine hydrochloride inhibits virus uncoating (5), and the antiviral effect is exerted on M2 function (2, 14), it is likely that the M2 molecules
incorporated intovirions play a role in viral infectivity. Ithas been suggested that the M2 protein may be associ-ated with an ion channel activity (47, 49), and direct evi-dence for a pH-activated ion channel has been obtained by
electrophysiological measurements of Xenopus laevis oocytes injected with M2 mRNA (41). Alterations in the
intracellularenvironment induced by M2 might affect virus assemblyindirectly, as the late stages of protein transport aresensitive to ionic conditions (30, 47). On the basis of the HAconformational changes that can occur in the presence ofamantadine (47), it was suggested that M2 influences the
stability of HA trimers duringtransportand that M2might
function to offset intracellular
pH
changes induced by the shutdown of host protein synthesis during virus infection(34, 47).
Inhibitionby
amantadinemight
be due to an interaction between the drug and M2 in virion or cellmembranes that alters the ion channel function of M2 (41)
and may
disrupt
eitherearlyorlate stages ofreplication(5,
14, 47, 49). Current evidence suggests that M2 might func-tion to influence interactions among virion components duringvirus entryorbudding.A monoclonalantibody to the externaldomain of M2 inhibitedinfluenzavirusreplicationin a strain-specific manner (50, 57, 58), and the mutation ofspecificresidues in the cytoplasmic domain of M2 or in the
Mlproteinconferred resistancetotheM2antibody (58).The
mechanism that targets Ml to sites of virus budding is not yet known, but immunoelectron microscopic studies indi-cated thatonceMl and thenucleocapsidassociate with the
plasma membrane,virusbuddingisrapidly completed(39).
TheMlprotein is reported to associate spontaneously with
lipids by inserting specific regions of the molecule into the
bilayer (13, 56), and the association of Ml with the virus
envelopeappearstobepHsensitive(60).The presentfinding
thatM2 labelwasfrequentlyassociated with nascentvirions is consistent with the hypothesis that the affinity of the Ml protein for the plasma membrane may be influenced by M2 ionchannelsorbyatargeting signal in the M2 molecule. In
addition, virion-associated M2 may facilitate uncoating in secondary endosomes by permitting a flux of ions into virions and altering interactions between Ml and the viral envelope orribonucleoproteincomplexes (5, 41,60).
There-fore, it is conceivable that M2 may function, perhaps in a pH-sensitive mechanism, to regulate the association be-tween internal virion components and the viral envelope during virus uncoating and assembly stages ofreplication.
ACKNOWLEDGMENTS
Wethank Eugene Arms and Skip Melsen forexpert assistance withphotography and electronmicroscopy. We thankYumi Mat-suoka andRandy Owens for providingrecombinantvacciniaviruses and David Kilpatrick for advice on constructing the recombinant vaccinia virus W-M2. We also thank Marianne Accavitti, Fran Allen, and Diane Williams of the University of Alabamaat Birming-hamHybridomaCoreFacility for help inpreparinghybridomasand BettyJeffreyforsecretarial assistance.
This work was supported by research grants Al 12680 and Al 20201 from the National Institutes ofAllergy and Infectious Dis-eases andCA 18611 fromtheNationalCancer Institute.P.G.H. was supported in part by Institutional Research Service Award HL 07553 from theNational Heart, Lung, and BloodInstitute.
REFERENCES
1. Bartles, J. R., H. M. Ferracci, B. Stieger, and A. L. Hubbard. 1987. Biogenesis of the rat hepatocyte plasma membrane in vivo: comparisonofthe pathways taken by apical and basolat-eral proteins using subcellular fractionation. J. Cell Biol. 105: 1241-1251.
2. Bean, W. J., S. C.Threlkeld,andR. G.Webster.1989. Biologic potentialofamantadine-resistant influenzaAvirus in an avian model. J.Infect. Dis. 159:1050-1056.
3. Brandli,A. W., and K. Simons. 1990. Transcytosisin MDCK cells: identificationofglycoproteins transported bidirectionally between both plasma membrane domains. J. Cell Biol. 111: 2909-2921.
4. Brewer, C. B., and M. G. Roth. 1991. A single amino acid change in thecytoplasmicdomain alters thepolarized delivery ofinfluenzavirushemagglutinin.J. Cell Biol. 114:413-421. 5. Bukrinskaya,A.G.,N. K.Vorkunova,G. V.Kornilayeva,R. A.
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.62.302.98.172.2]Narmanbetnova, and G. K. Vorkunova. 1982. Influenza virus uncoating in infected cells and effect of rimantadine. J. Gen. Virol. 60:49-59.
6. Caplan, M., and K.S. Matlin. 1989. The sorting of membrane andsecretory proteins in polarizedepithelial cells, p. 71-127. In K. S. Matlin and J. D. Valentich (ed.), Functional epithelial cells in culture. Alan R. Liss, Inc., NewYork.
7. Caplan, M. J., H. C. Anderson, G. E. Palade, and J. D. Jamieson. 1986.Intracellular sorting and polarized cell surface delivery of (Na+,K+) ATPase, an endogenous component of MDCK cell basolateral plasmamembranes. Cell 46:623-631. 8. Casanova, J. E., G. Apodaca, and K. E. Mostov. 1991. An
autonomous signal for basolateral sorting in the cytoplasmic domain ofthe polymericimmunoglobulin receptor. Cell 66:65-75.
9. Chakrabarti, S., K. Breechling, and B. Moss. 1985. Vaccinia virus expression vector: coexpression of beta-galactosidase provides visual screening ofrecombinant virus plaques. Mol. Cell. Biol. 5:3403-3409.
10. Collawn, J. F., M. Stangel, L. A. Kuhn, V. Esekogwu,S.Jing, I. S.Trowbridge, and J. A. Tainer. 1990.Transferrin receptor internalization sequence YXRF implicates a tight turn as the structuralrecognition motif for endocytosis. Cell 63:1061-1072. 11. Fuller, S. D., R. Bravo, and K. Simons. 1985. An enzymatic assayrevealsthat proteins destined for the apicalorbasolateral domains of an epithelial cell line share the same late Golgi compartments.EMBO J.4:297-307.
12. Gething, M.-J., K.McCammon, and J. Sambrook. 1986. Expres-sion of wild-type andmutantforms ofinfluenza hemagglutinin: therole offoldinginintracellular transport. Cell46:939-950. 13. Gregoriades, A., and B.Frangione. 1981.Insertion of influenza
Mprotein into the viral lipid bilayerandlocalization of site of insertion.J.Virol. 40:323-328.
14. Hay, A. J., A. J.Wolstenholme,J. J. Skehel, and M. H. Smith. 1985.Themolecular basis ofthespecific anti-influenza action of amantadine. EMBO J. 4:3021-3024.
15. Holsinger, L. J., and R. A. Lamb. 1991. Influenza virus M2 integral membrane protein is a homotetramer stabilized by formation of disulfidebonds. Virology 183:32-43.
16. Hopkins, C. R. 1991. Polaritysignals. Cell 66:827-829. 16a.Hughey, P. G., et al.Unpublisheddata.
17. Hull, J. D., R. Gilmore, and R. A.Lamb. 1988. Integration of a smallintegral membrane protein,M2, ofinfluenza virus into the endoplasmic reticulum: analysis of the internal signal-anchor domain ofaproteinwith anectoplasmic NH2terminus. J. Cell Biol. 106:1489-1498.
18. Hunziker, W., C. Harter, K. Matter, and I. Mellman. 1991. Basolateral sorting in MDCK cells requiresadistinct cytoplas-mic domain determinant. Cell 66:907-920.
19. Hunziker, W., andI. Mellman. 1989. Expressionof macroph-age-lymphocyte Fc receptors in Madin-Darby canine kidney cells: polarity and transcytosis differ for isoforms with or without coatedpit localization domains.J.Cell Biol. 109:3291-3302.
20. Jones,L.V., R. W. Compans, A. R. Davis, T. J. Bos, and D. P. Nayak.1985. Surface expression of influenzavirus neuramini-dase, anamino-terminally anchored viral membrane glycopro-tein, inpolarizedepithelial cells. Mol. Cell. Biol. 5:2181-2189. 21. Ktistakis,N.T.,D.Thomas,and M.G.Roth. 1990. Character-istics of the tyrosine recognition signal for internalization of transmembrane surface glycoproteins. J. Cell Biol. 111:1393-1407.
22. Lamb,R. A. 1989. The genes andproteinsofinfluenzaviruses, p. 1-87. In R. M. Krug(ed.), The influenza viruses. Plenum, NewYork.
23. Lamb,R.A.,and C.J.Lai. 1981. Conservation of the influenza virusmembraneprotein(Ml)amino acidsequence andanopen readingframe ofRNA segment 7 encoding a second protein (M2)inHlNl andH3N2 strains.Virology 112:746-751. 24. Lamb, R. A., S. L. Zebedee, and C. D. Richardson. 1985.
Influenza virus M2 protein is an integral membrane protein expressedontheinfected-cell surface. Cell 40:627-633. 25. LeBivic,A.,A.Quaroni,B.Nichols,and E.Rodriguez-Boulan.
1990. Biogenetic pathways of plasma membrane proteins in Caco-2, a human intestinal epithelial cell line. J. Cell Biol. 111:1351-1361.
26. Le Bivic, A., Y. Sambuy, K. Mostov, and E. Rodriguez-Boulan. 1990. Vectorial targeting of an endogenous apical membrane sialoglycoprotein and uvomorulin in MDCK cells. J. Cell Biol. 110:1533-1539.
27. Le Bivic, A., Y. Sambuy, A. Patzak, N. Patil, M. Chao, and E. Rodriguez-Boulan. 1991. Aninternaldeletionin the cytoplasmic tail reverses the apical localization of human NGF receptor in transfected MDCK cells. J. Cell Biol. 115:607-618.
28. Lisanti, M. P.,I. W. Caras, M. A. Davitz, and E. Rodriguez-Boulan. 1989. A glycophospholipid membrane anchor acts as an apical targeting signal in polarized epithelial cells. J. Cell Biol. 109:2145-2156.
29. Markushin,S., H. Ghiasi, N. Sokolov, A. Shilov, B. Sinitsin, D. Brown, A. Klimov, and D.Nayak 1988. Nucleotide sequence of RNA segment 7 and the predicted amino acid sequence ofMl and M2 proteins of FPV/Weybridge (H7N7) and WSN(HlNl) influenza viruses. Virus Res. 10:263-272.
30. Matlin, K.S. 1986. Ammonium chloride slows transport of the influenza virus hemagglutinin but does not cause missorting in a polarized epithelial cell line. J. Biol. Chem. 261:15172-15178. 31. Matlin, K. S., and K. Simons. 1984. Sorting of an apical plasma
membrane glycoprotein occurs before it reaches the cell surface in cultured epithelial cells. J. Cell Biol.99:2131-2139. 32. Matsuoka, Y., T. Ihara, D. H. L. Bishop, and R. W. Compans.
1988. Intracellular accumulation of Punta Toro virus glycopro-teins expressed from cloned cDNA. Virology 167:251-260. 33. Matter, K., K. Bucher, and H.-P. Hauri. 1990. Microtubule
perturbation retards both the direct and the indirect apical pathway but does not affect sorting of plasma membrane pro-teins in intestinal epithelial cells (Caco-2). EMBO J. 9:3163-3170.
34. Moore, L. L., D. A. Bostick, and R. F. Garry. 1988. Sindbis virus infection decreases intracellular pH: alkaline medium inhibits processing of Sindbis virus polyproteins. Virology 166: 1-9.
35. Mostov, K. E., A. D. B. Kops, and D. L. Deitcher. 1986. Deletion of the cytoplasmic domain of the polymeric immunoglobulin receptor prevents basolateral localization and endocytosis. Cell 47:359-364.
36. Owens, R. J., and R. W. Compans. 1989. Expression of the human immunodeficiency virus envelope glycoprotein is re-stricted to basolateral surfaces of polarized epithelial cells. J. Virol. 63:978-982.
37. Parks, G. D., J. D. Hull, and R. A. Lamb. 1989. Transposition of domains between the M2 and HN viral membrane proteins results in polypeptides which can adopt more than one mem-brane orientation. J. Cell Biol. 109:2023-2032.
38. Parks, G. D., and R. A. Lamb. 1991. Topology of eukaryotic typeIImembrane proteins: importance of N-terminal positively chargedresidues flanking the hydrophobic domain. Cell 64:777-787.
39. Patterson, S.,J. Gross, and J. S. Oxford. 1988. The intracellular distribution of influenza virus matrix protein and nucleoprotein ininfected cells and their relationship to haemagglutinin in the plasma membrane. J. Gen. Virol. 69:1859-1872.
40. Pfeiffer, S., S. D. Fuller, and K. Simons. 1985. Intracellular sortingandbasolateral appearance of the G protein of vesicular stomatitis virus in Madin-Darby canine kidney cells. J. Cell Biol. 101:470-476.
41. Pinto, L.H., L.J. Holsinger, and R. A. Lamb. 1992. Influenza virus M2 protein has ion channel activity. Cell 69:517-528. 42. Ray, R., and R. W. Compans. 1986. Monoclonal antibodies
revealextensive antigenic differences between the hemaggluti-nin-neuraminidase glycoproteins of human and bovine parain-fluenza 3viruses.Virology 148:232-236.
43. Rodriguez-Boulan, E. J., and D. D. Sabatini. 1978. Asymmetric budding of viruses in epithelial monolayers: a model for the study of epithelial polarity. Proc. Natl. Acad. Sci. USA 75: 5071-5075.
44. Roth, M. G., R. V. Srinivas, and R. W. Compans. 1983.
on November 9, 2019 by guest
http://jvi.asm.org/
5552 HUGHEY ET AL.
Basolateral maturation of retroviruses in polarized epithelial cells. J. Virol. 45:1065-1073.
45. Srinivas, R. V., N. Balachandran, F. V. Alonso-Caplen, and R. W. Compans. 1986. Expression of herpes simplex virus glycoproteins in polarized epithelial cells. J. Virol. 58:689-693. 46. Stephens, E.B., R. W. Compans, P. Earl,andB. Moss. 1986. Surfaceexpression of viralglycoproteins ispolarizedin epithe-lial cells infected with recombinant vaccinia viral vectors. EMBOJ.5:237-245.
47. Sugrue,R.J.,G.Bahadur,M.C.Zambon,M.Hall-Smith,A. R. Douglas, andA. J.Hay. 1990. Specific structural alteration of the influenzahaemagglutinin by amantadine. EMBO J. 9:3469-3476.
48. Sugrue,R.J.,R. B.Beishe,and A.J.Hay.1990.Palmitoylation of theinfluenzaAvirusM2protein. Virology 179:51-56. 49. Sugrue,R.J.,and A.J. Hay. 1991.Structuralcharacteristics of
theM2protein ofinfluenzaAviruses: evidence that it formsa tetramericchannel.Virology 180:616-624.
50. Treanor, J. J., E. L. Tierney,S. L.Zebedee, R. A. Lamb, and B.R.Murphy.1990.Passively transferred monoclonal antibody totheM2protein inhibits influenzaAvirus replication in mice. J.Virol. 64:1375-1377.
51. Valcarcel, J., A. Portela, and J. Ortin. 1991. Regulated Ml mRNAsplicingininfluenza virus-infected cells.J. Gen. Virol. 72:1301-1308.
52. van Meer, G., and K. Simons. 1986. The function of tight junctions in maintaining differences in lipid composition be-tween the apical and the basolateral cell surface domains of
MDCK cells. EMBO J. 5:1455-1464.
53. van Meer, G., E. H. K. Stelzer, and R. W. Wijnaendts-van-Resandt. 1987. Sorting of sphingolipids in epithelial (Madin-Darbycaninekidney)cells.J. Cell Biol.105:1623-1635. 54. Veit, M., H.-D.Klenk,A. Kendal, and R. Rott. 1991. The M2
proteinofinfluenza A virus isacylated.J.Gen. Virol. 72:1461-1465.
55. Wandinger-Ness, A.,M. K.Bennett,C.Antony,and K.Simons. 1990. Distincttransportvesicles mediate thedeliveryofplasma membrane protein to the apical and basolateral domains of MDCK cells. J. Cell Biol. 111:987-1000.
56. Ye, Z.,R.Pal,J.W.Fox,and R. R.Wagner.1987.Functional andantigenicdomains of the matrix(Ml) proteinofinfluenza A virus. J. Virol.61:239-246.
57. Zebedee,S. L., andR. A. Lamb. 1988. InfluenzaAvirus M2 protein: monoclonal antibody restriction of virus growth and detection of M2 in virions. J. Virol. 62:2762-2772.
58. Zebedee, S.L., and R. A. Lamb. 1989. Growth restriction of influenza A virusbyM2protein antibodyisgenetically linkedto theMl protein.Proc. Natl.Acad. Sci. USA 86:1061-1065. 59. Zebedee, S. L., C. D. Richardson, and R. A. Lamb. 1985.
Characterization of the influenza virus M2integral membrane proteinandexpression attheinfected-cell surface from cloned cDNA.J. Virol. 56:502-511.
60. Zhirnov, 0.P.1990.Solubilizationofmatrix proteinMl/M from virions occurs atdifferentpHfororthomyxo- and paramyxovi-ruses.Virology 176:274-279.
J.VIROL.