0022-538X/89/072941-10$02.00/0
Copyright © 1989, AmericanSociety for Microbiology
Neutralization
Epitopes of the F
Glycoprotein of Respiratory
Syncytial Virus: Effect of Mutation
upon
Fusion Function
JUDY A. BEELERt ANDKATHLEEN VAN WYKE COELINGH*
Laboratoty of Inifectious Diseases, National Institiute of Allergy and Infectious Diseases, Bethesda, Maryland20892 Received 12 January1989/Accepted 23 March 1989
Eighteen neutralizing monoclonal antibodies (MAbs) specific forthe fusionglycoproteinof the A2 strainof respiratory syncytialvirus(RSV)wereusedtoconstructadetailedtopological and operationalmapofepitopes
involved in neutralization and fusion. Competitive bindingassays identified threenonoverlapping antigenic
sites (A, B, and C)andonebridge site (AB). ThirteenMAb-resistant mutants(MARMs)wereselected, andthe
neutralization patterns oftheMAbswitheither MARMsorRSV clinical strains identified a minimumof 16
epitopes. MARMs selected with antibodies to six of the site A and AB epitopes displayed a small-plaque
phenotype,which is consistent withanalteration in abiologically active region ofthe Fmolecule. Analysis of MARMsalsoindicatedthat theseneutralizationepitopesoccupytopographically distinct but conformationally
interdependent regions with unique biologicalandimmunologicalproperties. Antigenic variation in Fepitopes
wasexamined by using 23 clinical isolates(18subgroupA and5 subgroup B) incross-neutralizationassayswith the 18anti-F MAbs. Thisanalysis identifiedconstant,variable, andhypervariable regionsonthe molecule and
indicated thatantigenic variation in the neutralization epitopes ofthe RSV Fglycoprotein is the result ofa
noncumulative genetic heterogeneity. Ofthe 16 epitopes, 8wereconservedon allorall but 1 of 23subgroup
Aorsubgroup B clinical isolates.
Respiratory syncytial virus (RSV) is the most important
cause of severe lower-respiratory-tract illness in young
children worldwide. Thisparamyxovirus causes yearly
epi-demics during which reinfection ofseropositive individuals iscommon. Although RSV is considered tobe amonotypic
virus, two antigenic subgroups have been characterized by cross-neutralization studies (8, 9, 54)with monoclonal anti-bodies (MAbs) (4, 32, 36, 46, 49) and polyclonal sera(50).
Subgroup-specific antigenic differences occur in several of
the RSV proteins, including the envelope glycoproteins, F and G, and the internal proteins N, M, NP, and the 22-kilodalton (M2) protein (2, 19, 20, 30, 32, 41, 46, 52). Ithas been suggested that this antigenic variation is a factor in
allowing reinfection to occur (8, 30, 37) and may be a
challenge to the development of a vaccine that will give
protection against disease from infection with all RSV strains.
Theprototypesubgroup Astrains(A2, Long, and RSS-2) and subgroupB strains (18537andWV4234) notonly differ antigenically but also have considerable nucleotide and amino acid sequence divergence between their envelope
glycoproteins (5, 13, 24a, 26). Despite these differences, postinfection sera from cotton rats contain antibodies that will neutralize heterologous strains in vitro, and
cross-protection against heterologous challenge has been shown followingimmunizationwith either the A2or18537 strain in
vivo (25, 40). This is due, in large part, to the immune
response tothe Fglycoprotein (28, 33, 34),asdemonstrated by the protection against heterologous challenge that was
given to animals immunized with a recombinant vaccinia virusexpressingthe Fglycoprotein ofRSV A2(Vac-F)and challenged with strain 18537 (35). These results, however, cannot accountfor theepidemiologicpatterns of RSV infec-tion within the humanpopulation: subgroupAand Bviruses
* Correspondingauthor.
tPresent address: Division ofVirology, Food and Drug
Admin-istration, Bethesda, MD 20892.
mayeithercocirculate orindependently dominate epidemics (21), and children so exposed appear to have only partial protection against infection, which isgreateragainstviruses of the same subgroup (31). It is likely that a better
under-standingof the complex epidemiology of RSV willbe facil-itatedby adetailedanalysisof the neutralization andfusion
domainsof the RSV envelopeglycoproteins.
Weinitiated thisanalysis by isolatingandstudyingalarge
panel ofneutralizinganti-F MAbsto subgroup Astrains of RSV in order to mapthe antigenic sites and neutralization
epitopes within these sites and to study the functional organization of the RSV Fglycoprotein. Competitive bind-ing assaysof18 MAbs delineated three antigenic sites, and neutralization assays of the MAbs with MAb-resistant
mu-tants (MARMs) and clinical isolates detected 16 epitopes,
some of which appear to be operationally interdependent although topographically distinct. Six of the MARMs
se-lected withfusion-inhibitingMAbsdisplayedasmall-plaque
phenotype, indicating that certain neutralization epitopes residewithindomainswhich affectthe fusion function ofthe molecule. These particular epitopes were shownto be
con-served among all subgroup A and B RSV clinical strains examined. Wealsosoughttoobtainan extensivemapof the
neutralization and fusionepitopes (i.e., epitopesrecognized by MAbs which inhibit fusion between infected cells) by usingbiologicalassayswhich couldbe usedtorelate biolog-ical functions of this proteinto specific antigenic sites.
MATERIALS AND METHODS
Cells and virus. Cellsused in these studies(HEp-2, Madin-Darby bovine kidney [MDBK], LLC-MK2, and BS-C-1)
were maintained in Eagle modified minimum essential
me-dium supplemented with 10%fetal bovineserum. Semicon-fluent cell monolayers were infected with RSV at a multi-plicity of infection of 0.1 in Eagle modified minimum essential medium-1% fetal bovineserum. Extracellular
vir-ions were harvested after extensive cytopathologywas evi-dent (48 to 72 h postinoculation) and stabilized by the
2941
on November 10, 2019 by guest
http://jvi.asm.org/
2942 BEELER AND COELINGH
addition of 100 mM MgSO4 (16). Viruses used for immuni-zation of mice or for radioimmunoassays were purified by sedimentation through 30% sucrose onto acushion of60% sucrose. Virus stocks used fortheselection of MARMs were concentrated by centrifugation to 108 50% tissue culture infectious doses
(TCID50)
perml.Theprototype human RSV strains (A2, Long, and 18537) wereplaque purified three times in HEp-2 cellmonolayers. Other human RSV strains, generously supplied by R. M. Hendry, Food and DrugAdministration, Washington, D.C., and H. W. Kim, Children's Hospital National Medical Center, Washington, D.C., were not plaque purified and werepassaged a limited number of timesin HEp-2 cells; they are identified according togeographiclocation, isolate num-ber, and year ofisolation. The bovine RSV strain was a gift from M. H. Smith,University of Iowa, Ames, Iowa, and was propagated and assayed in MDBK cellmonolayers.
Vaccinia virus recombinants expressing RSV F (Vac-F), RSV G (Vac-G), RSV N (Vac-N) or Escherichia coli I-galactosidase (Vac-,-gal) were agift from R. A. Olmsted, Georgetown University, Rockville, Md. Vaccinia virus re-combinants were propagated inBS-C-1 cell monolayers(35),
andextracellular virions wereused for mouseimmunization
and for infection of cellmonolayers whichserved asantigens in anenzyme-linked immunosorbent assay (ELISA).
Production of hybridoma cell lines. Female BALB/c mice were immunized by primary intranasal infection with 105
TCID50 of the A2 strain of RSV,and 3weeks later they were inoculated intraperitoneally with 108 PFU of the Vac-F recombinant. Four weeks later, they received an
intrave-nous injection with 104 TCID50 ofpurified RSV. Four days after thefinal immunization, spleniclymphocytes werefused
withthe NS-1 murine myeloma celllineby standard proce-dures (27). Hybridoma supernatantsproducingRSV-specific
antibodies were identified by anELISAof RSV-infectedcell monolayers. Allpositive hybridomas wereimmediately
sub-cloned twice by thelimiting-dilution method andwere used for the production ofascites fluid.
Seven additional hybridoma cell lines used in this study
werekindly provided by L. Anderson, Centers forDisease Control, Atlanta, Ga., E. Norrby, Karolinska Institute,
Stockholm, Sweden, and B.Fernie,GeorgetownUniversity;
Rockville, Md.; the methods used for theirproduction and characterization have been published(MAb 151 [31]; MAbs 43-1 and 13-1 [15]; MAbs 1436C, 1302A, 1308F, and 1331H [3]). The immunoglobulin subclass of each MAb was deter-mined by using the Boehringer-Mannheim Immunoglobulin
Subclass Determination Kit. Polyclonal antisera used in these studies included cotton rat post-RSV-infection serum, pooled adult human serum, and murine anti-RSV ascites
fluid. Murine ascites fluids were produced by infection with eitherRSV or Vac-F, followed by intraperitonealinjectionof pristane and Sarcoma 180 cells (47).
Serologic assays. An indirect ELISA was performed with RSV-infected LLC-MK2 cell monolayers or vaccinia virus recombinant-infected BS-C-1 cells. Cell monolayers in 96-well plates were infected at a multiplicity of infection of 1. After development of extensive cytopathology, the cell monolayers were desiccated at 37°C and stored at 4°C. Plates were washed three times with phosphate-buffered saline-0.1% Tween 20, blocked with 20% fetal bovine serum, and washed again prior to addition of the test MAbs. After an overnight incubation, peroxidase-conjugated goat-anti-mouse immunoglobulin G (Cappel Laboratories) was added
for
4h, and thetest was developedwithasubstratesolutioncontainingthe chromophore
2,2'-azinobis(3-ethylbenzthiaz-olinesulfonic acid) (ABTS).
Neutralization assays were performed as follows. Serial
twofolddilutions of heat-inactivated (56°C for 30min) ascites fluid of serum were mixed with 100
TCID50
ofRSV in the presence of5% guinea pig complement. After 60 min, the virus-antibody mixtures were transferred to monolayers of HEp-2 cells in 96-well plates; 6 days later, the monolayers were examined microscopically for cytopathology. Micro-scopic observations were confirmed by staining with 5% glutaraldehyde-1% crystal violet. Neutralization titers were expressed as the reciprocal of the highestantibody dilution which inhibited more than95% of theviralcytopathic effect present in thecontrol wells.Fusion inhibition (FI) assays were performedasdescribed previously (32) to identify MAbs that inhibit syncytium formation. Briefly, confluent monolayers of HEp-2 cells in 96-well plates were infected with 100
TCID50
of virus at 37°C. After 60 min the inoculum was removed, and the monolayers were washed for 2 h with Eagle modified mini-mum essential medium to remove unabsorbed virus. Serial twofold dilutions of MAbs or hyperimmune sera werethen added to each well. Six days later monolayerswere exam-ined for cytopathic effect, and antibody titers were deter-mined in the same manner as described for neutralization assays. Competitive solid-phase radioimmunoassays were performed aspreviously described (12). Briefly, MAbswere intrinsically labeled with [35S]methionine. Unlabeled ammo-nium sulfate-precipitated MAbs hadafinal concentrationof 1 mg of protein per ml and showed similar ELISA titers (<10-fold difference) when tested on the RSV A2-infected LLC-MK2 cells prior to use in the RIA (data not shown). Purified RSV virions were adsorbed to polyvinyl chloride plates at aconcentration of 0.15to1.4 ,ugofproteinperwell. Apreviously determined saturating amountof labeled MAb was mixed with increasing concentrations of unlabeled am-monium sulfate-precipitated MAbs and added to the wells. The amount of bound labeled MAb was measured for each well. Competitive binding titers were expressed as the reciprocalof the dilution ofcompetingunlabeled MAb which produced 50% inhibition in binding of the labeled MAb.Selection of MARMs. Themethod for selection of MARMs has been described previously for influenza A virus (18).
Briefly, undiluted ascites fluids were incubated with
undi-luted or decimaldilutionsof concentrated of RSV A2at37°C
for 1 h. Virus-antibody mixtures were adsorbed to HEp-2
cellsfor2hat37°Cand thenoverlaidwith0.8%agarose L-15 medium containing the selecting MAbat adilution of 1:100. After 6 days, plaques were visualized with neutral red. The frequency at which MARMs occurred was estimated as
describedpreviously (18, 44). MARMswere plaquepurified twice, amplified in the presence of the selecting MAb, and tested again in neutralization assays against the complete
panelof anti-F MAbs. MARMs areidentifiedbytheprefix V and by the MAb used for their selection (e.g., V1269).
RESULTS
Specificity of MAbs for RSV Fglycoprotein. To produce a large number ofneutralizing MAbs specific for the RSV F glycoprotein, we immunized mice by sequential infection
with RSV A2 and Vac-Frecombinant virus. This immuniza-tion protocol stimulated high levels ofcirculating antibodies
which reacted in the ELISA with F glycoprotein expressed by Vac-F-infected BS-C-1 cells. Following initial infection
with RSV, mice developed anti-F titers of ca. 1:640, which J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Assignment of RSV glycoprotein F-specific monoclonalantibodies toantigenic sitesby competitiveradioimmunoassay Anti-
[35Slmethi-
Reciprocaldilution ofindicattedunlabeled MAbwhichproduced50% inhibition of labeledMAb"
onine-genic labeled
site MAb 1436C 1153 1142 151 1200 1214 1237 1129 1121 1107 13-1 43-1 1112 1269 1243 1331H 1308F 1301A
A 1436C 64 256 64 256 64 16 64 64 256 64
1153 4,096 1,024 256 1,024 256 256 256 1,024 1,024 1,024 1142 16.384 4,096 1,024 16,384 1,024 4,096 4,096 4,096 1,024 4,096 151 4,096 1,024 256 4,096 256 64 64 256 1,024 1,024 1200 256 1,024 256 256 256 256 256 256 256 256 1214 1,024 1,024 256 4,096 256 256 256 1,024 1,024 1,024 1237 1,024 1,024 1,024 256 1,024 256 256 1,024 1,024 1,024 1129 4,096 1,024 256 1,024 1,024 256 1,024 256 4,096 256 1121 256 256 256 1,024 256 64 256 256 1,024 256 AB 1107 4,096 1,024 1,024 256 4,096 64 1,024 1,024 256
B 13-1 4,096 1,024 1,024 16,384 1,024
43-1 4,096 4,096 4,096 4,096 4,096
1112 64 256 64 256 64
1269 256 256 64 256 256
C 1243 256 256 1,024 1,024
1331H 4,096 16,384 65,536 16.384
1308F 64 256 256 64
1302A 16,384 16,384 16,384 4,096
Each MAb was radioactively labeled, added to dilutions of unlabeled MAbcompetitors. and testedinsolid-phalseradioimmunoassays. Blanks indicate no
competitionabove that observed with control ascites.
increased to a titer greater than 1:106 following subsequent infection with the Vac-F recombinant. Fusion of splenic lymphocytes from animals immunized in this way yielded 30 hybridoma cell lines which produced antibodies to RSV proteins (data not shown). Seventeen of the MAbs reacted specifically in the ELISA with Vac-F-infected BS-C-1 cell monolayers. These MAbs did not react with uninfected monolayers or with monolayers infected with G, Vac-N, or
Vac-3-gal
and are therefore designated RSV-anti-F MAbs. The specificity of these MAbs for the viral F glyco-protein was confirmed by radioimmunoprecipitation assays (data not shown).Hybridoma cell lines were grown as ascites tumors, and the neutralizing antibody titer of each was determined with the RSV A2 strain. Of the 17 F-specific MAbs, 11 were able to neutralize virus infectivity. Seven additional neutralizing anti-F MAbs kindly supplied by three other laboratories were also used in these studies to complete the panel of 18 RSV-anti-F neutralizing MAbs.
Topographical analysis of RSV-F antigenic sites. To deter-mine the number of nonoverlapping antigenic sites detect-able with the panel of 18 neutralizing MAbs, we performed
competitive-binding solid-phase radioimmunoassay. Each of the 18 MAbs was intrinsically labeled with [35S]methionine, and increasing amounts of unlabeled MAb were used for competition. CompetitionbetweentwoMAbsindicated that they boundtoepitopesthatwereinclose proximity,i.e.,the same site, whereas lack of competition indicated that they
bound to spatially separated sites. The level ofcompetition
between labeledandunlabeledMAbs isindicated inTable 1. The endpoints for 50% competition by agiven MAb varied over awide range ofantibody dilutions (e.g., theendpointat which MAb 1214 inhibits binding of other site A MAbs ranges from 1/16 to 1/4,096), presumably reflecting differ-ences inantibodyaffinitiesand proximityof epitopes. These differences areunlikelytobe duetoquantitative differences in the unlabeled MAbs, because they had similar ELISA titers when tested individually. As expected, in every case
thebindingof labeled MAb was inhibitedbythehomologous competitor. Ten MAbs exhibiting reciprocal competitions were assigned to antigenic site A, four MAbs (13-1, 43-1, 1112, and 1269) were assigned to antigenic site B, andfour MAbs (1243, 1308F, 1331H, and 1302A) were assigned toa region ofthe F glycoprotein designated site C. MAb 1107 competedreciprocally withMAbsfromsiteAand nonrecip-rocally with each of the MAbs from site B, indicating that MAb 1107bound to anepitope that mayact asabridge (site AB) between thesetwo sites. Five nonreciprocal reactions aredefinedbythepairwisecompetitions between MAb 1107 (site AB) and MAbs1214(site A), 13-1, 43-1, 1112, and 1269 (site B). Such results may also be duetosubtledifferencesin antibody affinities. Alternatively, bindingof one MAb may cause conformational changes inotherepitopes, which pre-ventantibody binding of another MAb.
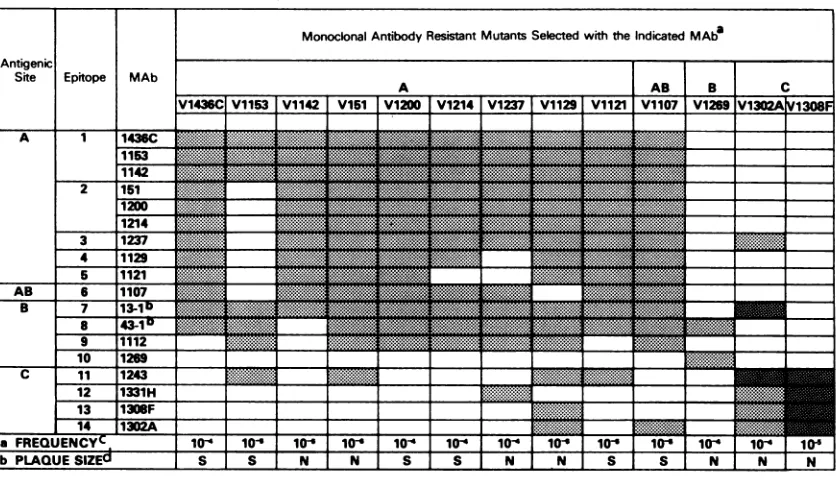
Selection of MARMs and construction of an operational epitope map. Analysis ofreactivity patterns of the panel of MAbs with MARMs identified 14 epitopes located within antigenicsitesA, B, AB, andC. MARMs were selectedfrom aconcentrated preparation ofplaque-purifiedRSV (A2)and were shown to be true mutants by their resistance to neutralization by the selecting antibody. Cross-neutraliza-tion studies were done by using 13 MARMs and the com-plete panel of18RSV-anti-F MAbs(Table2). These neutral-ization reactivity patterns were used to construct an operational mapof the F-glycoprotein epitopes.
Aninterestingfeature of thisanalysisof theFglycoprotein was that MAbs frequently did not react with MARMs selected by using MAbsassigned todifferent antigenic sites by competitive-binding assays. This phenomenon was ob-servedfor MAbs mappingtoall of the Fantigenic sites and may reflect the sensitivity of neutralizing MAbs to alter-ations in areas of the protein other than the actual epitope. For example, the nine MAbs defining antigenic site A by competitive-bindingassays(Table 1) clearly recognizedfour epitopes (Al, A2, A4, and A5), as indicated by their reac-tivity patterns with MARMS selected by usingMAbs
on November 10, 2019 by guest
http://jvi.asm.org/
2944 BEELER AND COELINGH
TABLE 2. Determination of thenumber ofepitopeswithinantigenic sitesby neutralization ofMARMs
Antigenic
Site Epitope {MA
MonoclonalAntibody Resistant Mutants Selected with the Indicated MAba
A AB B C
V1436C V1153 V1142 V151 V1200 V1214
I
V1237I V11291
V1121 V1107 V1269 V1302A|V1308FI~~I~
A 1 1430C
1153 1142
2 151
1200 1214
3 1237
4 1129
5 1121
AB 6 1107
B 7
9 10 11 12 13 14 a FREQUENCY .
1112
1243
1308
1302 b-
b7-H F A
b PLAQUESIZEU
'Symbols:
HII.
neutralization titer c8t):
LIII neutralization titer>80.
h MAbs 43-1and13-1 wereproducedfollowingimmunization of mice with theLong
str-ain
of RSVanddonotneutralizethe A2strain,whichwasusedasthe parentforthe MARMs.' ThefrequencyatwhichMARMSwer-e selectedwascalculatedaccordingtothefollowingformula: PFUin the presence of RS MAb/PFU in the absence of
RS MAb.
"Plaque size:N, normalplaquesize(2.5mm);
S.
smallplaquesize(0.5mm).ping topographically to site A. However, the lack of reac-tivity of MAb 1237 with variant 1302A(selectedwithasite C MAb) indicated that it recognizes a fifth epitope (A3) and that its neutralizing activity is influenced by alterations in site C. These results support the view that in some cases, topographically distinct antigenic sites are operationally or
conformationally interdependent.
In addition to their altered antigenic properties,
approxi-mately half of the MARMs analyzed also exhibited an altered plaque morphology (Table 2). Six of the site A and AB MARMsproduced pinpointplaques under agarose over-lay, but this phenotype didnotstrictlycorrelate withspecific
siteAepitopes. For example, onlytwoof the three MARMs selected with MAbs recognizing epitope Al exhibited the small-plaque phenotype. Whether these phenotypic changes reflect differences in the nature or location of mutations in theMARM Fproteins is currently unknown.
Antigenic site B is a complex site which was clearly
distinguishedfrom siteAby MAbs 13-1, 43-1, 1112,and 1269 incompetitive-binding assays (Table 1). These results were confirmed bytheability ofMAb 1269to neutralize all site A MARMs and its inability to neutralize a site B MARM (V1269).Inseveral cases, site BMAbsdid not neutralize site AMARMs. This lack of neutralization can be explained for MAbs13-1and43-1: these MAbswereproduced to the Long strain ofRSV and donot neutralize either the A2 strain or mostofoursiteAMARMs (which are mutants selected from the A2strain). The neutralizing activity ofMAbs 13-1, 43-1, and 1112isclearly influenced by mutations within both sites Aand B, asevidenced by theirreactivity patterns with site A and B MARMs. Therefore, the results of competitive-binding assays (Table 1), in combination with analysis of mutants(Table 2), indicated that MAbs13-1, 43-1, 1112. and
1269recognized four epitopes in siteB and thatmutations in
site A can, in some cases, influence the neutralizing activity of antibodies directed to site B epitopes. For example, V1142 has a site A mutation but is now neutralizedby MAb 43-1 (neutralization tier 1/2,560).
MAbs 1243, 1308F, 1331H, and 1302A were unable to neutralize MARMs V1302A and V1308F, confirming our assignment of these MAbs to antigenic site C based on competitive-binding data(Table 1). However, as described above for site B MAbs, the site C MAbs showed unique
reactivity patterns withMARMs selected with siteA MAbs (V1153, V151, V1237, V1129, and V1121), indicating that they recognized four different site C epitopes which are influenced by changes within site A.
The mutation rate in the RSV-F epitopes was estimated from the frequency of isolation of MARMs (Table 2). MARMs wereselectedat afrequencyof10-4to10-6,which is similarto results withotherRNAviruses (44).
All MARMs wereneutralized by hyperimmuneantiserum (datanotshown),asdescribed for influenza virus(53).Itwas noted, however, that MARMs with alterationsaffecting the bindingof MAbs in all threeantigenic siteswereneutralized lessefficiently than MARMs with alterations affecting only one site.
Antigenicvariation among RSV clinical isolates from 1956 to 1986. In an attempt to identify additionalepitopes within the antigenic sites and in order to determine the degree of antigenicvariationthat occursnaturallyamongRSV strains,
we tested subgroup A and B viruses recovered during the past 29 yearsincross-neutralizationassayswith thepanel of anti-F-MAbs and with pooled human adult sera. The neu-tralizationtest wasused todefine thispatternbecause it isan important functional assay that might have more biological relevance than binding assays have. There were nine reac-tivity patternsforthe 18subgroupA viruses tested and five
J. VIROL.
fb
on November 10, 2019 by guest
http://jvi.asm.org/
RESPIRATORY SYNCYTIAL VIRUS F GLYCOPROTEIN 2945 -~-P,- --~1 >-01a: O00- ON< No ,.0, 0~1-, k
Ot NL
0L
-0, 1- ./h
0 0000 O0000 0
N ONO OJ C CD CD
0D 000 So0 0
0 0000 00,
C'--Bos ACO-_IA
vJ ON00'. k) ' ON0 C
0 0000 0000o
4.- tJk.3 t.IQ
I-
k.3k.-
I'J --7~
ONONONO ON ON ON ON. ON1
C71 k.3- C 00
00 ON CD k) CD-.)D~ . C)
000000000S
ON C JJON Jl. O(JO
00 O ON ON ON 00 ON
00o0000000s
000000000
ON ON t. k. k ON ) k. 00
000000000
_ _ _ u UI ~t-i (-A ti _- t-i ti uA 91 t1 _- t" >
t'
00 0000t') t-) IC'.tIa, ) ON 00OC alC-. . .00OC'ON )
000 E0D00 0 0 0 0 0 0 0 0 0 0
" i(-A " (.AIA _
o ININIo Io IoNo I k
0-
0000Lt
0L ]00 _P,ON00t-.J 1 00
0)C000)C LBBJ00 0
(ww w r'wJc w
ON ONON OC.)C) ON ) C-)
"-A~t(-A-.--'-" )~A
~-00 ON - . . . 00 CN )
00 00 00 0 00
F- f- 4- W1t
w ON A to O NL"CUN--
II--t') 4'-1 00ON I" 4)1 00 "C.I v00ONO t)Ct)C1
0D 0= C 0°°°
CDOCDCD)ICD IAAI
J w wC
0 0000 000
0
0~--0
L-,
J00000 00 A-1 tjC7,>
C,*IC )C C'-) ()C
0'. 000 -00I-000004-C-.
~~/, t-.)C'J
t-O oCo) NO
0 00 00j00000
wI.- w (.AJ w
OC') (111 0 " -P CIS oN "-CJON
_(OItC)'C.eN =C'-.)
0o [Sv z czC E2I 00s
00 -)C') C')
00 C'-.)1 0 0000 0000 0 0000 _ C) -= ~CO
00 00 0
C'-.C- C'D)
_ 0n
|A
A A A
I m
C'0
-A -P0D
00 0oc= x x) x
-- (AC'.) -- (.A b- t-ol -- ~- ~. t-
,-00ON 0 0oCN ON00
°) °1 °1° °I ° °°=
A A A A A A A A A t-) t-) t-) t-) I" t-) tI-. t-)
01 1
ON A A A\1 ONC'-.)I ON IC'.) C'. C-) C'-.- 00 ON
0t4z,00C
00o o o
uc > C O. CO -1 0r tz CD > 00 0N >OD r 0 00
_
-o o. P, 0 .P.>
C4C
00 00_ w ~~~~~~CD
00
CD
CDP00 0
00
00
Uu
~~~~~~~O
00
or
W0 |0,==
CD C
VOL.63, 1989
fr
3 _. 0, 0, _. _. CO 0E CO 0 _. -O_. 0 -o _. o, -o CO 0 CO 0. 0 _.0 00 C -1 E. r-V) E. _. CA.
co CL CK CD CA CD CD D In _.' E CD U7 _. 2. CD C. _. _2. CD 0CD CO 0CD C XCO C 0,C CD~ 0-F C7 Q0 _00 C (A CD Ct' C CD CD CD (A CD 0 la 0 5 3 CD _. -1 El. CD CD C. _. -a CDn tz >
tz
(-n
on November 10, 2019 by guest
http://jvi.asm.org/
2946 BEELER AND COELINGH
TABLE 4. Fl andneutralization activities of anti-FMAbsagainsttwosubgroupAstrainsof RSV"
Antigenic Epitope MAb_ImmunizingImmunoglobulin Aus/A2/6__SL/863/84_
sitesiteEpitope MAb Immuniingstrain smunolbulinAsass"subclass N titer Fltiter N titer FlL8I4titer N/FI
A 1 1436C A2 1K 3,200 1,280 5,120 2,560 2
1153 A2 1K 520 320 2,560 320 8
1142 A2 1K 600 320 1,280 320 4
2a 151 A2 1K 2,400 1,280 5,120 2,560 2
2b 1200 A2 2BK 1,500 1,280 5,120 640 8
2c 1214 A2 2BK 1,700 640 2,560 640 8
3 1237 A2 1K 700 640 1,280 320 4
4 1129 A2 1K 650 320 1,280 320 4
5 1121 A2 1K 640 640 640 320 2
AB 6 1107 A2 1K 1,200 320 5,120 160 32
B 7 13-1 Long 1K <20 <20 2,560 <20 >128
8 43-1 Long 1K 20 <20 5,120 <20 >256
9 1112 A2 1K 200 <20 5,120 <20 >256
10 1269 A2 1K 600' 320" <20 <20 1.87c
C 11 1243 A2 2AK 120 40 5,120 40 128
12 1331H A2 2AK 1,300 800 5,120 80 64
13 1308F A2 1K 1,050 640 5,120 320 16
14 1302A A2 1K 900 640 5,120 64 64
"Anti-Gcotton rat seraneutralizedAus/A2/60 withN=2,560andFl<20;itneutralizedSL/863/84 with Fl <20. ClinicalisolateWash/53/1960 neutralization titers,Fltiters,and ratios are as follows: MAb 43-1(2,560, <20, >256),MAb 13-1(80, <20,>4),MAb 1112(5,120, <20, >256),MAb 1269(5,120, 2,560, 2).
bEach value represents thereciprocalof the neutralization(N)titer or
Ft
titer of the isolatebythe MAb.Theratio of neutralization toFtactivityfor each was determinedby usingthe titer forSL/863/84,except thatAUS/A2/60wasused to determine the ratio for MAb 1269.
patternsfor the 5subgroupBviruses(Table3).The neutral-ization patterns confirmed theoperationalmapderived from MARM analysis (Table 2) and indicated that MAbs 151,
1200, and 1214 recognized differences inepitopes (Table 3,
A2a,A2b,andA2c,respectively). Thus,the total number of neutralization epitopes detectable by the MAb panel is 16. The distribution of neutralization epitopes within strains indicated that constant,variable, and hypervariable regions
existed onthe Fglycoprotein.
Of the detectable F neutralization
epitopes,
threewithin siteC(C12, C13,andC14)werestrictlyconserved among all RSV strains examined. In addition, there appears to be ahighdegree ofantigenic stability in site AandABepitopes
amongsubgroupAviruses: noalterations in siteAepitopes
were detected. On the other hand, subgroup B viruses showedconsiderable antigenic variation in site Aepitopes: epitopes
Al,
A2a, A2b,A4, and A5wereunaltered inonly60 to80%ofthe subtype Bstrainsanalyzed(Table3, footnotec). EpitopesA2candA3were not detectableby neutraliza-tion of any of the B strains, indicating that these epitopes
weresubgroup A-specific neutralization epitopes. One sub-group B clinical isolate(W/171/1985) was notneutralizedby anyof the MAbsreacting with site A or ABepitopes.
In contrast to the antigenic stability in site A andsite C
epitopes among subgroup A viruses, site B appears to be hypervariable among subgroup A strains. For example, MAbs to site B epitopes neutralized only 33 to 61% of the
subgroup A viruses. In contrast, the site B epitopes were
relatively conserved among subgroup B viruses (Table 3, footnotec).
One bovine RSV isolatewas evaluated in asimilar man-ner.This bovine strain, likesubgroup Bviruses, lacked site A epitopes identified by MAbs 1200, 1214, and 1237. In
addition, it had alterations in all four epitopes associated withantigenicsiteB.This indicatesthat at least two
immun-dominant
regions
of the Fglycoprotein of bovine RSV aredistinctlydifferent fromthecorrespondingregionsof human RSV strains.
The results of thisanalysis indicated thatantigenic varia-tion occurs both between andwithin the human RSV sub-groups. However, of the 16 neutralization epitopes
identi-fied, 11 were present on each subgroup A virus tested, indicating the high degree of conservation of these F
epitopeswithin typeAstrains.Subgroup-A-specific neutral-ization determinants were found in antigenic site A, and
hypervariable epitopes clustered in antigenic site B. There was no correlation between the reactivity patterns of the clinical isolates and their year ofisolation, indicatingthat the
antigenicvariation detected doesnotrepresent the progres-sive accumulation ofantigenicalterations with time.Rather,
the observed variation appearstobe due togenetic hetero-geneity among RSV strains that infect humans.
Biological activity of RSV anti-F-MAbs. (i) Fusion inhibi-tion.Previousanalysisof otherparamyxovirusglycoproteins
indicated that distinct functional activities can often be
assigned to different antigenic regions of the glycoprotein,
e.g., Newcastle diseasevirus, HN (23); Sendai virus, HN
(38); parainfluenza virus type 3, HN, (11); and Newcastle diseasevirus, F (1). Asimilaranalysis of the antigenic and functionalorganization of the RSVfusionglycoprotein was
performedby comparing the ability of theMAbs to neutral-izeinfectivityandpreventsyncytium formation. Expression of neutralization andFl titers(Table4)as aratio(N/Fl)was usedtocompare theefficiency of each MAb in neutralization and/or Fl and differentiate functional domains on the mole-cule. Since several siteB MAbs exhibited low neutralizing
activity against the A2 strain, the assays were also per-formed with a subgroup A virus (SL/863/84), which was neutralizedto high titerby 17of 18 MAbs (Table 3). These MAbs had similar titersofneutralizingactivity but differed
substantially in their capacity to prevent the formation of
syncytia (Table 4).
J.VlIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 5. Comparison of neutralizing and binding activities of anti-F MAbs to RSV clinical isolates
Antigenic Neutralizing/binding activity of indicated clinical RSV
strain'
site
Epitope
MAbAUS/A2/61 W/18537/62 W/3327166 W/3199/66 W/343/67
A 2C 1214 +l+ -l- +l+ +l+ +l+
AB 6 1107 +/+ +l+ +l+ +l+ +l+
B 9 1112 +/+ +l+ -l- -l+ -l+
10 1269 +l+ -l+ +l+ +l+ +l+
C 11 1243 +l+ +l+ -l- -l+ +l+
aNeutralizing MAbswere testedwiththe indicated clinical strains in ELISAbindingand neutralizationtests,and the results of both tests arereported.
Each of the MAbs defining site A epitopes and one MAb
defining a site B epitope (B10) inhibited fusion very
effi-ciently, asindicated by the lowN/Fl titer ratios (ratios of 2 to8). Themajority of site B MAbs did not exhibit Fl activity. MAbs toepitopes in site C had intermediate ratios,
suggest-ing that perturbation of this site can also affect fusion
function. Thus, it appears that two of the topographically
distinct antigenic domains of the F protein correspond to
regions that are at or near the site of most important
biological activity. Furthermore antigenic sites A and C are
relatively wellconserved (Table 3), whereas site B is more variable and contains only one epitope that might be at or nearthefusion site. Also, it should be emphasized that 6 of the 10 MARMsselected with site A and site AB MAbs had the small-plaque phenotype.
(ii) Neutralization and binding of RSV Anti-F MAbs. The capacity of the RSVF-MAbs to neutralize subgroupAand
subgroup B viruses and to bind to the F proteins of these
viruseswascompared (Table 5). In severalinstances, MAbs
whichdid notneutralize agivenvirus also did not bind to the Fproteinof that particular strain. For example, the lack of
binding of MAb 1214 with the 18537 virus explained the
failure ofthis MAb to neutralize this strain. On the other
hand,antibody binding didnotalways correlatedirectlywith theability to neutralize infectivity. For example, analysis of
five other subgroup A clinical isolates yielded additional
examples ofadequate antibody binding without
neutraliza-tion.Thereactivity ofMAb 1112 withisolatesW/3199/66and
W/343/67,
MAb 1269with W/18537/62, and MAb 1243 withW/3199/66 illustrate this pattern (Table 5). These results
indicate that dissociation ofimmunological and biological
activity can occur for F epitopes and probably reflects the morestringent requirements for antibody-mediated
neutral-izationcompared with antibodybinding.
DISCUSSION
Thegoal ofthis project was to construct detailed
opera-tional, topological,andfunctionalmapsof neutralization and fusion epitopes on the RSV Fglycoprotein. Since
immuni-zation of micewithpurified RSVproteins appeared toyield
few neutralizing MAbs, we devised a novel immunization scheduleinvolving sequentialinfection with RSV and Vac-F
recombinants. This immunization stimulatedhigh levels of F
antibody in these animals and resulted in 11 hybridomas which secreted F-specificneutralizing MAbs. Thesuccessof this protocol may be due to both the nature of antigen presentationand thedegreeof stimulusobtainedbyinfecting
mice with two different viruses in a specific order. First, immunization byvirus infectionallowed the Fglycoprotein
to be delivered in an undenatured form.
Second,
mucosalinfectionwith RSV primedtheanimalforan Fresponsebut didnotrestrict the replicationof Vac-F.
Priming
bynatural infection with RSV also appeared to be important. Anti-Ftiters obtained after immunization with Vac-F alone or induced by infection with Vac-F followed by RSV were much lower thanwe have reported here (data not shown).
Competitive radioimmunoassay with these MAbs identi-fied threenonoverlapping antigenic sites involved in neutral-ization. We also observed a potential bridge site (AB), suggesting that sites A and Bareincloseproximityonthe F molecule. These results are in good agreement with those obtained for RSV and otherparamyxovirus fusion proteins,
which have delineated three or four nonoverlapping anti-genic regions (4, 10, 14, 15, 42, 50). Furthermore, neutrali-zation tests involving MARMs and RSV clinical isolates proved to be the most sensitive method to detect subtle
antigenicdifferences among these viruses;theyidentified 16
epitopes. Seven of theseepitopes arelocated within site A, fourarelocated within site B,fourarelocated within siteC, and one is located in the bridge site AB. One of the
neutralization epitopes of the RSV F-glycoprotein has
al-readybeenlocalizedtoresidues 221 to 236 on the F1subunit by Trudel et al. (48). Nucleotide sequence analysis ofour MARMs will give additional information about the specific
amino acids involvedin 13 of the neutralization epitopesof theRSV Fglycoprotein.
This study also yielded additional insights into the
behav-ior of F neutralization epitopes in response to mutations
within specific antigenic sites. We present evidence that mutations distantfromtheantigenicsite caneffectresistance toneutralizationorenhancement ofneutralization,
presum-ably mediated by conformational changes. MARMs with
mutations that abolish reactivity with site A MAbs also increased the reactivity of certain site B epitopes or abol-ished neutralization mediated by a site C epitope. One MARM, V1107, sustained a mutation in the bridge epitope AB, and,as aresult, neutralizationat adistantepitope,C14,
wasabolished. Similarly,onesite C MARM, V1302A, failed tobeneutralized bythe MAbdirectedagainst antigenicsite Aepitope 3 and wasalso neutralized by site B MAbs 13-1 and 43-1attiters of1/1,280and 1/5,120, respectively. These resultsindicate that localchanges in theFglycoproteincan result in profound changes in the structure of epitopes
located on more distantantigenic regions.
This is also the first report of MAb-selected
small-plaque
variants of RSV. These six viruses implicate four site A
epitopes and the bridge site, AB, as important functional
regionsontheFglycoproteinthatmayregulatethefusion of infected cells. Specific mutations within theF glycoprotein
of other paramyxoviruses have been associated with alter-ations in plaque size. Indeed, a MAb-selected
small-plaque
variantof Sendai virus has anamino acidchange in Sendai virus Fl residue 407 (Pro->Gln), and it has been
proposed
that this epitope is part of the fusion-active site
(39).
Pro-tease-resistantmutantsof Sendaivirusareanotherexample
of small-plaque variants. It has been determined that the
on November 10, 2019 by guest
http://jvi.asm.org/
2948 BEELER AND COELINGH
alteration in thebiological properties of these viruses occurs as a result of specific amino acid changes at or near the cleavage site (22, 24). It is anticipated that our small-plaque-phenotype RSV MARMs will likewise have specific amino acid changes in their F glycoproteins. It is unlikely,
although still possible, that these phenotypic changes are the result ofmodifications in other envelope glycoproteins or the matrix protein as described formumps virus, measles virus, and bovine parainfluenza virus type 3 (7, 29, 43). The mechanism by which these mutations affect plaque size remains to be studied. One can postulate several causes, including aquantitative decrease in the amount of F glyco-proteinexpressed at the surface of infected cells. The overall stability ofthe F glycoprotein may be altered as a result of changes in charge or hydrophobicity, and this variant F molecule might be less likely to interact with adjacent cell membranes or have decreased activity during syncytium formation.
We were able to delineate antigenic sites and epitopes involved in syncytium formation by fusioninhibition assays. Thus, two of theantigenically stable neutralization sites on the F glycoprotein (sites A and C) correspond to protein domains important for viral fusion activity. Athird neutral-ization site (site B) has minimal involvement in syncytium formation, which may account for the high degree of varia-tion tolerated in this domain. The fact that site A MAbs inhibit syncytium formation more efficiently than site C MAbs and most site B MAbs do has no effect on syncytium formation may reflect differences in the proximity of these three antigenic sites to the fusion-active site(s). Thus, anti-genic site A may be at or near afusion-active site, because it appears to have an important effect on plaque size and is involved in syncytium formation. In addition, we have been able to characterize both MARMs and a clinical isolate which escaped neutralization by all of the MAbs directed to site A or site C epitopes. These viruses form syncytia in vitro, suggesting that major antigenic changes in the F glycoprotein can be tolerated without loss of the fusion-active site.
Anotherinteresting structural property of F-glycoprotein epitopes became apparent when we compared the reactivity patterns of MAbs in neutralization and binding assays. We discovered that there is not necessarily a direct correlation between these two activities. As was expected, absence of reactivity in ELISA did indicate that the epitope for neutral-ization was absent; however, there were several cases, particularly for site B epitopes, that demonstrated dissocia-tion in biological and immunological reactivities, i.e., MAbs could bind but no longer neutralize. Similar findings have been noted in the course ofdifferentiating strains of Sindbis virus (45), yellow fever virus (6), rabies virus (17), and human parainfluenza virus type 3 (10). Currently, we lack a satisfactory explanation for this common but puzzling ob-servation. It may imply that sequence changes in these epitopes result in escape from neutralization but that the overall conformation has not deviated from the wild type sufficiently to preclude binding. This may also be an indica-tion that major conformational changes in these epitopes would not be tolerated, because they contribute to the stability of biologically functional regions. Dramatic changes would produce lethalmutations and may explain, in part, our failure to select MARMs in three of four site B epitopes despite repeated attempts. Furthermore, it also indicates that the use of only binding assays may lead to an underes-timate of the true epidemiological potential of certain RSV strains.
Thus, the combination of MARMs and biological assays allowed us not only to map F neutralization epitopes more
thoroughly, but alsotocharacterizeimportantstructural and functional relationships. Comparison of the F gene nucleo-tide sequences of the RSV MARMs described here with thoseofhumanparainfluenzatype3 fusionproteinMARMs (10) will identify specific areas of paramyxovirus F
glyco-proteins critical for neutralization and fusion inhibition by
monoclonal antibodies.
Cross-neutralization studieswerealso usedtoevaluate the
antigenic variation in these Fepitopesthatoccurs innature. RSV strain variation was initially observed by Coates et al. (9) and has been confirmed in more recent epidemiological studies by Mufson et al. (31) and Hendry et al. (21), using
binding assays. We wereabletodetectantigenic variation in the F epitopes of RSV clinical strains and to identify
constant, variable, and hypervariable regions of the mole-cule (Table 3, footnote c) as well as subgroup-specific neutralization epitopes. Of 11 epitopes of antigenic sites A and C, 10 were constant among subgroup A viruses; how-ever, several epitopes in site A were highly variable among subgroup B viruses. In contrast to the generally conserved nature of sites A and C, site Bvaries among both subgroup A and B strains and appears to be hypervariable among subgroup A isolates. Successful vaccine design should in-clude, as a minimum, the epitopes of antigenic site C to stimulate neutralizing antibody that will cross-react with all subgroup A and B viruses.
Although the clinical relevance of RSV F-epitope varia-tion is unknown, it does raise interesting questions regarding the immune response to natural infection and immunization. Which F epitopes should be present in a vaccine to stimulate complete and durable protection against disease in vivo? Is there adesirable repertoire of responses that will prevent the spread of infection? Are antibody responses to specific epitopes antagonistic or detrimental in vivo? Wang et al. have indicated that such restrictions do occur in individual antibody responses in their investigation of anti-influenza hemagglutinin epitope-specific responses following natural infection (51). They have observed that individuals may responddifferently to infection by producing antibodies with a wide range of specificities and thereby demonstrated that no single region of the hemagglutinin molecule was immu-nodominant. A similar situation may exist for other viral envelope glycoproteins, including RSV.
The spectrum of the immune response of infants and children to the neutralization epitopes of the RSV F glyco-protein following initial infection is unknown at present. If these epitopes are not equally immunogenic during infection of humans, we can postulate, on the basis of the results or the present study, that primary exposure to a subgroup A virus with alterations in all site B epitopes might not confer adequate protection in vivo against subsequent infection with asubgroup B virus which has the commonly conserved site B epitopes but not the conserved epitopes of antigenic sites A and AB. Thus, individual epitope-specific responses may explain, in part, the reinfection of seropositive children such as those described in the study by Mufson et al. (31). What is needed is a study that would combine the evaluation of the specific response of infants and children to the neutralization epitopes on the RSV F glycoprotein with F-epitope analysis of the RSV strains shed from each child during the initial and any subsequent infection. This would allow the identification of F epitopes that are immunodomi-nant during human infection and would give additional J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
insightintothedevelopmentof aprotectivehumoral immune response.
This study hasprovidedadetailed characterization ofthe neutralization and fusion epitopes on the F glycoprotein of RSV. In addition, we have defined important structure-function relationships that suggest that neutralization
epitopesoccupyoperationally interdependentdomainson an extremely flexible molecule. We are currently sequencing the F genes of our MARMs to identify the amino acids involved in specific epitopes, especially those that are criti-cal tofusion activity.
ACKNOWLEDGMENTS
We acknowledge Chris Winter for expert technical assistance; Brian R. Murphy for helpful discussions; Erling Norrby, Larry Anderson, andBruceFernie formonoclonalantibodies; Robert M. Chanock foracareful review of themanuscript, and Linda Jordan foreditorial assistance.
Thiswork was supportedin part by World Health Organization grantV21/181/12.
LITERATURE CITED
1. Abenes, G., H. Kidd,and R. Yanagawa. 1986. Antigenic map-ping and functional analysis of the F protein of Newcastle disease virus using monoclonal antibodies. Arch. Virol. 90: 97-110.
2. Akerlind, B., and E. Norrby. 1986. Occurrences ofrespiratory syncytial virus subtypes A and B strains in Sweden. J. Med. Virol. 19:241-247.
3. Anderson, L.J., R. A. Coombs,C. Tsov, andJ. C. Hierholzer.
1984. Useofthe biotin-avidin systemto studythe specificity of antibodies against respiratory syncytial virus. J. Clin. Micro-biol. 19:934-936.
4. Anderson,L.J., J.C.Hierholzer,C.Tsov,R. M.Hendry,B.F. Fernie, Y. Stone,andK. McIntosh.1985. Antigenic characteri-zation ofrespiratory syncytial virus strains with monoclonal antibodies. J. Infect. Dis. 151:626-633.
5. Baybutt, H.N.,andC.R. Pringle.1987. Molecularcloningand sequencing of the F and 22K membrane protein genes of the RSS-2 strains of respiratory syncytial virus. J. Gen. Virol. 68:2789-2796.
6. Buckley, A., and E. A. Gould. 1985. Neutralization ofyellow fevervirus studiesusingmonoclonal andpolyclonalantibodies. J. Gen. Virol. 66:2523-2531.
7. Carrigan, D. R. 1985. Round cell variant of measles virus: spontaneousconversion fromproductivetocell-associatedstate
of infection. Virology 144:337-350.
8. Coates, H. V., D. W. Ailing, and R. W. Chanock. 1966. An
antigenic analysis of respiratory syncytial virus isolates by a
plaque reduction neutralization test. Am. J. Epidemiol. 83: 299-313.
9. Coates,H.V.,L.Kendrick,and R. M.Chanock. 1963.Antigenic differences between twostrains ofrespiratory syncytial virus. Proc. Soc. Exp. Biol. Med. 112:958-964.
10. Coelingh, K. L. V. W., and E. Tierney. 1989. Antigenic and functional organization of human parainfluenza virus type 3 fusionglycoprotein. J.Virol. 63:375-382.
11. Coelingh, K. L. V. W., C. Winter, E. Jorgensen, and B. R.
Murphy.1987.Antigenicandstructuralpropertiesof the
hemag-glutinin-neuraminidase glycoprotein of parainfluenza type 3: sequenceanalysisof variants selected with monoclonal antibod-ies which inhibit infectivity,
hemagglutination.
andneuramini-daseactivities. J. Virol. 61:1473-1477.
12. Coelingh, K. L. V. W., C. Winter, and B. R. Murphy. 1985. Antigenic variation in thehemagglutinin-neuraminidase protein of humanparainfluenza type 3 virus. Virology 143:569-582. 13. Collins,P.L.,Y. T.Huang,andG. W.Wertz. 1984.Nucleotide
sequence of the gene encoding the fusion (F) glycoprotein of respiratory syncytial virus. Proc. Natl. Acad. Sci. USA 81:
7683-7687.
14. Fenner,M., and H. Binz. 1986. Monoclonal antibodiesspecific
forSendai virus. 1. Production andcharacterization of
mono-clonal antibodies. Scand. J. Immunol.24:335-340.
15. Fernie, B. F., P. J. Cote, Jr., and J. L. Gerin. 1982. Classifica-tion ofhybridomastorespiratory syncytialvirusglycoproteins.
Proc. Soc. Exp. Biol. Med. 171:266-271.
16. Fernie, B. F., and J. L. Gerin. 1980. The stabilization and purification ofrespiratory syncytial virus using MgSO4.
Virol-ogy 106:141-144.
17. Flamand, A., T. J. Wiktor, and H. Koprowski. 1980. Use of hybridoma monoclonal antibodies in the detection ofantigenic differences between rabies andrabies-related virusproteins. II. Theglycoprotein.J. Gen. Virol. 48:105-109.
18. Gerhard, W., and R. G. Webster. 1978. Antigenic drift in influenza A viruses. 1. Selection and characterization of anti-genic variants ofA/Pr/8!34 (HONI)influenzaviruses and
mono-clonal antibodies. J. Exp. Med. 148:383-392.
19. Gimenez, H. B., P. Cash, and W. T. Melvin. 1984. Monoclonal antibodies to human respiratorysyncytial virus andtheir use in the comparison ofdifferent virus isolates. J. Gen. Virol. 65: 963-971.
20. Gimenez, H. B., N. Hardman, H. M. Keir, and P. Cash. 1986. Antigenic variation between humanrespiratory syncytial virus isolates. J. Gen. Virol. 67:863-870.
21. Hendry, R. M., A. L. Talis, E. Godfrey, L. J. Anderson,B. F. Fernie, and K. McIntosh. 1980. Concurrent circulation of anti-genically distinct strains of respiratory syncytial virus during community outbreaks. J. Infect. Dis. 153:291-297.
22. Hsu, M. C., A. Scheid, and P. W. Choppin. 1987. Protease activation mutants of Sendai virus: sequence analysis of the mRNAof the fusion protein (F) gene and direct indentification of thecleavage-activation site.Virology 156:84-90.
23. Iorio, R. M., and M. A. Bratt. 1984. Monoclonal antibodies as functionalprobes of the HNglycoprotein of Newcastledisease virus: antigenic separation of the hemagglutinating and neur-aminidase sites. J. Immunol. 133:2215-2219.
24. Itoh, M., H. Shibitu, and M. Homma. 1987. Single aminoacid substitution of Sendai virus at the cleavage site of the fusion proteinconfers trypsin resistance. J. Gen. Virol. 68:2939-2944. 24a.Johnson,P.R., and P.L. Collins.1988. Thefusionglycoproteins of human respiratory syncytial virus of subgroups A and B: sequence conservation provides a structural basis for antigenic relatedness. J. Gen. Virol. 69:2623-2628.
25. Johnson, P. R., R. A. Olmsted, G. A. Prince, B. R. Murphy, D. W. Alling, E. E. Walsh, and P. L. Collins. 1987. Antigenic relatedness between glycoproteins of human respiratory syncy-tial virus subgroups A and B: evaluation of the contributions of FandG glycoproteins to immunity. J. Virol. 61:3163-3166. 26. Johnson, P. R., M. K. Spriggs, R. A.Olmsted,and P. L. Collins.
1987. TheG glycoprotein of human respiratory syncytial viruses ofsubgroups A and B: extensive sequence divergence between antigenically related proteins. Proc. Natl. Acad. Sci. USA 84:5625-5629.
27. Kohler, G., and C. Milstein. 1976. Derivation ofspecific anti-body-producing tissue culture and tumor lines by cell fusion. Eur.J. Immunol. 6:511-519.
28. Merz, D. C., A. Scheid, and P. W. Choppin. 1980. Importance of antibodies to the fusion glycoprotein of paramyxoviruses in the prevention of spread of infection. J. Exp. Med. 151:275-288. 29. Merz, D. C., and J. S. Wolinsky. 1983. Conversion ofnonfusing
mumps virus infections to fusing infections by selective prote-olysisof the HN glycoprotein. Virology 131:328-340.
30. Morgan, L. A., E. G. Routledge, M. M. Wilicocks, A. C. R.
Samson, R. Scott, and G. L. Toms. 1987. Strain variation of respiratory syncytial virus. J. Gen. Virol. 68:2781-2788. 31. Mufson, NI. A.,R. B. Belshe, C. Orvell, and E.Norrby. 1987.
Subgroup characteristics of respiratory syncytial virus strains recovered from children with two consecutive infections. J. Clin. Microbiol. 25:1535-1539.
32. Mufson,M.A., C. Orvell,B.Rafnar,and E. Norrby. 1985.Two
distinct subtypesof humanrespiratory syncytialvirus. J.Gen. Virol. 66:2111-2124.
33. Murphy, B. R., and E. E. Walsh. 1988. Formalin-inactivated
respiratory syncytial virus vaccine induces antibodies to the
on November 10, 2019 by guest
http://jvi.asm.org/
2950 BEELER AND COELINGH
fusion glycoprotein that are deficient in fusion-inhibiting activ-ity. J. Clin. Microbiol. 26:1595-1597.
34. Norrby, E., and Y. Gollmar. 1975. Identification of measles virus-specific hemolysis-inhibiting antibodies separate from hem-agglutination-inhibiting antibodies. Infect. Immun. 11:231-239. 35. Olmsted,R.A., N. Elango, G. A. Prince, P. R. Johnson, B. Moss, R. M. Chanock, and P. L. Collins. 1986. Expression of the F glycoprotein of RSVby arecombinantvaccinia virus: compar-ison of theindividual contribution ofthe FandG glycoprotein to hostimmunity. Proc. Natl. Acad. Sci. USA 83:7462-7466. 36. Orvell,C., E. Norrby, and M. A. Mufson. 1987.Preparation and
characterization ofmonoclonalantibodies directed against five structural components of human respiratory syncytial virus subgroupB.J. Gen. Virol.63:3125-3135.
37. Porthier, P., S. Ghim, T. B. Bour, J. B. Gouyon, and M. Dauvergne. 1987. Antigenic variation ofrespiratory syncytial virusinrecurrent infection. Eur. J. Clin. Microbiol.2:212. 38. Portner, A., R. A. Scroggs, and D. W. Metzger. 1987. Distinct
functions ofantigenic sites on the HNglycoprotein ofSendai virus.Virology 158:61-68.
39. Portner, A., R. A. Scroggs, and C. W. Naeve. 1987. The fusion glycoprotein of Sendai virus: sequence analysis ofanepitope involved in fusion and virusneutralization. Virology 157:556-559.
40. Prince, G. A., R. L. Horswood, D. W. Koenig, and R. M. Chanock. 1985. Antigenic analysis ofaputative new strain of respiratory syncytial virus. J.Infect. Dis. 151:634-637. 41. Routledge, E. G., M. M. Willcocks, L. Morgan, A. C. R.
Samson,R.Scott, and G. L. Toms. 1987.Heterogenicity of the respiratory syncytial virus 22K protein revealed by Western blotting with monoclonal antibodies. J. Gen. Virol. 68:1209-1215.
42. Sato,T.A.,A.Fukuda, and A. Sugiura. 1985. Characterization of themajor structural proteins of measles virus with monoclo-nalantibodies. J. Gen. Virol.66:1397-1409.
43. Shioda, T., S. Wakao, S. Suzu, and H. Shibuta. 1988. Differ-ences in bovine parainfluenza 3 virus variants studied by
se-quencing of the genes of viral envelope proteins. Virology 162:388-396.
44. Smith, D. B., and S. C. Ingliss. 1987. The mutation rate and
variabilityof eukaryotic viruses: an analytical review. J. Gen. Virol. 68:2729-2740.
45. Stanley, J., S. J. Cooper, and D. E. Griffen. 1985. Alphavirus neurovirulence: monoclonal antibodiesdiscriminating wild-type from neuroadapted Sindbis virus. J. Virol. 56:110-119. 46. Storch, G. A., and C. S. Park. 1987. Monoclonal antibodies
demonstrate heterogeneity in the Gglycoprotein of prototype strains and clinical isolates of respiratory syncytial virus. J. Med. Virol. 22:345-346.
47. Tikasingh, E. L., L. Spence, and W.G. Downs. 1966.The use of adjuvant and sarcoma 180 cells in the production of mouse
hyperimmune ascites fluids toarboviruses. Am. J. Trop. Med. Hyg. 15:219-226.
48. Trudel, M., F. Nadon, C. Sequin, G. Dionne, and M. Lacroix. 1987. Identification of a synthetic peptide as part ofa major neutralization epitope of respiratory syncytial virus. J. Gen. Virol. 68:2273-2280.
49. Walsh, E. E., M. W. Brandriss, and J. J. Schlesinger. 1987. Immunological differences between the envelope glycoproteins of two strains ofhuman respiratory syncytial virus. J. Gen. Virol.68:2169-2176.
50. Walsh, E. E., P. J. Cote, B. F. Fernie, J. J. Schlesinger, and M. W. Brandriss. 1986. Analysis ofthe respiratory syncytial virusfusion protein using monoclonal andpolyclonal antibod-ies.J.Gen. Virol.67:505-513.
51. Wang, M.-L., J. J. Skehal, and D. C. Wiley. 1986. Comparative analyses of the specificities of anti-influenza hemagglutinin antibodies in human sera. J. Virol.57:124-128.
52. Ward, K. A., J. S. Everson, P. R. Lambden, and P. J. Watt. 1984.Antigenicand structuralvariation in themajor nucleocap-sid protein ofrespiratory syncytial virus. J. Gen. Virol. 65: 1749-1757.
53. Webster, R. G.,W.G.Laver, G. M.Air, C. Ward, W.Gerhard, and K. L. vanWyke. 1982.The mechanism ofantigenic driftin influenza viruses: analysis of Hong Kong (H3N2) variantswith monoclonal antibodies to the hemagglutinin molecule. Ann. N.Y. Acad. Sci.354:142-161.
54. Wulff, H., P. Kidd, and H. A. Wenner. 1964. Respiratory syncytial virus: observationsonantigenic heterogeneity. Proc. Soc. Exp. Biol. Med. 115:240-243.
J. VIROL.