Copyright C) 1985, AmericanSociety forMicrobiology
Rous
Sarcoma
Virus
Variants That
Encode
src
Proteins with
an
Altered Carboxy
Terminus
Are
Defective
for
Cellular
Transformation
VICTORIA W. WILKERSON, DEBRA L. BRYANT,t ANDJ. THOMAS PARSONS* Department of Microbiology, University ofVirginiaSchool of Medicine, Charlottesville, Virginia22908
Received 11February 1985/Accepted 13 April 1985
ThesrcgeneofRoussarcomavirus (v-src)anditscellular homolog, thec-srcgene,share extensivesequence
homology. Themostnotable differences betweenthesegenesreside intheregion encodingthe carboxy terminus of thesrcproteins. We constructed mutationswithin the3'endofthev-src genetodetermine thesignificance of thisregiontothe transforming potential ofthev-srcprotein, pp6O-src. ThemutantsCHd1300 and CHisf511 containmutationsthatalter thelast23 aminoacids ofpp6ovsc, whereas the mutantCHis1545-C containsa
linkerinsertion that alters the last11amino acids of pp6Ov-src, and themutantCHisl545-HIcontainsa linker
insertionthatresults ina9-amino-acid insertionatposition 415. Plasmids bearingeachof these mutationswere
unable to transform chicken cells when introduced into these cells by DNA transfection. In addition, the structurally altered srcproteins encoded by the mutantshad much-reduced levels of tyrosine protein kinase activity invivo, asmeasuredbyautophosphorylation and phosphorylation of the34,000Mrcellular protein,
andinvitro,asdetermined by measuringthe levelof pp6osc autophosphorylation. Thesedataindicate that the carboxy-terminal aminoacidsequences play an important roleinmaintaining thestructure of the catalytic
domain ofpp6ov-src. In contrast, the transfection of chicken cells with plasmidDNA containing a chimeric v-c-srcgeneresultedinmorphological celltransformation andthesynthesis ofanenzymatically active hybrid protein. Therefore, the carboxy-terminal sequencealterationsobservedinthec-srcproteindonotaloneserve
toalter the functional activity ofa hybridv-c-srcprotein appreciably.
Theinduction of cellular transformation byRous sarcoma virus (RSV) requires the functional expression of the viral src (v-src) gene product pp605rc (1, 3, 25).
pp6Osrc
is a tyrosine proteinkinaseandisamemberofafamily ofrelated oncogene products, each of which is either a functional tyrosine protein kinase or is structurally related to the tyrosine kinases(1, 10, 20,27, 34). Invitro,pp6Osrc catalyzes the tyrosine-specific phosphorylation of a variety ofsub-strates(9, 11, 24), including phosphatidyl inositol lipids (36,
55).
In vivo, the cellular expression of functional pp6Osrc results in thetyrosine-specific phosphorylation ofanumber ofcellular proteins (12, 13, 19, 44,47). The intrinsic kinase activity of pp6Osrc, coupled with thetransformation-specific
tyrosine phosphorylation of certain cellular proteins and lipids, suggests that the src protein plays a critical role in cellular transformation via the phosphorylation of cellular targetmolecules.
The biochemical characterization of
pp6Osrc
andstructur-ally alteredsrcproteins encoded byRSVmutantshas ledto theconclusion that pp6Osrc contains several distinguishable functional domains (4,5, 15, 40). Considerable evidencehas shown thatpp6osrc isassociated with the plasma
mnembrane
via an amino-terminal domain (14, 15, 32) and that this associationwith themembraneisrequiredfor cellulartrans-formation (17). The observation that
pp6Osrc
contains lipid covalentlyboundto anamino-terminalresidue, glycine (45), suggests thatthismoietymaycontributetotheanchoring of the src protein in the plasma membrane (7, 8, 17, 48). Inaddition,
theanalysis
ofdeletion mutations within the 5' half of the src gene has indicated the presence of a second* Correspondingauthor.
tPresent address: Department of Molecular Biology, Unogen, Inc.,Charlottesville, VA 22901.
amino-terminal domain located between residues 130 and 169thatisrequired for morphological cell transformation(4, 40; V. W. Wilkerson and J. T. Parsons, manuscript in preparation). A variety of experimental evidence has indi-catedthat the
carboxy-terminal
half of pp60src contains the functional domain forprotein kinase activity (5, 16, 39, 52, 53).Theextensive amino acid sequence homologybetween
thecarboxy-terminalhalfof pp60srcandother
viral
tyrosine protein kinases furthersupportsthe conceptthat thisportion ofpp6Osr(
contains the functional catalytic domain or do-mains necessary for tyrosine protein kinase activity (1). Recently, we showed that mutants containing singleamino
acidchangeswithin aregion that isconserved among virtu-ally all ofthe
tyrosine
kinases (the A-P-E orala430-pro431-g1u432
sequence)aredefectivefor celltransformation (5,40). The biological andbiochemicalproperties
ofthese mutantsstrengthenthe argument thatthisregion ofthesrc
protein
is part of a functional domain whose structuralintegrity
is required forcellular transformation. Recently, Takeya and Hanafusa (56) determined the nucleotide sequence of thecoding
region
ofthe normal cellularhomolog (c-src)
ofthe v-src gene,pp60c-src',
andcompared it with the sequenceof the v-src gene. The deduced amino acid sequences(56)
exhibit virtual homology except within the
carboxy
termi-nus,where thecarboxy-terminal 19aminoacidsofpp60c-src
are replaced by a new set of 12 amino acids ofpp60vsrc. Recently, several groups have shown that
overexpression
of the c-src gene product does not lead to overt cellular transformation(29, 39, 49,50),indicating
thatthestructural differences between the c-src and v-src geneproducts
are essential formediating cellulartransformation.
We investigated the role ofthe
carboxy-terminal
amino acid sequences in the maintenance offunctional
srcprotein
activity by introducing deletions and
oligonucleotide
inser-314
on November 10, 2019 by guest
http://jvi.asm.org/
3' src GENE MUTATIONS 315 Bgl 11
1530
Bgl I Mst 11 Pvu 11
I I I
1550
PuII
1570
. CGGMGGACCCTGAGGAGCGGCCCACCmAAGTACCTGCAGGCCCAGCGCTCCCTGGCTTGThGGGAGGTCGCTGAGTAA..
ArgLysAspProGluGluArgProThrPheLysTyrLeuGInAlaGInLeuLeuProAlaCysVa1LeuGluValAlaGlu***
S00 510 520
4j300
jxl5ll
jlS545-C
1&545-H
ArgLysAspProProLeuSerThrCysAr Pr oSerCY sSe rL.uLeuValCysTrDArtSerLeuSe rLvsTvrG 1uA1&**
...CGGAAGGACCCtgacatcgatgtgaGGAGCGGCCCACCflAAGTACCrGCAGGCCCAGCTGCrCCCTGCITGlTlTGGGMGTCGCrGiTAAGTACGAGGfTGAA... ArgLysAspProAs_1lCAsoVArPAlyGlyrLeuSl rThaGi_y sArgPrSerCvS.r-LeuVal CysTrvArnScrLeuS.rLvsTvrGluAMj*0*
..CGGAAGGACCCTGAGGAGCGGCCCAC=AATGTACCTGCAGGCCCAGcat cgat&CTGCTCCCTGCITTTG'GTTGMGAGGTCGCTGAGTAAGTACGAGGCGTAG..
ArgLysAspProGluGluAirgProThrPheLysTysLeuGInA aGInHisAr&CvstCysSer-LeuLeg_VlCysTryA.riSerLeuSer-LxsTvrlu*
..CGGAAGGACCCTGAGGAGCGGCCCACCIAAhGTACCTGCAGGCCCAGcc caagc ttgggcca*a*gcttggcgTCCCTGCMUTGTliGlGAGGTCGCrGA;TAA.. ArgLysAspProGluG1uArgProThrPheLysTyrLeuGI1nAla*G1nProLvsLeuG lyProSe rLeuGlXVIlProAla*CysVal LeuGl1uVaA1 *G 1u***
CGGAGGGACCCrGAGGAGCGGCCCAClGTGAGTACCTGCAGGCuCCTGGAGGACrACrTCACCTCGACAGAGCCCCAGouCCAGCCTGGAGAGACCTATAG AtrgAOrgAspProG1 Gl1uArgProThrPheG 1uTyrLeuGI1nAla*PheLuGl1uAspTyrrPheThrSerThrGl1uProGl1uTyrG1nProGlvGluAsnLeu***
FIG. 1. Nucleotide sequence ofmutations within the 3' terminus of the RSV src gene. Insertions ofsynthetic oligonucleotides are designated bylowercase letters.Aminoacidsequencesthat differ fromwild-typeviralsrcareunderlined. Amino acidsequencedifferences betweenv-srcandc-src areindicated withadashed line.Sequencealterationswereidentifiedbydigestionwithrestriction endonuclease and bydirectDNAsequenceanalysis (37).Relevant restriction endonuclease recognition sites within thesrcgene areincluded.
tions into the 3' end of the src gene. Mutant viruses generated bythetransfectionofmutagenizedviralDNA into chicken cells encode src proteins containing alterations in
thecarboxy-terminal 23 amino acids of pp60src. Analysis of the proteins encoded by these viruses showed that gross structural alterations of the carboxy terminus of pp6osr( resulted in transformation-defective viruses that encode enzymatically inactive srcproteins. However, the
substitu-tion ofsequencesfrom the 3' end of the c-srcgenedoesnot lead to an appreciable loss of transforming activity in the
hybrid virus, and thesrc protein encoded bythesechimeric
genes retains wild-type levels of tyrosine protein kinase activity.
MATERIALS ANDMETHODS
Cells, viruses,andplasmids. Chickenembryocell cultures
wereprepared from gs-negative embryos (Spafas) and
main-tained as previously described (23). Prague A (PrA) RSV
DNA (4, 26) used for mutagenesis was contained in a
nonpermutedmolecularclone of RSVinserted intoapBR322
plasmid vector (pJD100). The transfection of chicken cells withpJD100 or mutagenized pJD100 DNA was carriedout
by applying 1to2 p.gof purifiedplasmid DNA (28)tocultures of chicken cellsas describedpreviously (26). Since pJD100 and itsmutagenized derivatives containnonpermuted copies of theRSVgenome,transfected cellsproduce high levels of mutantvirus, whichwas routinely used for subsequent
ex-periments.
Restrictionenzymedigestion andagarosegel
electrophore-sis. Restrictionenzymes werepurchased fromNewEngland
Biolabs, Inc., Beverly, Mass.,orBethesda Research
Labo-ratories, Inc., Gaithersburg, Md.,andusedasrecommended
by the supplier. DNA restriction fragments were purified
either from agarose gels containing low-gelling-temperature
agarose (SeaPlaque FMC Corp., Marine Colloids Div., Rockland, Maine) or by adsorption to and elution from DEAE-membrane paper (Schleicher and Schuell, Inc., Keene, N.H.). Southern blotanalysis(54)of cellularproviral DNAwas carriedoutas describedpreviously (23).
Construction of mutations. Deletion mutationswere intro-duced at each of the two MstII restriction sites in the src
geneof the RSVgenome (nucleotidepositions25 and 1509) (46) bythepartial digestionofpJD100DNA withMstII,the purification of linear molecules, and treatment with the exonuclease BAL 31 (Bethesda Research Laboratories) as
described previously (4). After BAL 31digestion, staggered endswere repaired with Escherichia coli DNA polymerase Klenow fragment (New England Biolabs) and the linear DNA moleculeswere ligated with T4 DNAligase and used
totransformE. coli HB101 cells(18). Theresultingcolonies
were screened for deletion mutations in the src gene by
isolating plasmid DNA from 1-ml cultures (28), digesting with MstII,andanalyzingthe DNAfragments byagarosegel
electrophoresis. Insertion mutations were constructed by partiallydigesting pJD100DNA with either MstII orPvuII,
purifying linear DNA, and religating in the presence ofan
8-base-pair (bp) ClaI linkeror a 12-bpHindIII linker(New England Biolabs). In thecase of DNAdigested with MstII, the staggered ends were repaired with Klenow fragment beforeligation. Plasmid DNAwasused totransform E. coli HB101 cells,and theresultingcolonieswerescreenedfor the Mst 11
1S10
NstII
A I I I I
-E---
0
VOL.55, 1985
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.61.540.67.355.2]absenceofeither a MstII or a PvuII restriction site and the acquisition of either a ClaI or a HindlIl restriction site. All mutations were confirmedby direct DNA sequencing (37).
Immunoprecipitation and polyacrylamide gel analysis of labeled cell proteins. Infected chicken cells were labeled with [35S]methionineor
32p,
for 4 to 6 h asdescribed previously (4, 41, 43). Immunoprecipitationof src protein from labeled cell extracts wascarried out as follows. Cells were washed twice with STE (0.15 M NaCl, 50 mM Tris hydrochloride [pH 7.2], 1mMEDTA) and lysed in 2 ml of 10 mM Tris hydrochloride (pH7.2)-0.1 MNaCI-1 mM EDTA- 0.5%deoxycholate-1% Nonidet P-40. Lysates were clarified at 100,000 x g for 30 minand incubatedwith rabbitanti-p60srcserum (22) or rabbit anti-36K serum (19) at 0°C for 60 min. Immune complexes wereadsorbed to Formalin-fixedStaphylococcusaureus (30) and washed once in lysis buffer, once in 10 mM Tris hydrochloride (pH 7.2)-i M NaCl-1% deoxycholate-1% Triton X-100-0.1% sodium dodecyl sulfate-1 M urea, and once in the original lysis buffer. The immune complexes were then suspended in sample buffer, boiled, and electrophoresed on a 9.0% polyacrylamide gel (33). Gels werefixed, stained, dried, andautoradiographed. Gels con-taining 35S-labeled proteins were treated with En3Hance (New England Nuclear Corp., Boston, Mass.) to provide fluorographic enhancement before drying. Toquantitate the relativeamountofpp6srcorstructurallyaltered forms of src protein in the labeling experiments,we immunoprecipitated equalamounts of labeledprotein extract. Immune complex protein kinase assays were performed as described previ-ously (4, 42). To compare the levels of kinase activity of wild-type and structurally altered src proteins, weim-A.
1 2 3 4
I">
_W.B.
1 2 3
5 6 7 8 9 10
---r"0
-mI4I
pp8osrc
"z.1-;PP52"
-w
_I-4FW # ....30 _ppASr
4 5 6 7 8 9 10
FIG. 2. Immunoprecipitation ofsrc protein froi
with[35S]methionine(A)orwith32p,(B). Chickencel
PrARSV or mutant viruses werelabeled with [35S 32p,, harvested, immunoprecipitated with rabbitant andanalyzed by polyacrylamide gel electrophoresi ography as described in the text. Lanes: 1, unin'
PrA-RSV-infected cells; 3, d1300-infected cells; 4, cells; 5, uninfected cells; 6, PrA-RSV-infected c infected cells; 8,pm9-infected cells; 9, isl545-C-inf is1545-H-infectedcells.
A.
1 2 3 4 5 6
pp8 _kt
-B.
123 4 5
-._ -~~~p368
FIG. 3. In vitroand in vivo protein kinase activity of src protein from cells infected with PrA RSV or mutant viruses. Parallel cultures of cells were either harvested, immunoprecipitated with rabbit anti-p60src serum, and assayed for autophosphorylation as described in the text (Panel A) or labeled with 32p,, harvested, immunoprecipitated with anti-36K-protein serum, and analyzed by polyacrylamide gel electrophoresis and autoradiography. Lanes: 1, uninfected cells; 2, PrA-RSV-infected cells; 3,d1121-infectedcells; 4, pm9-infected cells; 5, is1545-C-infected cells;isl545-H-infected cells.
munoprecipitated parallel cultures of unlabeled and
[35S]methionine-labeled
cells with antiserum, and deter-mined the amountofpp6Osrc autophosphorylationrelative to the level of35S-labeled
srcprotein in immune complexes.Indirect immunofluorescence staining of cells. RSV-infected anduninfectedchicken cellswere seeded on cover slipsandallowedtogrowovernight. Cells werefixed in 3% paraformaldehyde, permeabilizedin0.4% TritonX-100(35), and incubated with 100 1LI of anti-src monoclonal antibody EB7(42) for30 min at roomtemperature. Thebindingof EB7 antibodywas localized by incubation ofthe cells withgoat F(ab')2 anti-mouse immunoglobulin G (Jackson Immuno Research) (10,ug/ml) for30 minfollowedbyincubationwith fluorescein-conjugated rabbit F(ab')2 anti-goat F(ab')2 (Jack-son ImmunoResearch) (10
p.g/ml)
foran additional 30 min. Between incubations with antibody, the cover slips were washed three times with phosphate-buffered saline. The stainedcells wereexamined withaLeitz fluorescence micro-scope(Leitz/Opto-Metric
Div. ofE. Leitz Inc., Rockleigh, N.J.) equippedwith epi-illumination.RESULTS
Isolation of 3' src mutations. In ourinitialexperiment, we soughttoprobetheeffects ofstructuralmodificationswithin the carboxy-terminal region ofthe src protein. Therefore, deletion mutations near the 3' end of the src gene were
constructedbyutilizingaMstIIrestriction site located 71bp upstream from the 3' terminationcodonofthesrcgene
(Fig.
1). Deletionswereintroduced by exonucleaseBAL31 diges-tion,repair,andreligationwithT4ligaseasdescribed above. -
ppeOSrC
DNA from individual transformants wasdigested
withMstII,
andplasmids
containing deletions oftheMstIIsite inp
p52arc
the 3' end of the src gene were identifiedby
agarosegel
electrophoresis. DNA sequence
analysis
ofone suchmuta-m cells labeled tion, d1300, revealed a deletion of bp 1509 to bp 1518 in the lls infectedwith src gene resulting in a deletion ofGlu504to
Arg506
followed ]methionine or by a shift to an alternate reading frame (Fig. 1). Whenti-p6rc
serum, chicken cells were transfected withdl300DNA,
nomorpho-es
and
autoradl-
logical
transformation wasobserved. Two additionalmuta-islS11-infected
tions within the carboxy-terminal sequences werecon-iells-
7,dll21-
structedbylinker insertion. A mutationwasobtained by the Fectedcells; 10, insertion ofaneight-bp
ClaI linker at the same MstII site utilized above (Fig. 1). This mutation, isl511, has anon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.324.562.69.174.2] [image:3.612.68.306.399.630.2]3' src GENE MUTATIONS 317
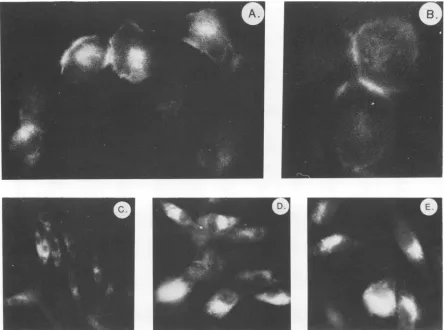
FIG. 4. Immunofluorescent staining of cells infected with PrA RSV or mutant viruses. Cells were grown on cover slips, fixed, permeabilized, and stained withamonoclonalantibodytopp60src(EB7)asdescribedinthetext.(A) and (B), PrA-RSV-infectedcells; (C)and (D),pm9-infected cells; (E), isl545-H-infected cells.
tion of11 nucleotides (3 nucleotides fromthe
repair
ofthe MstII staggered ends), which resulted inaframeshiftaltering the last 23 amino acids ofpp60src.
The insertion ofan 8-bp ClaIlinkerataPvuIIrestriction site36bp fromthe3' endof the src gene (Fig. 1) yielded the mutationis1545-C,
which contains a shift to the same reading frame as the is1511 mutation butresults inanalteration of onlythe last 11amino acids ofpp6Osrc. Transfection with DNAs from isl511 andis1545-C
did not induce a morphological transformation of chicken cells.Finally,a12-bp HindlIl linkerwasinsertedat the 3'-terminal PvuII restriction site, yielding an inframe insertion of24nucleotides(the result of the insertion oftwo linker molecules). This mutation,is1545-H,
results in aninframe insertion of eight amino acids (Fig. 1).Transfection withDNAfromCHisl545-Halso did not result in a morpho-logical transformation of chicken cells.
Characterization of structurally altered src proteins. The expression ofthe mutant viruses encoding proteins having
an altered carboxy terminus was examined in chicken cells infected with high-titered stocks of the mutant viruses
CHdI300,
CHisl511,
CHisl545-C,andCHisl545-H. In addi-tion, cellswerealsoinfected with two other transformation-defectiveviruses,CHdl121 andCHpm9.CHdl121
encodes a srcproteincontainingadeletion of sequencesg1y82
toarg169 (40), and CHpm9contains a single point mutation resulting in a proline-to-serine alteration at residue 431 (6). Infected cellswerelabeled with[35S]methionine
(Fig.2A) or32p,
(Fig. 2B), labeled cell extracts were immunoprecipitated withrabbit anti-p60src serum, and the immune complexes were
analyzed by polyacrylamide gel
electrophoresis. Labeling
with [35S]methionine revealed that each of the carboxy-terminal mutants encoded an approximately 60,000-mole-cular-weight (60K) protein,asexpectedfrom the nucleotide sequenceanalysis(Fig. 2A,lanes3, 4, 9, and10). However, additional, slightly smaller, src-related proteins (molecular weight, 57,000 to 59,000) were often detected in im-munoprecipitatesof cellsinfected withd1300 andisil11
(Fig.
2A, lanes 3 and 4). These lower-molecular-weight proteins appear to be degradation products of pp60src', based on S. aureus V8partial peptide mapping (datanotshown). Label-ing of parallel cultures with
32Pi
and immunoprecipitation withanti-p60src serumrevealed that thesrcproteins contain-ing carboxy-terminal alterations were significantly underphosphorylated comparedwithwild-typepp6Osrc
(Fig. 2B, lanes 2 and6), pm9pp6Osrc (lane 8),or pp52srcencoded bythedeletionmutantd1121 (lane 7).We havealso observed that[35S]methionine-labeled
immunoprecipitates from cells infected with virus containing carboxy-terminal src muta-tions exhibited diminished amounts of the cellular protein pp9O (D. L. Bryant, V. W. Wilkerson, and J. T. Parsons, unpublished results; L. A. Lipsich and J. Brugge,un-published results). Sincepp9Oandasecond cellularprotein, pp5O, arenormally associated with soluble pp6Osrcin RSV-transformed cells (2, 38; J. Brugge, Curr. Top. Microbiol. Immunol., inpress), it appears that thesrcproteins contain-ing carboxy-terminal alterations do notassociate withpp9O
VOL. 55, 1985
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.81.526.72.402.2]318 WILKERSON ET AL.
* *
Bgi I
m
=mum
Bam HI
E *EUEI I
*
U-...-c-src
1I
4
1N
- j
~~~~~~~~I
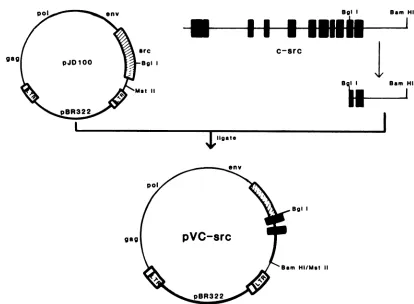
Sam HiFIG. 5. Construction of the plasmid pVCsrc containing a chimeric src gene. The plasmid pJD100 was digested withMstII, the ends were repaired withKlenow fragment of DNA polymeraseI,and then theplasmid was digested withBgIlyielding a vectorfragment missing the 3'end of the viralsrcgene. The cognate c-src fragment was prepared from the plasmid pSVOHCS11 by digestion withBamHI and repaired withKlenowfragment as describedpreviously. Digestion withBgll, which cleaves the c-src DNA at the identical site as v-src, yielded a BglI-blunted BamHIc-srcfragment, which was then inserted into theBglI-bluntedMstII pJD100 vector to form the chimeric plasmid pVCsrc.
andppSOorthat the complex is unstable in mutant-infected cells. Thelack of association of pp9O and pp5O with thesrc
protein encoded by is1545 has been confirmed by im-munoprecipitation with antiserum to the pp9O protein (Lipsich and Brugge, personal communication).
To determine the effects of alterations of the carboxy terminus on tyrosine protein kinase activity, we infected cells withmutantorwild-typeRSV andimmunoprecipitated the lysates with anti-p6Osrc serum; we also incubated the
immunecomplexes with-y-[32P]ATPto measurethelevel of
src autophosphorylation. The 3' mutations isl545-C and
isl545-H encode src proteins with significantly reduced
levels of invitro phosphorylation activity (Fig. 3A, lanes 5 and6). Thelevelofkinase activity observedwas similarto thatexhibitedbyimmunecomplexesfromcellsinfectedwith the point mutation pm9 (lane 4). Immune complexes from cells infectedwith d121, an amino-terminal deletion muta-tion, also exhibited a level of tyrosine kinase activity
re-duced fromwild-typelevels(lane 3). Thephosphorylation of the cellular 36K protein, which is phosphorylated
pre-dominantly on atyrosineresidue in RSV-infected cells (19,
46), was also examined in mutant-infected cells (Fig. 3B).
Parallel culturesofinfectedcellswere labeled with
32Pi
and[35S]methionine, and extracts were prepared and
im-munoprecipitated with anti-36K-protein serum. In cells
in-fected withvirusbearingthecarboxy-terminalsrcmutation,
the 36K protein was underphosphorylated compared with
dl121-infected or wild-type RSV-infected cells (Fig. 3B).
Similar levelsof 36Kproteinwerepresentineach celllysate
as determined by the immunoprecipitation of [35S]methio-nine-labeledcells(data not shown). These results reveal that structuralperturbation ofthecarboxy-terminalsequence, as delineatedby themutations showninFig. 1, results in RSV mutants defective for cell transformation. Such mutants encode structurally altered src proteins significantly dimin-ished in their capacity to phosphorylate substrates both in vitro and invivo.
Intracellularlocalization of structurally alteredsrcproteins. Wild-type
pp6Osrc
in RSV-infected cells isprimarily located in the plasma membrane (14, 32). In addition, there is evidencesuggestingthat theassociation ofpp6Osrc with pp5O andpp9Ois involved in the transportofpp6Osrc
totheplasma membrane (14; J. Brugge, in press). We examined the intracellular distribution ofthe altered src proteins by im-munofluorescence localization techniques (35). Cells in-fected with PrA RSV, CHpm9 (6), and CHisl545-C were grown on coverslips, fixed with formaldehyde, permeabil-ized,andstained with monoclonalantibodytopp6OSrc. Cells infected with either CHpm9(Fig. 4C and D)orCHisl545-C (Fig. 4E) routinely exhibited an asymmetric perinuclear patternof fluorescence with no apparentlocalization ofsrcproteinattheplasmamembraneorwithinadhesionplaques
or cell-cell junctions. In contrast, wild-type virus-infected cellsexhibitedstainingattheplasmamembraneandcell-cell junctures (Fig. 4A and B). The altered. pattern of
pp6Osrc
distribution within the cell, asrevealedby the fluorescence localization experiments, suggests that structural modifica-tions resident in thesrcproteins encodedby CHisl545-Cor J. VIROL.
.l
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.101.515.62.366.2]3' src GENE MUTATIONS 319
1
2
3
4
rn-p
FIG. 6. Southernblotanalysis of chicken cells infected withPrA RSV orpVCsrc. Total cell DNA was isolated from cells infected withPrARSVorpVCsrc, digested withEcoRI,electrophoresed on an agarosegel, and transferredto nitrocellulose by the methodof Southern (54). A 32P-labeled 1.7-kilobase MstII fragment from pJD100containing the entirev-src gene wasusedtoprobe the cell DNA blot for src-specific fragments. Lanes: 1, pJD100 DNA; 2, pVCsrcDNA;3,PrA-RSV-infected cell DNA; 4,pVCsrc-infected cell DNA.
CHpm9 influence the localization of src to the plasma membrane. Preliminarybiochemicalanalysis ofthe localiza-tion ofthe src protein encoded by CHisl545-C revealed a less stable association with cellular membrane compared with wild-type pp6Osrc (LipsichandBrugge, unpublished).
Construction andanalysis of a chimeric src genebearingthe 3'endof thec-srcgene. Direct DNA
sequencing
of thev-src and c-src genes has shown that the carboxy-terminal 19 aminoacidsofpp6ocsrcwerereplaced byanewsequenceof 12amino acids of pp6Ov-sr, (Fig. 1). Becauseofour observa-tions thatmutations introduced intothe v-src geneproximal to the site ofsequence divergence between v-srcand c-src (Fig. 1) result in a substantial reduction in transforming activity and protein kinase activity, we undertook to con-struct a hybrid src gene containing nucleotide sequences derived from v-src and c-src genes. The hybrid src gene (v-c-src) (Fig. 5) encodes an src protein containing amino acid residues 1 to 431 ofv-src and residues 432 to 533 of c-src. The transfection ofchicken cells with the plasmid DNA containing the hybrid src gene resulted in cellular transformation; however, foci were detected 7 to 10 days later than in cellstransfected withplasmid DNA containing only the wild-type viral DNA. Because ofthe delay in the appearanceof transformed foci in cells transfected withthe hybridsrcgene,thestructureof theintegrated proviralDNA wasexamined to determine ifrearrangementof the srcgene hadoccurredduringthe infection process.Figure6 shows a Southern blot ofDNA isolated fromcells transfected with either wild-type PrA DNA (pJD100) or the hybrid v-c-src DNA (pVCsrc) and digested with the restriction enzyme EcoRI.Cells transformed withwild-type DNAcontainedthe characteristic 3.09-kilodalton EcoRI src fragment (23), whereas cells transfected with pVCsrc DNA contained a 4.2-kilodalton EcoRI srcfragment
(Fig.
6).Immunoprecipi-tation of cells infected with pVCsrc and labeled with [35S]methionine revealed the presence of a 61-kilodalton protein whose electrophoretic mobility was slightly slower than pp6Ov-src (Fig. 7A). The change in the electrophoretic
k
b
p
mobility
ofpp`v-c-src
is consistent with thepredicted
se-quence alteration of the c-srcprotein. In a parallel experi-ment, theprotein kinase activityofpp61v-c-srcwascompared with that of pp60v-src. Both src proteins exhibited similar activities measured by either the src autophosphorylation (Fig. 7B) or thephosphorylationof the exogenous substrate casein (data not shown). These data clearly indicate that
4. 1
0
replacement
ofthecarboxy-terminal
sequencesofv-srcwiththe cognate sequencesofc-srcdoes not significantly alter the ability ofthe src geneproduct tomediate cellular
transfor-3.0
9
mationor to function invitroas atyrosine protein kinase.DISCUSSION
We used site-directed mutagenesis techniques to isolate deletionmutations and linkerinsertionmutationswithin the 3' endoftheRSVsrcgene. Analysis of a deletion mutation, insertion-frameshift mutations, and an inframe insertion mutation revealed that structural alteration of the carboxy-terminal portion of the pp60src protein leads to the loss of cell-transforming function and a loss in tyrosine protein kinaseactivity. The minimal mutationrequiredtoinactivate the src proteinin this studywas aninsertion of nine amino acids at
position
415 of the src protein sequence. Interest-ingly, this insertion mutation occurredadjacentto thepoint ofsequencedivergence
betweenthev-srcandc-srcproteins. However,whenachimericv-c-srcgene wasconstructed and introduced into chicken cells in the context of an RSV vector, we observed transformation ofthe cells and identi-fied an enzymatically active hybrid srcprotein
by im-munoprecipitation of cellextracts. Theprecisenature of the structural motifthatwasalteredbyourmutations is unclear at this time; however, a comparison of the deduced amino acid sequences of thetyrosine
kinases encoded by thefps andyes oncogenes revealsa highly conserved leucine resi-due atposition
517 (31, 51). This residue was present inpp60`c5src
but was altered(replaced)in the mutantis1545-H (Fig. 1).Itispossiblethatthis residue iscritical intheproperA.
ppsOsrc
B.
1
2
3
4
1
2
3
4
ppe60rc-4 W
FIG. 7. Identification ofsrc proteinsin cells infected withvirus containing a v-c-src gene. Parallel cultures of chickencells were eitherlabeledwith[35S]methionine, harvested,immunoprecipitated with rabbitanti-p6O0r" serum, andanalyzed by polyacrylamidegel electrophoresis (A) or harvested, immunoprecipitated with anti-p64Yr" serum,andassayed forautophosphorylationasdescribedin Fig. 2. Lanes: 1, uninfected cells; 2, PrA-RSV-infected cells; 3, pVCsrc(clone2)-infectedcells; 4, pVCsrc (clone15)-infectedcells. Clones 2 and 15 ofpVCsrc representindependent isolates of the plasmidsbearingthechimericv-c-srcgene.
VOL.55, 1985
404.;
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.52.290.71.286.2] [image:6.612.311.551.513.633.2]folding
of thecarboxy-terminal portion
of the src protein sequence.srcproteins havingastructurallyalteredcarboxyterminus exhibited a
significant
reduction intyrosine protein
kinaseactivity
both in vitro, by the functional immune complex kinase assay, and in vivo, by thedegree oftyrosinephos-phorylation
of the 36Kprotein,aprotein readily phosphory-latedontyrosine invivo inRSV-transformed cells. That theunderphosphorylation
of the 36Kproteinisaconsequence of amutation withinthecatalyticdomaingivingriseto inactive mutantsrcprotein
issupported bytheobservationthat cells infected with the amino-terminal deletion mutant CHdl121 containsignificant
levels ofphosphorylated
36Kprotein.It isinteresting
to note that, as we reported earlier with muta-tions within the A-P-E-A sequence ofpp6Osrc (6), src pro-teins which contain an alteredcarboxy
terminus and have reducedtyrosine protein
kinase activity are alsounderphosphorylated
(Fig. 1B) on both serine andtyrosine(Bryant, unpublished
observations).Ourobservations that the alteration of thecarboxy termi-nus of
pp60vsrc
has adeleterious effect on cell transforma-tion as well asprotein
kinaseactivity
suggests that thecarboxy-terminal portion
ofpp60vsrc
is involvedinmaintain-ing
theoveralltertiary
structure oftheenzymatically
active srcprotein. Gentry
etal.(21)
haveobserved thatanantibody
to a
pentadecapeptide corresponding
insequence to residues 498 to 512 of the src protein inhibits pp6Osrc protein kinaseactivity.
These data are consistent with the view that thecarboxy-terminal portion
ofthe srcprotein
isimportant
inmaintaining
the structure ofa functionalcatalytic
domain. We have considered thequestion
ofwhether the structuralchanges
manifest inpp60csrc (56)
alter thefunctionalactivity
of the c-src
protein.
In agreement with Iba et al. (29), we found that viruscontaining
a chimeric src geneencoding
residues 1 to 431 ofv-src and residues 432 to 533 ofc-src
induced the transformation of chicken cells, albeit more
slowly
than didwild-type
RSV. Todate,
we have been unable to demonstrate any clearbiological
or biochemical differences between thesrcproteins
encodedby
thehybrid
and
wild-type
src genes. Our observations appear to differ from those ofShalloway
et al.(49),
whoreported
that asimilar
hybrid
src gene wasinefficientin the transformation of mouse 3T3 cells. Transfection of mouse 3T3 cells withpVCsrc
DNA hasconsistently yielded
foci of transformedcells,
butadetailedcharacterization ofcelllines established fromsuch foci remains tobe carried out.ACKNOWLEDGMENTS
We thankBetty Creasy, Amy Coffman, andRenee Renaud for excellenttechnicalassistance and JonMorganandMarkHeaneyfor many helpful discussions. We are particularly indebted to Sarah Parsonsforprovidingthe antiserumtopp6OsrcandtoRay Erikson for providing the antibody to the 36K
protein.
The c-src plasmidpSVOHCS11wasgenerously provided byTonaGilmer.
J.T.P. is a recipient of a Faculty Research Award from the American Cancer Society. This investigation was supported by PublicHealth Service grants CA29243 and CA27578 awardedbythe National Cancer Institute and grant NP-462G from the American CancerSociety.
LITERATURE CITED
1. Bishop,J. M.1983. Cellularoncogenesand retroviruses.Annu. Rev.Biochem. 52:301-354.
2. Brugge,J.S.,E.Erikson,and R. L. Erikson.1981.Thespecific interaction ofthe Rous sarcoma virus transforming protein, pp6Osrc,withtwocellular proteins. Cell25:363-372.
3. Brugge, J. S., and R. L. Erikson. 1977. Identification of a transformation-specific antigen induced by an avian sarcoma virus. Nature (London)269:346-348.
4. Bryant, D.,and J. T. Parsons. 1982. Site-directed mutagenesis of the src gene of Rous sarcoma virus: construction and characterizationof adeletion mutanttemperature sensitive for transformation.J. Virol. 44:683-691.
5. Bryant, D., and J. T. Parsons. 1983.Site-directedpoint mutation inthe src geneofRoussarcomavirus results inaninactive src geneproduct. J.Virol. 45:1211-1216.
6. Bryant, D. L.,andJ.T. Parsons. 1984. Amino acidalterations withinahighly conserved region of theRous sarcomavirussrc geneproduct pp6Osrc inactivate tyrosine protein kinase activity. Mol. Cell. Biol. 4:862-866.
7. Buss, J. E.,M.P.Kamps, and B. M. Sefton.1984.Myristicacid isattachedtothe transforming protein ofRous sarcomavirus during or immediately after synthesis and is present in both soluble and membrane-bound forms of the protein. Mol. Cell. Biol.4:2697-2704.
8. Buss, T., and B. M. Sefton.1985.Myristic acid,a rarefatty acid, isthelipid attachedtothetransforming protein ofRous sarcoma virusand its cellularhomolog.J. Virol.53:7-12.
9. Collett,M. S.,E.Erikson, and R. L. Erikson. 1979. Structural analysis of the avian sarcomavirustransforming protein: sites ofphosphorylation. J. Virol. 29:770-781.
10. Collett,M.S.,and R. L. Erikson. 1978.Protein kinase activity associated with the aviansarcomavirussrc geneproduct.Proc. Natl. Acad. Sci. U.S.A.75:2021-2024.
11. Collett,M. S., A. F. Purchio, and R. L. Erikson. 1980. Avian sarcoma virus-transforming protein, pp6fisrc, shows protein kinase activity specific for tyrosine. Nature (London) 285: 167-169.
12. Cooper, J. A., and T. Hunter. 1981. Changes in protein phos-phorylation inRous sarcomavirus-transformed chicken embryo cells. Mol. Cell.Biol. 1:165-178.
13. Cooper, J. A.,N. A.Reiss,R.J.Schwartz,and T. Hunter.1983. Threeglycolyticenzymes arephosphorylatedattyrosine in cells transformed by Rous sarcoma virus. Nature (London) 302:218-223.
14. Courtneidge, S.,andJ. M. Bishop. 1982. Transitofpp60v-srcto theplasma membrane. Proc.Natl.Acad. Sci. U.S.A. 79:7117-7121.
15. Courtneidge, S.,A. D. Levinson, andJ. M. Bishop. 1980.The protein encoded by the transforming gene of avian sarcoma virus (pp60rc) and a homologous protein in normal cells (pp60prwo-src) areassociated with the plasma membrane. Proc. Natl.Acad. Sci. U.S.A. 77:3783-3787.
16. Cross, F., and H. Hanafusa. 1983. Localmutagenesis ofRous
sarcomavirus:themajorsites oftyrosineandserine
phosphory-lation of p60s'' are dispensable for transformation. Cell 34:597-608.
17. Cross,F.R.,E.A.Garber,D.Pellman,andH. Hanafusa.1984. Ashortsequencein thep6osrcNterminus isrequiredforp6Osrc myristylationand membraneassociation andfor cell transfor-mation. Mol. Cell. Biol. 4:1834-1842.
18. Dagert, M., andS. D. Ehrlich. 1979. Prolonged incubation in calcium chlorideimproves the competence of Escherichia coli cells. Gene 6:23-28.
19. Erikson, E.,and R. L.Erikson. 1980. Identificationofacellular proteinsubstrate phosphorylated by theavian sarcoma virus-transforminggeneproduct. Cell 21:829-836.
20. Erikson, R. L., M. S. Collett, E.Erikson, and A. F. Purchio. 1979. Evidence that the aviansarcomavirus-transforminggene product isacyclic AMP-independent proteinkinase.Proc.Natl. Acad. Sci. U.S.A.76:6260-6264.
21. Gentry, L. E., L. R. Rohrschneider, J. Casnellie, and E. G. Krebs. 1983.Antibodiesto adefinedregion of pp6Osrcneutralize the tyrosine-specific kinase activity. J. Biol. Chem. 258: 11219-11228.
22. Gilmer, T. M.,and R. L. Erikson. 1981. Rous sarcoma
trans-forming protein, p60src, expressed in E. coli, functions as a
proteinkinase. Nature(London)294:771-773.
23. Gilmer, T. M., and J. T. Parsons. 1979. Analysis of cellular
on November 10, 2019 by guest
http://jvi.asm.org/
3' src GENE MUTATIONS 321 integration sites in avian sarcoma virus-infected duck embryo
cells. J. Virol. 32:762-769.
24. Guild, B. C., R. L. Erikson, and J. L. Strominger. 1983. HLA-A2 and HLA-B7 antigens are phosphorylated in vitro by Rous sarcoma virus kinase (pp60s'-r) at a tyrosine residue encoded inahighly conserved exon of the intracellular domain. Proc. Natl. Acad. Sci. U.S.A. 80:2894-2898.
25. Hanafusa, H. 1977. Cell transformation by RNA tumor viruses. p. 401-483. In: H. Fraenkel-Conrat and R. R. Wagner (ed.), Comprehensive virology, vol. 10. Plenum Publishing Corp., New York.
26. Highfield, P. E., L. F.Rafield, T. M. Gilmer, and J. T. Parsons. 1980.Molecular cloning of avian sarcoma virus closed circular DNA: structural and biological characterization of three re-combinant clones. J. Virol. 36:271-279.
27. Hunter, T., and B. Sefton. 1980. Transforming gene product of Roussarcoma virusphosphorylates tyrosine. Proc. Natl. Acad. Sci. U.S.A. 77:1311-1315.
28. Ish-Horowicz, D. 1982. Alkaline lysis method, p. 368. In: T. Maniatis, E. F. Fritsch, and J. Sambrook (ed.), Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, ColdSpring Harbor, N.Y.
29. Iba, H., T. Takeya, F. R. Cross, T. Hanafusa, and H. Hanafusa. 1984. Rous sarcoma virus variants that carry the cellular src gene instead of the viral src gene cannot transform chicken embryo fibroblasts. Proc. Natl. Acad. Sci. U.S.A. 81:4424-4428.
30. Kessler, S.W. 1975. Rapidisolation of antigens from cells with astaphylococcal protein A-antibody absorbent: parameters of theinteraction ofantibody-antigen complexes with protein A. J. Immunol. 115:1617-1624.
31. Kitamura, W., A. Kitamura, K. Toyoshima, Y.Hirayama, and M. Yoshida. 1982. Avian sarcoma virus Y73 genomesequence andstructural similarity of itstransforming gene product to that of Rous sarcoma virus. Nature (London)297:205-208. 32. Krueger, J. G., E. Wang, and A. R.Goldberg. 1980. Evidence
that the src gene product of Rous sarcoma virus is membrane associated. Virology 101:25-40.
33. Laemmli, U.K.1970.Cleavage ofstructural proteins during the assembly of the head ofbacteriophage T4. Nature (London) 227:680-685.
34. Levinson, A. D.,H.Opperman, L.Levintow, H. E. Varmus, and J. M. Bishop. 1978. Evidence that the transforming gene of avian sarcoma virus encodes a protein kinase associated with a phosphoprotein. Cell 15:561-572.
35. Little, C.D.,and W.T. Chen. 1982. Masking ofextracellular collagen and redistribution of collagen and fibronectin during matrix formation by cultured embryonic fibroblasts. J. Cell Sci. 55:35-50.
36. Macara, I., G.Marinetti, and P. C.Balduzzi. 1984. Transform-ing protein of avian sarcoma virus UR2 is associated with phosphatidylinositol kinase activity: possible role in tumorigenesis. Proc. Natl. Acad. Sci. U.S.A. 81:2728-2732. 37. Maxam, A.M., and W. Gilbert. 1980. Sequencing end-labeled
DNAwithbase-specific chemical cleavages. Methods Enzymol. 65:499-560.
38. Oppermann, H., A. D. Levinson, L.Levintow, H. E. Varmus, J.M. Bishop, and S. Kawai. 1981. Two proteins that im-munoprecipitate with the transforming protein of Rous sarcoma virus. Virology113:736-751.
39. Parker,R.C., H. E.Varmus, andJ.M.Bishop. 1984. Expres-sion ofv-src andchicken c-src in rat cellsdemonstrates qualita-tivedifferences betweenpp60v-src andpp60csrc.Cell37:131-139. 40. Parsons, J. T.,D.Bryant, V.Wilkerson, G.Gilmartin, and S. J.
Parsons. 1984.Site-directed mutagenesis of Rous sarcoma virus pp6Osrc: identification of functional domains required for
trans-formation, p. 36-42. In: G. F. Vande Woude, A. J. Levine, W. C. Topp, andJ. D. Watson (ed.), Cancer cells: oncogenes and viral genes. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
41. Parsons, S. J., D. J.McCarley, C. M. Ely, D. C. Benjamin, and J. T. Parsons. 1983. Isolation and partial characterization ofa monoclonal antibody to the Rous sarcoma virus transforming protein pp60sr'.J. Virol. 45:1190-1194.
42. Parsons, S. J., D. J. McCarley, C. M. Ely, D. C. Benjamin, and J. T. Parsons. 1984. Monoclonal antibodies to Rous sarcoma viruspp6O$r' react with enzymatically active cellular pp6Osrc of avian and mammalianorigin. J. Virol. 51:272-282.
43. Parsons, S. J., S. C. Riley, E. E. Mullen, E. J. Brock, D. C. Benjamin, W. M. Kuehl, and J. T. Parsons. 1979. Immune response to thesrcgene product in mice bearing tumors induced byinjection of avian sarcomavirus-transformed mouse cells. J. Virol. 32:40-46.
44. Radke, K., T. Gilmore, and G. S. Martin. 1980. Transformation byRous sarcoma virus: a cellular substrate for-transformation-specific protein phosphorylation contains phosphotyrosine. Cell 21:821-828.
45. Schultz, A. M., L. E.Henderson, S. Droszlan, E. A. Garber, and H.Hanafusa. 1985. Amino terminal myristilation of the protein kinase p6Osrc, a retroviral transforming protein. Science 227:425-427.
46. Schwartz, D. E., R. Tizard, and W. Gilbert. 1983. Nucleotide sequence of Roussarcoma virus. Cell32:853-869.
47. Sefton, B. M., T. Hunter, E. H. Ball, and S. J. Singer. 1981. Vinculin: a cytoskeletal target of the transforming protein of Rous sarcoma virus. Cell24:165-174.
48. Sefton, B. M., I. S. Trowbridge, and J. A. Cooper. 1982. The transforming proteins of Rous sarcoma virus, Harvey sarcoma virus and Abelson virus contain tightly bound lipid. Cell 31: 465-474.
49. Shalloway, D., P. M. Coussens, and P. Yaciuk. 1984. c-src and src homologoverexpression inmouse cells, p. 9-17. In G. F. Vande Woude, A. J. Levine, W. C. Topp, and J. D. Watson (ed.), Cancer cells: oncogenes and viral genes, Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
50. Shalloway, D., P. M. Coussens, and P. Yaciuk. 1984. Overexpressionof c-srcprotein does not inducetransformation ofNIH 3T3 cells. Proc.Natl. Acad. Sci. U.S.A.81:7071-7075. 51. Shibuya, M., and H. Hanafusa. 1982. Nucleotide sequence of
Fujinami sarcoma virus: evolutionary relationship of its trans-forming gene withtransforminggenes of other sarcomaviruses. Cell30:787-795.
52. Smart, J. E., H.Oppermann, A. P. Czernilofsky, A. F. Purchio, R. L. Erikson, and J. M. Bishop. 1981.Characterizationofsites fortyrosinephosphorylationin thetransformingprotein of Rous sarcoma virus (pp60I' rc), and its normal cellular homologue (pp60c-sr). Proc. Natl. Acad. Sci. U.S.A. 78:6013-6017. 53. Snyder, M. A., J. M. Bishop, W. W. Colby, and A. D. Levinson.
1983. Phosphorylation oftyrosine-416 is not required for the transforming properties and kinase activity ofpp6O-src. Cell 32:891-901.
54. Southern, E. M. 1975. Detection of specific sequences among DNAfragments separatedby gelelectrophoresis. J. Mol. Biol. 98:503-517.
55. Sugimoto, Y., M. Whitman, L. Cantley, and R. L. Erikson. 1984. Evidence that the Rous sarcoma virus transforming gene prod-uct phosphorylates phosphatidylinositol and diacylglycerol. Proc. Natl. Acad. Sci. U.S.A.81:2117-2121.
56. Takeya, T., and H. Hanafusa. 1983. Structure andsequence of the cellular gene homologous to the RSV src gene and the mechanism for generating the transforming virus. Cell 32: 881-890.
VOL.55,1985