JOURNAL OF VIROLOGY, Sept. 1993, p. 5692-5697 Vol. 67,No. 9 0022-538X/93/095692-06$02.00/0
Copyright© 1993, American Society for Microbiology
Conformational
Changes Affecting the
V3
and
CD4-Binding Domains
of
Human
Immunodeficiency
Virus Type 1
gpl20
Associated with
env
Processing and with Binding
of
Ligands
to These Sites
ABRAHAM PINTER,* WILLIAM J. HONNEN, AND SHERMAINE A. TILLEY
PublicHealth Research Institute, 455 First Avenue, New
York,
NewYork 10016 Received7April1993/Accepted 18 June 1993Two neutralizing human monoclonalantibodies (HuMAbs) directed against epitopes located near the tip of theV3 loop of humanimmunodeficiency virus type 1 env protein recognized solubilized gPrl60, but not gpl20, inradioimmunoprecipitation assays. Efficient immunoprecipitation of solubilizedgpl20by these antibodies did occur in the presence of HuMAb 1125H, directed against a conformational epitope overlapping the CD4-binding site, or its F(ab')2 fragment.Incontrasttothe
inability
oftheanti-V3 antibodies to immunopre-cipitate solubilizedgpl20, these HuMAbs did bind togpl20inintactvirions; this level of binding increased severalfold in the presence of theF(ab')2 fragment of 1125H. These results demonstrate that neutralization epitopesinthe V3 looparesequesteredinsolublegpl20 butpartly
exposed ingPrl60 and in virion-associatedgpl20and that binding of antibodies to the discontinuous CD4-binding site leads toconformational changes thatresult inthe exposure of V3epitopes in solublegpl20 and their enhanced
accessibility
ingPrl6O and in virion-associated gpl20. Enhanced bindingof suboptimal concentrations of 1125Htosoluble gpl20wasalso inducedby the presenceof an anti-V3 HuMAb, indicating the occurrence ofreciprocal allosteric interactions between theV3 loop and the CD4-binding site. It islikely
that these effects contribute to the synergisticneutralization of human immunodeficiency virus type 1
previously
reported for antibodies directed against these tworegions.Theenvproteinsof humanimmunodeficiencyvirus(HIV)
play essential roles in the initial phases of the infectious
cycleandaremajortargetsof the immune response. Potent neutralization epitopes have beenidentified in twodomains
of gpl20 that appear to be involved in different stages of receptorbinding and entry. The CD4-binding site ofgp120
has been shown to becomposed of discontinuous sequences, withmajor contributions from the third and fourth constant
domains, C3 and C4 (28). Monoclonal antibodies (MAbs)
that compete forbinding ofgpl20to CD4 have been previ-ously described; these include antibodies directed against
linear epitopes located within the C4 domain ofgp120that have weakneutralizing activity(1,20,36)and MAbsagainst highly conformational epitopesthat involve sites inmultiple
constant domains (25, 37) that have considerably more potent neutralizing activity (13, 31, 40). Both the epitopes recognized by the latter MAbs and the CD4-binding site
itselfaredestroyed uponreductionof disulfide bonds(13,23,
31, 40).The V3domain isnotrequired for CD4binding,but itis believedtoplayanundefined role in viralpenetration(8, 35). This domain exists as a highly immunogenic disulfide-linked loop whichacts asthe principal neutralizing domain of thevirus(16, 19, 21).Antibodiesagainstthe V3loopblock HIV-inducedsyncytiumformation andinfectivity(reviewed
in reference27),andmutations within thisregion inhibit the
fusogenicityofHIVtype1(HIV-1)env(9, 10, 14, 15, 29, 42) and affect thetropism of the virus for T cells and monocytes
(6, 14, 34,43).
Recently, we and others have reported that antibodies
againstthe V3loopandCD4-bindingsite of HIV-1gp120act
synergisticallyinneutralization ofmultiplestrains of HIV-1
(4, 24, 32, 38, 39, 41). This phenomenon has important
*Corresponding author.
implicationsforunderstanding mechanismsof viral infection and antibody-mediated neutralization in vivo and for the
developmentofpassive and activeimmunotherapies. Other evidence suggests theoccurrenceofallostericeffects
involv-ing the V3 loop and CD4-binding site of gpl20 that may relate to the cooperative activities of antibodies to these domains. Binding of soluble CD4 (sCD4) resulted in in-creasedbindingof anti-V3 antibodies to HIV-1IIIB-infected
cells (33) and virions(24) and increasedcleavage of the V3 loop on virions by exogenous proteases (7). In addition,
Wyattetal.(45)demonstratedaV3loop-dependentmasking
ofepitopesin theCD4-binding domainof HXB2gpl20in the presenceofionic detergents andan increasedbindingofan anti-V3 MAb uponintroductionofsingleamino acidchanges
in the C4 region of gpl20. In order to determine whether similar interactions between these domains occurred for other strains of HIV-1 andtoclarifythepotentialrelevance of such interactions towards synergistic neutralization by
antibodies againstthesesites, weperformed an
analysis
of thereactivity of various forms of HIV-1env proteinsfrom three strains of HIV-1 with human MAbs(HuMAbs) against
the V3loopandCD4-bindingsite thatpotentlysynergize in neutralization of HIV-1infectivity.
Threedifferent HuMAbswereused in theseexperiments.
1125H is directed against the conformational CD4-binding
site (39, 40). Its epitope is broadly conserved, and an
analysis of the reactivity of this antibody with a panel of
gp1204XB2
point mutants (37) identified sites in all five constantdomains that affectbindingofthis MAbtogpl20.4117C isan anti-V3HuMAb which recognizes the broadly
conserved sequence IXIGPGR presentnearthetipof the V3
loopof virusescontainingtheNorth American V3consensus sequence (39).Both of these HuMAbsarepotently
neutral-izing antibodies. 41148D, an anti-V3 HuMAb that reacts with multipleHIV-1 strains, including IIIB, SF2, andMN, 5692
on November 9, 2019 by guest
http://jvi.asm.org/
NOTES 5693
1 2 34 5 67
gp1
20-SF2
gplW2-
4
IIM
[image:2.567.68.261.71.195.2]MN
FIG. 1. Reactivity of HuMAbs with NP-40-solubilized HIV-1 gpl20.3H-glucosamine-labeledvirions ofthe SF2(22) and MN (11) strains ofHIV-1, grown in H9 cells, were solubilized with0.5% NP-40 and incubated withMAbs for 1 hat37°C.Immune complexes were collected with fixed S. aureus(Pansorbin; Calbiochem Inc.), washed in NP-40-containing buffer, and analyzed by SDS-PAGE (18) andfluorography (3). Thefollowingantibodieswereused:lane 1, HIV-1-seropositive human serum; lane 2, anti-CD4-binding site HuMAb 1125H; lane 3, F(ab')2 fragmentof1125H; lane 4, F(ab')2 fragmentof 1125H plus goat anti-human IgF(ab')2; lane 5, anti-V3 HuMAb4117C; lane 6, 4117CplusF(ab')2 fragment of 1125H; and lane 7, 4117C plus sCD4(obtained from Genentech, Inc.).
1 2 3 4
M.-. ..
5 6 7
[image:2.567.317.518.72.193.2]*::; ...
...
.. ..
^
^
.^.
= ....;r.
_ ..:..._.
_ :E57
w :e.... P.;.0.'..,.:
gPrl 60-
gpl20-gPrl 60-
gpl20-370
00
FIG. 2. Analysis of reactivity of cell-associated HIV-1env pro-teins with HuMAbs. H9 cells infected withHIV-1m werelabeled with3H-glucosamine for 4 h and lysed with0.5% NP-40 in phos-phate-buffered saline. Aliquots of cell lysate were incubated with HuMAbs ateither 37 or0°C, as indicated, andimmunoprecipitates werecollected and analyzed by SDS-PAGE. Thefollowing antibod-ies were used: lane 1, HIV-1-seropositive human serum; lane 2, 1125H; lane 3, F(ab')2 fragment of 1125H; lane 4, anti-V3 HuMAb 4117C;lane 5, 4117C plusF(ab')2 fragment of 1125H; lane 6, anti-V3 HuMAb41148D; andlane 7, 41148D plus plus F(ab')2fragmnentof 1125H.
has a less potent neutralizing activity. The epitope
recog-nizedby this MAbwas determinedaspreviously described
(39)tobe KRIHIGP for the MNsequence.
Differentialreactivity ofHuMAbs with solublegpl20. The ability oftwoHuMAbstorecognize gp120 solubilized from HIV virions of theSF2 and MN strainswas determinedby
radioimmunoprecipitation analysis of 3H-glucosamine-la-beledgp120 presentinNonidet P-40 (NP-40)-treated
super-natants ofinfected H9 cells (Fig. 1). A polyclonal human
serumefficiently precipitated gp120s from both strains under
these conditions (lane 1). Anti-CD4-binding site HuMAb 1125H also recognized both MN and SF2 gp120 (lane 2). In contrasttothis, equivalent levels of
4117C,
knownto react withhighaffinity with V3 peptides of bothMNenvand SF2env and to efficiently neutralize both viruses (39), did not precipitate gpl20 molecules from either of thesetwostrains (lane 5).
In ordertoexamine the effect of theanti-CD4-bindingsite HuMAb 1125H on theability of 4117C torecognize gpl20,
an F(ab')2 fragment of 1125H was prepared by pepsin
digestion. This resulted inahomogeneous product that did
not bind to Staphylococcus aureus that contained a trun-cated heavy chain and an intact light chain, as shown by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).Asexpected, the 1125H F(ab')2 fragmentwas
unable to immunoprecipitate labeled gp120 with S. aureus
(Fig. 1, lane 3). However, the F(ab')2 fragment did bindto gpl20, as demonstrated by the efficient precipitation of
labeled gp120 when goat anti-human F(ab')2 serum was
added priorto S. aureus (lane 4). When the 1125HF(ab')2
fragment and anti-V3 MAb 4117C were both present, effi-cient precipitation of MN and SF2 gpl20s was observed
(lane 6). Binding of the anti-V3 HuMAb togp120 was not detectedwhen sCD4wasused inplace of the 1125HF(ab')2
fragment (lane 7). These results suggested that binding of the
anti-CD4-binding site MAb fragment, but not sCD4, to solubilized gp120 led to a conformational change in gp120
that resulted in the exposure of a previously cryptic V3
epitope and consequent binding of 4117C. Similar results
were obtained when soluble gp120, present in clarified
supematants of infected cells, was immunoprecipitated by these antibodies in thecomplete absence of NP4O(data not
shown), indicating that these effects were not artifacts in-ducedby thepresence ofdetergent.
To obtain additional insight into the ability of anti-V3 antibodies to bind todifferent forms of HIV envprotein,the
reactivitiesofdifferent HuMAbs withenvproteinspresent in
lysatesofHIV-infectedcells weredetermined(Fig. 2).Since the extentofbinding of gpl20tosCD4has beenreportedto change with temperature (26), theseexperimentswere
per-formed at both 0 and 37°C. The
anti-CD4-binding
site HuMAb 1125H reactedequallywell withgPrl60andgp120(lane 2), while the anti-V3 HuMAbs 4117C and 41148D were able toimmunoprecipitate gPrl60but notgpl20 (lanes4 and
6). As wasobserved for soluble extracellular gp120, in the presence of the 1125HF(ab')2 fragmentbothanti-V3 HuM-Absgainedthe abilitytoimmunoprecipitate cell-associated gp120, and a modest enhancement in their reactivity with
gPrl60 was also apparent (lanes 5 and 7). Similar effects wereobservedatboth0and37°C.
Binding studies with recombinant gpl20 and gpl60 of
HIIVB.
Mostprevious
biochemicalanalyses
ofgp120
havebeenperformedwithproteins derived from strainsrelatedto
theoriginalLAIisolate. The V3loopsof thesegpl20sdiffer
fromthe moretypical MN-like isolatesby the insertion ofa
dipeptide, Gln-Arg, nearthe centerof theV3loop,and this
change has a profound effect on the
immunoreactivity
of thesegp120s. In orderto seewhetherthe effects describedabove alsooccurredforLAI-typeenvproteins,
immunopre-cipitation analyses
wereperformed
with recombinant forms ofgpl20 and gp160(rgpl20
andrgpl60)
derived fromLAI-related strains. rgpl20 was derived from the IIIB-derived clone pv.22 and expressed in CHO cells
(2). rgpl60
wasderivedfromtheBRUstrain and contains deletions ofboth
thegpl20-gp41
cleavage
siteandthehydrophobic
transmem-brane domain of
gp4l (17).
Bothmoleculesefficiently
bindCD4,
andthusthey
possessrelatively
nativeconformations.These molecules were radioiodinated
(30)
and incubatedwith biotinylated anti-V3 HuMAb 41148D, and immune
complexes were
precipitated
withstreptavidin-agarose
beads. Under these
conditions,
thisantibody
didnot precip-VOL.67, 1993on November 9, 2019 by guest
http://jvi.asm.org/
5694 NOTES
1 1
25H
0 1.6 0.4 0.1 0.025
CD4-Ig
1.6 0.4 0.1 0.025 jAg/ml
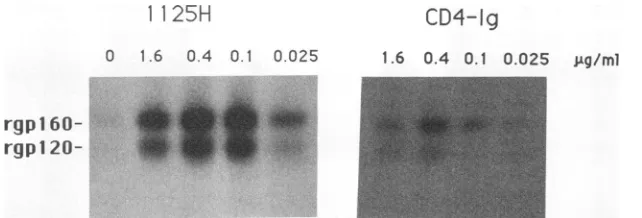
FIG. 3. Effects of HuMAb 1125H and sCD4-Igonreactivity of rgpl2O and rgpl60 with anti-V3 HuMAb 41148D. 125I-labeled rgpl20 and
rgpl60wereincubated with biotinylated 41148D (0.5 p,g/ml)in thepresenceof the indicated concentrations of either 1125HorsCD4-Ig(5).
Immunecomplexeswerecollected with streptavidin-agarose beads and analyzed by SDS-PAGE and fluorography.
itatergpl20 and reacted only minimally withrgpl60 (Fig. 3). In the presence of various concentrations of intact 1125H,
efficient precipitation of both rgpl20 and rgpl60 was
in-duced. Asimilar, thoughlessefficient, effectwasobtained in
the presence of soluble CD4-immunoglobulin (Ig)
immu-noadhesion (5). The concentrations of 1125H and CD4-Ig requiredtoprecipitate the maximumamountof recombinant proteinswere similar, inthe range of0.1 to 0.4 ,ug/ml. No streptavidin precipitation of labeled proteinwasdetectedfor
the unmodified 1125HorCD4-Ig alone (not shown).
Consis-tent with the results of the experiment described in the legend to Fig. 1, sCD4 itself did not enhance binding of 41148D to the recombinant env proteins (not shown). This
difference inactivityofsCD4 andCD4-Igmaybe relatedto either the largersizeordimericstructureofCD4-Ig.
Enhancement ofbindingof anti-V3 MAbstointact virions by antibodytotheCD4-bindingsite. The inabilityofanti-V3 HuMAbs 4117C and 41148D to immunoprecipitate soluble gpl20wasunexpectedinview of theneutralizing activityof these antibodies. One possible explanation for this effectis that, in contrast tothe sequesteredconformation of the V3 loop in soluble gpl20, this region is accessible in virion-associated gpl20. To test this possibility, we performed
similar immunoprecipitation experiments with these HuM-Abs usingintact virions.
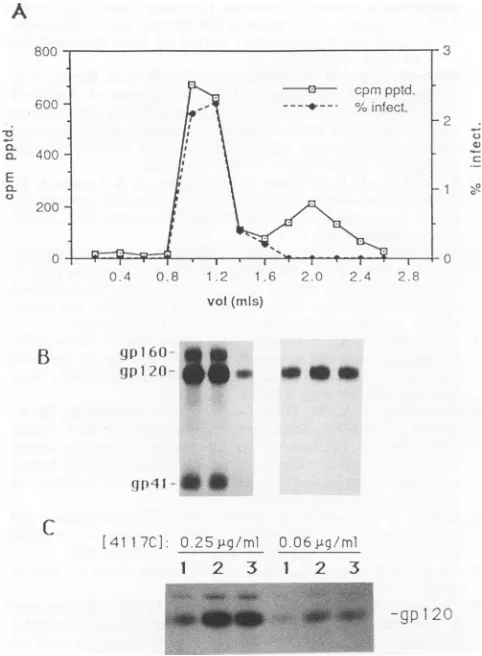
To separate intact virions from soluble gpl20, labeled culture supernatants were fractionated on a Sephacryl
S-1000 gel permeation column. This resulted in two peaks containingviralglycoproteins,asassayed by
immunoprecip-itation withan immune humanserum. Infectious viruswas
associated with the first peak (Fig. 4A), and this peak containedgPrl60, gpl20,andgp4l (Fig. 4B),consistentwith the presence of intact viral membranes. The second peak containedonlygpl20 and thus apparently correspondedto soluble protein that had fallen off the virus. The abilityof anti-V3 HuMAb 4117C to immunoprecipitate gpl20 from intact virions present in the first peakwas determined for
twodifferentconcentrations of 4117C (Fig. 4C).Quantitation of immunoprecipitated bands was performed by scanning
fluorographs with an Appligene CCD imaging system and integrating bands with the image analysis program NIH
Image. 4117C was able to immunoprecipitate a fraction of
the gpl20 from intact virus, and the amount of gpl20 immunoprecipitatedincreased two- to fourfold in the pres-enceof0.1 to 1.0p,g of1125HF(ab')2 fragmentperml.
Enhancement of binding of suboptimal concentrations of 1125Hbyan anti-V3 HuMAb. We havepreviouslyreported anincrease inbindingof1125Htorgpl60in thepresenceof an anti-V3 HuMAb in an enzyme-linked immunosorbent
assay (39). Inorderto seewhethersimilar enhancementof binding mightoccurforsolublegp120, abindingassaywas
performed usingasuboptimal concentration of 1125H. The
ability of different concentrations of purified anti-V3 HuMAb 41148Dtoenhancebinding of biotinylated 1125Hto soluble
gp120MN
was measured by precipitation assaysperformed with streptavidin-agarose beads (Fig. 5). At the concentration ofbiotinylated1125H used in thisexperiment (approximately 50 ng/ml) very inefficient precipitation of gp120wasobserved(lane 1);thiswasincreasedsignificantly
bythe addition of anti-V3 HuMAb 41148Dataconcentration
of either 0.10 or 1.0 p,g/ml (lanes 2 and 3). As expected, unbiotinylated41148Dbyitself did notleadtoprecipitation ofgp120 bythestreptavidinbeads(not shown).This
exper-iment indicates thatbindingof antibodiestotheCD4-binding site ofgpl20 can be enhanced bythe presence of anti-V3 MAbs. This result is consistent with our previous data
showing an approximately twofold increase in binding of 1125Htorecombinantgp160in thepresenceofanti-V3MAb
4117C(39) andwith datareported byThalietal.indicatinga
similarenhancement ofbindingofF105, aHuMAbdirected against the CD4-binding site, by an anti-V3 MAb (38). In
contrast to this, McKeating etal. did not detect enhanced bindingof39.13g, aratMAbagainstarelatedepitopein the
conformationalCD4-binding site, byanotherratMAbtothe V3loop,41.1i (24).Thereasonfor this difference isnotclear and may relate to differences in the concentrations or
specificitiesof theantibodies used.
The enhancedbindingof anti-V3 MAbstoHIV-1virions
or HIV-infected cells in thepresence of sCD4 or MAbsto the CD4-binding site has previously been described for LAI-related strains of HIV-1 (24, 33). Our present demon-stration of similar effects fortwoadditional strains suggests that this is a general property of HIV-1env and therefore morelikelytopossessfunctionalsignificance.The differen-tial reactivities of the anti-V3 HuMAbs described in this studyindicate that theV3loopexists in different conforma-tions in solublegpl20,in virion-boundgpl20,andinsoluble gPrl60. Other studies have documented interactions be-tween elements of theV3loop and sites within the C2(44) and the C4 (45) domains of gpl20. Both of these constant domains influence the CD4-bindingsite and the bindingof MAbstothe CD4bindingsite(28, 37).Evidencesuggesting that the C4region is associated with sites ingp41 has also been presented, since mutations in C4 lead to a reduced affinity of the gpl20-gp41 association (12). Apossible
sce-narioaccountingforthese results is that the V3 loop inter-actswith elements oftheCD4-bindingsite. This interaction is presumably strong in soluble gp120 but weaker when
rgpl60-rgpl
20-i.
:.1 A";;;
... W.
I!,
:7 .I1.
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.145.466.76.185.2]A
~p
0~
1
2
3
2
C1
-0-4 0.8 1.2 1.6 2.0 2.4 2.8
Vol(mIs)
B
l)160.qPI20- _-0 f 4
qp4i
-0Sf
C
[41 17C] 0O 25kg/mi. 006ig/ml
l 2 3 l 2 3
FIG. 4. Reactivityofanti-gpl20HuMAbswithpurified virions. (A) Supernatant mediumofHIVmN-infectedH9 cellslabeled
over-night with 3H-glucosamine was fractionated by gel permeation chromatography, and individualfractionswereanalyzed for infec-tious virus and for radioactivityimmunoprecipitable with a
poly-clonal human anti-HIVserum.Twopeaks of env-containing
mate-rialweredetected, centeringat1.1and 2.0 ml. Infectious viruswas
associatedonlywith thefirstpeak. (B) SDS-PAGE analysis of the
columnfractionselutingat1.0, 1.2,and 1.4 mland1.8, 2.0, and 2.2
ml immunoprecipitated with human HIV-seropositive sera. The samplesanalyzedarealignedunder thecorrespondingpoints of the
graphinpanelA.(C)Immunoprecipitation ofenvproteinspresentin purifiedvirionswith anti-V3 HuMAb 4117 in the absenceof any
additionalantibodies(lanes 1)andwith4117Cin thepresenceof the F(ab')2 fragment of HuMAb 1125Hateither 1.0 ±g/ml (lanes 2)or
0.1 ,ug/ml (lanes 3). This analysiswasperformedat two different concentrationsof4117C,asindicated.Antibodyincubationsand S.
aureus absorptionswere performed with intactvirionsin the
ab-senceofdetergent,with NP-40addedtothefinalS. aureuswash. Immunoprecipitated env proteins were detected by fluorography
afterSDS-PAGE.
gpl20 is associatedwithgp4l, whichmayinteractwithsome
of the sameconstant domain sequences. High-affinity
anti-bodiestotheconformationalCD4-binding sitemaycompete for theseV3associationsites ingpl20, thereby resultingina
structural rearrangement that exposes epitopes near the
crownof theV3loop.
Aconsiderable body of evidence suggests a critical role
forthe V3 domain inthe fusion step leadingto viral
[image:4.612.408.470.81.175.2]pene-tration (27). The data presented in this paper and in the previous studies showingenhancedexposureof theV3loop
FIG. 5. Enhancementof 1125H binding to soluble
gpl20mN
by anti-V3HuMAb.Biotinylated 1125Hat aconcentration suboptimal for immunoprecipitation (50 ng/ml) was incubated with 3H-glu--o cosamine-labeled soluble gp120 in the absence or presence of 41148D, andcomplexed gpl20wasprecipitated with streptavidin-agarose beads and analyzed by SDS-PAGE and fluorography. Concentrationsof added41148Dwere0(lane1), 0.1 (lane 2), and1.0 (lane3) p,g/ml.upon binding of ligands to the CD4-binding site of gpl20
suggest a model in which the fusogenic activity of the V3 loop is modulated by its intramolecular association with
other domains ofgpl20. When virionsbind to cellsurface CD4, aconformational rearrangement whichfullyactivates thefusogenic potential of the V3 domain occurs,resultingin viral penetration and subsequent infection of target cells. The relatively inefficient exposure of the V3 domain in virion-associated gpl20prior toits activation may serve to limit thecytopathogenicityof HIVenvproteins in inappro-priate cellular locations. Another effect of the sequestering
of the V3 domain would be less-efficientbinding of
poten-tiallyneutralizing anti-V3antibodies to freevirions,thereby reducing the sensitivity of virions to such neutralization. This effect is perhaps balanced by the inability of soluble
gpl20 tobind such anti-V3 antibodies andthereby act as a decoy. Theability of antibodiestotheconformational
CD4-bindingsite toincreasethe exposure ofneutralizingepitopes
in the V3 loop, together with the enhanced binding of
anti-CD4-binding site antibodiesthat occurs in the presence ofantibodies to the V3 loop, presumably accounts for the
synergistic neutralization observed for combinations of these antibodies and provides further support for an en-hanced therapeutic potential of combinations of such re-agents.
Thiswork wassupported by PHSgrantsAI-23884andAI-26081, CFAR grant AI-72659, and a grant from the Aaron Diamond Foundation.
WethankMary Rachofor valuabletechnical assistance andSam Kayman for helpful discussions and for a critical review of the manuscript.
REFERENCES
1. Ardman, B., M. Koowalski, J. Bristol, W. Haseltine, and J. Sodrosid. 1990. EffectsonCD4binding ofanti-peptidesera to
the fourth and fifth conserved regions of HIV-1 gpl20. J. AcquiredImmuneDefic.Syndr.3:206-214.
2. Berman, P.W., J. E. Groopman,T.Gregory, P. R.Clapham, R. A.Weiss, R. Ferriani, L. Riddle, C. Shimasaki, C. Lucas, L. A. Lasky, andJ. W. Eichberg. 1988. Human immunodefi-ciencyvirus type 1 challenge ofchimpanzees immunized with recombinant envelope glycoprotein gp120. Proc. Natl. Acad. Sci. USA 85:5200-5204.
3. Bonner, W. M., and R. A. Laskey. 1974. A film detection method fortritium-labeled proteins and nucleic acids in poly-acrylamidegels.Eur. J.Biochem.46:83-88.
4. Buchbinder, A., S. Karwowska, M. Gorny, S. Burda, and S. Zolla-Pazner. 1992. Synergybetween human monoclonal
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.62.303.82.410.2]5696 NOTES
bodiestoHIV extendstheir effectivebiological activity against homologous and divergent strains. AIDS Res. Hum.
Retrovi-ruses8:425-427.
5. Byrn,R.A.,J.Mordenti,C.Lucas,D.Smith, S. A.Marsters, J. S. Johnson, P. Cossum, S. M. Chamow, F. M. Wurm,T. Gregory, J.E. Groopman, and D.J. Capon. 1990. Biological propertiesofaCD4immunoadhesin. Nature(London) 344:667-670.
6. Chesebro, B., K. Wehrly, J. Nishio, and S. Perryman. 1992. Macrophage-tropic human immunodeficiency virus isolates from different patients exhibit unusualV3 envelope sequence homogeneityin comparisonwith T-cell-tropic isolates: defini-tion of critical amino acids involved in celltropism. J. Virol. 66:6547-6554.
7. Clements, G. J.,M.J. Price-Jones,P. E. Stephens, C. Sutton, T. F.Schulz,P.R.Clapham, J.A.McKeating,M.0.McClure, S.Thomson,M. Marsh, J. Kay,R. A.Weiss, andJ.P. Moore. 1991.TheV3loops of theHIV-1andHIV-2surface glycopro-teins contain proteolytic cleavage sites: a possible functionin viral fusion? AIDSRes. Hum.Retroviruses 7:3-16.
8. Freed,E.O.,D.J. Myers,and R. Risser. 1991.Identification of the principal neutralizing determinant of human immunodefi-ciencyvirus type 1as afusion domain. J. Virol. 65:190-194. 9. Freed,E. O., and R. Risser. 1991. Identification of conserved
residues in the humanimmunodeficiencyvirus type 1principal neutralizing determinant thatareinvolved infusion.AIDS Res. Hum.Retroviruses7:807-811.
10. Grimaila,R.J., B. A. Fuller, P. D. Rennert, M. B. Nelson, M.-L. Hammarskjold,B.Potts,M.Murray, S.D.Putney,and G.Gray. 1992. Mutations in theprincipal neutralization determinant of humanimmunodeficiency virustype 1affectsyncytium forma-tion, virus infectivity, growth kinetics, and neutralization. J. Virol.66:1875-1883.
11. Gurgo, C., H. Guo, G. Franchini,A.Aldovini, E. Collalti,K. Farrell,F.Wong-Staal,R.C.Gallo,and M. S.Reitz,Jr. 1988. Envelope sequences of new United States HIV-1 isolates. Virology164:531-536.
12. Helseth, E., U. Olshevsky,C. Furman,andJ. Sodroski. 1991. Human immunodeficiency virustype 1gpl20envelope glyco-protein regions important forassociationwith thegp4l
trans-membraneglycoprotein.J. Virol. 65:2119-2123.
13. Ho,D. D., J.A.McKeating,X. L.Li,T. Moudgil,E.S.Daar, N.-C.Sun,andJ.E. Robinson.1991.Conformationalepitopeon
gp120importantinCD4binding and human immunodeficiency virustype 1 neutralization identifiedby ahuman monoclonal antibody.J. Virol.65:489-493.
14. Hwang,S.S.,T.J. Boyle,H. K.Lyerly,and B.R.Cullen.1991. Identification of theenvelopeV3loopastheprimary
determi-nantof celltropismin HIV-1. Science 253:71-74.
15. Ivanoff, L.,D. Looney,C.McDanal, J.Morris,F.Wong-Staal, A.Langlois, S.Petteway,and T. Matthews.1991. Alterationof HIV-1 infectivity and neutralization by a single amino acid replacementinthe V3loopdomain. AIDS Res. Hum.
Retrovi-ruses7:595-603.
16. Javaherian, K.,A.J. Langlois, C. McDanal, K. L.Ross, L. I. Eckler,C. L.Jellis,A. T.Profy,J.R.Rusche,D. P.Bolognesi, S. D.Putney, and T.J.Matthews. 1989. Principal neutralizing domain of the humanimmunodeficiencyvirus type 1envelope protein. Proc.Natl. Acad.Sci. USA86:6768-6772.
17. Kieny, M. P., R. Lathe, Y. Riviere, K. Dott, D. Schmitt, M. Girard,L.Montagnier,andJ. Lecocq. 1988. Improved antige-nicityof the HIVenvprotein by cleavagesite removal. Protein Eng.2:219-225.
18. Laemmli,U.K.1970.Cleavageof structuralproteins during the assemblyof the head ofbacteriophage T4. Nature (London) 227:680-685.
19. LaRosa, G. J., J. P. Davide, K. Weinhold, J. A.Waterbury, A. T.Profy, J.A.Lewis,A.J.Langlois,G. R.Dreesman,R.N. Boswell,P.Shadduck,L. H.Holley,M.Karplus,D. P.Bolognesi, T.J. Matthews,E. A.Emini,andS. D.Putney.1990. Conserved sequence and structural elements in theHIV-1 principal
neu-tralizingdeterminant. Science249:932-935.
20. Lasky, L. A., G. Nakamura, D. H. Smith, C. Fennie, C.
Shimasaki, E. Patzer, P. Berman, T.Gregory,and D.J.Capon. 1987. Delineation of a region of the human immunodeficiency virus type 1gpl20 glycoprotein critical for interaction with the CD4 receptor. Cell 50:975-985.
21. Leonard, C. K., M. W. Spellman, L. Riddle, R. J. Harris, J. N. Thomas, and T. J. Gregory. 1990. Assignment of intrachain disulfide bonds and characterization of potential glycosylation sitesof the type 1 recombinant human immunodeficiency virus envelope glycoprotein (gpl20) expressed in Chinese hamster ovarycells. J. Biol. Chem. 265:10373-10382.
22. Levy, J. A., A. D. Hoffman, S. M. Kramer, J. A. Landis, J. M. Shimabukuro, and L. S. Oshiro. 1984. Isolation of lymphocyto-pathic retroviruses from San Francisco patients with AIDS. Science 225:840-842.
23. McDougal, J. S., J. Nicholson, G. Cross, S. Cort, M. Kennedy, and A. Mawle. 1986. Binding of the human retrovirus HTLV-III/LAV/ARV/HIV to the CD4 (T4) molecule: conformation dependence, epitope mapping, antibody inhibition, and poten-tial foridiotypic mimicry. J. Immunol. 137:2937-2944. 24. McKeating, J.A., J. Cordell, C. J. Dean, and P. Balfe. 1992.
Synergistic interaction between ligands binding to the CD4 bindingsite and V3 domain of humanimmunodeficiencyvirus type Igpl20. Virology191:732-742.
25. McKeating, J. A., M. Thali, C. Furman, S. Karwowska, M. K. Gorny,J.Cordell,S.Zolla-Pazner, J.Sodroski,and R. A.Weiss. 1992. Amino acid residues of the human immunodeficiency virus type 1 gpl20 critical for the binding of rat and human neutralizing antibodies capable of blocking the gpl2O-sCD4 interaction. Virology190:134-142.
26. Moore, J. P., and P. J. Klasse. 1992. Thermodynamic and kineticanalysis ofsCD4bindingtoHIV-1virionsand ofgpl20 dissociation. AIDSRes. Hum.Retroviruses8:443-450. 27. Moore, J. P., and P. L. Nara. 1991.The role of theV3loop of
gpl20in HIV infection.AIDS 5:S21-S33.
28. Olshevsky,U., E. Helseth, C. Furman, J. Li, W. Haseltine, and J. Sodroski. 1990. Identification of individual human immuno-deficiencyvirus type 1 gpl20 amino acids important forCD4 receptorbinding.J.Virol.64:5701-5707.
29. Page, K. A., S. Stearns, and D. R.Littman. 1992.Analysis of mutations in the V3 domain of gpl60 that affect fusion and infectivity.J. Virol.66:524-533.
30. Pinter, A., and W. J. Honnen.1988. Asensitive radioimmuno-precipitationassayforhumanimmunodeficiencyvirus(HIV).J. Immunol.Methods112:235-241.
31. Posner, M. R., T. Hideshima, T. Cannon, M.Mukherjee,K.H. Mayer, and R. A. Byrn. 1991. An IgG human monoclonal antibodythatreactswithHIV-1/gpl20,inhibits virusbindingto
cells,andneutralizes infection. J.Immunol.146:4325-4332. 32. Potts, B. J., K. G. Field, Y. Wu, M. Posner, L. Cavacini, and M.
White-Scharf.Synergistic inhibition of HIV-1 by CD4 binding domain reagents and V3directedmonoclonalantibodies. Virol-ogy, in press.
33. Sattentau,Q.J., and J. P. Moore. 1991. Conformationalchanges induced in the humanimmunodeficiencyvirusenvelope glyco-protein bysoluble CD4binding.J. Exp.Med. 174:407-415. 34. Shioda, T., J.A.Levy,andC.Cheng-Mayer. 1992. Small amino
acidchangesin the V3hypervariable region of gpl20canaffect theT-cell line andmacrophage tropismof human immunodefi-ciencyvirus type 1. Proc. Natl. Acad.Sci.USA89:9434-9438. 35. Skinner,M.A.,A.J. Langlois, C. McDanal, J. S. McDougal, D. Bolognesi, andT.J.Matthews. 1988.Neutralizingantibodies to
animmunodominantenvelope sequence do notpreventgpl20 bindingtoCD4. J.Virol.62:4195-4200.
36. Sun, N., D. D. Ho, C. R. Y. Sun, R. Liou, W. Gordon, M. S. C. Fung, X.Li, R. C. Ting, T. Lee, N. T. Chang, and T. Chang. 1989.Generationandcharacterization of monoclonal antibodies
to the putative CD4-binding domain of human immunodefi-ciencyvirus type 1gpl20.J. Virol. 63:3579-3585.
37. Thali, M., C. Furman, D. D. Ho, J. Robinson, S. Tilley, A. Pinter, and J. Sodroski. 1992. Discontinuous, conserved
neu-tralizationepitopes overlapping the CD4-binding region of the human immunodeficiencyvirus type 1 gpl20envelope glyco-protein.J. Virol.66:5635-5641.
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
38. Thali, M., C. Furman, B. Wahren, M. Posner, D. D. Ho, J.
Robinson, and J. Sodrosld. 1992. Cooperativity of neutralizing antibodies directedagainst the V3 and CD4 binding regions of the human immunodeficiency virus gpl20 envelope glycopro-tein. J. Acquired Immune Defic. Syndr. 5:51-59.
39. Tilley, S. A., W. J. Honnen, M. E. Racho, T. C. Chou, and A. Pinter. 1992. Synergistic neutralization of HIV-1 by human monoclonal antibodies against the V3 loop and the CD4 binding site ofgpl20.AIDS Res. Hum. Retroviruses 8:461-467. 40. Tilley, S. A., W. J. Honnen, M. E. Racho, M. Hilgartner, and A.
Pinter. 1991. A human monoclonal antibody against the CD4-binding site of HIV-1gpl20 exhibitspotent,broadly neutralizing activity. Res. Virol. 142:247-259.
41. Tilley, S. A., W. J. Honnen, M. E. Racho, M. Hilgartner, and A. Pinter. 1991. Human monoclonal antibodies against the putative CD4binding site and the V3 loop of HIV gpl20actinconcert to
neutralize virus, abstr. 70. VII International Conference on
AIDS, Florence, Italy.
42. Travis,B., T. Dykers, D.Hewgill,J.Ledbetter,T. Tsu,S.Hiu,
andJ. Lewis. 1992. Functional roles of the V3 hypervariable region of HIV-1 gpl60 in the processing of gp160 and in the formation ofsyncytia in CD4+ cells. Virology 186:313-317. 43. Westervelt, P., D. B. Trowbridge, L. G. Epstein, B. M.
Blum-berg, Y. Li, B. H. Hahn, G. M. Shaw, R W. Price, and L.
Ratner. 1992. Macrophage tropism determinants of human im-munodeficiency virustype1 in vivo. J. Virol. 66:2577-2582. 44. Willey, R. L., E. K. Ross, A. J. Buckler-White, T. S. Theodore,
and M. A. Martin. 1989. Functional interaction ofconstantand variable domains of human immunodeficiency virus type 1 gpl20.J. Virol. 63:3595-3600.
45. Wyatt, R., M. Thali, S. Tilley, A.Pinter, M. Posner, D. Ho, J. Robinson, and J. Sodroski. 1992. Relationship of the human immunodeficiency virustype 1gpl20 third variable loop to a
component of the CD4 binding site in the fourth conserved region. J. Virol. 66:6997-7004.