Copyright 01975 American Society for Microbiology Printed inU.SA.
Heat-Stable Variant
of
Human
Adenovirus Type
5:
Characterization and Use
in
Three-Factor Crosses
C. S. H. YOUNG1 * AND J. F. WILLIAMS
Medical Research Council, Virology Unit, Institute of Virology,GlasgowGll5JR,Scotland Receivedforpublication20January 1975
A variant ofhuman adenovirus type 5 which is heat stable (hs) invitro has
beenisolatedfollowingthreeroundsofheat inactivation at 52 C.Thevariant is
geneticallystable, both throughvegetative viral passageandthrough
recombina-tion into othergenetic backgrounds, which suggests that it arises from asingle
mutation.Three-factor crosses, using this mutantinconjunctionwith previously
described temperature-sensitive mutants, suggest the hs mutation lies near the
left-handend of the genetic map. The mutant has beenusedtodemonstratethe
production of reciprocal recombinants in two-factor crosses. The mutational
lesion isunknown, but phenotypicmixing occurs inhs x hs+ infections, which
suggests that it lies in a gene specifying a virion structural protein. Other
biological parameters examined have shownno differences from thewild-type hs+.
A set oftemperature-sensitive(ts) mutants of
human adenovirus type 5 (Ad5), isolatedinthis
laboratory (16), has been partially
character-ized by complementation analysis (17) and
examinedphysiologically intermsofthe
struc-tural antigens (8), polypeptides (9), and viral
DNA(14)made in HeLa cells on infectionatthe
restrictive temperature. These and other
inves-tigations(3) arebeginningtogive an estimate of
the number of genes in this virus and an
indication of their functions. Knowledge of the
relative positions of the genes on the genome
will be important in understanding how their
functions areco-ordinated duringviral
replica-tion. Accordingly, the ts mutants have been
crossed to obtain recombination data from
which a genetic map hasbeenconstructed(J.F.
Williams, C. S. H. Young, and P. E. Austin, Cold Spring Harbor Symp. Quant. Biol., in
press).
Inany set oftwo-factor crosses, good
additiv-ity of recombinant frequencies leads to an
unambiguous genetic map, but such additivity
isnotfound in allcrosses, especially for markers
which are close together, so that the gene order
is to some extent uncertain. The uncertainty
canbe resolvedif anadditional third markeris
used in the cross along with the two markers
being mapped. This approach has been used
with poliovirus (2) and herpes simplex virus
type 1 (1). Theoretically, it could be appliedto
the adenoviruses, where, in addition tots
mu-'Presentaddress:DepartmentofMicrobiology, Collegeof PhysiciansandSurgeonsofColumbiaUniversity,NewYork, N.Y. 10032.
tants, other classes have been described; for
example, cytocidalmutantsintype 12 (13) and
host range mutants in type 5 (12).The
observa-tion that two of our Ad5ts mutants, ts 18 and ts
19, were extremely heat labile (S. Ustacelebi,
Ph.D. thesis, University ofGlasgow, 1973)
sug-gested the possibility of searching for virion
heat-stable mutants which could be used in
classical three-factor crosses. This report
de-scribes the isolation and partial
characteriza-tionof one such mutant andillustrates the use
of this marker in asetofthree-factor crosses.
MATERIALS AND METHODS
Virus and cells. The wild-type Ad5 and the ts mutants derived from it have beendescribed previ-ously (16),ashavethe methodsfor viruspropagation andtitrationby plaqueassay inHeLacells (15).
Heat inactivation. Thekinetics ofheat inactiva-tion of virus at 52Cwas measured inthefollowing way. A 0.1-mlaliquot of virus was added to 0.9 ml of prewarmed Tris-hydrochloride buffer (pH 7.4) in a 20-mlcylindricalbottleinawaterbath. Atintervals, 0.1-ml samplesweretaken and diluteddirectly into ice-cold medium. Whencomparing the heatstability ofdifferentplaqueisolates,0.1ml ofeach isolatewas added to 0.9 ml ofprewarmedbuffer in 5-ml bottles held in arack in the waterbath,and the inactivation wasstopped at a given timebyplacing all the bottles directly in an ice-water mixture. Inactivations were carriedoutinaGrantstirringwaterbath in which the temperature could be maintainedat52.0 + 0.5C.
Recombination. The details ofsetting up mixed infections and single-parent controls have been de-scribedpreviously(17).Therecombinantfrequencyis expressed as: titer at 38.5C/titer at 32.5C x 2 x 1168
on November 10, 2019 by guest
http://jvi.asm.org/
100%.Ingeneral, cellssinglyinfectedyielded negligi-ble titers at 38.5C compared with doubly infected cells; thustherecombinantfrequencies didnothave to be corrected for reversion or leakiness in any parent. The factor of 2 intheexpression correctsfor theproductionofundetected doubletsrecombinants which areexpectedto arise with the samefrequency asthets+class.
RESULTS
Isolation of a heat-stable (hs) mutant. Selection of a heat-stable variant was made
0 2 4 6
from a temperature-sensitivemutant so that it
wouldbe possible immediately to analyze
three-factor crosses of the type tsx-hs+ x tsy-hs.
Mutant tsl, which has much the same heat
stabilityasthe wild type (Fig. 1A), was chosen
as starting material because it lay near the
middle of the two-factor genetic map as it
existed at that time. Since the location of an hs mutation was not known a priori, there was a good chance that the mutant would not be so distant from the ts marker as to be of no use in three-factor analysis.
FIG. 1. Heat inactivationofAd5wildtypeandmutants at52C. The dataareexpressedasthelogarithmof thesurviving fraction.
Minutes of inactivation
8 10 12 0 2 4
b
\ -~
tsl-hs \
VOL.15, 1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.505.102.386.211.654.2]Themutantwasisolated in thefollowingway.
Asample ofafourth-passagestock ofmutant ts
1 was inactivated at 52C to give a surviving
fraction of about 2.5%. A0.1-ml aliquotofthis
fraction was seeded on HeLa cells to obtain a
high-titer stock. A sample of this stock was
inactivatedtoabout5%,andasecondhigh-titer
stock was obtained. Finally, an aliquot from
this second stockwasinactivated, andsamples
were taken after 5, 10 and 15 min of
inactiva-tion, giving 0.4, 0.03, and 0.007% survival,
respectively. Ten plaques were isolated from
each sample, and aliquots weretested for heat
stability by heatingfor 10min at52 C. Several
plaque isolatesshowedanenhanced heat
stabil-ity compared with a tsl control, and one from
the 0.04%survivingfractionwasplaque purified
and grown in HeLa cells to give a high-titer
stock. This putative heat-stable mutant
(tsl-hs) was tested by heating at 52 C and
com-pared to tsl and Ad5 wild type. Figure 1A
shows heat inactivation curves for tsl-hs, tsl,
wild type, and tsl8; clearly tsl-hs is heat
stable, whereas tsl8 is heat labile. The heat
stability oftsl-hs remainedunchanged through
numeroussubsequent vegetative viruspassages
sothatwecanconclude thatthehsmutationis
geneticallystable.
An important question is whether the
heat-stablephenotypeis causedbyasinglemutation
or by several mutations with more or less
additive phenotypic effects. The way in which
tsl-hs was selected, which was designed to
en-hance the frequency of any pre-existing hs
mutant,couldfavour the enrichment of
"multi-ple" mutants. Repeated back-crossing of the
mutant to its parent would be expected to
reveal itsmultiplenature, but this isextremely
time consuming in adenovirus. So, we
pro-ceeded on the assumption that the hs
pheno-type results fromasingle mutation, and thatif
it does not, the fact would be revealed in
subsequenttwo-factorand three-factorcrosses.
Construction of derivative hs strains by
genetic recombination. For complete genetic
analysis by three-factorcrosses, it is necessary
tohave the thirdunselected marker in all thets mutantsoftheset tobe tested.Accordingly,the
hs marker was transferred to some of our ts mutants,thoughnotall, sincenorapidmethod was available to facilitate the transfer. To
obtain ts-hs strains, the hs marker was
trans-ferredtoawild-typebackground fromwhich it
could be transferredtoothertsmarkers. Trans-fertowild typewasfirst made in thecross
tsl-hs x tsl7-hs+ in which several of the ts+
progeny proved tobehs. One oftheseplaques
was purified, grown to high titer, and tested
again for heat stability. The new derivative was
found tobeasheatstable as the original tsl-hs
parent(Fig. 1B).This isolate was used as thehs
parent in crosses designed to transfer the
marker to various ts mutants. In such crosses the ts+-hs parent was normally in excess of the
ts-hs+ parent. The infectedcellswereincubated
at32.5C for4 days, and plaques were isolated
fromthe yield titrated at the same temperature.
Each plaque was checked for its temperature
sensitivity and, if ts, was checked for heat stability. The data are given in Table 1, which
shows clearly that it is possible to transfer hs
into a variety of ts backgrounds and that it is
notclosely linked to any of the ts markers used.
Some ofthese ts-hs plaques from the different
crosses were purified, grown to high titer, and
tested forheatstability. Inactivation curves for
two of the derivatives, ts5-hs and tsl7-hs, are
shown inFig. 1C and D. Although they appear
tobe slightly less heat stable thants 1-hs, the
difference is considered to be within the limits
of experimental variation. The extent of this
variation can be seen when the inactivation
curvesfor ts+-hs in Fig. 1B and E arecompared.
Nosegregants displayingintermediate levels of
heatstability wereobservedamong the progeny
fromany of thecrosses, so we conclude that the
hs marker may betransferred intoanumberof
geneticbackgrounds without loss of heat
stabil-ity. Theseobservationsdonotsupport the view
that the hsphenotype results from a number of
additive mutations but suggest rather that it
arises from asingle mutational lesion.
Use of the hs marker inthree-factorcross
analysis. The most ambiguous region of the
current two-factor genetic map (Fig. 2) is near
tsl, where five complementation groups are
closely linked and where also there are a large
number of individualmutants in
complementa-tion group A (Williams et al., Cold Spring
Harbor Symp. Quant. Biol., in press). Thus it
was ofconsiderable interest to determine if the
hs marker could be usedtoorderthese mutants.
Crosses were set up in the usual way, but
parents were added at a number of different
input multiplicities of infection, each parental
stock being titrated at the time of infection.
TABLrE 1. Analysis oftsprogenyfrom two-factor crossesinvolvingts-hs+ xts+-hsviruses
No. of ts No. of ts
Cross progeny which %
tested werehs
ts+-hsx ts3-hs+ 13 2 15
ts+-hs x ts5-hs+ 12 3 25
ts+-hsxtsl3-hs+ 10 1 10
ts+-hsxts17-hs+ 18 1 5.6
on November 10, 2019 by guest
http://jvi.asm.org/
HEAT-STABLEVARIANT OFADENOVIRUS
Only those crosses which had an approximate
equality of input were analyzed fully.
Well-isolated ts+ plaques were picked from assay
plates incubated at 38.5 C, and the heat
stabil-ity was compared with that of ts+ plaques
obtained from the isogenic cross in which
nei-ther parent contained the hs marker. In many
cases they were also compared with plaques
from astandard ts+-hs strain. Arepresentative
setofinactivation data is shown in Fig. 3, where
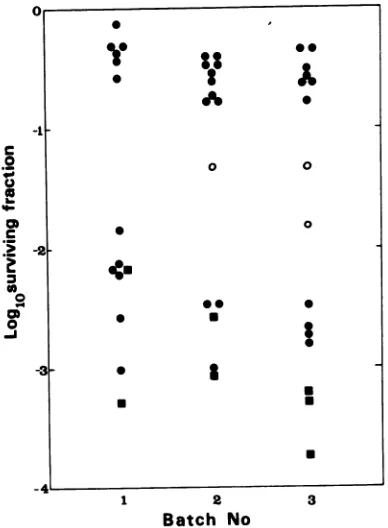
itcan be seen that the plaque isolates fall into
two clearly distinguishable categories, hs and
hs+. There are a few exceptions which, on
further analysis, prove to be mixed plaques.
Thiswill be discussed below.
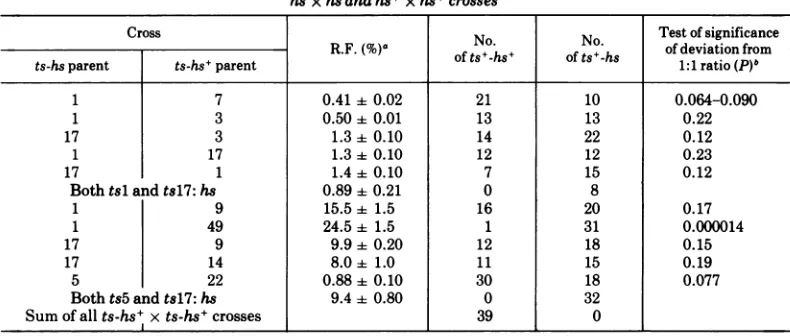
The results of crosses between ts mutants which are closely linked are shown in the top six
linesofTable2, from which it can be seen that
the ratio of hs to hs+ phenotypes among ts+
recombinantsis notsignificantlydifferent from
1.0. This istrue for mutants in the same or in
different complementation groups (line 1 and
lines 2 to 5, respectively) and for a pair of
reciprocal crosses (lines 4 and 5). Since the hs
marker cannot liebetween all the ts markers in
all possible pairwise combinations, the results
stronglysuggest that it lies a long distance away
from any of the ts markers involved. It is
important to note that the high frequency of
both hsand hs+ phenotypes amongthe
recom-binants cannot be accounted for by (i) a high
rateofmutation from hs to hs+, since the cross
tsl-hs x tsl7-hs (line 6) only yields ts+-hs recombinants,norby(ii)the reverse from hs+ to
hs, since the accumulated data for ts-hs+ x
ts-hs+ crosses
(line
13) revealonly
ts+-hs+offspring. Furthermore, the results cannot be
explained byselective enrichment for hs or hs+
progeny intheyields, since in two-factor crosses
of both ts+-hs x ts-hs+ and ts+-hs+ x ts-hs,
there is an approximate equality of the input
and output ratios ofhs/hs+ (data not shown).
In the other crosses in Table 2, mutants
spanningthetwo-factor genetic map wereused.
Inallcases except one, thereis anhstohs+ ratio
among the ts+ recombinants of approximately
1.0. The exception, cross tsl-hs x ts49-hs+
(line 8), which has a ratio which deviates
significantly from 1.0 (P = 0.000014), involves
or--I
c
.2
0
o
-c
.5
-2
-3
2 3
Batch No
FiG. 3. Data from heatinactivationofts+ plaques from the crosses ts3-hs+ x ts17-hs and ts3-hs+ x ts 17-hs+.Plaqueswereanalyzed in batchesondifferent
days. Inactivationwasfor4minat52C.Symbols:0,
Plaquesfrom ts3-hs+ x ts17-hs; *, plaques from ts 3-hs+ x ts17-hs+; 0, plaques of intermediate heat
stability whichwereprogenytested(Table 3). 30
1l
171
7
1
21°0,
0,
%, 0-76 0-6
* N 4- , to
49
3.1 3,6
11-9
I,-14 "' 30 '
10 \ 7,
8, 12 4'.1-I
54
13 9 2.2,5
6-5
[image:4.505.246.440.168.432.2]10-10
FIG. 2. Summaryofthecurrenttwo-factormap.Distancesare2xts+frequencyexpressedaspercentages.
0
y
~~~~0@*
A
o 0
* 0
* 0
* * S
0
*
a
* :
U
VOL.15, 1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.99.394.509.658.2]mutant ts49, which lies atthe left-hand end of
the two-factor genetic map. This result is
con-sistent with the view that hs itself lies atthe
left-hand end, close tobut separable fromts49.
The location ofhswould explain the
approxi-mately randomdistributionofhsamongthe ts+
recombinantsfromcrossesinvolvingmutantsin
the right-hand end of the map.
Twopointsare illustrated by thecrossts5-hs
x tsl7-hs (line 12). (i) The rate of reversion
fromhstohs+isnothigh enoughtoconfusethe
three-factorcrossresults, since all 32ts+
recom-biantsarealso hs. This extends the observation
made in the cross tsl-hs x tsl7-hs (line 6). (ii)
Although thetwo parentsaresecond-generation
derivatives from the original isolate, no ts+
recombinants of intermediate heat-stable
phe-notype segregated in the cross. If the tsl-hs phenotype resulted from the additive effects of
a number ofrandomly located mutations, two
independently constructed derivatives, suchas
ts5-hs and tsl7-hs, would be expected to
contain different sets of mutations. Hence in
recombination between these ts-hsderivatives,
either intermediate or wild type, heat-labile
phenotypes might arise by recombination,
de-pendingonthe numbersofmutations and their
linkage relationships. This expectation is not
borne out, nor is there any segregation of
true-breeding intermediates in any of the
three-factor crosses. These observations lend
support tothe view that the hsphenotype isnot causedby multiple mutation.
Mixed plaques. As mentioned earlier, a few
ts+ plaques from three-factor crosses showed
intermediate hs phenotypes, such as those
shown in thedata taken from the cross ts3-hs+
x tsl7-hs (Fig. 3). The progeny of the three
intermediate plaques from this cross were
tested. All were found to be mixed in that the
intermediate phenotype didnotbreedtrue but
segregated intwo cases into hsand hs+ and in
one case into hs+ and a further intermediate
class which was presumed to be mixed also
(Table 3). In most three-factor crosses, a few
plaque isolates were found to be slightly less
heatstable than the standard ts+-hs or slightly
more heatstablethan the standard ts+-hs+ used
(examples ofthe latter can be seen in Fig. 3,
batch 3). When tested, the progeny of these
plaques always proved to be either ts+-hs or
ts+-hs+, suggestingthat theslightinitial
devia-tions from the standard values were due to
experimental error. In no case did any isolate
with intermediate hs phenotype breed true;
[image:5.505.267.465.302.417.2]plaques were either mixed or completely hs or
TABLE 3. Analysis of progeny from plaques of intermediate heat stability from thecrossts3-hs+ x
tsl7-hs
No. ofprogeny No. of progeny No. of progeny Plaque no. plaques plaques plaques
which were which were which were ts+-hs ts+-hs+ intermediate
21 5 5 0
27 0 2 2
34 1 3 0
aThese two plaques are assumed to be mixed also
sinceother two progenyplaques were genuinely hs+. TABLE 2. Analysis of ts+progenyfrom three-factorcrossesofthe typets-hs+ x ts-hsand fromsomeisogenic
hsxhseandhs+xhs+crosses
Cross No. No. Test ofsignificance
R.F.(%)a of
ts.-hs+
of ts+-hs ofdeviationfromts-hsparent ts-hs+parent of ts-hs of ts-hs 1:1 ratio(P)5
1 7 0.41+0.02 21 10 0.064-0.090
1 3 0.50+ 0.01 13 13 0.22
17 3 1.3+ 0.10 14 22 0.12
1 17 1.3± 0.10 12 12 0.23
17 1 1.4 0.10 7 15 0.12
Both tsl and tsl7:hs 0.89 ± 0.21 0 8
1 9 15.5± 1.5 16 20 0.17
1 49 24.5 ± 1.5 1 31 0.000014
17 9 9.9 0.20 12 18 0.15
17 14 8.0 ± 1.0 11 15 0.19
5 22 0.88 0.10 30 18 0.077
Both ts5 andts17: hs 9.4 ± 0.80 0 32
Sumofallts-hs+xts-hs+crosses 39 0
aR.F.,Recombinantfrequency:2x frequency of ts+ in theyield (see Materials andMethods).The valuesare
meansoftwoduplicates performedatthesametime± theexperimental range.
The probability iscalculated from Fisher's exact testsince thenumbersofobservationsare small. The probabilityisforthe nullhypothesis that hsis notlinkedtoeithertsparent.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.70.465.460.628.2]hs+.Thisisfurther
experimental
evidencethaths is not caused
by multiple
mutations withadditivephenotypic effects.
The nature of the mixedhs/hs+ plaques is not
known, but several explanations are possible. The plaques might arise from clumps of hs and
hs+ virusparticles, but moreinteresting
possi-bilities are that they represent segregants from virions containing more than one complete genome or from partially heterozygous
struc-tures. The latter situation is foundinphage (4)
and hasbeen proposed for herpes simplex type 1
(1).At present, we cannot choosebetween these
possibilities.
Reciprocalrecombinants.For mapping pur-poses, it is usual to assume that where only one
class ofrecombinantfromatwo-factor cross can
bescored easily, as, forexample, ts+ from ts x
ts crosses, the other class is present at
equal
frequency. Thus to obtain the recombinant
frequency fromsuch a cross, the ts+ frequency is
doubled.However, the hs marker can be used to check this assumption since crosses of the type
ts-hs x ts+-hs+
(or
the reciprocals) will yieldrecombinants ts+-hs and ts-hs+, both of which
canbe detected. Accordingly, testcrosseswere setup between ts5-hsandts+-hs+, andplaques
wereisolatedfromtheyieldtitrated at 32Cand
tested for heat stability. Of the 160 plaques
tested, four were ts+-hsand three were ts-hs+. Thus there would appear to be no gross
devia-tion fromreciprocality, atleastinthis cross.
Oneofthe ts+-hsrecombinantplaqueisolates
was purified and grown to high titer, and its heat stability was compared with that of the original ts+-hs strain. No difference in the heat
stabilitieswasfound (Fig. 1E), despitethe fact
that the hs marker had been transferred from
the original ts+-hs strain to ts5 and then back
to
tsW.
In addition, no isolates of intermediatephenotype segregatedfromthets5-hs x ts+-hs+ crosses. These observations areyetfurther
evi-dence that hs iscausedby asingle mutation.
Phenotypic mixing. At present we do not
know thephysiologicalbasis for the heat
stabil-ity of the hs marker. Heat-stable mutants ofX
and the T-odd bacteriophages are known tobe
DNA deletion mutants (5-7).Inthe case of T4,
onthe otherhand, heatstabilityarises froman
alteration to a virionprotein (11). As a
biologi-cal approach to this problem we have tested
whether or not phenotypic mixing occurs in
mixedinfections ofhs andhs+ strains. Should
phenotypic mixing occur, and particles of
inter-mediate sensitivity arise, this would be strong
evidence that heat stability results from a
change in a virus structural component. Ac-cordingly, cellswereco-infectedat a low
multi-plicity with ts+-hs and at increasing
multiplici-ties with tsl-hs+. As a control, cells were
infectedwith ts+-hsandatincreasing
multiplic-ities with tsl-hs. Both sets of infections were
incubated at 32.5 C for 4 days, and the yields
were titrated at 38.5 C to score for
tsW.
Aliquotsfrom theyields were heated for 10 min at 52 C
andtitrated at 38.5 C to score for surviving ts+
progeny. As shown in Fig. 4, the effect of
including virus of wild-type heatstability is to
lower the proportion of phenotypically ts+-hs
virusparticles emerging from the mixed
infec-tion. Todetermine that this was not due to the
production of largenumbers of ts+-hs+
recombi-nants, 10 ts+ plaques were isolated from the
yield of the mixed infection in which tsl-hs+
was in greatest excess and tested for heat
stability. Nine of these plaques were ts+-hs;
thus the lowering of the proportion of ts+
heat-stable particles emerging from the cross
must result from the encoating of ts+-hs DNA
with hs+ andhs+lhs protein. This evidence for
phenotypic mixing strongly suggests that the
underlyingcauseofthe heatstability in the hs
mutant is an alteration in a virionprotein.
Otherphenotypic properties.The following
phenotypic characteristics were found to be
identical in hs and hs+ viruses: (i) ability to
transform ratembryo cells;(ii)abilitytoinduce
interferon on chick embryo fibroblasts; (iii)
frequency of ts+ recombinants incrosses ofthe
general types tsx x tsy, tsx-hs x tsy, tsx x
tsy-hs,andtsx-hs x tsy-hs; (iv)inactivationby
neutralizing antiserum to Ad5; (v) stability on
storage at -20and -70 C; (vi) abilityto form
i
-2I._
0
cm
c
:F
-td5-hs alone tsl-hs alone
tst-healone
10 20 30 40 50
Ratio of input multiplicities
FIG. 4. Heat inactivation of the yields from the mixed infections ts+-hs x ts1-hs+ and ts+-hs x
tsl-hs. Yields were assayed at 38.5C, both before and after 10-mininactivationat52 C.ts+-hswasaddedat
amultiplicity of0.1PFUpercell in allcases,whereas tsl-hs+ and tsl-hs were added at increasing multi-plicities. Symbols: 0, ts+-hs x tsl-hs+; *, ts+-hs x
tsl-hs.
U
* U
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.254.445.338.597.2]infectious centers; (vii) particle to PFU ratios (20 10);and(viii)yields ofvirus at 32C(and,
where applicable, 37 and 38C) (100 to 1,000
PFU per cell). There isno loss of, oralteration
in, any ofthewild-type functions examined.
DISCUSSION
The aim of this workwas todevelop a third
marker suitablefor inclusion in ts x ts crosses
so that unambiguous gene orders could be
deduced. This marker mustfulfiltwo essential
criteria; it should be genetically stable and
should result from a single mutation. The
phenotype of the hs mutant isolated here is
stable through many vegetative passages, and thus the first criterion is fulfilled. The marker
can be transferred to a number of different
genetic backgrounds, and genetically stable
intermediate phenotypes donot segregate from
either two- or three-factor crosses. From these
results weconclude that the second criterion is
also upheld. Other criteria, desirable but not
absolutelyessential, forathird markerarethat
thephenotype is easily scored and that the locus
is not too distant from the markers to be
ordered. These latter criteria allow us to
esti-mate, rapidly and with statistical reliability, thedistribution ofthe third unselected marker
amongtheselectedrecombinants. Withrespect
to these two criteria, the hs marker is less
useful. (i) Because of phenotypic mixing, the
yield ofa three-factor cross cannot be assayed directly by heat inactivation to measure the frequency of ts+-hs among the ts+ progeny.
Instead, the analysis depends onthe
time-con-suming process of picking ts+ plaques and
testing their heatstability. (ii)hs mapscloseto
ts49atthe left-hand endofourcurrent
two-fac-tor genetic map and is thus a considerable
distance from those markers towards the
right-hand end whose order ismostuncertain.
Conse-quently, we need to analyzea large number of
plaques from any one cross to obtain a reliable estimate of the positions of the ts markers
relativeto hs.
The hsmarkerhas been usedtoexamine the
reciprocalityofrecombinant frequenciesin
two-factor crosses and, although the data are
lim-ited,itindicatesthatthereis no grossdeviation
from reciprocality. Thus we feel justified in
doubling the ts+ frequencyin ts x ts crosses to
obtaintherecombinant frequency.
Thenatureof themutation to heatstabilityis unknown, but theobservation that phenotypic
mixingoccurs inhs x hs+ crosses suggeststhat
the lesionresultingfrom the hs mutationliesin
a virionprotein.It is known thatpentonbase is
the first protein to be expelled from thevirion
when it isheatedat 56C (10),andit is tempting
to speculate that the hs mutation lies in the
appropriate structural gene.
The successful isolationof a new type ofAd5
mutant which is stable, can be transferred to
other genetic backgrounds, and can be used in
three-factorcrossesencourages usin oursearch
forclasses ofmutantswhichmap inotherareas
of the genome. Recently, T. Harrison, in our
laboratory, has isolated a host-range mutant
which we hope will be useful in three-factor
crosses. Eventually we hope to obtain other
markers which would enable all areas of the
genome tobe mappedreliably.
ACKNOWLEDGMENTS
We should like to thank Lesley Fraser for excellent technical assistance,J. H.Subak-Sharpeforhisinterestand criticism,and P. E. AustinandE. A.C.Follett forperforming the cell-transformation assay and virus particle counts, respectively.
LITERATURE CITED
1. Brown, S.M., D. A. Ritchie,and J. H. Subak-Sharpe. 1973.Genetic studieswithherpes simplexvirustype1. The isolation oftemperature-sensitivemutants, their arrangement intocomplementationgroups and recom-bination analysis leadingto a linkage map. J. Gen. Virol. 18:329-346.
2. Cooper,P. D. 1968. Ageneticmap ofpoliovirus tempera-ture-sensitive mutants.Virology 35:584-596. 3. Ensinger, M. J.,andH.S.Ginsberg.1972.Selection and
preliminary characterization oftemperature-sensitive mutantsof type 5adenovirus.J.Virol. 10:328-339. 4. Levinthal, C. 1954. Recombination in phage T2: its
relationship to heterozygosis and growth. Genetics 39:169-184.
5. Parkinson, J. S., and R. J. Huskey. 1971. Deletion mutants of bacteriophage lambda. I. Isolation and initial characterization. J. Mol.Biol. 56:369-384. 6. Ritchie,D.A.,andF. E.Malcolm.1970.Heat-stableand
density mutants of phages Ti, T3 and T7. J. Gen. Virol. 9:35-43.
7. Rubenstein,I. 1968.Heat-stablemutantsofT5phage.I. Thephysical properties ofthe phage andtheir DNA molecules.Virology36:356-376.
8. Russell, W. C.,C. Newman, andJ. F. Williams. 1972. Characterization oftemperature-sensitive mutantsof adenovirustype5-serology.J.Gen. Virol. 17:265-279. 9.Russell, W.C., J. J. Skehel,and J. F. Williams. 1974.
Characterization oftemperature-sensitivemutantsof adenovirus type5:synthesisofpolypeptidesininfected cells. J.Gen.Virol.24:247-259.
10. Russell, W.C.,R. C.Valentine,and H.G.Pereira.1967. The effect of heat on theanatomy of the adenovirus. J. Gen. Virol. 1:509-522.
11. Streisinger,G. 1956.Phenotypic mixingofhostrange and serological specificitiesin bacteriophagesT2and T4. Virology 2:388-398.
12. Takahashi, M. 1972. Isolation of conditional lethal mu-tants(temperature sensitive andhost-dependent mu-tants) of adenovirus type5.Virology49:815-817.
on November 10, 2019 by guest
http://jvi.asm.org/
13. Takemori,N., J. L.Riggs, andC.Aldrich. 1968.Genetic studies withtumorigenic adenoviruses. I. Isolation of cytocidal(cyt)mutantsof adenovirustype12.Virology 36:575-586.
14. Wilkie, N. M., S. Ustacelebi, and J. F. Williams. 1973. Characterization oftemperature-sensitive mutantsof adenovirus type 5: nucleic acid synthesis. Virology 51:499-503.
15. Williams,J. F. 1970. Enhancement of adenovirusplaque
formation on HeLa cells by magnesium chloride. J. Gen.Virol.9:251-255.
16. Williams, J. F., M. Gharpure, S. Ustacelebi, and S. McDonald. 1971. Isolation of temperature-sensitive
mutantsofadenovirustype5. J.Gen.Virol.11:95-101.
17. Williams,J. F., and S. Ustacelebi. 1971. Complementa-tion and recombination with temperature-sensitive
mutants of adenovirus type 5. J. Gen. Virol.
13:345-348.
1175
on November 10, 2019 by guest
http://jvi.asm.org/