JOURNAL OF VIROLOGY, Feb. 1975, p. 355-362
Copyright©1975 AmericanSociety for Microbiology Printed in U.S.A.Vol. 15, No. 2
Scanning Electron Microscopic Studies of Virus-Infected Cells
I.
Cytopathic
Effects and Maturation of Vesicular Stomatitis Virus in
L2
Cells
KATHRYN V. HOLMES
Department ofMicrobiology, University of Texas Southwestern Medical School, Dallas, Texas 75235
Received for publication 12 September 1974
L2 cells infected with vesicular stomatitis virus under single-cycle conditions
have been studied by scanning electron microscopy after preparation by the
critical point drying technique. Three dimensional images of intact cells show
bullet-shapedvesicular stomatitis virus virions budding singly and in radiating
clusters both from the plasma membrane between cellular microvilli and from
the sides of microvilli. Virus-induced cytopathic effects observed by scanning
electron microscopy include intermeshing of microvilli, loss of filipodia which
attach cells to the substrate, and rounding up and detachment of infected cells
from the substrate.
The maturation ofvesicular stomatitis virus
(VSV)has been characterized in manyelectron
microscope studies of thin sections of
VSV-infectedcells (4, 7, 19-21). Longstrands of viral
nucleocapsid appear inthecytoplasm and
char-acteristicbullet-shapedvirions develop by
bud-ding from the plasma membrane, or, in some cell types (21), into cytoplasmic vesicles.
McFadden et al. (10) have estimated that as
many as10,000 virions maybudsimultaneously
from asingle cell.
In negatively stained virus preparations the
nucleocapsid strandappearsto bearrangedina
tightly coiled helix within the virion, and the
viralenvelopeiscovered withprojections10nm
longand 3.5 to 4.5nm indiameter (2, 3, 7, 12,
15). Althoughinfectious (B) virionsmeasure70
nm indiameter and175nminlength,
prepara-tions of VSV may contain noninfectious
trun-cated VSV particles which are shorter than B
virions(9).
Single-cycle growth curves of VSV in many
cell types show that after a latent period of
about 2h the titer of infectious virus increases
at an exponential rate until 8 to 10 h after
infection (21). Synthesis of cellular RNA and
protein is rapidly inhibited ininfected cells (8,
11, 16-18).Thecharacteristiccytopathiceffects
(CPE) of VSV infection are rounding of cells,
detachment fromthesubstrate, andlysis.
This communication describes a scanning
electronmicroscope (SEM) studyofVSV
matu-ration and CPE. The unique stereoscopic im-ages of the surfaces of infected cells have
re-vealed that virions bud both singly and in
radiatingclusters,virionsdevelopboth from the
plasma membrane over the cell body and from
microvilli, and that major changes in cell
sur-faceorganelles such as microvilli, filipodia, and
ruffled membranes result from virus infection.
MATERIALS AND METHODS
Cell cultures. The L2 variant of the L929 line of mouse fibroblasts, obtained from L. Sturman, New York State Department of Public Health, Albany, N.Y., was grown in monolayers on glass coverslips in Dulbecco's modification of Eagle minimal essential medium (5) with 10% heat-inactivated fetal calf serumwith 100 U of penicillin, 100
Ag
ofstreptomycin, and 0.25tsg
offungizone per ml in 5%CO2at 37 C.Virus strain. The Indiana serotype of VSV was obtained from E. Sulkin's laboratory, Southwestern Medical School, Dallas, Tex., where it had been passaged in L2 cells.
Virus inoculation. Confluent monolayers of L2 cellsonglass coverslipsin 60-mmplastic petridishes wereinoculated with 0.5 ml ofVSVat amultiplicity ofinfection ofabout 10 PFU per cell. Control mono-layers were sham inoculated with0.5ml of medium. After absorption for 1 h at 37C, the inoculum was removed and the cell monolayerswere overlaid with growth medium and incubated in 5%CO2at37C. At
intervals, monolayers of control and infected cells werefixed andprepared for SEM.
Preparation of specimens for SEM. Cells were fixed insitu onglass coverslips with glutaraldehyde
(14).Glutaraldehyde was addedtothe mediuminthe petri dishto afinal concentrationof1%,andthecells werefixed for 1 h at 37C.Inallsubsequentsteps,care was takento keep the surfaceofthe coverslipunder fluid to prevent distortion of cells. The medium containing glutaraldehydewasreplacedwith 1%
glu-taraldehyde inphosphate-bufferedsaline(6)andthe cellswereheld at4C for1 to4days.Fixedmonolayers weregraduallydehydratedafterthemethod of Ander-355
on November 10, 2019 by guest
http://jvi.asm.org/
HOLMES
son (1)inserial 15-minchangesof30, 50, 70, 95, and 100%ethanolfollowedbytwo15-mintreatmentswith
amyl acetate. In a Polaron critical point drying
apparatus, the amyl acetate was replaced byliquid
CO2. The CO2was brought tothe critical pointand
the carbon dioxide gas was released. The specimens
werethen coated with a layerapproximately 10nm thick of gold orgold andpalladiuminaMikros carbon
evaporator with a rotary coater attachment and observed withaJEOL JSM-35Uor anETEC model 1000 scanning electron microscope at 20 to 25 kV
accelerating voltage. Stereoscopic photograph pairs were taken with 60 difference in specimen tilt and observed with a'fixed focusstereoscopic viewer
(Ful-lam, Inc., Schenectady, N.Y.).
RESULTS
Morphology
ofnormal L2 cells. Insham-inoculated control monolayers, L2 cells
ap-peared as spindle-shaped or polygonal cells
(Fig. 1). Several different types of surface
pro-jections could be identified (13). The cellswere
attached to the glass substrate and to each
other bylong, rigidfilipods. Ruffledmembranes
were observed at the edges of most cells and
the cell surface was studded with curving,
cylindrical,unbranched microvilli with rounded
tips. The microvilli wereofvarying lengths but
most had a diameter ofabout 100 nm. On the
surface of some cells were
large
roundedpro-jections
which have been calledblebs. Betweenthese
projections,
the cell surface was smooth.In subconfluent
monolayers
ofuninfectedcells,
rounded
dividing
cells could be observedat-tachedtothe
coverslip
by
numerousfilipods.
Budding
of VSV virions. About 4 h afterinfection with
VSV,
numerousshort,
rigid,
bullet-shaped
projections
could be seenbud-ding
perpendicular
to the cell surface betweenthemicrovilli. Thediameteroftheshort
projec-tions was
usually
slightly
lessthan that ofthemicrovilli. These short
projections
werebe-lieved to be VSV virions
budding
from theplasma
membrane,
since at this time after infection both transmission electron micros-copy(TEM)
andinfectivity
assays indicatedthat extensive virus
budding
wasoccurring.
Also, comparable
numbersofsuch shortprojec-tions were not observed in uninfected control
cells. In
single
photographs
thepresumptive
virions
appeared
tobeofvaryinglength
becauseofthe different
angles
at whichthey
projected
from the cell. When stereoscopic images were
observed,
however,
the virionsappearedtobeofuniform
length,
suggesting
that there werefewtruncated
particles
in thispreparation.
Figure
2 shows many VSV virionsbudding
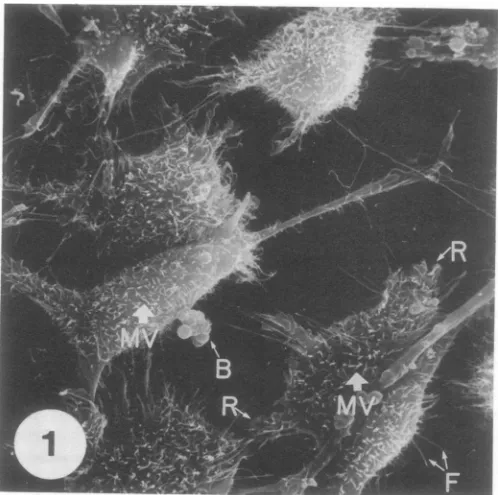
FIG. 1. Uninfected L2 cell monolayerfixed with glutaraldehyde, prepared by the critical point drying
technique, coated with gold andpalladium, and viewed by SEM. The uninfected cells are elongated or
polygonal withasmoothsurface studded withnumerous curving, cylindrical, unbranched cellularmicrovilli (MV). The cells areattached to the substrate byelongated, rigidfilipodia (F). Ruffledmembranes (R) are presentatthe cell borders. Large rounded blebs (B) areobserved on themembranesofsome cells. x2,000.
356 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.503.131.380.371.619.2]SEM STUDIES OF VIRUS-INFECTED CELLS
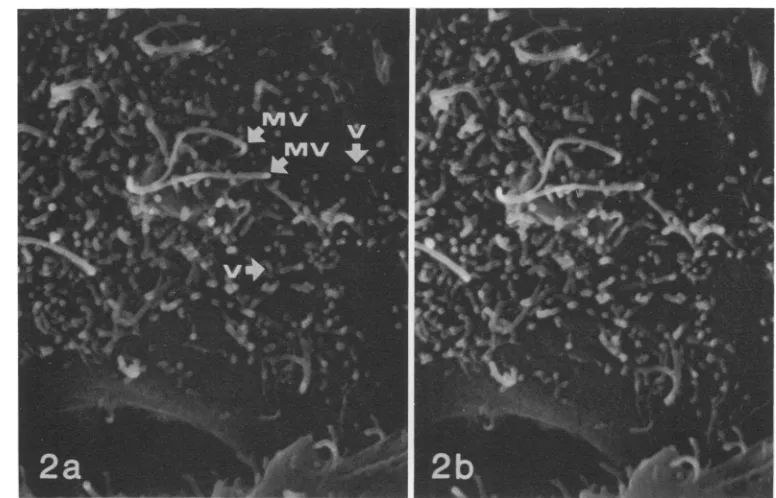
FIG. 2. VSV virions budding fromtheplasmamembraneofaninfectedL2cell8haftervirus inoculation. Thevirions (V) appearas numerousshort,rigid rods with rounded endsbuddingperpendiculartothesmooth,
flatportion ofthe plasma membrane between thelongercurvingcellular microvilli(MV).The horizontalarrow labeled Vindicates a virion budding from amicrovillus.Fig. 2aand barestereoscopic imagesphotographed
with adifference of 6° inspecimen tilt. To obtaina three-dimensional imagethey should be observed witha stereoscopic viewer. x 13,000.
from the cell surface between microvillias well
asfromthe sidesofmicrovilli at 8 h aftervirus
inoculation. Virions budding from the cellular
microvilli gave them a branching appearance
not seen in microvilli on control cells. VSV virions budded perpendicular to the plasma
membrane andwerereadily distinguished from
absorbed virions whichwereobservedin
prepa-rations held at 4 C to be attached to the cell
membranebythe sidesofthevirionratherthan
by the end. VSV virions frequently were
ob-served budding in clusters radiating from a
single point on the surface of an infected cell
(Fig. 3). Buddingclusters of two to five or more
virionshave been observed bySEM.
As thecellsbegantoroundup,the numberof
VSV virions seen budding from the plasma
membrane increased. The stereoscopicimage of
the rounded fibroblastic cell in Fig. 4makes it
possible to seethat extensive buddingofvirions
from cellular microvilli caused them to appear
branched. As the virions budded, they often
appeared to adsorb to other cellular microvilli
from which viruses could be budding, forming
an intricate network of adsorbed viruses and
cellular processesaroundthe cell. In Fig. 5 at a
higher magnification, the intimate association
ofbudding virions with microvilli can be seen
without the stereoscopic image. Unfortunately,
it was not possible to resolve viral surface
projections on the budding virions because the limit of resolution of the microscopes used in
this study was 10 nm. Unequivocal
identifica-tion of any individual surface projecidentifica-tion as a
virusrather thananunusuallyslender and short
microvillus is therefore not possible by this
technique.Virus-specific antigens might be
vis-ualized on infected cells incubated with hybrid
antibody directed against the antigens and
against avisible marker suchastobacco mosaic
virus.Thistechniquehas beensuccessfullyused
to localize antigens on the surface of
lympho-cytes observedbySEM(M.L.Lipscomb,K.V.
Holmes, E. S. Vitetta, U. Hammerling, J. W.
Uhr, submitted for publication).
Thesurface of infected cellsby17h after virus
inoculationwasalmostcompletelycovered with
budding VSV virions (Fig. 6 and 7). It seems
probable that thesevirions werearrested inthe
process of budding because little infectious virus wasreleased from infected cellmonolayers at this time. Projecting from the cells were a
small numberofmarkedlyelongated microvilli
from which clusters of VSV virions were
bud-357 VOL. 15,1975
on November 10, 2019 by guest
http://jvi.asm.org/
J. VIROL.
HOLMES
FIG. 3. Stereoscopic SEMimages oftwo VSV-infectedL2 cells showing virus-induced CPE and budding virions. AsaresultofVSVinfection, the cellatthe upper lefthas roundedupand lost most of thefilipodia whichattachedittothe substrate. The centralcell,which isbeginningtoroundup as aresult of VSVinfection, isstillattachedtothesubstratebynumerousfilipodia.The virions shownatthetipsof the arrows and in the inset(c)arebuddingin clusterswhich radiatefromasingle pointonthe cellsurface.Such multiple virus buds
arefrequentlyobservedinVSV-infectedcells. Themultiplebudsareusuallyof equallength whenseeninstereo
images. x2,500,inset x8,400.
FIG. 4. Stereoscopic SEM imagesofafibroblasticcell17hafterVSV inoculation. Numerous virions budding from the cellular microvilli give these abrushlikeappearance not seen in uninfected control cells. The budding virusesareoften absorbedto thesurface of the cellortoothermicrovilli,forming anintermeshing network of viruses and cellularprocesses around theinfected cell. Compare with control cells in Fig. 1. x2,000.
358
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.56.464.67.310.2] [image:4.503.142.378.383.625.2]SEM STUDIES OFVIRUS-INFECTED CELLS
[image:5.503.117.365.63.315.2]FIG. 5. Highmagnification of a VSV-infected cell like thatshown in Fig. 4. The numerous projections from the cellsurface include both microvilli(MV) and VSV virions (V). The microvilli are longer and are slightly larger in diameter than the virions. Virions budding from the sides of microvilli make them appear to be branching. Fine details of the virion surface suchasvirusspikes arenotresolved inthispreparation. x45,000.
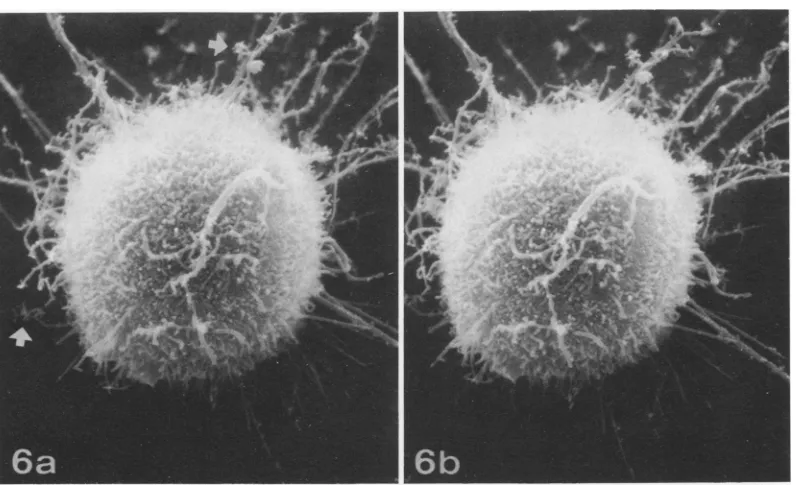
FIG. 6. Stereoscopic SEM images of VSV-infected L2 cell 17 h after virus inoculation. Theinfected cell has rounded up and is attached to the substrate by a few filipods. Most of the cell surface is covered withbudding
virions (See Fig. 7), which havereplaced most of the microvilli. Theremaining microvilliareelongated and
frequentlyappearbrushlike due to the budding of virions from the sides of the microvilli.Arrowsindicateareas whereradiating clusters of virions are budding at a great distance from the cell body. x5,000.
359
VOL. 15,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.47.445.363.606.2]HOLMES
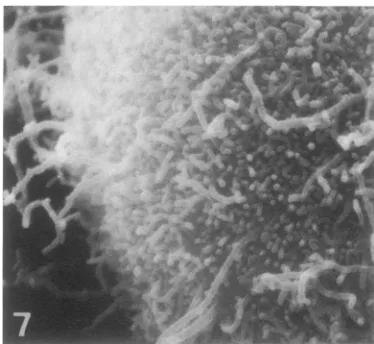
FIG. 7. Highermagnification of the lower left
por-tion of the cell shown in Fig. 6. The budding VSV virionscanbeclearlyseenbothonthe cell surface and
budding from microvilli. x10,000.
ding. Someofthesemicrovilliweremorethan5 ,umlong. In TEM studies virusesbudding from microvilli at such great distances from the cell
body might beincorrectly interpretedasvirions
that had been released into the medium. SEM,
by permitting observation of the entire cell, shows that these budding virions are still
at-tached tothe cell.
Development of VSV-induced CPE. As
moreandmorebuddingvirionsappearedonthe
surface of infected cells, major changes in cell
surface organelles occurred. By 8 h after virus
inoculation, manycellswere beginningtoshow
CPE. These cells (like that in Fig. 4) had no
ruffled membranes and showed numerous
mi-crovilli withbudding viruses.
The nature of the microvilli changed
exten-sivelyasinfectionprogressed.Inaddition to the
branching andsometimes brushlikeappearance
of microvilli due to the presence of budding
virions(Fig.4and6), infected cells often showed
veryelongated microvilli.These mayhave been remnantsoffilipodiawhich had detached from
the substrate. Because there is no identifiable
surface substructure by which virions can be
distinguished from microvilli, SEM alone
can-not prove that these long structures are of
cellularrather than viral origin. In thin sections
of VSV-infected cells, unusually long budding
virions with nucleocapsid extending the full length of the virion have occasionally been observed (10). However, since these represent
an unusual occurrence and the present study
reportselongated projections from almostevery
celllateininfection, itappearsmorelikelythat
the elongated structures were indeed cellular
microvilli.
Virus-induced changes in cell shape have
been visualized in three-dimensional images
with the SEM. Some infected cells hadbegunto
round up by 8 h after VSV inoculation. Asthe
previously flattened cells began to round up,
numerous filipods could beseen attaching the cells to the substrate (Fig. 3). However, as the
CPEprogressedandthe cellsassumeda
spheri-calshape, the number offilipodsanchoring the
cells was markedly reduced (Fig. 3 and 6). At the final stage of infection the few remaining
filipods detached and the infected cells,
stud-ded with virus particles, floated free in the medium.
DISCUSSION
Ultrastructural studies of virus-infected cell
monolayers by SEM offers several unique
ad-vantages overconventionaltechniques of TEM.
(i)The SEM permitsthe studyoflarge numbers
ofwhole cellsin situ andtherefore reduces the
samplingerrorsinherentinstudies doneonthin
sections of infected cells. (ii) The development
ofvirus-induced CPE can be studied in intact
cells with stereoscopic photographs.
Interac-tionsofinfected cells with each other and with
the substrate can readily be observed.
Virus-induced alterations in the normal cell surface
and its organelles such as microvilli, filipodia,
ruffled membranes, etc., can be detected best
by SEM. (iii) The maturation ofvirionscan be
observed in three-dimensional images and the
distribution of virus budding atdifferent sites
onthe cellsurface can readily be mapped.
These advantages are, however, dependent
upon the technique used to prepare infected
cells for SEM.When infectedcellswerefixedin
glutaraldehyde, dehydrated and air-dried from
avariety ofsolvents,grossdistortionofthe cells
resulted. Virions were obliterated or appeared
collapsed onto the surface of the cell (K. V.
Holmes, unpublished data). Gluteraldehyde
fixation(14) and subsequentdehydrationby the
critical point dryingmethod (1, 13) yielded cells
in which normal three-dimensional structure
was retained and delicate surface organelles
such as microvilli and ruffled membranes were wellpreserved (Fig. 1). With these techniques it waspossible to seethree-dimensional images of
whole virus-infected cells showing budding of
individualvirions (Fig. 2 to 4, 6).
The SEM studies reported in this paper
demonstrate that major structural changes in
the plasma membrane of L2 cells occurred as a result of VSV infection. After infection, virus
360 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.65.252.70.242.2]SEM STUDIES OF VIRUS-INFECTED CELLS buds developed and progressively covered more
and more of the cell surface. In preparations
such as these, SEM shows only those virus
buds which were still attached to the cell.
Virions that had matured and had beenreleased
would have been washed away during
prepara-tion for SEM. Late in infection, when the cellular metabolic activity is known to be mark-edly reduced (8, 11, 16-18), and the yield of
releasedvirus isverysmall (21), thesurfacesof
all the cells were covered with budding virions.
It is possible that maturation of these virions
wasarrested because they could not bereleased
from the infected cell. The process of viral
budding and release may be much faster during the period of exponential rise of released virus than in the rounded up cells at the end stage of VSV-induced CPE. In addition, the observation
that 17 h after infection most ofthe budding
virions seen in three-dimensional images were
about the same length suggests that the
devel-opment of the bullet-shaped virion may be
faster than its release. Similarly, the presence of
clusters of budding virions on the surface of
infected cells could represent aberrant develop-mental forms that could not be released nor-mally from cells and therefore stayed at the cell membrane for a longer time. Although such
multiple buds of VSV have occasionally been
observed by TEM (20), the frequency ofthese
aberrant forms can be determined best by
SEM.
Previous reports of CPE induced by VSV
indicated that infected cells rounded up, de-tached from the substrate, and floated away
into the medium. This study shows that as
infection progressed, there was a great
reduc-tion in the number of cellular filipodia, the
delicateorganelles ofthe plasma membrane by
which cells attach to the substrate orto each
other. Whether this resultedfromusurpation of
cell membrane by virus buds or an internal
metabolic alteration caused by VSV infection
mustbe determined. SEM studiesofpersistent
virus infections in vitro inwhich virus
produc-tion can proceed athighlevelsfor longperiods
would be useful to show the interactions
be-tween virus budding and organelles of the
plasma membrane that are compatible with
long life ofthe infected cell. Clearly, nutrients
neededforboth cell and virus-specificsyntheses
of nucleic acids, lipids, and proteins must be
able to enter via the plasma membrane ifthe
infected cell istoremain viable. What
propor-tion ofthe normal cell membrane isneeded to
preserve these functions remains to be
deter-mined.
ACKNOWLEDGMENTS
I thank Patricia Wiesner for expert technical assistance and A. Kabaya for help and advice. Scanning electron microscopes were generously made available by the Mostek Corp.,Carrollton, Tex., JEOL Inc., Medford, Mass., and the Department of Pathology at Southwestern Medical School.
These studies were supported by Public Health Service grantCA14517 from the National Cancer Institute and grant PO1-AI-11851 from the National Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Anderson, T. F. 1951. Techniques for thepreservation of three dimensional structure in preparing specimens for the electron microscope. Trans. N.Y. Acad. Sci. Ser. 2 13:130-134.
2. Bergold, G. H., and K. Munz. 1967. Ultrastructure of Cocal, Indiana and New Jersey serotypes of vesicular stomatitis virus. J.Ultrastruct. Res. 17:233-244. 3. Cartwright,B.,C.J.Smale, F. Brown, and R. Hull. 1972.
Model for vesicular stomatitis virus. J. Virol. 10:256-260.
4. David-West, T. S., and N. A. Labzoffsky. 1968. Electron microscopic studies on the development of vesicular stomatitis virus. Arch. Gesamte Virusforsch. 23:105-125.
5. Dulbecco, R., and G. Freeman. 1959. Plaque production bythe polyoma virus. Virology 8:396-397.
6. Dulbecco, R., and M. Vogt. 1954. Plaque formation and isolation of pure lines with poliomyelitis viruses. J. Exp. Med.99:167-182.
7. Howatson, A.F., and G.F. Whitmore. 1962.The develop-ment and structure ofvesicular stomatitis virus. Virol-ogy16:466-478.
8. Huang, A. S., D. Baltimore, and M. Stampfer. 1970. Ribonucleic acidsynthesisofvesicular stomatitis virus. III. Multiple complementary messenger RNA mole-cules.Virology42:946-957.
9. Huang, A.S.,J. W.Greenwalt, and R. R.Wagner. 1966. Defective T particles ofvesicular stomatitis virus. I. Preparation,morphologyand somebiologicproperties. Virology 30:161-172.
10. McFadden, G. K.,R. L.Truitt, andI. L.Schechmeister. 1973. Ultrastructural examination of
phytohemag-glutinin stimulatedlymphocytesinfectedwith vesicu-lar stomatitis virus. Arch. Gesamte Virusforsch. 41:229-237.
11. Mudd,J.A.,and D. F.Summers.1970. Proteinsynthesis
invesicular stomatitisvirus-infectedHeLa cells.
Virol-ogy42:328-340.
12. Nakai,T.,and A. F. Howatson. 1968. The finestructure
ofvesicular stomatitisvirus.Virology35:268-281.
13. Porter, K.R.,D.Kelley,and P. M. Andrews. 1972.The preparationofculturedcells and soft tissues for
scan-ning electron microscopy, p. 1-19. Proc. Fifth Annu.
Stereoscan Colloq. Kent Cambridge Scientific Co., MortonGrove,Ill.
14. Sabatini, D. D., K. Bensch, and R. J. Barrnett. 1963. Cytochemistryandelectronmicroscopy.The
preserva-tion ofcellular ultrastructure andenzymatic activity by aldehydefixation. J. Cell Biol. 17:19-58.
15. Simpson, R. W., and R. E. Hauser. 1966. Structural components of vesicular stomatitis virus. Virology 29:654-667.
16. Wertz, G. W., and J. S. Youngner. 1970. Interferon production and inhibition ofhost synthesis in cells infected with vesicular stomatitis virus. J. Virol. 6:476-484.
17. Wertz, G. W., and J. S. Youngner. 1972. Inhibition of protein synthesis in L cells infected with vesicular
VOL. 15,1975 361
on November 10, 2019 by guest
http://jvi.asm.org/
362 HOLMES
stomatitisvirus. J.Virol.9:85-89.
18. Yamazaki, S., and R. R. Wagner. 1970. Action of inter-feron: kinetics and differential effects on viral func-tions. J. Virol.6:421-429.
19. Yang, Y. J., D. B.Stoltz,and L.Prevec. 1969.Growthof vesicularstomatitis virus in a continuous culture line of Antheraea eucalypti moth cells. J. Gen. Virol.
J. VIROL.
5:473-483.
20. Zajac, B.,and K.Hummeler.1970.Morphogenesisofthe
nucleoprotein ofvesicular stomatitis virus. J. Virol. 6:243-252.
21. Zee,Y.C.,A. J.Hackett,and L.Talens.1970.Vesicular stomatitis virus maturation sites in sixdifferenthost cells. J.Gen. Virol. 7:95-102.
on November 10, 2019 by guest
http://jvi.asm.org/