CopyrightC1977 American Society forMicrobiology Printed inU.S.A.
Temperature-Sensitive Defect
of Vesicular Stomatitis Virus
in
Complementation Group II
ARLETTE COMBARD,* CHRISTIANE PRINTZ-ANE, CLAIRE MARTINET, ANDPIERRE PRINTZ Institut deMicrobiologie, Universitg de Paris-Sud, Centre d'Orsay, 91405 Orsay, France
Received for publication5August1976
The prototype member of the complementation groupIItemperature-sensitive
(ts)mutantsofvesicularstomatitisvirus, tsII052,has been investigated.In ts
II052-infectedHeLacellsatthe restrictive temperature (39.50C), reduced viral
RNA synthesis was observed by comparison with infections conducted at the
permissivetemperature (30'C). It wasfound that foran infection conductedat
39.50C,no38S RNA or intracytoplasmic nucleocapsids were present. For
nucleo-capsids isolated from ts II052purified virions or fromts II052-infected cells at
300C, the RNAwassensitive to pancreaticRNaseafter an exposure at 39.50Cin
contrast totheresistance observed for wild-typevirus. The nucleocapsid
stabil-ityof wild-typeviruswhenheatedto 63°Corsubmittedto varyingpHwas not
foundinnucleocapsids extractedfrom tsII 052purifiedvirions. The datasuggest
that fortsII052there isanaltered relationship between the viral 38S RNA and
the nucleocapsid protein(s) by comparison with wild-type virus. Such results
arguefor thecomplementation groupIIgene product being Nprotein, so that
thetsdefectints II 052representsanalteredN protein.
Early analyses of the temperature-sensitive
(ts) mutants of vesicular stomatitis virus
(VSV), Indiana serotype, demonstrated that
mutants belonging to complementation group
II could not be classified clearly as RNA+ or
RNA-. Inourhands,atthe nonpermissive
tem-peraturethesemutants neversynthesizedmore
than 50% of their normal RNA yield (9). For
those belonging to the Glasgow ts collection,
the prototype ts II G22 has a complete
RNA-phenotypeand ts G21induces a poor and
varia-ble amount of RNA synthesis (16). Studies of thetsmutantsof group IIhave beenhampered
by certain characteristicsregularly associated
with preparation of mutant stocks, i.e., low
yield, leakiness, and high reversionrate.
Nev-ertheless, the results obtainedin two laborato-ries(9, 23)haveledtothe conclusion that thets
defect in group II mutants might concern the
replication process of the virus. To date, this
aspect of the VSV infectioncycle hasnot been
defined. On the assumption that studies of the
group II ts mutants might help to elucidate
some stepsof the replication process, we have
designed experimentstodetermine the nature
of thets II052defect. Toobviatetheproblemof
leakiness andmultiplicationofrevertants, both
invitroandinvivostudieshave beenused.The
results obtainedrevealthatthenucleocapsid of
the ts II 052 mutant is unstable andprobably
due to some defective association between the
protein and the genome RNA.
(This work istakeninpartfrom athesisby
C.M. submittedtothe Universitkde Paris-Sud
in partial fulfillment of the requirements for
thePh.D. degree.)
MATERIALS AND METHODS
Cells andviruses. All analyses ofin vivo RNA syntheses used HeLa cell monolayers grownas de-scribedpreviously (18) or, for experimentsinwhich alow level ofresidual cellular protein synthesis was required,inchicken embryo cultures (3).
VSVs (Indiana serotype) investigated were: a heat-resistant wild type with equivalent growth ca-pabilitiesat 39.5and 30°C;a tsmutantbelongingto complementation groupIV,tsIV0111 (8);a ts mu-tant belonging to complementation group II, ts II 052 (8);andtwoindependent revertants of wild-type phenotype obtained from subclones of ts II052. All virusstocksweregrown onchicken embryo cells and contained only standard B particles (18). Virus stocks were verified by suitable complementation tests and checked for their inability to grow at 39.5°Corlowcontentofrevertants. Usually progeny virusproduced by thetsII052mutant atthe nonper-missive temperature represented 1%that obtained atthepermissive temperature. Sincetheyieldsof ts II 052 werefrequently of low titer, a concentration stepwasusually used, which involved pelletingthe virions(22,000 rpmfor1h in theSpinco SW27 rotor). Analysesofuridine-labeled products. Aftera 45-minadsorptionperiodat roomtemperature,infected cellsweretreated with 10,ug ofactinomycin Dper ml. Whenprimarytranscriptionwasstudied,
cyclo-heximide (100 jg/ml) was also added. Cells were
labeled byincorporating [3H]uridine (20 ,uCi/ml)in 913
on November 10, 2019 by guest
http://jvi.asm.org/
ET AL.
the culturemedium. Cytoplasmic extractswere pre-pared as described previously (3). Nucleocapsids wereobtained from these extractsby successive ad-dition of deoxycholate and Brij-58, both to a final concentrationof 0.5%,and centrifugationin alinear sucrose gradient (15 to 30%)ineither NEB(0.01 M Tris, pH 7.4, 0.01 M NaCl, 0.02 M EDTA) or HSB (0.01MTris, pH 7.4, 0.5MNaCl, 0.05MMgCl2) at 27,000 rpm for 3 h at4VC in the Spinco SW27.1 rotor. RNA species wereisolated from the cytoplasmic ex-tract by addition of sodium dodecyl sulfate (SDS) (1%, wt/vol, final concentration). The RNA species were fractionated at 21,000 rpm for 15 h in the SW27.1 rotor, usinga 15 to30% sucrosegradientin SDS-TENbuffer (18).
Analysis of viralproteins. Chicken embryocells werepreferred for viral protein studies because the shutoff of cellular protein synthesis is both rapid and total (3).
Conditions for labeling viral proteins within in-fected cells werethe same aspreviously described (3). After the recovery from reticulocyte standard buffer (RSB)-disrupted cells, cytoplasmic extracts were separated into soluble and insoluble compo-nents bycentrifugation at 42,000 rpmfor90 minin the Spinco SW65rotor.Proteinswereextracted from the supernatantand from thepellet by boiling sam-plesin 1%SDS, 1%mercaptoethanol, and1Murea, andwereanalyzedon 7.5%neutralSDS-acrylamide
gels. Electrophoresiswasperformedand radioactiv-ity wasestimated asdescribed elsewhere (3).
Isolation of nucleocapsidsfrom virions. Pellets of concentrated virionswereresuspendedin avolume of RSB such that theprotein concentrationdidnot exceed0.2mg/ml, andnucleocapsidswereextracted following the proceduredescribedby Emerson and Wagner (5), except that the 1-h solubilization of viruses was conducted at room temperature. Nu-cleocapsids were separated from the solubilized viral componentsbycentrifugationat45,000 rpm(Spinco SW65 rotor) for90 min. Thepelleted nucleocapsids
wererinsed several times withasaline buffer and thenhomogenized inthemedium required for fur-theranalysis.
Chemicals. Mostcompoundswerepurchasedfrom sourcesmentioned elsewhere (3, 18). ActinomycinD was agift from Merck, Sharp & Dohme. Triton X-100and Brij-58 were obtained from Sigma Chemical Co., St. Louis, Mo., and sodium deoxycholate was purchased from BDH, Poole, England. Radioiso-topes were obtained from CEA, Saclay, France. Pancreatic RNase(crystalline 5x)wasfrom Nutri-tionalBiochemicals Co., Cleveland, Ohio, and Reno-grafin-76wasfromSquibb, Princeton, N.J.
RESULTS
Characteristics of ts II 052 infections. (i)
Viral RNAsynthesized at39.5°C. Sucrose
gra-dient separations of [3H]uridine-labeled viral
RNAinducedat39.5°C by ts II 052 in HeLa cells
were found to produce an unvarying pattern. Peaks of 28S and 13 to158mRNA were always
observed, but not the peak of388 RNA (Fig.
1A). No qualitative variation was observed
0
10 20 30 40
FRACTIONS
FIG. 1. In vivo RNA synthesis directed by ts II 052. HeLa cells were infected for 45 min at room temperature by tsII 052 at a multiplicityof infection
of -40PFU/cell. Theythen receivedcomplete Eagle
minimumessential medium with 10pgof actinomy-cinDper ml and[3H]uridine(20,uilml) from1to4 hpostinfection. Cells were washed, scraped off the plates, pelleted, and disrupted in RSB. SDS was added (1%, wt/vol,final concentration) to solubilize cytoplasmic RNAs, which were thenanalyzed by cen-trifugation in 15 to30% sucrose gradients (21,000 rpm for 15 h in the Spinco SW27.1 rotor at 17°C). Arrows onthe figure show the peak positions of 18S and 28S ribosomal markers from HeLa cells. (A)
CompleteRNAsynthesis; (B)restricted RNA
synthe-sisafteradditionof100 pg of cycloheximideper ml justafteradsorption.Symbols: (@) 30°C; (0)39.5°C.
after changing the lengthortime of the labeling
period(1h or3hbetween 0to7 hpostinfection)
or the input multiplicity (10 to 100 PFU/cell), eventhough these factors do affect the apparent quantities of mRNA synthesized (unpublished
observations). First, along with the timeof
in-fection, a slight amplification was demon-stratedinthequantityof mRNAprogressively
produced at 39.5°C. Nevertheless, at all times
transcription at the nonpermissive
tempera-ture was severely depressed compared with
thatat30°C; this was noticed also for the early stages ofprimary transcription (Fig. 1B). In
these features, ts II 052 withstood a high
tem-perature less well inHeLacells than in BHK cells (1) andslightly differed fromtsIIG22 (23). In addition, in our hands, after blocking by
cycloheximide less and less RNA was
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.501.263.451.55.313.2]sized at39.50C, whereas at300C alinear
accu-mulationoccurred for several hours, asit does
normally (11).
Aneffect of themultiplicityof infectionupon
theamountof RNAsynthesis obtained withts II052-infected chicken embryo cells(9)wasalso
observed in the HeLa cell system. For HeLa
cells, the quantity ofviral RNA produced at
39.50C byts II052wasdirectly proportional to
the multiplicity of infection used (.100 PFU/
cell). At higher multiplicities, the amount of
RNA synthesis plateaued (50% of the 300C
yield).
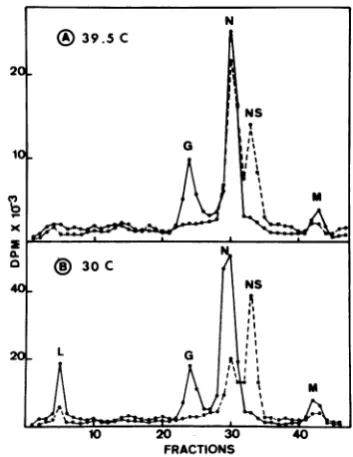
(ii) Intracellular nucleocapsids. To
deter-mine whether the absence of 3&S virion-like
RNA at39.50Cwasduetothe absence of
prog-eny nucleocapsids, cytoplasmic extracts
ob-tained 4hpostinfection from ts II 052-infected cells were separated in 15 to30% sucrose
gra-dients, and radioactivitywassoughtinthe
re-gionof thegradient where VSV nucleocapsids
should berecovered (Fig. 2). A120S nucleocap-sid peak was obtained from cytoplasmic ex-tracts made from cells infected at 300C, but
none wasdetected fromextractsobtained from cells incubated atthe nonpermissive
tempera-ture. Inaddition, nucleocapsids have notbeen
seenby electronmicroscopy of cells infected at
39.50C(V. Deutsch, unpublished observations).
Our results corroborate those of Unger and Reichmann (23), who also observed no
virus-i. 6
NC
I.
10 20 30 40 sI
Bottom FRACTIONS
FIG. 2. Intracellularnucleocapsidsproc II052. HeLa cells were infected in the r
actinomycin D,labeled with[3H]uridinefc processedtoobtaincytoplasmicextracts as Thedetergents deoxycholateandBrij-58u
totheextracts (finalconcentrationsof0.5 for each) just before layeringtheextractson
sucrosegradientsin NEBbuffer.Centrifuh
performedin theSpincoSW27.1 rotor at4 at27,000 rpm. Asa marker, unlabeled
sides extractedfrom wild-typevirions were
in the samegradient; theirpeak position
minedby optical densityisrepresentedby inthefigure. Symbols: (-)30°C; (0)39.5
like RNAornucleocapsidsintsIIG22-infected
cells incubated at the nonpermissive
tem-perature.
(iii) Cytoplasmic compartmentalization of
the N protein. Cells infected with ts II 052 at
either 30 or 39.50C were labeled for 3 h with
[3H]leucine. Cytoplasmic extracts were
re-solved accordingtothe procedure of Wagneret
al. (26) intoasedimentable anda
nonsediment-able fraction.
At300Cthe distribution of viralproteins
pro-ducedints II052-infected cells wassimilarto
that obtained from thewild-typevirusinfection
(Fig. 3B). The pellet contained both viral
pro-teins that were inserted into cellular
mem-branes (G and M) and those thatwere
associ-ated with genome RNA in nucleocapsids (N,
FRACTIONS
FIG. 3. Separation intosedimentable and
nonse-dimentable components of the cytoplasmicproteins
produced bytsII052.Afteranadsorption period of
Top 40 min at room temperature (multiplicity of
infec-Top tion, -50PFU/cell), plates(100mmindiameter) of
duced byts chicken embryo cells werecovered with5ml of1/10
presence of (vollvol)-dilutedminimumessential medium and
in-vr4h,and cubatedatthe indicatedtemperature;[3H]leucine (20 sinFig.1. uXCi/ml)wasaddedfrom1to4hpostinfection.At the
vereadded end ofthe labeling period,cytoplasmicextracts were
%, wt/vol, obtained by disrupting cells inRSB witha Dounce 215to30% homogenizerand separating them into sedimentable nationwas and nonsedimentable fractions by centrifugation at £°C for3 h 42,000 rpmfor 90 min in the Spinco SW65 rotor. nucleocap- Proteins were extracted from thesupernatantfluid alsospun (0)and thepellet(*)andanalyzedon a7.5%neutral as deter- SDS-acrylamide gelat5mA/gel for6 h.(A) Infected
thearrow cellsincubatedat39.5°C;(B)infected cells incubated
F0C. at30°C.
I
a
4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.260.437.259.488.2]916 COMBARD ET AL.
NS, and L). In the supernatant, the proteins present asfree molecules within thecytoplasm
wereevident. Itwasfound that theNSprotein
remained essentially in the nonsedimentable
fractiontogether with about30%of theN
mole-cules.
For ts II052-infected cells incubated at39.50C
(Fig. 3A), 50%of theN proteinremainedinthe
soluble fraction and the amount of labeled N
proteinthereinwas greaterthan theamountof
labeled NS protein (compare Fig. 3A and B).
These observations, together with the lack of
nucleocapsids observedinthets II052-infected
cells described above, suggest that possibly N
proteins were present in the pellet fraction
(Fig. 3A) notbecausetheywereassociated with
RNA, but because they were associated with
some other structural entity in the infected
cells (e.g., polysomes, membrane components,
etc.). Other possible explanations could be
en-tertained;however,itisclearthat(i)N, G, NS,
andMproteinsweresynthesized at39.50Cin ts
II052-infected cells and (ii) the distribution of
N protein wasabnormal.
Studyofts II 052nucleocapsidsinvitro. The
data obtained from the in vivo experiments
reported above suggest that ts II 052 may be
defectivein someaspectofitsreplicative
capac-ity. One possibility, as suggested previously
(9), is that the nucleocapsids synthesized at
39.50Carerapidly destroyed duetoanunstable
featuretheypossess atthattemperature. This
possibilitycanbe tested duetotheproperty of
VSVnucleocapsidsto befullyresistantto
pan-creaticRNasedigestion(19, 25)and other
dena-turing agents. Therefore, we examined the
ef-fect ofsome of thesefactors onthe stability of
nucleocapsids from ts II 052 virions and those
from wild-type virus. Three of these
experi-ments are reported, which demonstrate that
thereare infact significant differences between
thenucleocapsids ofts II 052and those of
wild-typevirus.
(i) Preparation of virionnucleocapsids.
Nu-cleocapsids labeled with suitableradioisotopes
wereobtained by treating a dilutedsuspension of virions in RSB by the high-salt solubilizer cocktail of Emerson and Wagner (4). It was
found that this treatment gave nucleocapsids
containing 38S RNA and Nprotein (no L pro-tein). The amountofNS protein stillassociated
withthepurifiednucleocapsidswasdetermined
by labeling the virion stock with
[32P]phos-phoric acid and determining their content of
NSphosphoprotein (14, 20)bycomparisonwith
the original virus preparation. Only5% of the
initialNSprotein wasrecoveredinthe
nucleo-capsid pellet (eitherofwild-type virus orofts
II052), whichsuggests that each nucleocapsid
containing some 700 to 2,300 molecules of N protein probably possessed no more than 2 to
11 NS molecules (2, 24). For one specified
ex-periment, the recently described extraction
procedureofEmersonand Yu (6) wasused; in
thiscase, virtuallyno NS phosphoprotein was detected in thenucleocapsid cores.
(ii) Action of pancreatic RNase on nucleo-capsids. Labeled nucleocapsids obtained from wild-type or ts II 052virionswerediluted in 2 x SSC (SSC = 0.15 M NaCl + 0.015 M sodium
citrate), pH 7.4, in order to obtain the same
final RNAconcentration (5 ,ug/ml). Theywere
incubated at 30 or 39.5°C for varying periods
ranging from 10 to 240 min. At the indicated
time, aliquots were removed and divided into
two samples. One was immediately cooled in
ice and trichloroacetic acid precipitated; the
second was digested with pancreatic RNase
(100 ,ug/ml, final concentration) for 30 min at
370Cand assayed for acid-precipitable
radioac-tivity.The RNase resistance obtainedwas plot-tedagainstthetimeof incubation(Fig. 4).
Nu-cleocapsids from the wildtyperemained
essen-100
0 0
so o -_
0
4-1
1 2 3 4
Time (h)
FIG. 4. RNase resistance ofts II 052 and wild-type nucleocapsids upon incubation at 30 and 39.50C. [3H]uridine-labeled nucleocapsids were
ex-tracted from ts II 052 or wild-type virions (=101°
startingPFU)asdescribedinMaterials and Meth-ods.Theywerehomogenizedin 2xSSC, pH 7.4, and divided into two batches, one incubated at 300C and the other incubatedat39.50C. At theprescribed
time, aliquots were cooled and assayed for their trichloroaceticacid-precipitable radioactivity, either before or after30 min of subsequent incubation at
37'C with100pgofpancreatic RNase per ml.
Sym-bols: *, wild-type nucleocapsids (the same curves were observed upon incubationat39.5 and30°C);
*, ts II 052 nucleocapsids incubated at 30°C; 0, tsII 052nucleocapsids incubatedat39.5°C.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.283.443.332.517.2]tially completely resistant to RNase at both 30
andat39.50C. Thiswas also the case for ts II 052
nucleocapsids heated at3000; however, the ts II
052nucleocapsids were much more labile
dur-ing their incubation at 39.50C. Nevertheless,
total degradation of the RNA was not obtained
and a plateau (20 to 60%) of residual RNase
resistance was observed (dependingon the
ex-periment). Anexplanation for this plateau was
obtained byexamining thesedimentation
prop-erties ofts II 052 or wild-type nucleocapsids
either in their native form or after in vitro
incubation at 39.50C with or without RNase
treatment (Fig. 5). Freshly isolated
nucleocap-sids from ts II 052sedimented only at 120S in sucrose gradients, as did nucleocapsids from
K'
a
standard wild-type particles. Radioactivity in
this peak was insensitive to pancreatic RNase
(Fig. 5A). After incubation for 2 h at 39.50C,
nucleocapsids from wild-type virus also were
recovered in the form of a120S peak resistant to RNase (Fig. 5B). However, nucleocapsids
from ts II 052 gave a more heterogeneous
pat-tern with a bimodal distribution around two
peaks (Fig. 5C), which were estimated to
pos-sess sedimentation coefficients of 120S and
140S.The120S peakwasessentiallyeliminated
bypretreatment withRNase,whereas the140S peakwas not affectedbysuch treatment. These
results suggest that the explanation for the
residual RNase resistance seeninthe previous
experiment (Fig. 4) was associated with the
BOTTOM- FRACTNS _TOP
FIG. 5. Sucrosegradient separation oftsII 052 orwild-typenucleocapsids exposedor notexposedto39.5°C andthensubmittedor notsubmittedtopancreaticRNase.Nucleocapsidswereextractedfrom [3H]uridine-labeled virionsofts1 052mutant orrevertants orofthewild-typestrainand dilutedin 2xSSC.For each virus, two equivalent samples (-5 Pg of RNA) were analyzed: one sample consistedof freshly isolated nucleocapsids (A, D), and the other was incubatedat39.5°C for2h(B, C, E, F). Eachsample wasthen dividedin twoaliquots. Onewasimmediately cooledinice(0);the otherwastreatedwith100pgofpancreatic RNaseper mland incubatedat37°C for30min(0),after which the aliquotwascooled andreceived40p1 of
diethylpyrocarbonate to inhibit the RNase activity. All samples were centrifuged on15 to 30% sucrose
gradientsinHSBat 27,000 rpmfor3h (SW27.1 rotor).Exogenous wild-typenucleocapsidswereaddedto
eachsampletolocate the 120Speak position of the normal VSVnucleocapsid(arrow).In AandD,the wild-typenucleocapsid sedimentationpatterns werenotreportedbecause oftheirsimilaritytothoseoftsII052;
andfor thesamereason,onlyone revertantcontrolprofileisgiven.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.52.443.246.559.2]918 COMBARD ET AL.
occurrence and amountofthe 140Speak.
With various preparations of ts II 052, we
haveregularly observed the RNase sensitivity
of the 120S nucleocapsid and the occurrenceof
the 140Speakuponincubation at39.50C;
how-ever, the relativeproportion of 140S and 120S
varied with the preparation (compare Fig. 5C
andFig. 6). It varied also for each preparation
with the time of incubation: the radioactive
material was first mainly in the 120S region,
and then the140S peakprogressively appeared
and was often thehighestpeakafter 2 to 4 h of
incubation at39.50C.
Other minorpeaks have also been
occasion-ally observed; they are believed to be
aggre-gates of one formoranother.
It hasbeendetermined that thenearly total
degradation of the 120Sts II 052nucleocapsids
by RNase doesnotleadtosmallderivativesin
the top half of the gradient but only to
acid-soluble material. The RNase-resistant 120S
peak has been found to contain complete
nu-cleocapsids possessing 38S RNA andNprotein.
Possibly these resistant structures represent revertantnucleocapsids.
The 120S and 140S entities were shown to
possessthe sameratio of RNA to protein
con-tent, asjudged bydouble-labeling experiments
using 14C-labeled amino acids and [3H]uridine
(Fig. 6)and by theircommonbuoyant densities
inCsCl (1.32g/ml).The sedimentationinto two
distinct nucleocapsid categories could not be
related to the length of their isolated
compo-nents: both yielded only 388 RNA and N
pro-tein, withnodetectablechangeintheir
respec-06
5
0
a --I,
FRACTIONS
FIG. 6. Protein and nucleic acid content ofts II
052 nucleocapsids incubatedat39.50C for2 h. Nu-cleocapsids wereprepared from tsII052virions
la-beled with both '4C-amino acids and [3H]uridine. They were incubated at 39.5°C for 2 h and then centrifugedina15to30%sucrosegradientinHSB
as in Fig. 5. Symbols: (@)14C-labeledaminoacids;
(0)[3H]uridine.
tive electrophoretic mobilities (data not
shown); precise evaluation ofthe NS protein
contamination couldnotbe done. However,the
same experiment as in Fig. 5 was conducted
with ts II 052 nucleocapsids that were
com-pletely NS depleted by the purification
proce-dure of Emerson and Yu (6). Nucleocapsids fromtsIV 0111 werechosen as controls (Fig.7).
Nucleocapsids just recovered after the final
treatmentof both mutants sedimented as two
peaks, 120S and 140S, which were RNase
re-sistant; somepreferential loss of 120S material
wasevident withts II 052. Nochange occurred
after a 2-h incubation at 39.5°C ofts IV 0111
nucleocapsids, whereas the 120S peakwas
re-ducedtoasmallRNase-resistantamount inthe
ts II 052 pattern.
Itwasof interest todeterminewhether
cyto-plasmicts II 052nucleocapsids obtained at the
permissive temperature possess the same
sen-sitivity to RNase as do the virion
nucleo-capsids. Therefore, ts II 052-infected cells
la-beled with [3H]uridineat300Cfor3hwereused
topreparecytoplasmicextracts inRSBasusual.
Half of eachextract received anequal volume
ofa 4x SSC solution and then was incubated
with pancreatic RNase(finalconcentration, 100
,ug/ml)
for30min at37°C. The secondhalfwassimilarly digested after a prior incubation at
39.5° for2h. Parallel analysesonHSB-sucrose
gradients of the remaining acid-insoluble
radio-active material demonstrated that the invivo
nucleocapsids fromts II052-infected cells were
sensitive to the nuclease digestion when, and
only when, they were exposed to 39.5°C. Both
120S and 140S peaks were still generated, the
radioactivity of the 140S peak beingRNase
re-sistant and the 120S material being 60%
de-graded (data notshown).
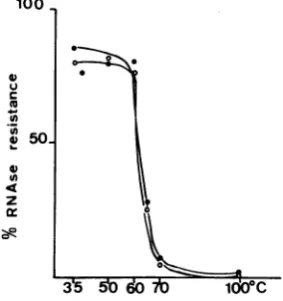
(iii) Melting of the nucleocapsidcomplex at
high temperature. "Melting" curve analyses
canbe used for studying interactions between
proteinsand nucleic acids. Byfollowing the fate
of RNA and protein usingwild-type
nucleocap-sidsheated for30min atvarioustemperatures,
M. Soria (Ph.D. thesis, Harvard University,
Cambridge, Mass., 1974)demonstratedarapid
dissociationofVSVnucleocapsids at 60 to650C
and a subsequent degradation of the nucleic
acid.
Wetherefore compared thestability of
wild-type and ts II 052nucleocapsids after30 minof
incubation in RSB (pH 7.4) at 37, 40, 50, 60, 65, 70, and 1000C. For both viruses, exactly the
same narrow temperature range for
dissocia-tion,60 to650C,wasobtained(Fig. 8).To study
the differentialthermolability of the twoviral
nucleocapsids, 630Cwaschosen. Nucleocapsids
extracted from 14C-amino acid-labeledts II 052
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.64.253.440.585.2]C)
U0
a.
FIG. 7. Analysisofts II052andtsIV0111 nucleocapsidsafter complete removal of the NS protein. The procedureofEmerson and Yu (6) was used to completely remove the NS protein from nucleocapsids ofts 1 052 andtsIV 0111.As inFig. 5, for each mutant, the nucleocapsid suspension in 2xSSCwasdividedin two samples;one wasmaintained inice(top),and theotherwasincubatedat39.50C for2h(bottom).One-half of
eachsample wasthen treated by pancreatic RNase (0), and the other halfwas untreated (-).
100
50_
z
35 51060 7001WC
FIG. 8. RNase resistance ofts II 052 and wild-typevirionnucleocapsidsincubated atdifferent tem-peratures. [3H]uridine-labeled nucleocapsids were obtainedfromts1 052andwild-type virions. After
suspension inRSB,pH 7.4, they were dividedinto aliquots of100 p1(-15,000 cpm), which were incu-batedfor30 min at the indicatedtemperatures. At the endof the incubation, all samplesweredigested
with25pgofpancreatic RNase permlat37 Cfor30 min before acid-precipitable radioactivity was
as-sayed. Symbols: (0)wild-type nucleocapsids; (0)ts
1 052nucleocapsids.
and [3H]leucine-labeled wild type were mixed
and incubated at 63TC. After incubation, the
remaining nucleocapsid structures were
puri-fiedfrom released radioactive material by
su-crosegradientcentrifugation;all fractions
sedi-mentingfasterthanorwith the 120Sexogenous
marker werepooled. The amount of each
iso-tope recovered in acid-precipitable material
versusthe time of incubationat630Cwas
deter-mined (Fig. 9). Other thanacommon 20%loss
inradioactivity ofbothlabels duringthe first 20
min of incubation at 630C (which could have
been duetodamageduringtheextraction
pro-cedure), itwasapparentthat the wild-type
nu-cleocapsidswere moreresistanttodegradation
than were those of ts II 052. We concluded,
therefore, that theinteractionsbetween the
ge-nome RNAand thestructuralprotein(s) of ts II
052 nucleocapsids were weaker than those
in-volvedinthe wild-type nucleocapsids.
(iv) Influence of pH on the nucleocapsid
configuration.Thechemical bonds involvedin
the stabilization of VSVnucleocapsids are not
known. However, ithasbeen found thatacidic
pH hampers nucleocapsid formation (7, 12).
The sedimentation properties ofnucleocapsids
fromwildtype or ts II 052 have been studied at
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.98.382.70.327.2] [image:7.501.66.208.399.550.2]920 COMBARD ET AL.
u
q, 100
IV
.0
X 50
G
0
4
0
v
b ~ ~~~~~9
10 20 30 40 50 60 Time (min)
FIG. 9. Stability oftsII052or wild-type
nucleo-capsids incubated at 63°C. Nucleocapsids were
ex-tracted from[3H]leucine-labeled wild-type virus and from 14C-amino acid-labeled ts II 052 virus. The preparations weremixed andhomogenizedinRSB,
pH 7.4. The suspension was incubated at 63°C 0.1°C, and at intervals aliquots were removed,
re-solvedon sucrosegradients, andassayed for label
remaining in the nucleocapsids. Symbols: (V) 3H-labeled wildtype;(V) "4C-labeledtsII052.
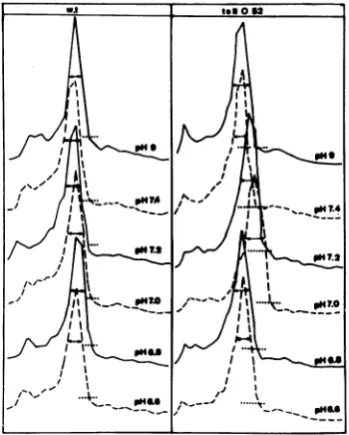
40CinsucrosegradientscontainingHSB buffer
adjusted tovariouspH values. ChangesinpH
wereobservedto exert two maineffectsonthe distribution of nucleocapsid material in the gradient: (i) achange in sedimentation veloc-ity; and (ii)greaterheterogeneityasjudged by
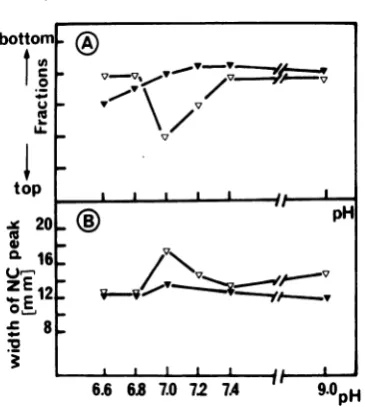
the spread of the radioactivity (Fig. 10). The variation obtainedversusthepHexposurewas
determined for both viruses and is plotted in reference to (i) the exact position in the
gra-dient of the maximum of thenucleocapsid peak and (ii) the width of this symmetrical peak at
half-height value (Fig. 11). For the wild typeno
particularchangeswereobservedinS valueor
peak width atpH values greater than 7.0. At lowerpHvalues, the nucleocapsids exhibiteda
lower S value. FortsII052nucleocapsids, both
anincreaseinpeakwidth(30%)andadecrease
inthe sedimentation coefficient (10%) were
ob-served around pH 7.0. These results indicate that thets II 052 nucleocapsids at neutral pH
possessabnormal features. DISCUSSION
Inagreementwith thestudies by Unger and Reichmann (23)on ts II G22, we have
demon-strated thatthe ts II 052 mutant at 39.5°C in
HeLa cells doesnot accumulate either nucleo-capsids or new 388 molecules. This
prop-erty, together with other characteristics of the viral RNA synthesis obtained at the
non-permissive temperature bytsII 052, makes the
group II mutants unique among the RNA+ ts mutants ofVSV; i.e., group III and group V mutants are clearly different (23; in
prepara-tion).
Theprevious data obtained withtsII052 (9)
suggested two alternative explanations for its failure to grow at39.50C: (i)there is no
synthe-sis of new genome RNA; or (ii) 388 molecules
arepolymerized but rapidly broken for lack of
stabilization. This latter possibility wasbased
onthe observation that for wild-type virus
in-tracellular 38S RNAisprincipally recoveredin
ribonucleoprotein structures involving the N
protein (10, 22). Calculations by Bishop and Roy (2) suggested that thereis atight packing
of the Nprotein along the RNAstrand, and this
probablyaccountsfor the remarkable stability
ofthe VSV nucleocapsid complextopancreatic
RNase and various other agents (10, 21, 25).
In theseanalyseswehavetested the stability
of the ts II 052 nucleocapsids under various
physical or denaturing conditions. We have
demonstrated that nucleocapsids isolated from thismutant are moresensitive toRNase when
preincubated invitro at39.50C by comparison
to those preincubated at 300C. This is in
con-trast tothe resultsobtainedat 30and39.50Cfor
wild-typevirusand for representativemutants
of all othercomplementation groups. The
[image:8.501.72.236.58.191.2]nu-cleocapsids ofanother group II mutant, ts II
FIG. 10. Effect ofpHonthesedimentation
proper-tiesofnucleocapsids from wild-typeorts 052
viri-ons. [3H]uridine-labeled nucleocapsids from
wild-typeandts 052 virionsweredivided in equivalent
batches andhomogenizedinHSB buffered with
var-iousTris-maleate bufferstogiveapHrangefrom6.6 to 7.4 or with Tris-hydrochloride to give pH 9.0.
They were then layered on15 to30% sucrose
gra-dients inHSB of thesamepHand spunat27,000
rpmfor3h in theSpincoSW27.1rotor at4°C.The profileradioactivity ofdistributionwasobtainedin
eachcase.
I.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.279.454.340.558.2]bottom
(A
Is
(U
._
x 20 C 16
zE12 8
OW
3a
v
a -I I I f
of pH
I I I I I I 1
6.6 6. 7.0 72 7A ' 9.0
FIG. 11. Demonstration from Fig.10. This
sum-marizesthemaineffects ofpHonthe sedimentation
propertiesof nucleocapsids from wild-typeorts 052 viruses.(A)Position inthe gradient where thetopof thenucleocapsid peak occurred is plotted against the pH used. (B) Width of the nucleocapsid peak (repre-senting the half-height value fromagraphin whicha
gradient fraction of0.3mlwasrepresented by5mm)
is plotted for each pH value. Symbols:(V)wild-type nucleocapsids; (V)tsII052 nucleocapsids.
063, were also observedto be RNase sensitive whenpreincubatedat 39.50C.
In addition, the fact that RNase-resistant 140S structures were derived for the ts II 052 nucleocapsids upon preheating at 39.5°C has been documented, but its significance is not
known. Any attempt to prevent its formation by using high-ionic-strength EDTA, dimethyl sulfoxide, mercaptoethanol, or Mg2+ ions was
unsuccessful. However, the unavoidable
con-version ofts II 052 120S nucleocapsidsto 140S
structureshasoccasionally been observed with the wild type or other mutant groups,
espe-cially with old stocksorwithpreparations
han-dled foralong time (see, for instance, in Fig. 7
the control pattern obtained with ts IV 0111 nucleocapsids extracted by the procedure of Emerson and Yu [6]). Unger and Reichmann (23) have reported the faster migration of the nucleocapsids ofone ts III mutant in sucrose
gradientsat39°0. Whether the140S form rep-resents aparticular physiological state of
nor-mal VSV nucleocapsidsisunclear. Inany case,
the fact that this change in the nucleocapsid
structure wasregularlyseenonly for mutants
ofgroup II (ts II052 and tsII 063) shows that
the ts II nucleocapsids are unstable in some way.
The results of analyses ofnucleocapsids
in-volvingtwoother physicalparameters,pH and
high temperature, suggest that the
interac-tionsbetweenthegenome anditssurrounding
proteins are quitedifferent forts II 052
nucleo-capsids by comparison with those ofwild-type
virus, and possibly thesedifferent interactions account for the weak stability of the mutant nucleocapsids.
Therefore,all intracellularfeatures observed
with ts II 052 at 39.50C are explained by the
incorrectpacking andprotection of ts II 052 new genomesissued froman enzymatically normal
replicationprocess, which isevidenced by the
progressive amplification oftranscription with
timeand by the depressive effect of added
cyclo-heximide.
Inrelationtothequestion of what component
isresponsible for thetsII052nucleocapsid
de-fects,somedata havetobediscussed. We have
demonstrated that the nucleocapsid coresthat
weanalyzedcontainonlytwoviralproteins: N
and NS. The general beliefisthat the N
pro-tein, present as thousands of units strongly
boundtothe genomeRNA, is endowed with the
ability to normally stabilizethe RNA. On the
otherhand, although95%of the NSprotein was
removed during the nucleocapsid extraction,
any possible role of the approximately 10
re-mainingNS molecules couldnot berejected: a
fewspecialfoldingpoints fastenedthrough the
NS protein mightsufficeto stabilize N
protein-RNA interactions. Thecomplete removal of the
NS protein from ts II 052 nucleocapsids by
Emersonand Yu'streatment (6) wasthen
car-riedout to determine whether the NS protein
must be taken into account. Insufficient
ma-terialwas available atthe end of thets II 052
nucleocapsid extraction to allow numerous
analyses, butthe effect of RNasewasthought
tobe themostconvincingtest.Two factsargue
for the preservation of characteristicanomalies
ofts II 052 nucleocapsids at 39.5° under these
conditions:occurrenceof the 140Speak and
in-stability of the 120S material. However, this
latter aspect did not involve the exogenous
RNasetreatment:the120S material
spontane-ously disappeared upon incubation at
39.50C.
Thiswas in contrast tothecompleteresistance
observedwith ts IV 0111nucleocapsids. Inthis
case, a 140S structure was also presentaswell as ts II 052 nucleocapsids notyet incubated at
39.5°C. But we have already mentioned that
the 140S form was occasionally observed with
differentmutants,especiallyafter along
prepa-ration procedure. The mutant ts IV 0111 was
chosenforcomparisonbecauseNganetal. (15)
havesuggestedthat thetemperature-sensitive
defect ofgroup IV mutants is relatedto the N
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.501.49.232.65.269.2]922 COMBARD ET AL.
protein. However, asreported by Hunt et al.
(13),the negativeresultsobtainedinthe
exper-imentsthat had ledtothis suggestiontell little about the actual lesion in group IV mutants.
The physiological behavior ofts IV mutants
reported previously (3, 18) would be more
ap-propriatelyrelated to a defect ina regulatory
protein implied in the transcription process,
which appearsto be thecase (6) for the
phos-phorylated NS protein. Similarly, our
stud-ieswithts IImutantsstronglysuggestalesion
intheir Nprotein. The defective glycosylation
of the Gprotein ints II 052-infected cells has
alsobeenreported (17). In HeLa cellswedidnot
usually encounter this defective event.
How-ever, the possible simultaneousness of
abnor-mal phenomena at both the N and G protein
levels-also reported by Wunner and Pringle
(28) on the New Jersey strain-would be
sig-nificant.
Inanycase, thepresentresults show thatts mutants belonging to complementation group
IIcould be of special interest in regardtoVSV nucleocapsid functions. Since several
independ-ent mutants have been selected (8), one may suppose that the different ts mutations have caused various changes in the nucleocapsid
structure undernonpermissive conditions. For instance, in the case of ts II 052, the defect interfering with the nucleocapsid allows the viral polymerase to act, although this enzyme
requires for its template thenucleocapsidcore
(4).Various degrees ofrelaxation in the ribonu-cleoprotein might be expected in other
mu-tants,whichin turnwouldmodifyinsome way
the template capacity of the nucleocapsid and thusleadtotheRNA-phenotype. Therefore, a
largersurveyofmutants ingroupIIwould help
to unravel thisaspectof the VSV cycle.
ACKNOWLEDGMENTS
This workwas partially supported bygrantsfrom the CentreNational de la Recherche Scientifique (L.A. 136and ATP4999.2.1),theCommissariat a l'Energie Atomique, and theFondationpourlaRechercheMedicaleFrangaise.
Wethank Annick Friedmann for her excellent technical assistanceandA. Berkaloff forcriticaladvice andreading ofthemanuscript.
C.M.isapredoctoral fellow.
LITERATURE CITED
1. Bishop,D.H.L., and A.Flamand.1975. The transcrip-tion ofvesicularstomatitisvirusandits mutants in vivoandin vitro,p.327-352. In B. W. J.Mahy andR. D. Barry (ed.), Negative strand viruses. Academic PressInc., London.
2. Bishop, D. H. L., and P. Roy. 1972. Dissociationof vesicular stomatitis virus and relationofthe viral proteinstotheviraltranscriptase. J. Virol. 10:234-243.
3. Combard, A., C. Martinet, C. Printz-Ane, A.
Fried-mann,and P. Printz. 1974.Transcriptionand
replica-tion ofvesicular stomatitis virus: effects of tempera-ture-sensitive mutations in complementation group IV. J. Virol.13:922-930.
4. Emerson, S. U.,and R. R. Wagner. 1972. Dissociation and reconstitution of the transcriptase andtemplate activities ofvesicular stomatitisvirusBand T viri-ons. J.Virol. 10:297-309.
5. Emerson, S. U., and R. R. Wagner. 1973. L protein requirement forinvitroRNAsynthesis byvesicular stomatitisvirus.J. Virol. 12:1325-1335.
6. Emerson, S. U.,and Y. H. Yu. 1975. Both NS andL proteins arerequiredfor in vitroRNA synthesis by vesicular stomatitis virus. J.Virol,15:1348-1356. 7. Fiszman,M., J. B.Leaute,C.Chany,and M.Girard.
1974.Mode of action of acid pH values on the develop-mentofvesicular stomatitis virus. J. Virol. 13:801-808.
8. Flamand, A.1970.Etudeg6ndtiquedu virus de la sto-matite v6siculaire: classementdes mutants
thermo-sensiblesspontan6sengroupesdecomplementation.
J. Gen. Virol. 8:187-195.
9. Flamand, A.,and F. Lafay. 1973. Etude des mutants
thermosensiblesdu virus de la stomatitev6siculaire
appartenantaugroupe decomplementation.II. Ann. Microbiol. Inst. Pasteur 124:261-269.
10. Huang, A.S., D. Baltimore, and M.Stampfer. 1970. Ribonucleicacid species of vesicular stomatitisvirus. III. Multiplecomplementary messenger RNA mole-cules. Virology 42:946-957.
11. Huang; A. S., and E. K. Manders. 1972. Ribonucleic acidsynthesisof vesicular stomatitis virus. IV. Tran-scriptionby standardvirusinthepresenceof defec-tiveinterferingparticles. J. Virol. 9:909-916. 12. Huang, A.S.,and E. L. Palma. 1974. Vesicular
stoma-titisvirus:defectivenessanddisease,p.87-99.InW. S. Robersonand C. F. Fox(ed.),Mechanisms ofvirus disease. W. A.Benjamin,Inc.,MenloPark, Calif.
13. Hunt, D. M.,S. U. Emerson,and R. R. Wagner. 1976. RNA- temperature-sensitive mutants of vesicular stomatitis virus: L-protein thermosensitivity ac-countsfor transcriptase restriction of group I mu-tants.J. Virol.18:596-603.
14. Imblum, R. L.,and R. R. Wagner. 1974. Protein kinase
andphosphoproteinsofvesicularstomatitis virus. J. Virol. 13:113-124.
15. Ngan,J. S. C., A. F. Holloway, and D. V.Cormack.
1974.Temperature sensitive mutantsof vesicular sto-matitis virus:comparisonof theinvitroRNA polym-erase of group I and group IV mutants. J. Virol.
14:766-.772.
16. Pringle, C. R., and I. B. Duncan. 1971. Preliminary
physiological characterization of
temperature-sensi-tivemutantsof vesicular stomatitis virus. J. Virol. 8:56-61.
17. Printz,P., andR. R.Wagner. 1971. Temperature-sen-sitivemutantsofvesicular stomatitis virus:synthesis ofvirus-specific proteins.J.Virol.7:651-662. 18. Printz-Ane, C., A. Combard, and C. Martinet. 1972.
Studyof thetranscription and thereplicationof vesic-ularstomatitisvirusby usingtemperature-sensitive mutants.J. Virol. 10:889-895.
19. Schincariol,A.L.,andA. F.Howatson.1970. Replica-tion of vesicular stomatitis virus. I. Viral specific RNAand nucleoproteinininfected L-cells. Virology 42:732-743.
20. Sokol, F., and H. F. Clark. 1973. Phosphoproteins, structural components of rhabdoviruses. Virology
52:246-263.
21. Soria,M., S.P.Little, and A. S. Huang.1974. Charac-terization ofvesicular stomatitis nucleocapsids. I. Complementary 40S RNA moleculesin nucleocap-sids.Virology 61:270-280.
22. Szilagyi, J. F., andL.Uryvayev. 1973. Isolation of an
on November 10, 2019 by guest
http://jvi.asm.org/
infectious ribonucleoprotein from vesicular stomati-tis virus containinganactive RNAtranscriptase. J. Virol. 11:279-286.
23. Unger, J.T., and M. E. Reichmann. 1973. RNA synthe-sis intemperature-sensitivemutantsof vesicular sto-matitis virus. J.Virol. 12:570-578.
24. Wagner, R.R.,S. U. Emerson, R. L.Imblum, and J. M. Kelley. 1975. Structure-function relationships of the proteins of vesicular stomatitis virus,p.1-20. In
B. W.J.MahyandR.D. Barry(ed.), Negativestrand viruses.Academic Press Inc., London.
25. Wagner, R. R.,T.C. Schnaitman, R.M. Snyder, and
C. A. Schnaitman. 1969. Structuralproteins of vesic-ular stomatitis virus. J. Virol. 3:611-618.
26. Wagner, R. R., R. M. Snyder, and S. Yamazaki. 1970. Proteins ofvesicular stomatitis virus: kinetics and cellular sites ofsynthesis. J. Virol.5:548-558. 27. Wertz,G. W., and M. Levine.1973.RNA synthesis by
vesicular stomatitisvirusandasmall plaquemutant: effects ofcycloheximide. J. Virol. 12:253-264. 28. Wunner, W. H., andC. R. Pringle. 1974.A
tempera-ture-sensitive mutant of vesicular stomatitis virus with two abnormal virus proteins. J. Gen. Virol. 23:97-106.
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG. 4.39.50C.startingtypeods.and37'Cdividedtractedtime,trichloroaceticbols:tswerebefore*, II ts RNase resistance of ts II 052 and wild-nucleocapsidsuponincubationat 30 and [3H]uridine-labeled nucleocapsids were ex- from ts II 052 or wild-type virions (=10](https://thumb-us.123doks.com/thumbv2/123dok_us/1547547.107281/4.501.283.443.332.517/startingtypeods-cdividedtractedtime-trichloroaceticbols-tswerebefore-resistance-nucleocapsidsuponincubationat-nucleocapsids-virions.webp)