0022-538X/80/06-0604/11$02.00/0
Gene
Expression
of
Herpes Simplex Virus
II.
UV
Radiological Analysis of Viral
Transcription
Units
ROBERT L.MILLETTE* ANDROSEMARIE KLAIBER
Department ofImmunologyandMicrobiology, WayneStateUniversitySchoolofMedicine, Detroit, Michigan 48201
Thetranscriptional organization of thegenomeofherpes simplexvirustype 1
was analyzedby measuringthe sensitivity of viralpolypeptidesynthesis to UV
irradiation of theinfectingvirus.Herpes simplexvirustype1wasirradiatedwith
variousdoses of UVlightandusedtoinfectxerodermapigmentosum fibroblasts. Immediate early transcription units were analyzed by having cycloheximide
presentthroughout the period ofinfection,removing the drugat8 hpostinfection,
and pulse-labeling proteins with [3S]methionine. Delayed early transcription unitswereanalyzed insimilarstudiesbyhaving9-fi-D-arabinofuranosyladenine
presentduringtheexperimenttoblockreplicationof theinputirradiatedgenome.
The viralpolypeptideswereseparatedby gel electrophoresisandquantitated by
densitometryofthegel autoradiograms. Thefollowingresultswereobtained. (i)
The UVtargetsizes for the viraltranscriptionunitsanalyzed rangedfrom 1.44 to 5.65kilobase pairs.Thisimpliesthat thecorresponding primary transcriptshave
minimummolecularweightsrangingfrom 0.46x 106to1.82x 106. (ii)Thegenes
for the four viralproteins, 165,145, 116, and 71(molecular weightx103),exhibited UV target sizes thatagreewiththeir calculatedgenesizeormeasured mRNAsize orboth and thusmustreside inpromoter-adjacentpositions. (iii) The
transcrip-tion units for theremaininggenesanalyzedshowedtarget sizesthatrangefrom 0.42to2.59kilobasepairsgreaterthan neededtoencode therespectiveproteins. Thisprobablyisareflection of their distances from promotersorthepresenceof
interveningsequencesorboth. It further suggests that thesegenesaretranscribed
asprecursorRNAmolecules thatarelargerthan their mRNA's.(iv)Theresults
indicate thatnoneofthe immediate earlygenesanalyzed canbecotranscribed,
whereassome of thedelayed early genes mightbe cotranscribed. No evidence wasfound fortheexistance oflarge, multigene transcriptionunits.
Previous studies on the characterization of
viral transcripts producedincellsinfected with
herpes simplex virus type 1 (HSV-1)
demon-strated that virus-specific transcripts found in
the nuclei ofinfected cells range from 10S to greater than 60S, whereas those found in the cytoplasm range from about 10S to 35S (31). This prompted the speculation that HSV mRNA's are synthesized as
high-molecular-weightprecursorsthatareprocessed into
lower-molecular weight cytoplasmic mRNA's (31). Subsequent sedimentation studies on HSV-1
transcripts selected by liquid hybridization
in-dicatedthatthenuclear transcriptshave onlya somewhatlargeraveragesize thanthe
cytoplas-mic transcripts (28). However,morerecent elec-trophoretic analyses of viral RNA on
methyl-mercury-agarose gels have shownthat early
nu-clearandpolyribosomalRNAshavesimilar size distributions, whereas late nuclear RNA has a considerably largeraveragesize than the
poly-somal RNA (9). Nevertheless, the true size of
the HSV-1primary transcriptsremainsin ques-tion since these studies have neither proven a precursor-product relationship between the
larger nuclear and smaller cytoplasmic HSV RNAs nor taken into account possible rapid
processing of larger nuclearprecursors.
Other recent studies have been directed
to-wardphysically mappingthe viralmRNA'sand
polypeptide coding sequences in the viral ge-nome. These haveutilizedliquid (13) and blot
hybridizationsof RNAtoHSV DNA restriction
fragments (3, 4, 9), R-loop mapping (29), and biochemicalanalysisofHSV-1xHSV-2genetic recombinants (18, 19, 21).Althoughthese stud-ies haveleadtothe mapping ofalarge number
of viralpolypeptides, RNAs,andfunctions, they
have notyetfully establishedthe number and precise location of promoters, the polarity of mosttranscription units, the existence of com-mon transcription units and intervening se-quences, orthesize andgenome mapcoordinates
of theviraltranscription units.
604
on November 10, 2019 by guest
http://jvi.asm.org/
TRANSCRIPTION
Toanswer some of thesequestionsregarding the
transcriptional
organization of the HSV-1 genome, we utilized the UVmappingtechnique developed by Sauerbier andco-workers (23,25). The principle of this method isthat the sensitiv-ityofexpression of a gene toUV irradiation isa function of the distance of that gene from its transcriptional promoter. The effect of UV ir-radiation onthe relative rate ofgeneexpression
may be quantitated either directly, by
measuring
thesynthesis of specific mRNA's, or indirectly, bymeasuring the synthesis of the specific poly-peptides. From the UV sensitivity of expression of a given gene, one can calculate theUVtarget size,in base pairs, of its transcription unit. This method has been used successfully to analyze thetranscriptional organization of a variety of procaryotic, eucaryotic, and viral genomes (for review, seereference 24).
By using host cells lacking excision repair activity, xeroderma pigmentosum (XP) fibro-blasts, toprevent the repair of the input irradi-atedviralgenome, we have been able to analyze thesensitivity of
synthesis
ofalarge number of HSV-1polypeptides
to UV irradiation of the infecting virus. The results of these studies pro-vide values for themuiimum sizes of viral tran-scription units and their primary transcripts and the distances of viralgenes from theirpromoters andplace
limitations on which genes may becotranscribed.
Moreover,they show that certain viral genesarepromoter adjacent, whereas oth-ers are considerably removed from their pro-moters and therefore must be transcribed as larger precursors.MATERIALS AND MEETHODS
Virusandcelis.HSV-1,Flstrain(7),was
propa-gatedinHEp-2 (humanepidermoid carcinoma) cells
byamodification ofpreviouslydescribed methods(26,
27).ConfluentHEp-2cellmonolayers inrollerbottles
were infected at amultiplicity of 0.02 PFU/cell in phosphate-bufferedsalinecontaining0.1%glucoseand 1%inactivated calfserum (PBS-gs).Afteradsorption at37°Cfor 2h,the mediumwasreplacedwith medium
199containing 1% inactivated calf serum, and incuba-tionwascontinuedfor2 to3daysat34°C.The cells
werescraped, centrifuged,andresuspendedinmedium
199plus1%inactivatedcalf serum,using2.5to3ml of medium per roller bottle. The cellsweredisrupted by
sonication for 2 min in an ice bath with an MSE UltrasonicPowerUnitatmaximumpower. The cell debriswasremoved by centrifugation for 30 min at
2,000xg.Viruswaspurifiedfrom thecellextractby
centrifugation through lineargradients of10to50%
(wt/vol) sucrose in VB (0.15 M NaCl-0.02 M Tris-hydrochloride[pH 7.5])for1hat25,000rpmand4°C inaSpincoSW27rotor.The virusbandwascollected,
diluted threefold in VB, and
centrifuged
for 1 h at25,000 rpm and4°C inan SW27rotor topellet the
virus.Thevirus, dissolvedinasmall volume of
PBS-gs,had a titer of 4x109 to 6x
10i
PFU/ml. Virus was plaque assayed on Vero (African green monkey kid-ney)cells and XP cells with an overlay medium of 1%methylcellulose inDulbecco-modified Eagle medium (DME) plus 1% inactivated calf serum. Virus and HEp-2and Vero cells were obtained from B.Roizman, University ofChicago,Chicago,Ill.
XPcells (AmericanType CultureCollection,
Rock-vile,Md.; no. CRL 1223, XP 12BE) were used for all UV mapping experiments. They were propagated in DMEplus 10% inactivated fetal bovine serum (Flow Laboratories Inc.,Rockville,Md.)andpassaged ata1:
3or 1:4dilution every 3 or4days. HEp-2 cells were
propagatedinmedium 199 plus 10% inactivated fetal bovine serum in 32-ounce (ca. 0.946-liter)bottlesor in medium 199plus5%calf serum inrollerbottles; Vero cellswerepropagated in DME plus 10% inactivated fetal bovine serum. Both kinds ofcellswerepassaged
every3 to4days at a 1:7 to 1:9 dilution.
UVirradiation of virus. Mixtures consisting of 1.8x 109to5x 109PFU of sucrosegradient-purified HSV-1 and9 x
10'0
to12 x1010PFUof bacteriophage T7(included asadose indicator) in 2.0 to 2.5 ml of PBS-gswereirradiated with constant stirring in a12-cmwatchglassonice. A UV lamp(Gelman Instrument Co., Ann Arbor, Mich.) that provided anoutput of 0.38to0.54
J/m2
per sat apeakwavelength of254 nmanddistance of 34 cm was used. Theviruswas irradi-ated with dosesvarying from0 to67J/m2.Samples of 20 to 50
Id
wereremoved at various times and diluted into1 to2ml ofPBS-gsonice. The UVinactivation ofphageT7wasmonitoredby removing 10-to20-ulsamplesandtitrating onEscherichia coliB., (a
re-pair-minus strain from W. Sauerbier, University of Minnesota,St. Paul, Minn.). This servedasaninternal dosestandard with 1.8 J/ m2 giving one T7 lethal hit (1).
AfterUVirradiation, alloperationsinvolving irra-diated virus and infected XP cells were performed
underyellow lightorindarkness until termination of the
experiments
toavoidphotoreactivatonof theUV-damagedDNA.
Infecting,labeling,andharvestingcells. Mono-layersof XPcells,85to99%confluent, wereprepared
in
9.6-cm2
cluster dishes. Thecellswereinfected with unirradiatedorUV-irradiatedHSV-1at20to50PFU/cell in0.4ml ofPBS-gs for 1 h at 37°C. Viruswas
removed, and the cells were overlaid with 2 ml of DMEplus1%inactivated calfserumcontainingeither
noinhibitors,50
Ag
ofcycloheximide (Calbiochem,LaJolla,Calif.) perml,or100 to 150
AM
9-.8-D-arabino-furanosyladenine(ara-A)and1jg
ofCovidarabine (2-deoxycoformycin) per ml (both were gifts from H. Machamer,Parke,Davis &Co., Detroit,Mich.).At2 to8hpostinfection (timesafteraddition of virustocells),thecellswererinsed three times with DMEplus 1%inactivated calfserum (when
cycloheximide
wasused) andtwotimes withMEM-1/100Met-1% DCS
(minimalessential mediumplus1%dialyzedcalfserum
andcontaining1/100theusualamountofmethionine) and then labeled with 0.4 ml of MEM-1/100 Met
containing[3S]methionine(Amersham Corp.,
Arling-tonHeights,Ill.)at20to30
1Ci/ml
for45minat37°C.
Thelabelingmediumwasthenremoved,andthecells
were rinsed two times with ice-cold
on November 10, 2019 by guest
http://jvi.asm.org/
bufferedsaline,lysedwith 0.2or0.3 ml oflysingbuffer
(2% sodium dodecyl sulfate,0.05 MTris-hydrochloride [pH 7.0], 0.7 M 8-mercaptoethanol, 5%sucrose, and bromophenol blue),and frozen at-90°Cuntil
electro-phoresis.
Polyacrylamide gel electrophoresis. Infected celllysateswerethawedand removed from theculture dishes with the aidofascraper,sonicatedbrieflyto reduce theviscosity,and heated for 2minat100°Cto denature proteins. Polyacrylamide slab gels (140 by
160 by 1.3 mm) were prepared essentially by the method ofLaemmli (15)but with0.36%
N,N'-diallyl-tartardiamide (Eastman OrganicChemicals, Roches-ter,N.Y.) as across-linker(8).Arunning gelof8.5%
acrylamideandastackinggel of 3.5or4%acrylamide
wereused. Proteinsamplescontaining 8,000to50,000 cpmof3Sin 20to50
pl
wereappliedandsubjectedtoelectrophoresis for 6.5 to 7.5h at200Vand 12to 16 mA. Gelswerestained for 4hwith0.25% Coomassie
brilliantblue (SigmaChemicalCo.,St.Louis, Mo.)in
ethanol-acetic acid-water (5:1:5), destained with wa-ter-acetic acid-isopropanol (8:1:1), dried by heating
undervacuum onWhatman3MMpaper,andexposed toKodak X-Omat RP film for 3to20daysfor
auto-radiography. Polypeptideswerequantitatedby
scan-ning filmswithaZienah soft-laserscanning densitom-eter (Biomed Instruments, Inc., Chicago, Ill.) and
measuring peak heights.Studies withlabeledprotein standardsrun ongelsshowed thatpeakheightswere
directly proportional to thequantityofproteinlabel under theconditions used.
Molecular weightsofviralpolypeptideswere deter-mined from mobilities relativeto protein standards
(Escherichiacoli RNApolymerase, bovineserum
al-bumin,ovalbumin, immunoglobulin G,E. coli DNase
I, papain, trypsin, chymotrypsin, and pepsin). Viral
polypeptideswerenumberedaccordingtotheir molec-ularweightsinthousandsandrelatedtoICP
(infected-cellpolypeptide)numbers ofHoness andRoizman(10,
11).
Calculations. The percentage of the amount of eachpolypeptide synthesized relative to the unirra-diatedcontrolsamplewascalculated andplottedon a
semilog plotasafunctionofUVdose to theinfecting
virus injoulespersquaremeter.Linesgivingthebest fittotheequationln(Rd/Ro) =-kdwerecalculated
bythe curve-fitting program on a Hewlett-Packard
(HP 97)calculator. In thisexpression, Rdistherateof synthesis of the individual polypeptideatUVdose d injoulespersquaremeter,Rois therateofsynthesis
ofthepolypeptideata zero UVdose,and k is the
first-orderrate constantorUVinactivationcrosssection in
squaremetersperjoule.Theslopesofthelines were
equal to the UV inactivation cross sections, k; the
reciprocalofkprovidedtheUVdoseyieldingonehit, or37%inactivationofpolypeptide synthesis. TheUV
target size of transcription units in base pairs was
calculatedby using the value of2.30 x 10-2 m2/Jas
theUVinactivation crosssectionfor 1,000 base pairs ofDNAunderourconditions. This valuewasobtained
fromaplot oftheUVinactivation crosssections, k,
versusthesizesofthegenesfor the individual
poly-peptidesinbasepairs (seebelow,Fig. 5). The number ofbasepairsrequiredtoencodeeachpolypeptidewas
J. VIROL. calculated from thefollowing relationship:number of basepairs=molecularweightofpolypeptidex3/115,
where115equalstheaverage molecularweight ofan
aminoacid.
RESULTS
UV radiological analysis of early viral
genes.In the first series of UVmapping
exper-iments,
weanalyzedthesensitivityofsynthesis
ofHSV-1immediate early (IE) polypeptidesto UVirradiationofthe virus. The mRNA for the
IE proteins accumulates in the cytoplasm of
infectedcellsduringacycloheximide block,and itcanbeassayedbyitstranslationinto
polypep-tidesimmediatelyafter removal of thedrug(11,
14).
The virus was irradiated withincreasing
doses of UV light and then used to infect XP cells.Cycloheximidewaspresentfrom 1 h after
additionof virus andthroughoutthe period of
infection.At7or8 hpostinfection , thedrugwas removed, the cells were pulse-labeled with
[3S]methionine, and the labeled polypeptides wereanalyzed by polyacrylamidegel
electropho-resis andautoradiography.SixIEviral
polypep-tidescanreadilybedistinguishedby the
sensi-tivityoftheirsynthesis to UV irradiation of the
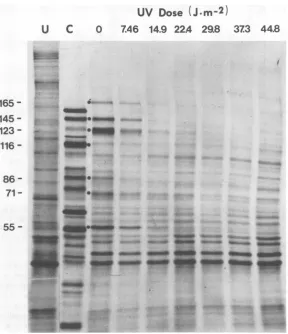
infectingvirus (Fig. 1). Thesearedesignatedas
viral polypeptides 165, 145, 123, 86, 71, and 55 in termsoftheir molecular weightsx 103 as deter-mined fromgel mobilities.Accordingtothe Ho-nessandRoizmannomenclature (10, 11), these presumablycorrespond tothe a and ,B
polypep-tidesICP 4, 6, 0, 20, 22, and 27. Insome
experi-ments, a delayed early (DE) polypeptide, 116, probablythe
ft
polypeptideICP 10,wasdetectedandanalyzed. This protein usually does not ap-pearwhencycloheximide is added with orbefore
the addition of virus. However, to achieve a better host turnoff in these experiments, the drug was added at the time ofvirusremoval, 1 h postinfection(30).
When the relative amount ofeach viral pro-tein synthesized wasdeterminedby
densitome-try of theautoradiograms and its logarithmwas
plottedagainst the UV dose given to the infect-ing virus, a series offirst-orderrelationshipswas
obtained (Fig. 2). The lines represent the best-fit plots for the datumpointsasdeterminedby computer analysis. It is apparent that the UV
sensitivity of synthesis of each polypeptide is not adirect functionofitsmolecular weightas would be expected if themRNA for each protein were transcribed from a continuous promoter-adjacent sequence.
The slopes of the lines yield the first-order rate constants for UV inactivation, orthe UV inactivationcrosssections, k, which are a
meas-ureof therelativeUVsensitivityof the
on November 10, 2019 by guest
http://jvi.asm.org/
UV
Dose
(J.m-2)
U C
0
7.46
14.9
224
29.8
165
145-
123-116- ;
86 S£*
71-
Op"-i .
55
-FIG. 1. Effect of UV irradiationonHSV-1 earlygeneexpression. HSV-1wasirradiated with UV lightat the indicated doses and used to infect XP cells. Cycloheximide (50pg/ml) waspresentfrom 1 to 8 h
postinfection.Thedrugwasremoved,and the proteinswerelabeled with[3S]methioninefor45min.Proteins wereanalyzedby gel electrophoresis and autoradiography..Viralpolypeptidesarenumbered in thousands of daltons.(U) Uninfected cell proteins; (C)control, infected cellsnottreatedwithcycloheximide.
sion of eachgene. From thesevalues, using the intrinsic dose-response factor of 2.30x 10-2
M2/
J (see below, Fig. 5), we calculated the target
sizes in basepairs of the transcription units of
the early genes analyzed (Table 1). Since the
UVsensitivityassaydoes not take into account
transcription "downstream" from thegenebeing analyzed (unlessthese sequences would be
re-quiredfortheproductionof functionalmRNA), the values shown represent minimum sizes for
the early transcription units. In Table 1, the
transcriptionunittargetsizesarecomparedwith
the number ofbase pairs calculated to encode
eachpolypeptide.Valid datawerenot obtained
forpolypeptide71 since itcomigrateswithahost
protein.
From these data we conclude the following.
(i) Theminimumtargetsizes for theearlygene
transcription unitsanalyzed rangefrom 3.05 to
5.35 kilobase pairs. (ii) The genes for proteins
116 and 165 have UV target sizes that
corre-spond closelytotheirpredictedgene sizes and thusmostlikely resideinpromoter-adjacent po-sitions.(iii) Thegenesfor viralproteins 145, 123, 86, and 55 show UV target sizes thatare0.46to
2.59 kilobasepairs greaterthan needed to encode
their respective polypeptides. This could be a
reflection of their distances frompromoters or
the presence ofintervening sequences or both
and indicates that theirprimarytranscriptsare
considerably larger than the size required for
their mRNA.
UV radiological analysis of DE HSV-1
genes.Inthis series ofexperiments,weallowed
viralproteinstobesynthesized from the onset
of infection, but to prevent the production of
nonirradiated progeny DNA molecules, we
in-hibited viral DNA replication with ara-A. As
373
44.8WA**
_4-..lg
opwo:
Q&
!.,: .:::w
4.
..p.:1 'r
....iiiiiii6lililL 17,::.
,"Wm"
dwa
'Wr--gw
'si"
400
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.106.394.75.409.2]608 MILLETTE AND KLAIBER
10
01O
XL1231
165
3 ~~~~~~~~~~~~~~~145
10 20 30 10 20 30
UV DoW (J-m2)
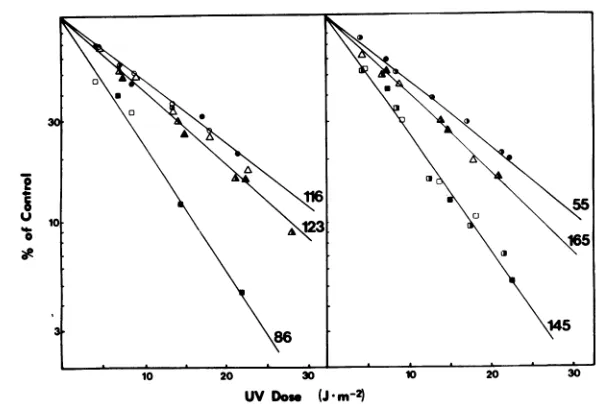
FIG. 2. Relative ratesof earlyprotein synthesisas afunction ofUV dose. Relativeratesof synthesiswere
deternined bydensitometryofseveralgel autoradiograhzs, includingthat shown inFig. 1. Linesrepresent
calculated best-fitfirst-order plotsfromthe averagesofseveralexperiments. Thedifferentkindsofdatum points indicate separateexperiments.Forabettercomparison,alllineswerenormalizedso as tointersect the
ordinateat100%. Viralpolypeptidenumbersaregivenin theright-handmargins.
TABLE 1. UVsensitivityandtargetsizesof early
HSV-Ipolypeptides
UVinacti- Base
Viral vationa Target pairs'
poly- crs e-
SZb
required Differencepeptide tion,k (base to en- (base
(mol t tion2k bas code pairs)
x10-3)
(xl0-2)
pairs)
pe°yd(x102)
~~peptide
165 9.66 4,200 4,300 -100
145 12.3 5,350 3,780 1,570
123 8.44 3,670 3,210 460
116 7.02 3,050 3,030 20
86 11.8 4,830 2,240 2,590
55 7.57 3,290 1,430 1,860
ak
values
weredeterminedbycomputerfrom theslopesof the UV inactivation plots. The values
repre-sentaveragesof threeorfour separateexperiments.
bTarget sizeswerecalculatedbydividingthe UV inactivation crosssection values, k, bythe intrinsic calibrationfactor,2.3 x10-2
m2/J
per1,000basepairs.'Number of basepairs=molecularweightof
poly-peptidex(3/115).
previously shown by Drach and Shipman (6), this drug, in conjunction with the adenosine deaminase inhibitorconvidarabine,
will
prefer-entially
andeffectively block HSV-1 DNAsyn-thesis. Futhermore,wehave found that at
con-centrations of these drugs that quantitatively
block HSV-1 DNA synthesis,
all
but the late classes of viral proteins are synthesized inin-fectedXP
cells
(Pedersen etal., submitted forpublication).
Monolayers of XP
cells
were infected withvirus that had been
irradiated
withvarious doses of UVlight
and then further incubated with mediumcontaining
ara-A andcovidarabine.
The infected cellswerepulse-labeled
for45minwith[3S]methionine
at 2 to 5 hpostinfection,
and the viralpolypeptides
wereanalyzed
by
polyac-rylamidegel
electrophoresis
andautoradiogra-phy.
Alarge number of viral
polypeptides
canbeidentified
by
correlating
their mobilities withpreviously
published gel
data(10,
11) and
from thesensitivity
oftheirsynthesis
toUV irradia-tion of theinfecting
virus(Fig. 3).
From the kineticsofsynthesis
of thesepolypeptides
(Ped-ersen et
al.,
submitted forpublication),
mostappear to
belong
to theDE,
orfl,
polypeptide
class (11). Since ara-A also causes a
delayed
turnoff
ofmanyof theearly
proteins
(Fig.
3and Pedersenetal.,
submitted forpublication),
sev-eralIEpolypeptidesalso
appeared
intheseanal-yses. In
addition,
a latepolypeptide,
154, wasconsistently
observed.The "survival"curves for the UV
sensitivity
of
synthesis
ofthesepolypeptidesareplotted
in Fig.4. As inthepreceding IEgenestudies,one canagain observethat nodirect correlationbe-tweenUVsensitivity and polypeptide size exists.
From the kinetic data, we calculated the UV inactivationcrosssections andtranscription unit
targetsizes for the viralgenesexpressed in the
presence of ara-A(Table 2). The resultsmaybe
summarized
asfollows.
(i) All transcription units analyzed in the ara-A experiments havemini-mum target sizes that range from 1.44 to 5.65
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.109.407.71.273.2] [image:5.504.66.257.336.476.2]609
UV
Dose
(J.m-2)
U
0
9.5
19
2&5 38
475
57
66.5
U
.~~~~~~
w1
e"
_t
_S.
Ins. ~ ~ ~ f
-~ ~
-1P#.usar -S
n - -.
--32
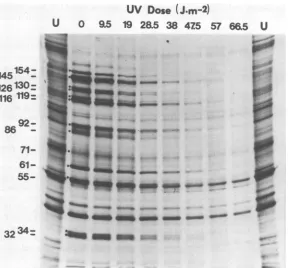
34-FIG. 3. Effect of UVirradiationonHSV-1earlyandlatepolypeptides synthesizedinthepresenceof ara-A. HSV-1 wasirradiatedwith LWlightatthedoses indicated and usedtoinfect XP cells. Ara-A (150 gA)
andcovidarabine(1 pg/ml)werepresentfromIto5.75hpostinfection. Cellswerelabeledwith[35rJmethionine
from 5to 5.75hpostinfection. Proteinswere analzed by gelelectrophoresis andautoradiography. Viral peptidesaredesignated by molecular weightx10-.(U) Uninfectedcellcontrols.
~~~~~~~~~~~~~~~~~~A
ae~~~~~~~~~~
10 20 30 10 20 30 10 20 0
[image:6.504.108.396.73.341.2]VDor (j m2)
FIG. 4. Relativeratesofsynthesisofviralpolypeptides,synthesizedin thepresenceofara-A,as afunction ofUV dose.Linesplottedrepresentcomputer-determined,best-fitfirst-order plots ofdatafromtwoorthree
experimentssuch astheoneshown in Fig.3. Thedifferent symbols representdifferent experiments. Viral
polypeptidenumbersareindicatedintheright margins.
kilobase pairs. (ii) The UV targetsizes for the
expressionof mostgenesanalyzedare
consider-ably larger than the number of base pairs
cal-culated to encode theirrespectivepolypeptides. Thedata indicate that thesegenesareremoved
from their transcriptional promoters by about 145
154-126
130:
116 119=
86
92-
71-
61-
55-_
go
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.52.453.404.589.2]610 MILLETTE AND KLAIBER
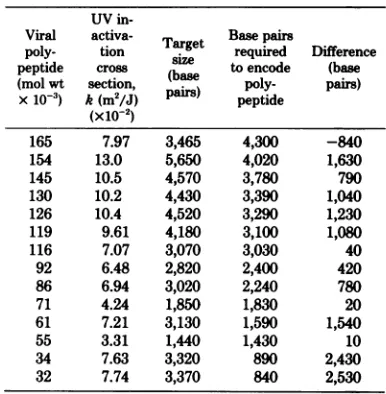
TABLE 2. UVsensitivity and target sizesofearly andlateHSV-1 genesexpressed in the presenceof
ara-Aa UV
in-Viral activa- Tat Basepairm
poly- tion
sairzge
required Difference peptide cross (b toencode (base (mol wt section, ase poly- pairs) x10-3) k(m2/J)pairs)
peptide(x10-2)
165 7.97 3,465 4,300 -840
154 13.0 5,650 4,020 1,630
145 10.5 4,570 3,780 790
130 10.2 4,430 3,390 1,040
126 10.4 4,520 3,290 1,230
119 9.61 4,180 3,100 1,080
116 7.07 3,070 3,030 40
92 6.48 2,820 2,400 420
86 6.94 3,020 2,240 780
71 4.24 1,850 1,830 20
61 7.21 3,130 1,590 1,540
55 3.31 1,440 1,430 10
34 7.63 3,320 890 2,430
32 7.74 3,370 840 2,530
aForcalculations,seeTable1andtext.The values
shown represent averages of four or five separate
experiments.
0.42 to 2.53 kilobase pairs and are, therefore,
mostlikely transcribed as larger precursor RNA
molecules.
(iii)
Thegenesfor viralproteins
116, 71,and55,however,
showaUVsensitivity
com-mensuratewiththeir predictedgenesize in base pairs. Thus these genes most likely reside in promoter-adjacent positions.Itshould be notedthat the UV target sizes for
several IE genes were about 450 to 1,800 base
pairs smaller in these studies thaninthe
preced-ing
cycloheximide
experiments. From kineticanalyses of the polypeptides synthesizedin the presenceandabsenceofara-A (Pedersenetal.,
submitted for publication) and from studies on
thepartial proteolytic peptidesofseveralofthe
IE polypeptides (S. Talley-Brown, unpublished
data),
itseemsthatwewereobservingthe same IEpolypeptidesinboth experiments.There are twolikely
explanations for this apparentshift in theUVsensitivityofthese genes: (i) expressionofthe DEgenesmight allowtranscriptionofthe IE genes from a more proximal promoter; and (ii) since DE geneexpressionisrequiredfor the turnoff ofthe IEgenes (11, 12), UVinactivation
ofthe DEgenes might cause adecreasedrate of turnoff and a lower apparent UVsensitivity of the IEgenes. With thepresentdata, we cannot
distinguish between these possibilities.
How-ever, it is clear from these considerations and from thehigherlevel ofIEgeneexpressionafter
cycloheximidereversal that the more valid data on the IE genes come from the cycloheximide experiments.
J. VIROL.
ConversionofUV inactivationcross sec-tions intotranscriptional distances.To con-vert UV inactivation cross sections into
tran-scriptional distances, ideally,oneshould haveas
a standard an internal transcription unit of known size that is expressed under the same
experimentalconditions.Lackingawell-defined
viraltranscriptionunit in thissystem,we estab-lished anintrinsiccalibrationby usingseveral of the genes analyzed. This is based on the
as-sumption that geneshaving minimum ratiosof UV crosssection togenesizereside in promoter-adjacent positions. Thus,in aplotof UV inacti-vation cross section versus RNA size in base
pairsormolecularweight,the genesadjacentto promoterswill establish aline of intrinsic cali-bration for the UV dose response under these
experimentalconditions (foradiscussion of
cal-ibration methods, see reference 24). In such a
plot with the present data, the genes for four
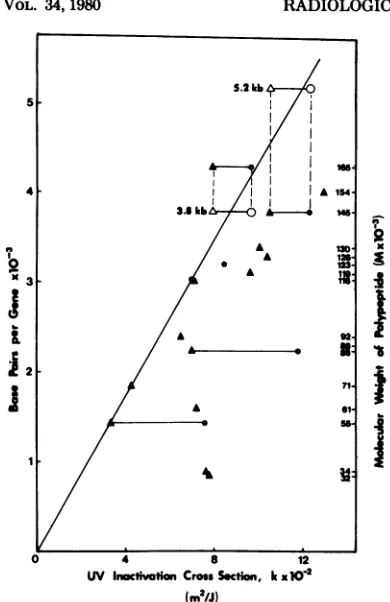
viralproteins, 55, 71, 116, and 165, establishan intrinsic calibration line(Fig. 5). Further support
for this calibration is obtained if we use the
actual measured sizes of several IE mRNA's
instead of the gene sizes calculated from poly-peptide molecularweights. Byelectrophoresisin
denaturing gels, wefound that the mRNA'sfor
viralproteins 165and145havesizes of3.8and
5.2kilobases, respectively (S. Talley-Brownand
R.
Millette,
submitted forpublication).
Using these values in the calibrationplot (opensym-bols, Fig. 5),wefound that the UVinactivation
crosssectionsforthesegenesfallvery near the
intrinsic calibration line. This line shows that
1,000 base pairs of HSV-1 DNA have a UV
inactivation cross section of 2.30 x 10-2 m2/J under theseconditions. In other
words,
it takes43.5 J/m2 to produce one
transcription-termi-nating lesion per 1,000 base pairs of HSV-1
DNA. We used this standard cross section to
calculatethetranscriptional distances.
DISCUSSION
In these studies we have demonstrated that XPcells that lackdetectable DNA excision re-pairactivitycan be usedeffectively toperform UVmapping experimentsonHSVgene expres-sion. These cells offer several advantages for such experiments. (i) They allow UV mapping
studies to be carried out at the translational level, providingthatviral DNAreplicationand
possible virus-induced DNA repair can be
blocked.Toobtainvalidresultsbythismethod, however, therate ofpolypeptide synthesismust be proportional to the amount of mRNA syn-thesized. This seemed to be the case in the presentexperiments since the UV inactivation
curves showed first-order inactivation kinetics toa 15% survivallevel or less inmost cases. (ii)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.62.255.102.299.2]-1
II
'I
UV 1noc*tvtionCrossSection, kx102 (m2/J)
FIG. 5. Plot ofgene sizeversus UV inactivation
crosssection. The datumpointsaretakenfrom
Ta-bles1and2. Thelinerepresentsanintrinsic calibra-tionforrelating UVdoseto
transcription-terminat-inglesionsproduced byUV irradiation in the viral
DNA,where k=2.3x10-2m2/Jper1,000basepairs
ofDNA.Symbols:0,datumpoints fromthe
cyclohex-imideexperiments; A,datumpointsfrom the ara-A
experiments; 0andA,datumpointsfor thegenesof proteins145 and 165replotted byusing the measured
sizesoftheirmRNA's.
In contrast to UV mapping studies in which infected cell complexes are irradiated, studies
with XP cellscan beperformed by irradiating
onlythe free virus. Thisassuresthatthe
meas-uredUV effectsare adirect result of UV lesions
in the viral genomeand allows a moreprecise
measurementof theactual UV dose delivered to
the DNA. (iii) XP cells permit UV mapping studiestobeperformedunderessentiallyDNA
repair-free conditions.With other celllines,this
condition can be approximated only by
pulse-labeling the infectedcellcomplexes immediately
after irradiation. However, this again involves dosemeasurementproblemsandprecludesUV mappingstudies at the translational level.
Themainconclusionsderived fromthese
ex-periments arethefollowing. (i) The UV target sizes for the transcription of all HSV-1 genes
analyzedrangefrom 1.43to5.65 kilobase pairs. This implies that the corresponding primary
transcriptshaveminimumsizes thatrange from
about 0.46 x
106
to 1.82 x106
daltons of RNA. (ii) Several of the genes studied, those forpro-teins 165, 146, 116, and 71, exhibit UV target
sizes that coincide with their calculated gene sizes or measured mRNA sizes or both. This suggeststhatthesegenes are promoter adjacent
and do not have extensive intervening
se-quences. (iii) The UV target sizes for the
tran-scription ofmost ofthe genes studied are larger
than calculatedtocodefor their respective
poly-peptides. Assuming thatthe transcription of
se-quencesdownstream froma gene is notrequired for the production of its mRNA,theexcess base
pairs probably reflect the distances from pro-moters orthepresenceof interveningsequences orboth. (iv)Acomparison ofthemeasured UV targetsizes with the number of basepairs needed tocode for the corresponding polypeptides
(Ta-bles1and2)reveals that few,ifany, of the HSV-1 genesanalyzed could reside incommon
tran-scription units. However, overlapping transcrip-tionunits, suchasthose observed foradenovirus
and simian virus40, cannot be ruledout.
Absence of HSV DNA repair and
repli-cation.
Tosuccessfully
apply
the UVmapping technique to the analysis of viral geneexpres-sion, it is essential that the irradiated viral ge-nomebe neitherrepaired norreplicatedduring
the experiment. The following considerations show that these requirements have been satis-fied.
(i) Hostcell DNA repair is negligible in this system. The XP cell line used in the studies
(CRL
1223,XP12BE) belongstotheA comple-mentation group, themost UV sensitive of theXP cell lines. Studies showing that these cells
have less than 2% of the rate of DNA repair found in normal human fibroblasts (22) have been verified by S.Talley-Brownin our
labora-tory with repair assays in
cell
lysates by themethod ofCiarrochi and Linn (2) (unpublished data).
Furthermore,
studies on theprolonged
survival of XP cells after UV irradiation by Maheretal. (17) and on therepair of UV-irra-diated adenovirus
by Day (5)
have shown that there isvirtually
noDNArepair
activity
in these cells.(ii) Viral DNA
repair
was not detectable inthe presence of
cycloheximide.
In theexperi-ments on IE viralgeneexpression,thepresence of cycloheximide
prevented
synthesis
of the HSV-1 DNApolymerase
andpotential
virus-specified repair enzymes. After removal of the
drug, thebrief 45-min
pulse-labeling
period
al-lowedmainlythe IEproteins
tobesynthesized.
In
experiments
notshown,
the UVsensitivity
of IE polypeptidesynthesis
wasdetermined after the irradiated viral genomes had been in theon November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.53.248.53.354.2]MILLETTE
cells for4,8, or 12h inthepresenceof
cyclohex-imide. For any given IE gene, no significant changeoccurred inthe
slope
of the UV inacti-vationcurvewithincreasing times postinfection (datanotshown). Thus,therewas nodetectablerepair or
replication
of viral DNA under these conditions.(iii) Viral DNA repair and replication were not
detectable
inthepresenceof ara-A. Tostudy
theUV inactivation of DE viralgenes, weused ara-A to block viral DNA
replication.
Under theseconditions, viralDNAsynthesis
wasneg-ligible (less than 0.15% of the control levels [Pedersen et
al.,
submitted forpublication]).
In additional UVexperiments,
viral polypeptideswere
pulse-labeled
for45minafter2to 12hof infection in the presence ofara-A.During
the first5hafterinfection,
theUV inactivationcross sections for each of the DE genes remainedconstant (data not
shown).
This indicates that there was no detectable loss oftranscription-terminating
lesions from the viral DNAduring
thisperiod.
Implications in the
mapping
of HSV-1genes. The measured UV target sizes
place
a number of limitationsonthetranscriptional
or-ganization of theHSV-1genome. Acomparison
ofthe UVtargetsizes of the IE geneswith the number of nucleotides needed tocode for their polypeptides
(Table
1) indicates that it is not possible foranyof theIE genesthatweanalyzed
tobe contranscribed. This conclusion would be validevenifweoverestimated themagnitude of thedose-response calibration factor.
Mapping data from several laboratories have shown that thegenefor IEprotein 165(ICP 4)
maps in the terminal and internal repeat se-quences
(TRs
andIRs) bracketing
the smallunique sequence (Us) and that the gene for
protein 123 (ICP 0) maps in the repeats (TRL andIRL) adjoining the large unique (UL) DNA sequence (3, 18-20). The data presented here eliminate the possibility of contranscription of
the genes acrossthe IRL-IRs junction and also preclude transcription of the IE genes from a
singlepromoterregionon acirculartemplate, as wasproposed recently by Clements et al. (3).
Acomparison of the UV target sizes with the number of base pairs needed to code for theDE
polypeptides (Table 2) reveals that there are
onlyafewpossibilities for contranscription. Al-though the data are compatible with the gene for protein 71 being promoter
proximal
tothat of32 or 34 and the gene for protein 55 lying promoter proximalto either 154, 61, 34, or 32, additional physical and UV mapping data will be needed to prove any cotranscriptions.Fur-thermore,the measured UV target sizesprovide
noindication for the existence oflarge multigene
transcription units suchas that found for ade-novirus latetranscription.
UVinactivation of
IE
genesdoesnot af-fect the UV senstivity data of the DEgenes.Thereisnowconsiderable evidence thatatleast
oneof the IEproteins is required for the
expres-sionof the DE and late viral
polypeptides (11,
12, 16,20). Itis
conceivable, therefore,
that the UV inactivation curves observed for the DEgenes representasummation oftwoeffects: the inactivation ofone or moreof the IEgenesand the inactivation of the individual DE genes. If this were the case, one can envision several possible modes of action of the IE gene prod-uct(s) onDEtranscription.
(i) An IE gene product is required in very
smallorcatalyticamountsfor theturn onof DE
genes.Letus assumefor thisdiscussion that this isanIEgenehavinganaverageUVtargetsize,
gene 165.Sinceweinfected thecells at a multi-plicity of30andinactivated thegene for poly-peptide 165 to about 10% of the controlatthe maximumUVdose, there remainedatthislevel ofinactivationapproximately three good copies ofgene 165productpercell.Accordingto
Pois-son distribution, the fraction of cells receiving
nointactcopies in thiscasewould be
e-&,
or0.05.Thiswouldcause, at most,
only
aslight down-wardcurvature totheobserved UV inactivationcurves atthehigher doses. The valuesat adose givinga90% reduction in
polypeptide
synthesis (28J/m2)
would be lowered byonly
about 5%. If theexpression
of three different IEgenes wererequired, assuming forsimplicity that each has the UV sensitivity of gene 165, the DE UV inactivationcurves at 28J/m2would be lowered by only 15%. This doesnotappear tobe thecase since nosignificant downward curvatures were
observed in theDEUVinactivationcurves.
(ii) A certain threshold level ofthe IE gene
product is required for maximum DE gene
expression; below that level theamount ofDE
expression isproportional tothe amount of IE geneexpression. In this case ourobserved DE geneinactivationcurveswould bebiphasic, first
showingaslope
intrinsic
totheUV inactivationof theDEgene and then exhibiting a slope that is the sum of that of the DE gene and that of the IE gene.Such a mechanism is very unlikely sincebiphasic curves for the UV inactivation of DE gene expression were not observed, and the target sizes for the inactivation of most DE genes are not large enough to include additional IE gene target sizes.
(iii) The amount of DE gene expression is
directly proportional to the amount of the IE gene expression. In this most extreme case we
on November 10, 2019 by guest
http://jvi.asm.org/
wouldexpect to see first-order UVinactivation
curvesfor theDEgenes, but their slopes would
be equalto thesum of the k values for the IE
plus the DE gene. This would yield apparent target sizes for the DE genes that would be at least 4,200 base pairs (the average target size for an IE gene) in excess oftheir coding require-ments. Inthe case in which more than one IE geneproduct wouldberequired for DE
expres-sion, the apparent DE gene target sizeswould beevenlarger. Theresults giveninTable 2 for
the DE genesshow that thiswas not thecase. From these considerations, we must conclude
that the UV inactivation parameters that we
observed for theDE and lateHSV-1 genes are notsignificantly alteredby the UVsensitivityof
the IEgenes.
In conclusion, we have shown that by using
exogenously irradiated virus withXPcells,one cananalyze the transcriptional organization of the HSV-1 genome by UVmapping. The data generated by this method have provided values for the minimum sizes of viraltranscription units
anddistancesof genesfrom theirpromoters and haveplaced restrictionsonwhichgenes maybe cotranscribed. This method should be
equally
applicableforstudying the transcription ofother
animal viruses, aslongasviral gene expression
canberestrictedtotheinput,unrepaired
paren-tal genome. The transcriptional distances
ob-tainedbythismethod shouldcomplement those obtained by
physical
and genetic mapping in that they should include all nontranslated se-quences required for gene expression.Futher-more, they can provide direct evidence for
co-transcription,
polarity
oftranscription, and pro-motershifts.ACKNOWLEDGMENTS
WearegratefultoSueTalley-BrownandMargaret
Ped-ersenforenlightening discussions,invaluable assistanceinthis
research, and many hours devotedtoreading manuscripts and drawing figures. Special thanks are due Walter Sauerbier, University of Minnesota, forhishelpfulcriticisms and
discus-sionsof this work.
Thisinvestigationwassupportedby Public Health Service grantCA21065awardedbytheNational Cancer Institute.
LITERATURE CITED
1. Brautigam, A. R., andW.Sauerbier. 1973. Transcrip-tionunitmapping inbacteriophageT7. I. In vivo tran-scriptionby Escherichiacoli RNApolymerase.J.Virol. 12:882-886.
2. Ciarrochi, G.,andS.Linn. 1978. A cell-freeassay meas-uringrepairDNAsynthesisinhuman fibroblasts. Proc.
Natl. Acad. Sci.USA75:1887-1891.
3. Clements,J.B.,J.McLauchlan,andD.J.McGeoch.
1979.Orientation ofherpessimplexvirustype1
imme-diateearlymRNA's. Nucleic Acids Res. 7:77-91.
4. Clements,J.B.,R.J.Watson,andN. M. Wilkie. 1977. Temporal regulation ofherpes simplex virus type 1 transcription: location oftranscripts on the viral
ge-nome.Cell 12:275-285.
5. Day, R. S.,M. 1974. Studies on repair of adenovirus 2 by humanfibroblasts using normal, xeroderma pigmento-sum,and xeroderma pigmentosum heterozygous strains. Cancer Res. 34:1965-1970.
6. Drach, J. D., and C. Shipman. 1977. The selective inhibition of viral DNA synthesis by chemotherapeutic agents: an indicator of clinical usefulness? Ann. N.Y. Acad. Sci.284:396-409.
7. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968. Characterization of herpes simplex virus strains differ-ing in their effect on social behavior of infectedcells. J. Gen. Virol. 3:357-364.
8. Gibson, W., and B.Roizman. 1974. Proteins specified by herpes simplex virus. X. Staining and radiolabeling properties of B capsid and virion proteins in polyacryl-amide gels. J. Virol. 13:155-165.
9. Holland, L. E., K. P. Anderson, J. R. Stringer, and E. K.Wagner. 1979. Isolation and localization of herpes simplex virus type 1 mRNA abundant before viral DNA synthesis. J. Virol.31:447-462.
10. Honess, R. W., and B. Roizman. 1973. Proteins specified by herpes simplex virus. XI. Identification and relative molar rates of synthesis of structural and nonstructural herpes viruspolypeptides in the infected cell. J. Virol. 12:1347-1365.
11. Honess, R. W., and B. Roizman. 1974. Regulation of herpes virus macromolecular synthesis.I.Cascade reg-ulation of the synthesis of three groups of viral proteins. J. Virol.14:8-19.
12. Honess, R. W., and B. Roizman. 1975. Regulation of herpes virus macromolecular synthesis: sequential tran-sitionofpolypeptide synthesis requires functional viral polypeptides. Proc. Natl. Acad. Sci. U.S.A. 72:1276-1295.
13. Jones,P.C., G. S. Hayward, and B. Roizman. 1977. Anatomy of herpes simplex virusDNA.VII. a RNA is homologous tononcontiguous sites in both the L and S components of viral DNA. J. Virol.21:268-276. 14. Kozak, M., and B. Roizman.1974.Regulation of
herpes-virus macromolecular synthesis: nuclear retention of nontranslated viral RNA sequences. Proc. Natl. Acad. Sci. U.S.A. 71:4322-4326.
15. Laemmli, U. K. 1970. Cleavage of structural proteins duringtheassembly of the head ofbacteriophage T4.
Nature(London)227:680-685.
16. Leung, W.-C. 1978. Evidence for a herpes simplex virus-specific factorcontrollingthe transcription of deoxy-pyrimidinekinase. J. Virol. 27:269-274.
17. Maher,V.M., D. J. Dorney, B.Konze-Thomas, A. Mendrala, and J. J. McCormick. 1979. DNA excision repairprocesses in humancellscaneliminate the cyto-toxicandmutagenic consequences of ultraviolet irradia-tion.Mutat.Res.62:311-323.
18. Marsden, H. S., N. D. Stow,V. G. Preston, M. C. Timbury, andN. M.Wilkie.1978.Physical mapping ofherpessimplex virus-induced polypeptides. J. Virol. 28:624-642.
19. Morse,L.S.,L.Pereira,B.Roizman,andP.A.
Schaf-fer. 1978. Anatomy of herpes simplex virus (HSV)
DNA. X.Mapping of viral genesbyanalysisof
polypep-tides andfunctionsspecified byHSV-1xHSV-2 recom-binants.J. Virol.26:389-410.
20. Preston,C. M.1979.Control ofherpessimplexvirustype 1mRNAsynthesisincells infectedwithwild-typevirus orthetemperature-sensitivemutanttsK. J. Virol. 29: 275-284.
21. Preston,V.G.,A. J.Davison,H. S.Marsden,M. C. Timbury,J.H.Subak-Sharpe, andN. M.Wilkie.
1978.Recombinantsbetweenherpessimplexvirustypes
1and 2:analysesofgenomestructuresandexpression
ofimmediateearlypolypeptides.J. Virol. 28:499-517. 22. Robbins,J.H.,K.H.Kraemer,M. A.Lutzner,B. W.
Festoff,andH.G.Coon. 1974.Xeroderma
on November 10, 2019 by guest
http://jvi.asm.org/
sum, aninherited diseasewithsunsensitivity, multiple
cutaneousneoplasms, and abnormalDNArepair.Ann. Intern. Med. 80:221-248.
23. Sauerbier, W. 1976.UVdamageatthetranscriptional level.Adv. Radiat. Biol. 6:49-106.
24. Sauerbier, W.,and K. Hercules. 1978. Gene and tran-scription unitmapping byradiationeffects. Annu.Rev. Genet. 12:329-363.
25. Sauerbier, W., R. L. Millette,andP. B.Hackett, Jr.
1970.The effects of ultraviolet irradiationonthe
tran-scription of T4 DNA. Biochim.Biophys.Acta 209:368-386.
26. Spear,P.G.,and B. Roizman. 1968. Theproteins spec-fiedbyherpes simplex virus.I.Timeofsynthesis, trans-fer into nuclei, and properties ofproteins made in productively infectedcells.Virology 36:545-555. 27. Spear,P.G.,and B. Roizman. 1972. Proteinsspecified
byherpes simplex virus.V.Purificationand structural
proteins of the herpes virion. J. Virol. 9:143-159 28. Stringer,J.R., L.E.Holland,R. I.Swanstrom, K.
Pivo,and E. K.Wagner. 1977. Quantitation of herpes simplexvirus type 1 RNA ininfected HeLa cells. J. Virol.21:889-901.
29. Stringer,J.R.,L.E.Holland,and E. K. Wagner. 1978. Mappingearly transcripts of herpes simplex virus type 1byelectronmicroscopy. J. Virol.27:56-73. 30. Talley-Brown,S., and R. L. Millette. 1979. Gene
expres-sion ofherpessimpley,virus. I. Analysis of cytoplasmic RNAs ininfected xeroderma pigmentosum cells. J. Vi-rol.31:733-740.
31. Wagner,E.K.,and B.Roizman. 1969. RNA synthesis in cellsinfected with herpes simpex virus. II. Evidence that a class of viral mRNA is derived from a high molecularweight precursor synthesized in the nucleus. Proc. Natl. Acad.Sci. U.S.A. 64:626-633.