0022-538X/80/10-0224/12$02.00/0
Densonucleosis Virus Structural Proteins
D. C.KELLY,* N. F. MOORE,1C. R SPILLING,1 A. H. BARWISE,2 ANDI.0. WALKER'
Natural Environment ResearchCouncil,Instituteof Virology, Oxford,OX) 3UB,1 andDepartment of
Biochemistry, Universityof Oxford, Oxford, OX) 3QU,2 United Kingdom
The proteincoatsoftwodensonucleosisviruses(types1and2)wereexamined byavariety ofbiophysical, biochemical, andserological techniques. The viruses were24 nmin diameter, containedatleast fourpolypeptides, wereremarkably
stable toextremes ofpH and denaturing agents, andwere serologically closely related. The two virusescould, however, be distinguished serologically and by differencesinmigrationof theirstructuralpolypeptides. For each virus the "top component" (i.e., the protein coat minus DNA, found occurring naturally in infections) appeared to have acomposition identical tothat ofthe coat of the
virus andwasa morestablestructure.Electrometric titrationcurvesofthe virus particles and top components demonstrated that the DNA phosphate in den-sonucleosisvirusparticleswasneutralizedbycations other than basicaminoacid sidechains of theproteincoat.Circular dichroism studiesshowed thattherewas a conformational difference between the protein coats oftop components and virusparticles.
Densonucleosis viruses (DNVs) are small
icosahedralviruses,isolated from
invertebrates,
whichcontainlinear
single-stranded DNA,
andthey sharemanystructural features incommon
with both the defective and the nondefective
parvoviruses
isolated from vertebrates(10, 17).
Structurally, the DNA contained
by
DNVs iscomparable
tothat of the defectiveadeno-asso-ciatedvirusgenome and isorganizedas
comple-mentary, separately packaged, linear
single-stranded DNAwhich possesses inverted
termi-nalsequencesandalimited nonrandomcircular
terminal
permutation (1,
9, 13,14).
Thegenomeis unlike the nondefective
parvovirus
genome,which (as
typified by
minute virusofmice andKilham ratvirus) is
organized
asalinear DNAmolecule
(of
onepolarity)
with shortpalin-dromicsequencesatthe ends
(2).
Little is known about the structure of the
protein coat ofDNVs.
Tijssen
et al. (33) havereported thataDNV of themothGalleria
mel-lonella contains four structural
polypeptides
withmolecularweightsof 98,000,69,000, 58,500,
and49,000and that the98,000-dalton
polypep-tide isadimerof the
49,000-dalton
polypeptide.
We have
previously
shown that theprotein
coat of DNVs has amolecular weight of3.55 x10'
and that themolecularweightofthevirus par-ticle (5.7x106)
represents theapproximatesum of the protein coat, the genome (molecularweight, 1.95x 106),and thepolyamines
(molec-ular
weight,
80,000) which constitute thecom-plete virus particle (13-15). In this study we
have examined theprotein coats of two DNVs
by exaning either the top components (i.e.,
the virusparticles minus DNA and polyamines)
orthecomplete virus particles.We have used a
variety of physical methodstoexaminethe
poly-peptide composition
ofthe coat, the secondarystructureofthe
protein,
andtheinteractions of thecoatwith the genome. Inaddition,we havecompared thetwoviruses serologically.The two
viruses studied wereDNVtype 1 (DNV1),
iso-lated from G. mellonella (24), and DNV2,
iso-latedfromthe
butterfly
Junonia coenia (29).MATERIALS AND METHODS Virusproduction andpurification. DNV1 was originallyisolatedby G. Meynadier. DNV1 and DNV2 wereprovided by C. F. Rivers. DNV1 and DNV2 virus particles and corresponding top components were pre-paredaspreviously described (13, 20).
Determination of extinction coefficients and concentrations. DNAwasestimatedbythe diphen-ylaminemethod asmodified by Burton (3). Calibration curves were prepared with standard solutions of 2-deoxy-D-ribose. RNA was estimated by the orcinol method as modified by Mejbaum (23). Calibration curves were prepared with standard solutions of D-ribose.
Due to lack of material, it was not possible to determine the extinction coefficients of virusparticles and top componentsby the dryweight method. The extinctioncoefficients weretherefore measured indi-rectly, usingarefractive index methodtodetermine protein concentration and estimating phosphorus of virusparticles after dialysis into 0.1 MNaCl by the method of Jones etal.(12).
The estimation ofprotein involved measuring the areaofasynthetic boundary ofasample of top
com-224
on November 10, 2019 by guest
http://jvi.asm.org/
ponentphotographed withSchlieren optics in a Spinco model E analytical ultracentrifuge and comparing this with the area obtained by the same method at the same bar angle for a standard solution oflysozyme, assuming the refractive index increment of both pro-teins to be 0.193/g per ml at 435.8 nm (5). The method gave a measure of the protein concentration of the solution of top component. Since the refractive index increment of DNA at 436.0 nm is also 0.193 (27), a similar method was used with solutions of virus par-ticles and top component sedimented appreciably at the speeds used (usually 12,590 rpm), photos were taken at various time intervals, and the measured areas were extrapolated to zero time to obtain the initialarea.
Routine analysis for protein was by the Folin method (21) as modified by Eggstein and Kreutz (6). Bovine serum albumin was used as a standard, and a correctionwasapplied by using the previous refractive index method and the Folin method on the same sample. Extinction coefficients of virus particles and top components wereobtained by measuring the ab-sorbanceofasample ofknown concentration in a
1-cm-path-length cellat260and 277 nm, respectively. Results were expressed in optical density units per milligram per milliliter for a 1-cm path length. UV spectraof virus particles and top components were obtainedat25°Coneither a Cary14recording spec-trophotometer oraUnicanS.P. 500 spectrophotome-ter.The extinctioncoefficients for virus particles and top componentswere 9.49±0.32/mg perml at 260 nm and 1.39±0.11/mg per mlat 277nm,respectively.
Electron microscopy. DNV particles and top componentsnegatively stained with 2% (wt/vol) ura-nyl acetate were examined on an AEI 6B electron microscopeaspreviously described (20).
Analytical ultracentrifugation. Analytical ultra-centrifugationwasperformedaspreviously described (13), using aBeckmanSpinco analytical ultracentri-fuge equipped with both Schlieren and absorption opticalsystems.
Preparationof antisera. Antiserawereprepared inboth rabbits andguinea pigs.Preimmunesera were obtained from rabbitsbybleedingfrom themarginal earvein andfromguinea pigs by cardiac puncture.
Rabbits were immunized by injecting at weekly intervals 1 ml of virus (containing 1 mg of virus) emulsified with 1 ml of Freund complete adjuvant (Difco Laboratories, Detroit, Mich.) subcutaneously, followedbyasimilar subcutaneousinjection with in-complete adjuvantandfinallyanintravenousinjection of1 ml of virus. Immune sera were taken4 weeks later.
Guineapigswereimmunizedbytwointraperitoneal injections (7 days apart) of1ml ofvirus (2 mg ofvirus) emulsifiedwith1mlofFreundcomplete adjuvantand werebled by cardiac puncture 3 weeks later to provide immunesera.
Gelimmunodiffusion.The reactions between
an-tibody and antigenwereexaminedby performing mi-crotests on glass slides (3 by 1 inch [ca. 7.6 by 2.5 cm]) coveredin0.75% (wt/vol) Ionagar 2 (Oxoid Ltd., London, United Kingdom) inphysiologicalsaline containing 0.5% (wt/vol) sodium azideand 0.001%(wt/ vol) trypan blue.
Polyacrylamide gel electrophoresis. Three polyacrylamidegel electrophoresis systems were used during these studies.
(i) Polypeptides were analyzed on 16-cm-long 12.5% polyacrylamide slab gels, using the buffer system of Laemmli (19) as previously described (7). Samples weredenatured by boiling in 0.1 M Tris-hydrochloride (pH 6.8)-2% (wt/vol) sodium dodecyl sulfate (SDS)-2% (vol/vol) 2-mercaptoethanol-10% (vol/vol) glyc-erol-0.01% (wt/vol) bromophenol blue. ,B-D-Galacto-sidase(molecular weight 132,000), phosphorylase (mo-lecular weight, 97,200),transferrin(molecularweight, 90,000), bovine serum albumin (molecular weight, 69,000), and ovalbumin (molecular weight, 46,000) wereusedtocalibrate thegels for molecular weight determinations.
(ii) Polypeptideswereanalyzedon10and 7% SDS-polyacrylamide cylindrical gels(6 by 100mm),using acontinuousphosphate-bufferedsystem aspreviously described (16). Thesamefive standardswereused for molecular weight determinations.
(iii) Native virus particles and top components wereanalyzedon3%polyacrylamide gels (5 by75mm) made up in 0.1 M Tris-hydrochloride, pH9.0, inthe absenceof SDS. Reservoir buffers contained0.025M Tris-glycine,pH9.0. Samplesin reservoirbufferplus 10% (wt/vol) sucrose were applied directly to gels. Electrophoresiswas at5mA pergelfor about70min. Nonreducing gelswere runby omitting mercapto-ethanol from the systemaspreviouslydescribed(25). Sugar residues were detected on proteins by Schiff stainingasdescribedby Fairbanksetal.(8).
Proteinswere located byCoomassie blue staining
aspreviouslydescribed.
'"I-labled
proteinwaslocated byautoradiography, using Kodirex X-ray film. Coo-massie blue-stained gels and autoradiographs were scanned inaJoyce-Loeblmicrodensitometerequipped withathin-layerattachment.lodination ofpolypeptides. Iodination of poly-peptideswascarried outessentially asdescribed by Mooreetal. (26). Intact virusortop componentwas iodinatedchemicallywithchloramine-Tor enzymati-callywith lactoperoxidase. Virus components previ-ouslydenaturedby boiling in1% (wt/vol) SDS for2 minwerealsoiodinatedbythe chloramine-Tmethod. Iodination catalyzed by lactoperoxidase was per-formed by adding 10 ILI (100
ACi)
of carrier-free 125I (Radiochemical Centre, Amersham, United King-dom),5pl
oflactoperoxidase (1 mg/ml)(Sigma Chem-ical Co.Ltd.,London,UnitedKingdom),and10pl
of freshlydiluted 0.05%(vol/vol) H202to 100pl
of phys-iologicalsalinecontaining20 ugof virusortopcom-ponent.The incubationwascarriedoutat room tem-peraturefor30minandstopped by dilutingthe
reac-tion40-fold withphysiologicalsaline.
Iodinationbychloramine-Twasperformedby add-ing 10,ul (100
jCi)
of"2I
to100pl
of intactor SDS-disrupted virus(20Agofprotein),followedby10pl
of chloramine-T (14mg/ml). The reactionmixturewasvigorously shaken for30s, andthereactionwas
ter-minatedbytheadditionof20
pi
of sodium metabisul-fite(28mg/ml).All iodinatedsamplesweredialyzed againstseveral 1-literchanges ofphysiologicalsaline before electro-phoresis. Intact virus componentswere rebandedon
on November 10, 2019 by guest
http://jvi.asm.org/
20 to50%(wt/vol)sucrosegradients made up in 7 mM sodiumphosphate(pH7.2)-0.1MNaCl. The gradients were run at 35,000rpmfor16hin a swing-out rotor (6 by 16.5 ml; M.S.E. Ltd., Crawley, Surrey, United Kingdom).
Circular dichroism spectrometry. Circular di-chroism spectraof DNV particles and top components wererecorded at200Cin aRousellJouan Dicrographe III, using cells of 0.01-, 0.1-, or 1.0-cm path length. Circular dichroism results arereported in terms ofe,
the mean residue ellipticity in unitsofdegrees per square centimeter per decimole. The mean residue weightof an amino acid residue was 108 as determined from amino acidanalysis(20).
Continuous electrometric titrations.
Continu-ouselectrometic titrationswerecarried outin a Ra-diometermicrotitrationassembly,consisting of a ther-mostatically controlled glass titration cell and mag-netic stirrer.HCl (0.1 N) or0.1 N NaOH was added fromanAgla glassmicrometersyringe.pH was mea-sured with anE.I.L. direct-reading pHmeterwith a glass electrode and a calomel electrodejoined by a KCl-saltbridge. Before titration a stream of nitrogen wasbubbledthroughthesample until the pH became constant,to removedissolved carbon dioxide. During thetitrationastreamofnitrogenwasblown over the sample.All titrationswerecarriedoutat25°C. Titra-tionswereperformed ontop component (2.5 mg/ml) in0.1 M KClandonvirusparticles (3.75 mg/ml) in 0.1MKClbelowpH5.7and in0.001 MKClabove pH 5.7.
The titration curve of top components or virus particles wasconstructed by subtracting the solvent titrationcurve(measured separately and corrected for dilution) from the sample titration. The results are expressedasmoles ofH+dissociatedorassociated per 100mol of amino acid residues. Todetemnine if titra-tionswerereversible, thesampleswere immediately retitrated from highorlow pH back to about pH 6.0 with0.1NHClor 0.1NNaOH.
RESULTS
Electron
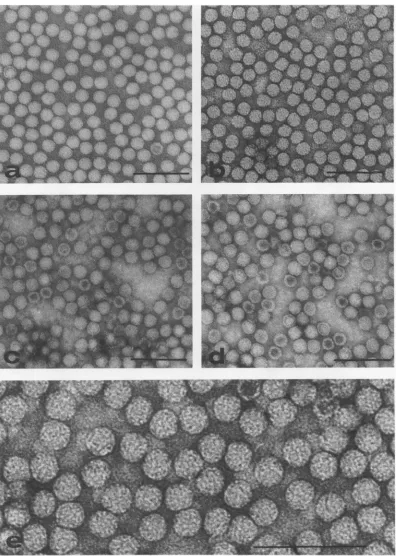
microscopy.
Preparations of purevirus
particles
and top components of DNVstained with 2%uranylacetateshowed that all
preparations comprised small, spherical,
pre-sumably icosahedral,
particles about 24nm indiameter (Fig. 1). Some of the virus particles
werepenetrated byavarietyofstain, including
uranyl acetate, ammonium molybdate, and
so-dium tungstosilicate. The extent ofstain
pene-tration oftop components varied from
experi-ment toexperiment,but an appreciable number
of particles remained inviolate. Structure was
observed, but not in sufficiently fine detail to
enumerate capsomeres with confidence. Such structureis shown in Fig.1.
Serology. Relationships betweenDNV1 and
DNV2 were examined by using antisera
pre-paredinrabbits andguineapigs against DNV1
andin guinea pigs againstDNV2.
Microimmu-nodiffusion
tests wereperformed
to conserveantigens and antisera.
Suspensions
of bothvi-ruses were
placed
inadjacent wells,
and theirreactions with the three antisera were
moni-tored.After 48 hadjacentbands ofprecipitation
in the gels became contiguous. When the
sus-pensionsweretestedagainstantiserumprepared
inrabbits,atinyspurwasformed where thetwo
bandsofprecipitationagainstDNV1 and DNV2
met (Fig. 2A). This spur was acontinuation of
the
precipitation
bandformed
by
DNV1. Toshow that this was not an
artifact,
the samevirus
suspension
wasplaced
inadjacent
wells;
therewere nospursontheprecipitationbands
produced
(Fig.
2BandC).
When the antiserumprepared in guineapigs
against
DNV2wasusedtocompareboth
viruses,
itwasfound thatsingle
bands of
precipitation
becamecontiguous,
withthe band formed
by
DNV2continuing
into alarge spur
(Fig. 2D).
Theguinea pig
antiserumprepared against
DNV1 wasof lowtiter,
but itstillgave avisible spuronthe band created
by
DNV1
(Fig. 2E).
The virusparticles of DNV1 andDNV2were
serologically identical to their
respective
topcomponents
(data
notshown).
Polyacrylamide
gel electrophoresis
ofDNV
structuralpolypeptides.
Polyacryl-amide
gel
electrophoresis
revealed that the virusparticles contained four main structural
poly-peptides,
designated
indecreasing
molecularweightasVP1
through VP4,
asshown inFig.
3.Adeno-associated
virustype 2polypeptides
areshown for
comparison.
The molecularweights
ofthese
polypeptides
areindicated in Table 1.Es-timates of molecular
weight
varied with thebuffersystemand
gel
concentrationsused. Thepolypeptide
compositions
of the virusparticles
andtop componentswereidentical for each virus(Fig. 4),
though
the relative amounts of eachpolypeptide
variedslightly (Table
2andFig. 4).
Additional
polypeptides,
intermediate in molec-ularweight
between VP1 andVP2,
weresome-times detected
by
Coomassie bluestaining
though they
were morereadily
detectedby
io-dination
procedures.
Omission of a
reducing
agent from thegels
failed to alter the
polypeptide profiles
of thecomponents,
showing
thatdisulfide
bonds arenot involved in
interpolypeptide
interaction.Schiffreagent failed to
positively
stain any ofthe
polypeptides;
this is tentative evidence that thepolypeptides
are notglycoproteins.
lodination
ofDNV polypeptides.Iodina-tion of DNV particles and top components was undertaken to explore the location of the various
structural polypeptides within these virus
par-ticles. The data presented here are for DNV1 components, though
similar
results were ob-tained with DNV2 components.AR
four main J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
-_ -L_w---t aM-* - .*-,_i
FIG. 1. DNVparticles negativelystainedwith 2%uranylacetate,showing generallyfeaturelessstructure. (a) DNV1particles; (b) DNV2particles; (c) DNVI top components; (d)DNV2 top components; (e) DNV2 particlesgreatly enlargedtoshow tentative substructure. Note stainpenetrationin some, butnotall, top
components. Bar=100nm.
.1 :
:6
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.510.54.450.67.624.2]FIG. 2. Serologicalrelationshipsbetween virusparticlesofDNV1 and DNV2 testedby microimmunodif-fusion. Wells: 1, rabbit antiserum againstDNVI; 2, guinea pigantiserum against DNV2; 3, guinea pig antiserum against DNV1;g,DNV1 antigen;j,DNV2antigen.Note the small butwell-definedspurin A.
A
B
C
[image:5.510.129.412.70.322.2]_ -_
FIG. 3. SDS-polyacrylamide gel electrophoresis of the structuralpolypeptidescontainedby DNVI (A), DNV2(B),and adeno-associated virus(C). The pro-teinswereresolvedon a12.5% slabgel.
structural polypeptides and the minor, fifth polypeptide were efficiently radiolabeled with 125I whenchemically iodinated with chloramine-T after disruption with SDS (Fig. 5), and the profilewassimilartothe Coomassieblue stain profiles. Both lactoperoxidase and chloramine-T radiolabeled the two high-molecular-weight
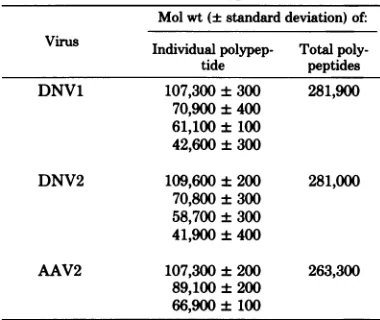
TABLE 1. Molecularweights ofstructural polypeptidescontained byDNV1,DNV2, and
adeno-associated virus type2(AA V2)a Mol wt(+standarddeviation)of: Virus Individual polypep- Total poly-tide peptides DNV1 107,300±300 281,900
70,900±400 61,100±100 42,600±300
DNV2 109,600 ±200 281,000 70,800±300
58,700±300 41,900±400
AAV2 107,300±200 263,300 89,100±200
66,900±100
aThe peptides were resolved on 12.5%
polyacryl-amide gels. ,B-D-Galactosidase (molecular weight, 132,000), phosphorylase (molecular weight, 97,400), transferrin (molecular weight, 90,000), bovineserum
albumin (molecular weight, 69,000), and ovalbumin (molecular weight, 46,000)wereusedasstandards.
polypeptides (VP1 and VP2) predominantlyand
aminorpolypeptide intermediate in molecular
weight between VP1 and VP2 of both virus
particles and top components. This
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.510.131.203.366.534.2] [image:5.510.273.463.394.554.2]DNV STRUCTURAL PROTEINS 229
MIGRATION DISTANCE (mm)
FIG. 4. Densitometric traces of the polypeptides contained by DNVparticles and top components
stained with Coomassie blueafter electrophoresison a10%SDS-polyacrylamideslabgel. (A) DNVIvirus particles; (B) DNVItop components;(C) DNV2virus
particles; (D)DNV2 top components.
stratesthatVP1, VP2, and the minor polypep-tide have accessible surface tyrosine residues, which may mean that these polypeptides are located predominantly at the exterior of the components. VP2 wasiodinated less efficiently invirusparticles than intop componentswhen lactoperoxidase wasused to iodinate, and this
may indicate a conformational difference be-tweenvirusparticlesandtop components.
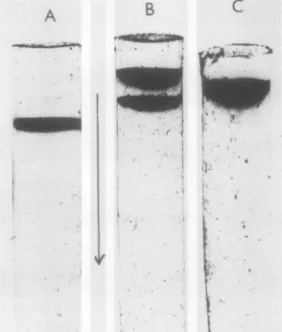
Acrylamide gelelectrophoresis ofintact
virus particles and topcomponents. Electro-phoresis of mixed DNV1 particles andtop com-ponentson3% acrylamide gelsatpH 9.0showed
two well-separated bands (Fig. 6), the faster moving with twice the mobility of the slower. Purified virus particles andtop components, re-spectively,comigrated with the bands migrating rapidly andslowly towards the anode; electron microscope observation of particles recovered from thebands detected inanatural mixture of virus andtop componentsconfirmed that virus particles migrated more rapidly. The rate of migrationwasdirectly proportional to the cur-rent applied. It also appeared that the propor-tion of topcomponent tovirus in partially
puri-fiedpreparationswas3:1 (as assessed by scan-ning negative photographs of Coomassie blue-stained gels), confirming earlier estimates (20). Thus, both virus andtop componentwere neg-atively charged at pH 9.0, the former having twice thenetcharge of the latter.
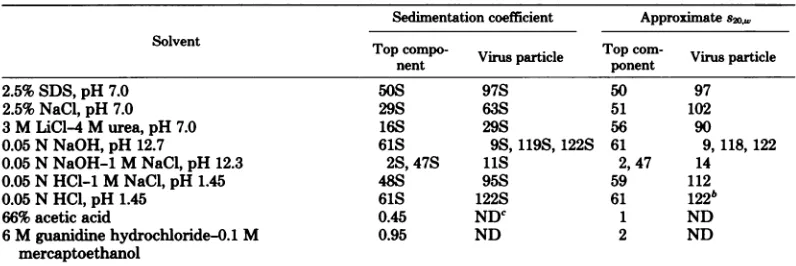
Dissociation of virus particles and top components of DNV1.Experiments dissociat-ing DNV1 particles and top components were carriedout togive information about forces sta-bilizing virus particles andtop componentsand toenablemeaningfultitrationstudiestobe car-riedout(seebelow). The virus proteinwas dis-sociated by incubating the material with an equal volume of twice-concentrateddenaturant at30°C for 24 h. Table 3 showsasummaryof
theeffects ofavariety ofdenaturing conditions onthe sedimentationproperties.
2.5% SDS, 2.5 M NaCl, and 3 M LiCl-4 M urea failed to break down DNV particles and top components and so were not suitable for preparing protein subunits from the virus.
Treatment with either 66% acetic acidor6 M guanidine hydrochloride-0.1 M 2-mercaptoeth-anolsuccessfullydissociatedtop componentsto
[image:6.510.56.235.65.498.2]low-molecular-weightcomponents(Table 3).
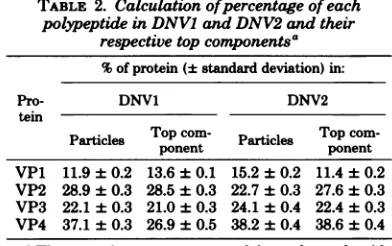
TABLE 2. Calculationofpercentageof each
polypeptideinDNVIand DNV2 andtheir
respectivetopcomponentsa
%ofprotein(±standarddeviation)in:
Pro- DNV1 DNV2
tein
Particles Topponentcom- Particles Top com-ponent VP1 11.9 ± 0.2 13.6 ±0.1 15.2±0.2 11.4±0.2 VP2 28.9 0.3 28.5±0.3 22.7±0.3 27.6±0.3 VP3 22.1 ± 0.3 21.0±0.3 24.1±0.4 22.4±0.3 VP4 37.1 ± 0.3 26.9±0.5 38.2±0.4 38.6±0.4 'The proteins were separated by polyacrylamide gelelectrophoresis,detectedbyCoomassie blue stain-ing, and scannedonaJoyce-LoeblChromoscan with anintegratorattachment.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.510.252.448.502.625.2]A
B
,,
ii
III iii
RELATIVEMiWATICN
FIG. 5. DensitometrictracesofDNV2
125I-Iabeledpolypeptides
resolvedbyautoradiographyof10% SDS-polyacrylamidegels ofvirusparticles (A)and top components(B)iodinatedbychloramine-TafterdisruptionbyheatinginSDS(i),lactoperoxidaselabelingofintactvirus(ii),and chloramine-Tlabelingofintact virus (iii).Note thatthe three
high-molecular-weightpolypeptides
wereradiolabeled in intactvirus.A
B C
FIG. 6. Polyacrylamide gel electrophoresis of
in-tactDNVIparticlesandtop componentson3% cylin-dricalgels. (A) Virusparticles; (B) top component andvirusparticles;(C)top component.
Ahigh pHhad a moremarkedeffect on virus
particlesthanontop components. Thetwo
en-tities (at =1mg/ml) weredialyzedinto 0.05 N
NaOHatpH 12.7or0.05NNaOH-1.0 M NaCl
atpH 12.3 at40Cfor48 h.The productswere
thenanalyzed byanalytical ultracentrifugation,
using Schlieren optics. Inlowsalt athigh pH,
thetopcomponent sedimentedas asymmetrical
peak with a sedimentation coefficient of 61S,
suggesting that it was
essentially
unchanged,whereas the virus particles sedimentedasthree
peakswithsedimentation coefficientsof9,119,
and 122S. In 1.0 MNaClandhighpH, thetop
componentsedimentedastwocomponentswith
sedimentation coefficients of 2 and 47S (the
latter wasthemajor component),andthe virus
particle sedimented as a single peak of 12S.
Thus, thetop components werefairly stableto
alkalineconditionsupto
pH
12.5, whereas undersimilarconditions ofhighpH and ionic strength,
virusparticleswereless stable thantop
compo-i I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.510.90.432.56.402.2] [image:7.510.95.236.459.625.2]DNV STRUCTURAL PROTEINS 231
TABLE 3. Dissociation ofDNVparticlesbyvariousdenaturing solvents: effects onsedimentation
coefficienta
Sedimentation coefficient Approximates20,w
SolventTocop-Tpom
Top comVo-
virus
particleTop
con-
Virus
particle2.5%SDS, pH 7.0 50S 97S 50 97
2.5%NaCl,pH 7.0 29S 63S 51 102
3MLiCI-4 M urea, pH 7.0 16S 29S 56 90
0.05 NNaOH, pH 12.7 61S 9S, 119S,122S 61 9, 118, 122
0.05NNaOH-1 M NaCl, pH 12.3 2S, 47S 11S 2, 47 14
0.05 NHCl-1MNaCl, pH 1.45 48S 95S 59 112
0.05 NHCl,pH 1.45 61S 122S 61 122b
66%acetic acid 0.45 NDC 1 ND
6Mguanidinehydrochloride-0.1 M 0.95 ND 2 ND
mercaptoethanol
aThe sedimentationcoefficients of virus particles and top components in 0.01 M phosphate buffer-0.1 M
NaCl,pH7.0, are117and 59 respectively (8). b Trace amounts detected.
cND, Not done.
nents. Thestabilities of both virus particles and
top components at a particular high pH
de-creased with increasing ionic strength. The
sta-bility oftopcomponent wasthuslittle affected
by the titration of basic groups onthe surface,
whichcaused theparticletohave a netnegative
charge. It is
likely
thatsomeof the stability oftopcomponentarises fromstrong
hydrophobic
protein-protein interactions. The virus maybe
less stable thantop component at apHgreater
than12duetotheionization of the bases in the
DNA.
Samples
of virusparticles
andtopcomponent,each
containing
1.2mg/rnl,
weredialyzed
at40C
for 60 hinto 0.05 N HCl, pH 1.45, and0.05 N
HCl-1.0 M NaCl at pH 1.45. The samples of
virusparticles and the corresponding samples of
top component werethen sedimentedinpairs in
theanalytical ultracentrifuge.AtlowpH and in
high
salt,
virusparticles
andtopcomponentbothsedimented as single
symmetrical
peaks withsedimentation coefficients of 112 and 59S,
re-spectively. Littleornodissociationhadoccurred
in either virus particles or top component. At
lowpH and in low
salt,
topcomponentshowedsomedissociationto
slow-sedimenting material,
but the main component was still
apparently
intact top component, with a sedimentation
coefficient of 61S. In contrast,
only
a trace ofintact virus
particles
was present in thecorre-spondingvirus
particle
sample. Thus,topcom-ponent isstabletoacidic conditions down topH 1.45. Under similar conditions of low pH and ionicstrength,virusparticlewasnomorestable
and
probably
less stable than top component.The stabilities of both virus particles and top
componentatalowpH decreasedwith
decreas-ing ionicstrength. The stability of top compo-nent waslittle affectedbythetitration of
side-chain carboxyl groups on the surface, which
caused the
particle
tohavea netpositive charge.Again,someof thestability oftopcomponentis
probably
duetohydrophobic protein-protein
in-teractions. Virus
particles
might be lessstablethan top component atpH 1.45if the DNA was
partially
accessible to protonsand,
hence, ifsome of the
negatively
chargedphosphate
groups onthe DNAwereneutralizedbyprotons.
ThepK of these
phosphate
groupsis about 1.0,and thusthey arenot normally titrated.
How-ever, apH of1.45might be low
enough
forsomeof these
phosphate
groups to be neutralized.Such neutralization
might
causedisrupting
forces sufficient to overcome the
stabilizing
forcesand, hence, dissociation of virus
particles
under conditions in which top component is
stable.
Alternatively,
ionization ofsome of thebasesontheDNAcould make ita
destabilizing
influence.Theeffect of ionicstrengthagain
sug-gests that ionic forces are
partly
involved indestabilizing
both virusparticles
and top com-ponent atthispH.Continuous electrometric
titration.Top
componentsafter exhaustivedialysis
into 0.1 MKCI hadapH of6.0.Itwasassumed that neither
K+ nor C1- ions bound
preferentially
to top components,andhencethispH
wastakentobe thepoint
ofzero netprotoncharge
for the pur-pose ofinterpreting
the electrometric titrationcurve. The titration curve of top components
obtained by
forward-titrating
samples
in 0.1 MKCI with 0.1 NHCIand then
back-titrating
with 0.1NNaOHisshown inFig.
7.Top
componentswere
partially
precipitated
betweenpH
4and6. Thetitrationcurves werereversiblebetweenpH
2.5and 6.0 and
partly
reversible,
butwithtime-dependent changes,in the
pH
range6.0 to 11.5.The acid endpoint waswell defined at
pH 2.5,
on November 10, 2019 by guest
http://jvi.asm.org/
o 2 m
oOH
OH
m0
OH U+ O
U)
8
4
4
2 4 6 8 10
pH
FIG. 7. Forward electrometric titration ofDNVI
top componentsin 0.1 MKCI.
butthe alkaliendpoint hadnotbeenreachedby pH 11.5. Titration curvesofanequivalent vol-umeof 0.1MKC0 wereobtained, and the num-ber ofH+ ions bound by the proteinwasobtained in the usualway.
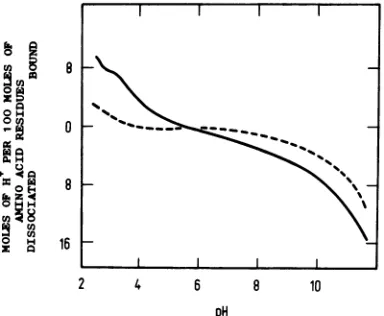
Apreliminary analysis of thegroupstitrated
was made as described by Tanford (32). Seven moles of H+ ions per 100 mol of amino acid residues were bound between pH 6.0 and 2.5. These may be assigned toside-chain carboxyl residues of aspartic acid and glutamic acid which have pK values of =4.0. The number of histidine, lysine, and arginine residues per 100 mol of aminoacid residues available for titrationabove pH 6.0 must therefore be seven, since at the isoionic pH (6.0)thenetnegativechargeequals thenet positive charge (32). Since 1 molofH+ ion per 100 mol of amino acid residues was dissociated betweenpH 6.0 and 7.3, presumably
from histidine, a total of 6 mol of lysine and
arginine residues per 100 mol of amino acid residueswas available fortitration. Amino acid analysis has shown that there is between 6 and 7mol oflysine and arginineper100 molof amino acidresidues inthe protein oftop components and virus particles (see Table 1 and reference 20); it islikely, therefore,thatallthese residues
arepositivelychargedatpH 6.0 andhence
avail-able for neutralizing negative charges on the DNA invirusparticles.
Between pH 7.3 and 11.5, between four and five H+ ions were dissociated per 100 mol of aminoacidresidues. The sidechains titrating in thisrangewouldhave been fromsome,butnot
necessarily all, of the cysteine, tyrosine, and lysine residues. The arginine residues wouldnot
beexpectedtotitratebelow pH12.Since only7 ofthe20aspartic acid andglutamic acid residues (20)per100molof aminoacid residues titrated,
13 of these residues in top component were
present in the amide form. Thus,all thelysine
and arginine residues in top componentare
ti-tratable andhencearepotentiallyavailable for
neutralizing negative charges on the DNA in virusparticles.
The forward titrationcurve of virusparticles
in 0.1 M KC0 is shown in Fig. 8. Since virus particles hadapH of 5.7 afterdialysis into0.1 MKCl,this wastaken asthepoint ofzeronet proton charge. From pH 2.5 to 5.5, virus was
partially precipitated. In contrast to top
com-ponents, the acid endpoint for virus particles wasnotreachedby pH 2.5. Between pH 3.0 and
5.7, 7 mol ofH+ ionper100mol of aminoacid
residues was dissociated. For top components
thecorresponding value between pH 2.5 and 6.0
was also 7 mol. Between pH 2.5 and 3.0, two
extra H+ ions were apparently dissociated per
100 mol of amino acid residues in the virus. Between pH 5.7 and3.0, therefore,thenumber oftitratablegroupsofvirusparticlesper100mol of amino acid residueswasthesame asthatof topcomponents, but below aboutpH 3.0 there
wereextratitratablegroups onvirusparticlesas
compared with top components. The alkali titra-tioncurveof viruswascarried outin mM KC1.
Fifteenorsixteen moles ofH+ ionper100 molof amino acid residues was dissociated for virus
particles between pH 5.7 and 11.5. The
corre-sponding value fortop componentswas 6 mol.
Clearly, thereareextratitratablegroupsinvirus particles as compared with top components in thispHrange.
Electrometric titrationswerecarried out un-der conditionswhich maintainedtop component
oUX
8uz
oQ.
OH
o H
Q
,o +
H
H
0
8
0
8
16
2 4 6 8 10
[image:9.510.69.261.49.234.2]pH
FIG. 8. Forward electrometric titration of DNVI particles ( ), pH 2.5to5.7 in 1.0 MKCI and pH 5.7to11.5 in 0.001MKCI. Thedifference (---) is also shownbetweentheforward electrometric titrations of top components(Fig. 7) andvirusparticles.
I I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.510.273.465.445.603.2]and virus
particles
intact. Thiswas confirmedby analyzing the titrated samples atboth high
andlowpHvalues inthe centrifuge. Inallcases
intact virus and top component wereobserved.
Therefore,anydifferencein the titrationcurves
betweentopcomponent and virus particles
can-not be due todissociation effects.
Thus, the electrometrictitrationexperiments
on top component (Fig. 7) and virus particles
(Fig.8)showed thatalargernumber ofhydrogen
ions per 100 mol of amino acid residues was
dissociated from intact virus than from intact
top componentinthe pHrange 2.5 to 11.0. The extratitratablegroupsof virus particles,as
com-pared with top component, weredissociated in
thepHrange 6.0 to 11.0andpossibly also inthe
pH range 2.5 to 3.0. No extra groups were
ti-trated from virus particles in the pH range 3.0
to6.0, ascompared withtop components.These
observations werenotduetothe differentionic
strength conditions used for the titrationsinthe
alkalinerange,since the effect ofincreasing ionic
strength would betodecrease the number ofH+
ions dissociatedathighpH (34).Thisis contrary
to the effect found. The difference in titration
curvefor virusparticles andtop component (see
Fig.8)suggested thatatpH greater than 6.0, the
twotypesof
particles
havedifferentcharges andhence different electrophoretic mobilities. We
showedearlier that virusparticles hadamobility
about twice that oftop component atpH 9.0.
Themobility difference wasconsistent with
vi-rus
particles'
having a negative charge abouttwicethat oftop componentatthispH. This is
confirmedby the titrationcurves.
The difference in titration behavior (Fig. 8)
for virus
particles
and top component may beascribed to titration of the DNA bases in the
virus. Thefour nucleotide bases ofDNAprovide
three titratable amino groups with pK'near 4
and two titratable
hydroxyl
groups with pK'near 11. The fact that little or no virus DNA
titrated between pH 3.0 and 6.0 but that viral
DNAapparently titratedinthepHrange 6.0 to
11.0 means thatthe titrationcurve for the nu-cleic acid invirusparticlesisshiftedto a lower pH range compared with DNA in
moderate-ionic-strength solutionsand issimilartothatof
denatured, and hence essentially
single-stranded, DNA under conditions ofvery high
ionicstrength (4). Thus, the DNA in virus par-ticles appears from its titration curve to be in an environment equivalent to a very high ionic
strength,where the netchargeonthephosphate
groups isalmostcompletely screened, aswould
be the case if the phosphates were
efficiently
neutralized
bycounterions.The nearequivalenceof the isoionicpHof the virus and topcomponent leadstotheimportant
conclusion that the net charge on the protein
doesnot change when it is combined with the
nucleic acid. This means that the negatively
charged phosphate
groups(pK
1)
ontheDNAmust be balanced by an equivalent number of
positivecounterionsin thevirus particlewhich
do not come from the viral protein. In other words, the majority, if not all, of the positive
chargeson the viralproteinare notinvolvedin
neutralizing DNA phosphates. These must be
neutralized, therefore, by small counterions
present within the virus particle. The
electro-static interaction between chargesontheprotein
and DNAappears tobeminiimal.
Acomparison of the titration curvesof virus
andtopcomponentshows that theprotein
side-chain
carboxyl
groupshaveverysimilar pKval-uesin bothcomplexes. They areprobably
situ-atedonthe surface of theprotein capsid andare
probably responsible for the solubility of the
virus in aqueous solution. Similar conclusions
mayalso be drawnconcerningthearginine,
his-tidine, and lysine side chains. These, too, are
freely accessible to H+ ionsin both complexes
and may therefore be situated on the outer
surface of theprotein capsid.
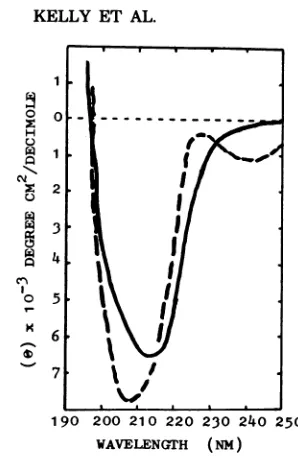
Circular dichroism studiesonDNV2
par-ticles andtop components. The circular
di-chroicspectraofDNV1, DNV2, and the
respec-tivetopcomponents intherange 230to320nm
have already been reported (11). The circular
dichroicspectra inthe195- to250-nm
region
ofvirusparticles andtop components areshown in Fig.9. The spectra were
significantly
different.Therewere decreases in negative dichroismof
the top component between 230 and 215 nm
when theproteinwasincorporated into the virus
capsid
and increasesintherange 215to198nmwhich might at first sight be
interpreted
aschangesin the secondary structure of the
pro-tein.However,itis also
possible
that thechanges
arosefrom the alteredenvironment of side-chain
aromatic chromophores, such as
tyrosine
ortryptophan, which might have interacted with
the DNA in the virus, or fromaltered
confor-mation of thebases inthe DNAitself.Itisnot
possible to distinguish between these various
possibilitiesonthe basis ofthese spectraalone.
It can be concluded, however, that there is a
definite interactionbetweenviralDNAand
pro-tein whichgives risetoalteredoptical properties.
DISCUSSION
DNV1and DNV2 are
similar,
butnotidenti-cal,virusesas
judged by
biophysical,
biochemi-cal, and
serological properties
of theprotein
coat. This basic conclusion
complements
the demonstration that the viruses possessclosely
homologousDNAsofsimilarstructure
(13).
on November 10, 2019 by guest
http://jvi.asm.org/
Cl)
l,
rz075 .
190 200 210 220 230 240 250 WAVELENGrH (NM)
FIG. 9. Circular dichroism spectra at room tem-peratureof DNV2 virus particles(-) andtop com-ponents( ).
Thetwoviruses show typical parvovirus
mor-phologyand size(9),althoughnodetailed
struc-ture could be determined. We were unable to confinn thededuction of Kurstak and Cot6 (18) that thevirus contains 42capsomeres.
Thepolypeptide compositions ofagiven DNV and itscorresponding top componentare iden-tical, and both containatleastfourpolypeptides.
The sum (265,000) ofthe molecularweights of
thepolypeptidesexceedsthecodingcapacity of
agenomewithnooverlappingorsplicedgenes the molecular weight of the DNV genome is
approximately 1.95 x 106, and so itpotentially
codes for about 2 x 105daltonsofprotein.
Pre-liminary peptide mapping experiments show
that thepolypeptides ofDNVsareinterrelated (N. F. Moore and D. C.Kelly, unpublished data).
Calculations show that the relative amountsof the various polypeptides are not easily accom-modatedinvirusparticles, assuming that parti-cles containing plus and minus strands have identical polypeptide compositions. The virus contains about 60 polypeptide molecules, of which about half are the major polypeptide
(VP4).The relativeamountsofeachpolypeptide
present differ considerably from the amounts calculatedby Tijssen etal. (33). This probably reflects theirunconventional method of calcu-lation. Under conditions which resolve three
polypeptidesforadeno-associatedvirusparticles
(30), four DNVpolypeptideswereresolved,and so itappearsthat thesevirusesareindeed dif-ferent despite similarities in the finestructures oftheirDNAs.Iodinationexperiments with both
J.
lactoperoxidaseandchloramine-T showedthat
only
the two high-molecular-weightpolypep-tides anda minor polypeptide were accessible
foriodination.Thisobservationagaincontrasts
with observations on the iodination of
adeno-associatedvirustype3capsid protein (22),where
allthreeadeno-associatedviruspolypeptidesare
iodinated.
However,
theobservationthatchlor-amine-T failstoiodinatethe two
lower-molecu-lar-weight
polypeptides does not necessarilymean that these are "core" polypeptides. It couldindicatemerelythat in the native
protein
state,theirtyrosine residuesareinaccessibledue
to tertiary folding of the polypeptide chain.
Omission ofa reducingagentfrom the sample
buffer failedto alter the polypeptide profileof
DNA polypeptides, confirmingthe observation
of
Tijssen
et al. (33), whichdemonstrates thatdisulfide bondsarenotinvolvedin thecreation
of theremarkablystable coat of these viruses. Gel
electrophoresis
ofintact virusparticlesatpH9.0 showedthat, in the case ofDNV1 (but
not DNV2, which failed to enter the gel), the
virusparticlescould beseparated fromtop
com-ponentandindicated that the virusparticleshad
a
higher
net negative charge than did the topcomponent; this isprobably attributableto the
negative chargeoftheDNAin thevirusarising
fromtitrationof theDNAbasesatpH9.0.
The conclusionthat most of theDNA
phos-phateisneutralized by cations other thanbasic
amino acids issupported by ourearlier obser-vation that the virusparticles,but not the top
component, are rich in polyamines, having a
sufficient amounttoneutralize 26% of the DNA
(15).
Thebiochemical,biophysical, and serological
tests described so far in this discussion have
failed to show that there is any difference
be-tweenthe top component
protein
and thepro-teincoatin situ in the virusparticle foragiven
virus. Althoughwehavepresentedindirect evi-dence that there is little direct interaction be-tweenthe viralgenome and itsprotein,
never-theless it is probable that interaction occurs,
albeiton alimitedscale, and thattheinteraction
hasacertain
specificity.
Todetermine whetheraconformational differenceoccurredin thecoat
whenpresent in topcomponentorin part ofthe
virus particle, circular dichroism studies were
performed, and differencesweredetected.
Anal-ysis of theresultsbystandardmethodsfailedto
demonstrate that the virusproteinexistedina
predominant
secondary
structuralform (similarobservations have been made for other
single-stranded RNA viruses[11, 28,31].Interactionof
single-stranded nucleic acid with the protein
induced amarked change in optical properties
of theprotein,nucleicacid,orboth.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.510.84.233.52.280.2]235
Theanalyses of the titration data suggest that
theelectrostatic interactions between DNA and
protein in the virus are minimal. The negative
charge on the DNA appears to be completely
screened by small counterions, such as
polya-mines.Althoughnonpolar interaction cannot be
excluded, and indeed thedifferences in the
cir-cular dichroic spectra between top component
and virus would support such interaction, the
picture that emerges is one where the DNA is
physically
contained within the protein, withlittle
direct interaction between them.ACKNOWLEDGMENTS
Wethank C.F.Rivers,M. H.Bew, M. D. Ayres, M. K. Arnold,andT. Lescottfor technicalasaistanceand R. H. Pain and R.Burgessfor use of theirDichrographe HI.
LITERATURE CITED
1.Barwise,A.H.,andL. 0.Walker. 1970. Studies on the DNA of a virus fromGaUeriamellonella.FEBS Lett. 6:13-16.
2. Bourguignon, G. J., P. J. Tattershall, and D. C. Wood.1976.DNA of minute virus of mice:selfpriming non-permutedsingle stranded genome with a 5' termi-nalhairpinduplex.J.Virol.20:290-306.
3. Burton, K. 1956. Astudy of theconditions and mecha-nism of the diphenylaminereaction for thecolorimetnc estimation ofdeoxyribonucleicacid. Biochem. J. 62: 315-323.
4.Cox, R.A.,andA. R.Peacocke.1956. Electrometric titration of thesodium salts ofdeoxyribonucleicacids. HI. Theeffectof sodiumchloride. J. Chem. Soc., p. 2499-2512.
5. Doty, P.,and J. T. Edsall. 1951. Light scattering in proteinsolutions. Adv. ProteinChem.6:35-121. 6. Eggstein,M.,andF. U. Kreutz. 1955. Protein
determi-nations.Klin.Wochenschr.33:879-889.
7. Elliott,R. M., T.Lescott,andD.C.Kelly.1977. Sero-logical relationships of an iridescentvirus (type 25) recently isolatedfromTipulaap. with two other
irdes-centviruses(types2and22).Virology81:309-316. 8.Fairbanks,C.,T.L. Stock, andD. F.H.Wallach. 1971.
Electrophoretic analysisofthe majorpolypeptidesof the humanerythrocytemembrane. Biochemistry10: 2606-2617.
9. Gerry, H. W., T. J. Kelly, and K. L. Berns. 1973. Arrangement ofnucleotidesequences in adeno-associ-ated virus DNA. J.Mol.Biol. 79:207-225.
10. Hoggan, M. D. 1971.SmallDNAviruses,p. 43-79. In K. Maramoroschand E. Kurstak(ed.), Comparative virol-ogy.AcademicPress,Inc.,New York.
11.Isenberg, H.,R. I.Cotter,and W. B.Gratzer. 1971. Secondarystructureand interactionof RNA and pro-teinin abacteriophage.BiochimBiophys.Acta 232: 184-191.
12.Jones,A.S.,W.Lee,and A. R.Peacocke.1951. Deter-mination of thephosphorous content of sodium deoxy-ribonucleate. J. Chem.Soc.,p. 623.
13.Kelly,D.C.,A. H.Barwise,and I.0.Walker. 1977.
DNAcontainedbytwo densonucleosisvinruses.J. Virol. 21:396-407.
14. Kelly, D.C., and H. K Bud.1978.Densonucleosis virus DNA: analysis of fine structure by electronmicroscopy and agarose gelelectrophoresis.J. Gen.Virol. 40:33-43. 15. Kelly, D. C., and R. M.Elliott. 1977.Polyamines con-tained by two densonucleosis viruses. J.Virol. 21:408-410.
16. Kelly, D. C., and T. W.Tinsley. 1972. The proteins of iridescentvirustypes 2 and 6. J.Invertebr. Pathol. 19: 273-275.
17.Kurstak,E. 1972.SmallDNAdensonucleosisvirus. Adv. Virus Res.17:207-242.
18. Kurstak,E.,and J. R.Cote. 1969.Proposition de clas-sification de virus de ladensonucleosebaseesurl'etude delastructuremol6culaireetdespropri6tes physioco-chimiques. C. R.Acad. Sci.268:616-619.
19. Laemmli,U. K. 1970. Cleavage of structural proteins duringtheassemblyof theheadofbacteriophage T4. Nature (London)227:680-685.
20. Longworth,J.R.,T.W.Tinsley,A H.Barwise, and I.0.Walker.Purification of a non-occludedvirus of Galkria mellonella.J.Gen. Virol. 3:167-174. 21. Lowry,0.H.,N. J.Rosebrough, A.L.Farr, and R. J.
Randall. 1951. Proteinmeasurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
22. Lubeck, M. D., and F. B. Johnson. 1977. Radiodination ofadenovirus-associatedvirusexternal structural pro-teins.Virology83:453-457.
23. Mejbaum,W. 1939. Z.Phys. Chem.(FrankfurtamMain) 258:117-120.
24.Meynadier, G., C. Vago, G. Plantevin,and P.Atger. 1964. Virose d'un typehabituel chezleLepidoptera Galleria mellonellaL. Rev.ZooLAgric. Appl. 63:207-209.
25. Moore, N.F.,and D.C.Kelly.1974.Sendai virus struc-turalproteins:analysisofpolypeptideslinkedby disul-phide bonds. Intervirology2:128-133.
26.Moore,N.F.,J. M.Kelley,andR. R.Wagner.1974. Envelopeproteins of vesicular stomatitis virions: acces-sibilitytoiodination.Virology61:292-296.
27.Peacocke,A. R., and I.0.Walker.1962.Thethermal denaturationof sodium deoxyribonucleate. III.Light scatteringstudies. J.Mol.Biol.5:564-569.
28.Piazzolla, P., V. Guantieri, C. Vovlas, and A. M. Tamburro.1977.Circulardichroism studies ofchicory yellowmottlevirus.J. Gen.Virol.37:359-372. 29.Rivers, C. F.,and J. F.Longworth. 1972.A
non-oc-cludedvirusofJunoniacoenia (Nymphalidae:Lepidop-tera). J.Invertebr. Pathol.20:369-370.
30. Rose, J.A.,J. V.MaizeLJr.,J. K.Inman,and A. J. Shatkin.1971.Structuralproteinsof adenovirus-asso-ciatedviruses.J.Virol.8:766-770.
31.Tamburro,A.M.,V.Guentieri,P.Piazzolia,and D. Gallitelli.1978.Conformationalstudiesonparticlesof turnipyellowmosaicvirus.J.Gen.Virol. 40:337-344. 32.Tanford, C. 1962. Theinterpretation ofhydrogen ion
titrationcurvesofproteins.Adv. ProteinChem. 17:70-165.
33.Tijssen, P.,J.vandenHurk,and E.Kurstak.1976.
BiochemicaLbiophysical, andbiological propertiesof densonucleosisvirus. I.Structuralproteins.J. Virol. 17:
686-691.
34.Walker,L.0.1965.Theelectrometric titrationcurvesof histone and nucleohistone. J. Mol. Biol. 14:385-393.