0022-538X/91/094636-09$02.00/0
Copyright © 1991,American SocietyforMicrobiology
Human
Immunodeficiency Virus
Integrase
Protein Requires
a
Subterminal
Position of
Its
Viral DNA
Recognition
Sequence for Efficient Cleavage
CORNELIS VINK, DIK C. VAN GENT, YPE ELGERSMA, AND RONALD H. A. PLASTERK* Division ofMolecular Biology, The Netherlands Cancer Institute,
Plesmanlaan 121, 1066 CX Amsterdam, The Netherlands Received 28 November1990/Accepted24May 1991
Retroviral integrationrequires cis-acting sequences at theterminiof linear double-stranded viral DNA and aproduct of theretroviral pol gene, the integrase protein (IN). IN isrequiredandsufficient forgeneration of recessed3' termini of theviral DNA (thefirststepinproviralintegration)and forintegration of therecessed DNAspecies in vitro. Human immunodeficiencyvirus type 1 (HIV-1) IN, expressedinEscherichia coli, was purified tonear homogeneity. The substrate sequence requirements for specific cleavage andintegration of retroviral DNA were studied in a physical assay, using purified IN and shortduplex oligonucleotides that correspond tothetermini ofHIV DNA. Afew point mutationsaround the INcleavagesitesubstantiallyreduced cleavage; mostother mutations did not have adrastic effect, suggestingthat the sequencerequirements are limited. Theterminal 15bp oftheretroviral DNAweredemonstrated tobesufficientforrecognition byIN. Efficient specific cutting oftheretroviralDNAbyINrequiredthat thecleavagesite,thephosphodiesterbond atthe3' side ofaconservedCA-3'dinucleotide,belocatedtwonucleotides awayfrom the end of the viral
DNA,
however, low-efficiency cutting was observed when the cleavage site was located one, three, four, or five nucleotidesaway from theterminus ofthedouble-stranded viral DNA. Increased cleavage byIN was detected when thenucleotides3' of theCA-3'dinucleotidewerepresentassingle-stranded DNA.IN wasfoundtohave astrong preference forpromoting integration into double-stranded ratherthansingle-strandedDNA.Efficient retroviral replication requires integration of a double-stranded DNA copyofthe viral RNA genomeintoa chromosome ofthe infected cell. Transcription ofthe
inte-grated provirus produces viral RNAs that function as ge-nomesof progenyvirionsor asthe template fortranslation of
viral proteins. Assembled virions bud from the cell mem-brane, and infection of other cells completes the retroviral replication cycle.
Studies ofintegration in the Moloney murine leukemia
virus(MoMLV), humanimmunodeficiency virus (HIV), and avian sarcoma-leukosis virus (ASLV) systems have indi-cated that thefollowing steps areinvolvedin theintegration
process.After reversetranscriptionandsecond-strand DNA
synthesis, a linear, double-stranded DNA molecule with
flush ends is formed. This DNA copy of the viral RNA contains atits termini direct repeat sequences, termed long
terminal repeats (LTRs). At the outer edges of the LTRs,
short, imperfect inverted repeats are present. These se-quences are necessary and sufficient for correct proviral
integration(6, 7).The two terminal nucleotides at the 3' ends
of the blunt-ended DNA are cleaved off, resulting in the putative immediate precursor for integration (2, 15, 26). This
cleavage reaction is executed by the viral integrase protein (IN), which is the only protein known to be required for
integration (3, 5, 8, 18, 19, 30). Inthe next step, the target DNA is cut in a staggered fashion and joined to the viral 3' OH ends (3, 8, 15, 18). The length of the staggered cut is virus specific (35). In the case of HIV integration,
presum-ablya5-bp staggered cut is generated, which is inferred from the characteristic 5-bp direct repeats in the target DNA that flankintegrated HIVproviruses (10, 21, 36). The last step in
*Correspondingauthor.
proviral integration is repair of the single-stranded DNA gaps presentin theintegration intermediate. Itispossiblyin this step that theterminal nucleotidesattheproviral5'ends are lost. The resulting provirus is now flanked by a short directduplication ofthe target DNAand terminates with the dinucleotides 5'-TG and CA-3'. These same dinucleotides
are also present at the termini of some eukaryotic and
prokaryotic transposable elements and in all retrotrans-posons.
The HIV-1 IN protein is encodednear the 3' end ofthe viral pol gene and isa32-kDaproteolyticcleavageproduct of
a gag-pol fusion protein precursor. Mutations in the
IN-coding regionhavenoinfluenceon reversetranscription,but
integration ofthe viral DNA canbe blocked (9,28, 34). An
understanding ofthe roleofthe INproteinin theintegration
processhas comefromthedevelopmentofdifferentinvitro integration assays. First it was shown that extracts from
retrovirus-infected cellswere able tointegrate endogenous
DNA aswell asexogenously added retroviral DNA (2, 10, 11, 14, 37). Thereafter it was demonstrated by a direct
physical assay that the IN proteins of HIV, MoMLV, and ASLV arethe only viral proteins that are required both to generate the recessed 3' ends of the viral DNA and to
accomplish theirintegration in vitro (4, 8, 18, 30). In this assay, oligonucleotides that represent the termini of the retroviralDNA were incubated with IN, and it was shown that besides specific cutting of the oligonucleotides, IN is alsocapable ofintegratingoneoligonucleotide into another. The INproteinof HIV-1 hasbeen overexpressed in insect cells (5). This protein was partially purified, and it was shown that it is able to promote integration of mini-HIV DNAinageneticassay. HIV-1 IN wasalsooverproduced in andpartiallypurifiedfrom Escherichia coli (4, 30). For this protein, it was demonstrated in a physical assay that it is
4636
on November 10, 2019 by guest
http://jvi.asm.org/
capable of generating the 3' recessed ends of the retroviral DNA. In this case, double-stranded, blunt-ended oligonucle-otides that represent the termini of the LTRs of HIV-1 DNA were used as substrate viral DNAs.
We overproduced HIV-1 IN in E. coli and purified it to near homogeneity. We show here that IN has specific
integrationactivity as well as specific cleavage activity. The sequence requirements for efficient specific cleavage and integration of HIV DNA were investigated by using a series of oligonucleotides that contain mutations relative to an
oligonucleotide that mimics the ends of HIV DNA. We
demonstratethat for specific cutting by IN, the cleavage site (the phosphodiester bond at the 3' side of the conserved
CA-3' dinucleotide) must be located at a subterminal
posi-tion in the viral DNA. We further investigated whether IN could carry out integration into single-stranded as well as double-stranded DNA.
MATERIALS AND METHODS
Construction of a plasmid expressing HIV-1 IN. The HIV-1 INexpression plasmid that we constructed is essentially the same as the plasmid described by Sherman and Fyfe (30). The HIV-1 IN-coding sequence was derived from plasmid
pHXB-2D (13). The IN gene was subcloned into plasmid
PiAN7 (29) to generate plasmid pRP177. The sequence 5'-ATAGAT-3' at positions 3822 to 3827 in the IN-coding sequence (24) was changed by oligonucleotide-directed
mu-tagenesis to the sequence 5'-ATCGAT-3', which is a ClaI
restriction site. In plasmid pRP177, the StuI site that is
locatedatposition4985 (24) was lost by fusion to the HincII
site of vector PiAN7. The adjacent BamHI site in the
polylinker ofPiAN7 and theClaI site intheIN-coding region
wereused to clone the IN gene into the NdeI and BamHI
sitesof theexpressionvectorpET-3c(25), together with the
syntheticoligonucleotide 5'-TATGTTTCTAGATGGAAT-3'
and its complement, having NdeI and ClaI cohesive ends. The ATG inthe oligonucleotide (in bold print) is the
trans-lation start codon and precedes the TTT codon, which encodes the actual INN-terminalaminoacid,phenylalanine.
In theresultingHIV-1 INexpressionvector,pRP274, the IN gene is under the control of a bacteriophage T7 RNA
polymerase promoter. E. coli BL21(DE3) was transformed withthe INexpressionplasmid.Inthisstrain,acopy ofthe T7RNApolymerasegeneispresentin thechromosomeand
isunder the control ofthe lacUV5 promoter.
PurificationofHIV-1IN. INwas purified asdescribed by
Sherman and Fyfe (30), with some modifications in the method ofgrowing and lysis ofthe bacteria. Bacteria har-boring plasmidpRP274 weregrownfor 16 hat37°C in LB
broth supplemented with 50 ,ug ofampicillin per ml. The culture was diluted 50 times in 1 liter of TB medium (27) containing 10 mM MgSO4 and 50 ,ug ofampicillin per ml. After 4 hofgrowthat37°C,isopropyl
P-D-thiogalactopyran-osidewas added to afinal concentration of 0.3 mM. Threehoursafter
induction,
cellswereharvestedby
centrifugation
and resuspended in cold 50 mM Tris-HCI
(pH
7.5)-S5 mMdithiothreitol-1mM EDTA-1 mgoflysozymeper ml. After
incubationon icefor15 min, thesuspensionwas sonicated. Thesonicextract wascentrifugedfor 1 hat
12,000
x g.Wefound that with these
modifications,
we obtainedvirtually
pure, solubleIN (see Results).
SDS-polyacrylamide gelelectrophoresis. Proteinswere sep-aratedon10%sodiumdodecylsulfate
(SDS)-polyacrylamide
gels (20). Gelsweresilver stained
by
using
asilver stain kit(Sigma Chemical Co.).
kD
1 2
97-66-~
43-31-
-22- OM
14-
-FIG. 1. HIV-1INexpressed in and purified from E.coli. Purified IN was subjected to SDS-polyacrylamide gel electrophoresis and silver stained(lane 2). Lane 1, molecular size markers.
Oligonucleotide substrates. Thesynthetic oligonucleotides
that weretested for specific cleavage and integration by IN aredepictedinFig.2to 4.Before labeling and annealing, the
oligonucleotides werepurifiedbyelectrophoresis ina
dena-turing 12% polyacrylamide (20:1 acrylamide/bisacrylamide)
gel. Ten picomoles of the oligonucleotide strand to be labeled with 32P at its 5' end was incubated with 40 ,uCi of
[.y-32P]ATP
(3,000Ci/mmol;AmershamCorp.) and 0.1 UofT4polynucleotide kinase (BoehringerMannheim) in a
reac-tion volume of10 ,ul. Afterincubation for1 h at37°C, 10
pLI
of formamide loading dye (95% formamide, 20 mM EDTA, 0.05%bromophenol blue, 0.05% xylene cyanol)wasadded. The samplewasincubatedat80°Cfor 3 min and loaded onto a12%denaturingpolyacrylamide gel.Afterelectrophoresis,an X-ray film (X-Omat AR; Eastman Kodak Co.) was exposed to the gel to localize the labeled oligonucleotide.
Theoligonucleotidewasthencutfrom thegel and eluted for 16 h at 37°Cin elution buffer (27). Afterprecipitation with
ethanol, the oligonucleotide was resuspendedin H20 toan endconcentration of0.2
pmol/I,l.
Then 1[lI
(10pmol) ofthe unlabeledcomplementary strandwasaddedto10RI
of thisoligonucleotide sample,and themixturewasheatedto900C. The DNAwas annealed by slowcooling toroom tempera-ture.
Cleavage andintegrationreactions.Reactionconditions for
specific cleavageandintegrationof the
oligonucleotide
sub-strates weregenerally asdescribed byCraigie
etal. for the MoMLV INprotein (8).Reaction mixtures(10,ul) contained75 mM
NaCl,
20 mMmorpholinepropanesulfonic
acid (MOPS; pH 7.2), 3 mM MnCl2, 10 mMdithiothreitol,
20%(vol/vol) glycerol, 100 ,ug of bovine serum albumin per ml,
0.2 pmol ofduplex
oligonucleotide,
andabout 50 ng of HIV INprotein. Thereaction mixtureswereat300C for1h,
and thereactionswerestoppedby addition of10,ulofformamide loading dye. Afterheating
to80°C
for 3 min, 5pul
of thesamples was loaded onto a 12%
polyacrylamide-8
M urea-1x TBE(89mMTris,
89 mMboricacid,
2 mMEDTA)
gel and electrophoresed.Thegelwas fixed
by
incubation in10% methanol-10% acetic acid for 30 min and dried on Whatman 3MM paper.Reaction
products
werevisualizedby
autoradiography.
RESULTS
Purification of HIV-1 IN. The IN
protein
of HIV-1 waspurified to near
homogeneity
essentially
as describedby
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.402.475.77.222.2]A 25 20 15 10 5 1
HlV-1 US 5t-TTAGTCAGTGTGGAAAATCTCTAGCAGT -3
EIIV-1 U3 TCGTTGGGAGTGAATTACCCCTTCCAGT
HTV-2 U5 AAACCGAGGCAGGAAAA'TCCCTAGCAGG lIV-2 U3 TCTTTCACTGTAAAACATCCCTTCCAGT
MoMIV U5 GACTACCCGTCAGCGGGGGTrCTTT1rCATT
HIV-1 U5 5t-TTAGTCAGTGTGGAAAATCTCTAGCAGT -3
mutants
(1--)>T---
---T-CA(2-1)>TG
---TG--A(1)>T ---
T--A)1)>G ---
G--C(2)>T
---T---C(2)>G
---(-;---ACG(4-3)>GA --- - .---
GA----T(5)>A
---A---C(6)>T ---
T---C(8)>G
---G---A(10)>G ---
G---AAATC(12-8)>GGGGG ---
----GGGGG---A(13)>C --
---c---T<->G,A<->C(26-14)
G,CTGACTGTGTT---MoMLV U15 mutants
T(j3)>G IT(4-3) >AG
GGGGG( 12-8)>AAA
C 13)>A
5'-GACTACCCGTCAGCGGGGGTlCIr'I'lCAEI-1
-_____AG
----XTC --- AAATC---A
---FIG. 2. Specific cleavageandintegration of oligonucleotide sub-stratesby HIV-1 IN. (A) Sequencesof the28-bpoligonucleotides
sp.cl.
it.
used and summary of results from the cleavage andintegration
assay. The HIV-1 sequences are from reference 38, the HIV-2 ++ ++ sequences are from reference 16, and the MoMLV sequences are
+ t4 from reference31.Numbers inparentheses refertopositions of the nucleotides shown at the top. Only one strand of the double-strandedsubstrates is shown. The 3' end of each strandrepresents - - theterminus ofthe viral DNA. The conserved dinucleotide CA-3' is indicated inbold print. sp.cl., specific cleavageatthe
phosphodi-ester bond 3' of the CA-3' dinucleotide. Specific cleavage was sp.cl.+ llt. scored when the band at the
position
on thegel'
of thespecific
productwasthe predominant product. Int., integration. ++, high 4 level of specific cleavage or integration; +, moderate level of t + specificcleavageorintegration; +/-,lowlevel ofspecificcleavage
+
t - or integration; -, nospecific cleavage or integration. (B) Specific 4+4 t cleavageandintegrationof thevarious substrates. The
oligonucle-++ + otideswereincubated in the presence (right lane) or absence(left ++ + lane) of HIV-1 IN. The top panels show a longer exposure of the
+ +/- region of the gel containing products longer than 28nucleotides, to ++ + identify integration products. Lengths of the oligonucleotide strands
areshown ontheright. Lanes contain the following substrates: 1, HIV-1 U5;2, HIV-1 U3; 3, HIV-2 U5; 4,HIV-2U3; 5, MoMLVU5; --- 6, AAATC(12-8)>GGGGG; 7, MoMLV T(3)>G; 8, C(8)>G; 9,
+ - - A(13)>C; 10,MoMLVGGGGG(12-8)>AAATC;11, MoMLV TT(4-+S_ _ 3)>AG; 12, A(1)>G; 13, C(2)>T; 14, A(10)>G; 15, G(-1)>T; 16, AG(4-3)>GA; 17, CA(2-1)>TG; 18, C(2)>G; 19, A(1)>T; 20, Mo-MLVC(13)>A; 21, C(6)>T; 22, T(5)>A; 23,T+-G,A+C(26-14).
I'
1 2 3 4 5 6 7 1 8 9 10 11
pw
48*
U'
I
i-H
,,
II 1L 13 14 13 lb 17 1I IY LU 1 LL 23
imp .4
Sherman and Fyfe (30). Purified IN is shown on a silver-stained SDS-polyacrylamide gel in Fig. 1. Onlyafewother minor bands can be detected in the IN preparation. The identity of the 32-kDa IN bandwas determined by immuno-blotting, using polyclonal antisera that were raised againstan oligopeptide that is identical to theC-terminal 11 amino acids ofHIV-1 IN (data notshown). Wefound that by using some modifications in the method ofinduction and lysis of the bacteria that express IN (see Materials and Methods), we obtained proteinpreparations that had ahigher puritythan
-28
those previously described (30).-26 Sequence requirements for specific cleavage and integra-tion. The sequence requirements for specific cleavage and
integrationof retroviral DNA by HIV-1 IN were determined in aphysical assay. Initially, the activities of HIV-1 IN were tested onsynthetic oligonucleotidesthatmimic the terminal 28bp of HIV-1, HIV-2, and MoMLV DNA. The sequences ofthese oligonucleotide substrates are shown in Fig. 2A.
Purified IN was incubated with the blunt-ended oligonucle-otides undertheappropriatereaction conditions (see Mate-rials andMethods), and the products were electrophoresed inadenaturing polyacrylamide gel. Reaction products were
visualized by autoradiography (Fig. 2B). As was shown
previously (4, 30), HIV-1 IN is capable of specifically
removingtwonucleotides from the 3' ends of the HIV-1 U3 andU5 termini.Afterincubation with IN, a major product of 26 nucleotides could be seen, in addition to the band
correspondingtothe substrate. A minor but clear product of 24nucleotides could also be detected, indicating that HIV-1 IN is also able to cleave, with a low efficiency, the phos-phodiester bond between the G nucleotide at position 3 and -28 the C nucleotide at position 2 of the U5 sequence and -26 between the two C nucleotides at positions 2 and 3 in the U3 sequence of HIV-1(Fig. 2A). The relevance of cleavage at these positions isunknown. The higher reactivity of HIV-1 U5 ends than of HIV-1 U3 ends, which has previously been described (4), was notseen; theHIV-1 U3 and U5 oligonu-B
I..
II-. m-
".-:1..f:
Sk.,AL.. .1;.L :z;."iiiis.l a .""Al IAAAW
fl. I..,:f .. :. I..::::,: T..
.7.f...f...lipillipi ... .-:..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.62.301.82.710.2]cleotides were cleaved at approximately equal levels (Fig.
2B, lanes 1 and 2). Oligonucleotides that represent the U3
andU5 ends ofHIV-2 werealsogood substratesforspecific
cleavage by HIV-1 IN; a major product with a length of 26
nucleotides could be detected in both instances. Reaction products that are longer than 28 nucleotides could be de-tectedafterincubationof IN withboththe HIV-1and HIV-2 substrates (Fig. 2B, upperpanel). Itwasshown forMoMLV IN (8), ASLV IN (18), and HIV-1 IN (4) that these bands represent productsofintegrationofoneoligonucleotideinto
another. Products that are shorter than 26 nucleotides can
also beexplained byintegration, becausethe 3' ends ofthe cutmade in the (5') labeled strandofthetarget DNAremain unjoined in the integration product. Specific cleavage and
integration were not observed when the oligonucleotides
were incubated with an extract from E. coli BL21(DE3) transformed with plasmid pET-3c (data not shown). This extract was subjected to the same purification procedure as was the HIV-1 IN preparation.
HIV-1 IN is not able to cleave the MoMLV U5 oligonu-cleotide with high specificity (Fig. 2). A nonspecific degra-dationladder couldbe observedwhenthe MoMLV substrate was incubatedwith IN.
We set out to determine the nucleotides present at the LTR ends ofHIV-1 that are required for specific cleavage
andintegration by IN. Forthis purpose, we tested oligonu-cleotides that contain mutations relative tothe oligonucleo-tide that represents the terminal 28 bp of the HIV-1 U5 sequence. The sequencesofthe different mutant substrates
areshowninFig. 2A.Theresultsobtained fromthecleavage
and integration assay are depicted in Fig. 2B and summa-rized in Fig. 2A. Mutations that are located in the vicinity
and at the 5' sideofthe INcleavage site(thephosphodiester
bond at the 3' side ofthe CA-3' dinucleotide at
positions
1 and 2 in Fig. 2A) all had areducing
effect onspecific
cleavage and integration. The most drastic effect was seen with the oligonucleotide that contains the CA(2-1)>TG mutation. This substratewas notcleavedatallbyIN. When the A nucleotide at position 1 and the C nucleotide atposition 2 were mutated
separately,
a reduced but still visible level ofspecific cutting was observed. Apparently,both positions are important, and the added effect of the double mutant
[CA(2-1)>TG]
is a total block ofcleavage.
Althoughintegrations
ofmutantsC(2)>G
andC(2)>T
wereobserved, no integrationswere seen ofthe
oligonucleotides
that are mutated at
position
1 [A(1)>T andA(1)>G]
(see Discussion). It should be noted that the changes that areintroduced into the mutant substrates could influence the
function ofthe
oligonucleotides
astargetsforintegration.
The A nucleotidesat
positions
10and13were alsoshownto be important for
cleavage
andintegration by IN;
the substrates A(10)>G andA(13)>C
were cleaved andinte-grated with a lower
efficiency
than thewild-type
substrate.Other point mutations did not reduce the
degree
ofsite-specific cleavageand
integration.
Whenall ofthenucleotidesat
positions
14 to 26 werechanged
[T+-G,A<-*C(26-14)
in Fig. 2A], no effect oncleavage
andintegration
could bedetected. This suggests that the terminal 15
bp
of theretroviral DNA are sufficient for
recognition by
IN(see
below).Toinvestigate whetherwecould
change
the MoMLV U5end sequence in such a way that it would become a target
for specific
cutting by
HIV-1IN,
we altered some of thenucleotidesin the MoMLV
oligonucleotide
tobases thatarepresent at similar
positions
in the HIV-1 U5 substrate and wereshown abovetobeimportant
forcleavage
andintegra-tion. For
example,
weshowed that theA(13)>C
mutationin HIV-1 U5 resulted inalower level ofcutting by
HIV-1 IN. The MoMLVU5
sequence contains a C nucleotide atposition
13(Fig. 2A).
Wechanged
this C nucleotide in the MoMLV U5sequence to an Anucleotide that ispresent at the sameposition
in the HIV-1 U5oligonucleotide.
Theresulting
oligonucleotide, C(13)>A,
was not agood
sub-strate for
cleavage
andintegration by
HIV-1 IN. The otherMoMLV mutantswere also not cleaved
efficiently by
IN. The resultspresented
here indicate that the conserved CA-3' dinucleotide isrequired
forspecific
cleavage
andintegration.
Mostother nucleotides atthe viral DNAends,
however,
arenotabsolutely
required
forspecific
INactivity.
The substrate
specificity
is thereforeprobably
determinedby
several nucleotidesatthe viral DNAterminithat all
contrib-uteto
specific recognition.
Thisexplains why
nosingle
point
mutation in the HIV-1 U5terminus
completely
blocked INactivity
andwhy
changing single
nucleotides oftheMoMLVU5end into nucleotidesthatare
present
at similarpositions
in the HIV-1 U5end did notresult in efficient
site-specific
cleavage
by
HIV-1 IN.Spacing
requirements
forspecific cleavage.
HIV-1 INnor-mally
cleavesthe 3' endoftheblunt-ended retroviralDNA at thephosphodiester
bond that islocated two nucleotidesaway from the 3' terminus. Since we showed that the sequence
requirements
forcutting
andintegration
are not very strong, we wanted to determine whether additionalspecificity
isprovided by
theposition
oftherecognition
site withregard
totheviral DNA termini. Wetestedwhether IN would tolerate additional nucleotides at the 3' end ofthe HIV-1 U5 sequence forspecific cleavage
andintegration.
We studieda series ofmutantdouble-strandedoligonucleo-tides that contain either no, one, two
(the
wild-type
situa-tion), three,
four, five,
orsix nucleotides3' of theconserved CA-3' dinucleotide. The sequences ofthese substrates aredepicted
inFig.
3A;
the resultsof thecleavage
andintegra-tionassayareshown in
Fig.
3B. Itis clear that all substrateswere cleaved at the
phosphodiester
bond that is located at the 3' side of the CA-3' dinucleotide. The blunt-ended substratethatdoesnotcontainanyresidues3'ofthe CA-3'(substrate
1/2)
cannotbespecifically
cleavedby
IN. How-ever,integrations
ofthis substrate couldbe detected(Fig.
3B).
Thewild-type
oligonucleotide,
having
twonucleotides(GT-3')
3' oftheCA-3'dinucleotide,
wasthe bestsubstrate forsite-specific cleavage
andintegration
by
IN.Specific
cleavage
gradually
decreased whenmore nucleotides wereaddedto the 3' end. The
oligonucleotide
having
six nucleo-tides 3' ofthe CA-3' sequence(substrate
13/14 inFig.
3B)
wasnota
good
substrateforcleavage.
Aspecific
bandattheposition
of26nucleotidescanbeseen, butthis bandis less intense than thenonspecific
bandsatthepositions
of31, 30,
29,
and 24 nucleotides. The substratecontaining only
one nucleotide(a
G)
3'ofthe CA-3' wascleavedlessefficiently
than thewild-type
substrate.Integration
events, as seen inFig.
3Bfrom theproducts
on thegel
thatarelarger
than 28nucleotides,
were mostfrequent
when thewild-type
sub-strate was used. The otheroligonucleotides
that have less than six nucleotides 3' of the CA-3' wereintegrated
atapproximately
equally
lowfrequencies. Integrations
ofthelargest
substrate(substrate
13/14)
werenotdetected.The
inhibitory
effectofadditionalbasepairs
atthe viral ends may result from steric hindrance of the addedse-quencesin the
recognition by
IN. Toinvestigate
thisfurther,
wealsotested substrates in which
only
oneDNAstrandwasextended. We first
investigated
whether HIV-1 IN would cleaveduplex
oligonucleotides
in which the sequence 3' ofon November 10, 2019 by guest
http://jvi.asm.org/
the conserved CA-3' dinucleotide was present as
single-stranded DNA. The substrate
containing
a 3' overhang of twonucleotides (substrate5/2)
wascleavedmoreefficientlythan its flush-endedcounterpart, the
wild-type
oligonucleo-tide
(substrate
5/6) (Fig.
3C). However,integration
wasslightly
reduced. The other substrates tested all showedsimilarpatterns of
cleavage;
when the sequenceatthe 3' sideof the CA-3' dinucleotide was present as single-stranded DNA,
cleavage
was greater than with the substrates thatcontaindouble-strandedextensions. Eventhesubstratethat contains sixbases 3' of the CA-3' sequence
(substrate
13/2)wascleavedandintegrated.Wetested whether the enhanced
cleavage
ofsubstrates that contain 3'protrusions
was the result ofspecific
INactivity
and not ofnonspecific,
3'single-stranded
exonucleaseactivity
causedby
contaminat-ing proteins
in the INpreparation.
For this purpose, thelabeled
single-stranded oligonucleotide
of30nucleotidesthatcontains fourbases at the 3' side ofthe CA-3' dinucleotide
(oligonucleotide 9)
was annealed with theoligonucleotide
that has the
wild-type
sequence(oligonucleotide 6).
IN-specific cleavage
ofthissubstrate would resultin aproduct of 26nucleotides,
whereasnonspecific
3'single-stranded
exonuclease
activity
would result in aproduct
of 28nucleo-tides. In
Fig. 3C,
amajor
band of 26 nucleotides can beobserved after incubation of substrate 9/6 with IN. This
finding
demonstrates that the enhancedcleavage
is indeedthe resultofIN
activity.
We also tested whether 5' single-stranded extensions in the DNA strand that is not cleaved
by
IN would influencespecific cleavage.
The labeledwild-type
strand of 28nu-cleotides
(oligonucleotide 5)
was annealed witholigonu-cleotides that are
longer
by
one(oligonucleotide
8), two(oligonucleotide 10),
three(oligonucleotide 12),
orfour(oli-gonucleotide
14)nucleotides,
and theresulting
substrates were incubated with IN(Fig.
3D). The level ofsite-specific
cleavage
andintegration (which
could be observed afterlonger
exposureofthegel;datanotshown)wascomparabletothe
wild-type
level in allcases.Consequently,
INdoesnot seemto be hinderedby
these 5'single-stranded
extensions.The
cleavage
andintegration
assays with the oligonucle-otides showninFig.
3A were alsoperformed
withpurified
HIV-2 IN. The
purification
of HIV-2 IN will be described elsewhere (34a). HIV-2 IN was indeed able tospecifically
cleave the HIV-1U5-derivedsubstrates and showeda cleav-age pattern similar tothat of HIV-1 IN (Fig. 3E). Also, in thiscasethe
wild-type
substratewascleavedmoreefficientlythan the other
oligonucleotides,
and cleavage decreasedgradually
whenmorenucleotideswereaddedtothe 3'sideofthe conserved CA-3' dinucleotide. Enhancedcleavage was seen when the bases 3' of the CA-3' were present as
single-stranded
DNA, as was observed for the HIV-1 INprotein.
Specific
cleavageand integrationof15-bp oligonucleotides. Weshowed that the terminal 15bpof the LTRs of HIV-1aresufficientforspecific cleavage andintegrationby IN. These terminal 15bp, however,werepresenton anoligonucleotide of 28
bp (Fig.
2).Therefore,wewantedtodetermine whether anoligonucleotide
ofonly the terminal 15bp of the HIV-1U5 sequence would be sufficient for specific cleavage and
integration by
HIV-1 IN. Cleavage of the15-bp
substrate wascompared with cleavage ofthe28-bp wild-type substrate(oligonucleotide
5/6)
andof the28-bpsubstrate thatcontains mutations at positions 26 to 14 (Fig. 2). We also testedduplex oligonucleotides
that have 15 nucleotides in one strand and 28 in the other strand. The structures of theoligonucleotides
are depicted in Fig. 4A; the results areA
01 2
3
4
5
6
7
8
9
10
lT
~~~~~~~~~-GT
0
0 1
\ ~~~~~~~-CAC;T
0
11
9-12
13 14
B
1/2
.,:i...
~~CGTCAAAAA
3/4 5/6 7/8 9/10 11/12 13/14
-32
.j..:
:-28
-26
C
5/6 5/2 3/4
-32 CAGTI' GTCAAAA
J
-rxlrrr
on November 10, 2019 by guest
http://jvi.asm.org/
D 5/6 5/8
5/10
5/12 5/14
3/4
3/2
5'6
5/2
7/8
7/2
9/109/2
11/12
11/2
13/1413/2
- 28
Vftto4a%a'v-26
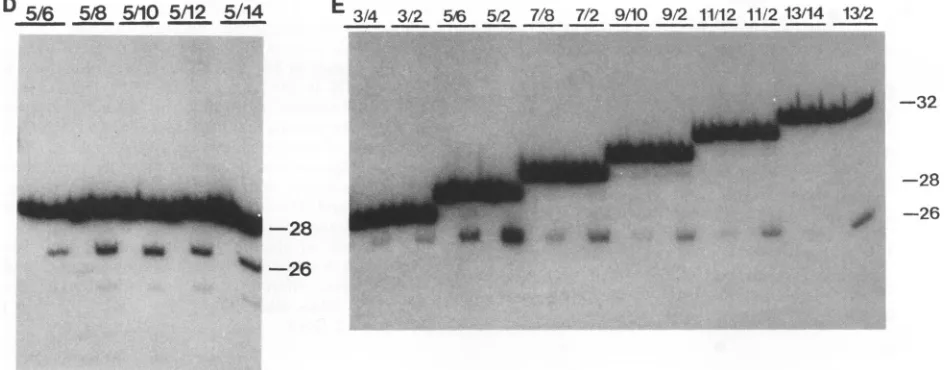
FIG. 3. Cleavage and integrationofmutantHIV-1U5 oligonucleotides that contain deletionsorextensionsatthe 3'sideoftheconserved CA-3'dinucleotide.(A)Structures oftheoligonucleotide substrates. In eachcase,thetoplinerepresentstheHIV-1U5sequence asdepicted
inFig.2A,andthebottom linerepresentsthe complementary strand. Thelinescorrespondtoalengthof 24 nucleotides. Each dot indicates
theradiolabeled phosphategroupatthe 5' end of the viral strand. EachINcleavage site is depictedbyan arrow. (B)Specificcleavageand
integration ofthe substrates shown in panelA.The substrateswereincubated in the absence (leftlane)orpresence(right lane)ofHIV-1 IN.
Theautoradiograph shownin lane 1 (substrate1/2)wasexposedtwiceaslongasthose in the other lanes. Substrate numbers (from panelA)
are indicated above the lanes. Lengths of the oligonucleotide strands are shown on the right. (C) Specific cleavage and integration of
substratesthatcontainsingle-stranded extensions intheoligonucleotidestrand that is cleaved by IN. Substrate numbers (from panel A)are
indicatedabovethe lanes. (D)Specificcleavage and integration of substrates that have single-stranded extensions in the strand that isnot
radiolabeled. Substrate numbersareindicated above the lanes. (E) Specific cleavage and integration of different substrates by HIV-2 IN.
Substratenumbersareindicated abovethe lanes.
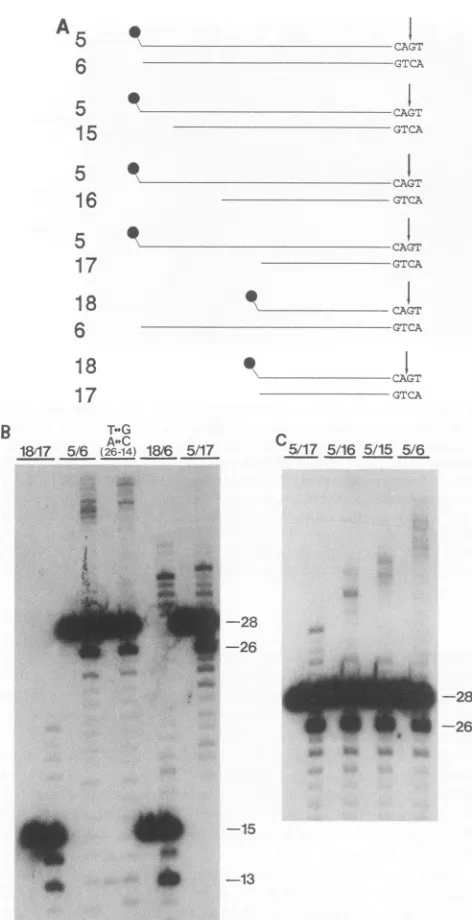
shown in Fig. 4B. The 15-bp substrate could indeed be cleaved by IN, but there were about equal amounts of cleavage product of 14 nucleotides (a nonspecific product) and of 13 nucleotides (the specific product). Integration products could also beobserved when the 15-bp substrate was used. Weconclude that the 15-bp oligonucleotide is a
substrate for specific cleavage and integration but not as
good a substrate as the 28-bp oligonucleotide. Specific cleavagewas more efficientwiththesubstrate in whichthe strand that iscleaved by IN hada length of 15 nucleotides
and thecomplementary strand hadalength of 28 nucleotides (substrate18/6;Fig. 4B).Inaddition,integrationeventswere
more frequent withthis substrate than with the 15-bp sub-strate. Itmightbesuggestedthatmorenucleotides than the terminal 15 bpof the HIV-1 U5 sequence are required for recognition byIN. We have shownabove, however, thatall residues located more than 15 nucleotides away from the viralterminicould be alteredwithout adetectable effecton
specific cutting. Incombination with the observation thata
single-stranded extension beyond position 15 from the ter-minus is sufficient to rescue high-efficiency cleavage, we
suggestthat IN is helped by the presence ofany (nonspe-cific)extensiontoholdontoitsviral recognition site.
Targetspecificity for integration. The largest products of integrationof the15-bpoligonucleotidewere24nucleotides inlength (Fig.4B anddatanotshown). Thisfinding demon-strates that integration can take place at phosphodiester bondsthatarelocated4nucleotidesormoreremoved from
the 5' end of the target DNA (13 nucleotides are obtained from the 3' recessed strand that integrates, andapparently the maximumlengthof thetargetDNAstrand is 11 nucleo-tides). Thelargestproductthatresultedfromintegration of the wild-type substrate had a length of 50 nucleotides (as observed after longer electrophoresis of reaction products
andoverexposureof the gel;datanotshown), whichimplies
that the 26nucleotidesoftherecessed donoroligonucleotide integrated4nucleotidesor moreremoved from the 5' termini
ofa 28-bp targetDNA. When substrate 18/6 was used, the
largestintegration products that could be detected were 38
nucleotidesinlength (Fig. 4B), whichmeansthatinthiscase
theintegration sitewas3nucleotides or moreremoved from
thetermini ofthetargetDNA. Theseexperiments showthat theintegration sitemustbelocatedataninternalpositionin
thetargetDNA andatleastthreenucleotides awayfrom the 5' end of the target DNA.
Aninteresting observationcanbemade when the maximal length oftheintegration product ofsubstrate 5/17 is consid-ered. The radiolabeled strand of the donor DNA that can integrateis 26nucleotideslong (aftersite-specificremoval of twobases from the initial 28-nucleotide strand by IN); the complementary strand is 15 nucleotides long. Ifthe target phosphodiester bondcould beanybond in either
oligonucle-otidestrand that is located 3 or morenucleotidesawayfrom
a 5' end (see above), then the maximum length of the integration productsisexpectedtobe51 nucleotides(26 plus 25). Asshown inFig. 4B, the ladder ofintegration products extends to aposition corresponding to40 nucleotides. This shows that thetarget site for efficient integration cannot be found in the 5' single-stranded region of the 28-nucleotide strand of substrate 5/17. Apparently, integration preferen-tially takes placeinto double-stranded DNA. Close inspec-tion of Fig. 4B reveals faint bands that correspond to reaction products longer than 40 nucleotides. We can
ex-clude that these bandsrepresenttheproductsofraredouble
integration events, since this explanation is not consistent with the observation thatproducts largerthan 40 nucleotides
were not generated when substrate 18/6 was used. An
explanation might be thatintegration can takeplace witha
-32
-28 -26
....
. se
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.73.545.80.265.2]A * --
CAG
5
CXG.6
5
15
5
16
5
17
18
6
18
17 0
-GTCA
GTCA
0 C
I-IT
0
.'
I
CAGT
GTCA
5/175/16 5/15 5/6
-28 -26
*~. W..
"l::.:e o
-15 -13
FIG. 4. Specificcuttingandintegrationof15-bpoligonucleotides and of oligonucleotides that contain stretches ofsingle-stranded
DNAofdifferentlengths. (A)Structures of the substrates. Substrate 5/6 isthe same as inFig. 3. Oligonucleotidestrands15, 16, and17 representtheterminal23, 19, and 15nucleotides ofthe HIV-1 U5 strandthat isnotcleavedbyIN. EachINcleavagesite isindicated byanarrow. (B)Specific cleavageandintegrationof15-and28-bp substrates and of substrates that contain both a 15- and a 28-nucleotide strand.Thesubstrateswereincubated intheabsence(left lane)orpresence(right lane)of HIV-1 IN.Substratenumbers(from panel A)are indicated above the lanes. Thestructureofsubstrate
T<->G,A-QC(26-14)
isshownin Fig. 2A. Lengthsof the oligonucle-otide strands are shown on the right. (C) Specific cutting andintegrationofsubstrateshavingdifferentstretches ofsingle-stranded DNA. Substratenumbersareindicatedabove thelanes.
low
frequency
into single-stranded DNA. However, we cannotruleoutthatsomebasepairing
ofthesingle-strandedDNAoccursunder the reaction conditions thatwereused.
The conclusion that IN
preferentially
usesdouble-stranded DNA as atarget for insertion wasfurther tested.
Oligonucleotides
containing
differentlengths
ofsingle-stranded DNA were studied. The structures of these sub-strates are shownin
Fig.
4A. In all cases, the strandtobe cleavedby
IN is the 28-nucleotide HIV-1 U5wild-type
strand. Inthe various
substrates,
this strand is annealedtocomplementary
strandsthat havealength
of15, 19, 23,
or28nucleotides
(the
complementary wild-type strand).
Thecom-plementary
oligonucleotides
thatare smaller than 28nucle-otides are recessed at their 3' ends relative to the
28-nucleotidestrand. The results of thecleavageand
integration
assay with these substrates are shown in Fig. 4C. The
maximal
length
of theintegration products
increases asexpected
with thelength
of thedouble-strandedregion
of theoligonucleotides.
Therefore,
we conclude thatintegration
preferentially
takesplace
into double-stranded rather thansingle-strandedDNA.
DISCUSSION
The IN
protein
of HIV-1 was expressed in E. coli andpurified
to near homogeneity. The sequence andspacing
requirements
forefficientspecific
cleavageandintegrationof HIV DNAby
IN were studied in a direct physical assay. HIV-1 IN was shown to be capable of specificallycutting
and
integrating 28-bp
oligonucleotidesthat mimic the U3 and U5 termini of HIV-1 as well as HIV-2 DNA. Despite thedifferences in sequence between the U3 and U5 end of HIV-1 or HIV-2, these substrates were cleaved and
inte-grated
withaboutequal efficiencies.Oligonucleotide
substrates that contain mutations relative to the oligonucleotide that represents the terminal 28 bp of the HIV-1 U5 sequence were tested. The results presented here demonstrate that there is only a limited sequencerequirement for site-specific cleavage and integration
by
HIV-1 IN. Only pointmutations in the direct vicinity andat the 5' side of the IN cleavage site could drastically reduce
cleavage
andintegration.
Mutation of the conserved CA-3' dinucleotide tothe sequence TG-3' totallyblockedcleavage
and, as aconsequenceintegration. When the Aatposition 1(Fig.
2A) was changed to a T, low-efficiency cleavage was detected. However, integrationeventscouldnotbedetectedwhenthissubstratewasused. This result could beexplained
by
the fact that the Aatposition 1 of the HIV 3' ends is the nucleotide that is coupled to the target DNA. Theimpor-tance of this A nucleotide for integration has also been observed for MoMLV (3). In this case, the integration of
plasmids, linearized by digestionwith EcoRI, was studied in agenetic assay. It was shown thatintegration was favored when the termini of the plasmids were modified in such a waythat an A nucleotide was present at the 3' ends.
A limited sequence requirement for integration has been describedpreviouslyfor the MoMLV INprotein, whichwas provided either as cytoplasmic extract from MoMLV-in-fected cells(37)or asapartiallypurified protein (3). MoMLV IN was shown to be capable of integrating HIV DNA,
despite the considerable difference in sequence between the termini of MoMLV and HIV DNA (3, 37); 7 of 15 nucleo-tides at the U3 end and 9 of 15 residues at the U5 end are different between HIV-1 and MoMLV. The integration of HIV substrates by MoMLV IN was less efficient than the
integrationof MoMLV substrates (3, 37). We show herethat
despite the limited sequence requirements for integration,
HIV-1 IN is not able to specifically cleave MoMLV DNA. Even when nucleotides 1 to 4 or 8 to 12 of the MoMLV U5 sequence in Fig. 2A were changed to the residues that
B
T-G18'17 5/6 (26-14) 18/6 5/17
J-s
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.62.298.72.532.2]are present at corresponding positions in the HIV-1 U5 sequence [substrates TT(4-3)>AG and GGGGG(12-8)
>AAATC, respectively],noefficientspecific cleavagecould be seen. It should beemphasized, however, that the crite-rion for specific cutting is rather stringent; low levels of
cleavage cannotbe detected inthe assay that we used. The limited sequence requirement for specific cleavage
andintegration ofretroviral DNA termini raises thequestion of how IN can specifically recognize its viral target site. Besides the sequence contextoftheviralends, the spacing between the IN cleavage site and the DNA terminus is important for cleavage and integration (see below). In vivo,
in the cytoplasmofthe infectedcell,the viralDNAis partof
anucleoprotein complex that containsthe INproteinasthe
only detectable protein component (12). It is
possible
that the IN proteins are helped, by their interaction with eachother, to recognize the retroviral DNA ends. Thus, the
interaction between the IN proteins orthe continued pres-ence ofIN molecules in the right position inthe viral core
particle might circumvent the need for
high-specificity
rec-ognition ofthe viral endsby IN.
The distance between the INcleavage site (the phospho-diester bond 3' ofthe CA-3' sequence) and the retroviral DNA terminus is important for specific cleavage by IN.
Efficient specific cleavage occurred when the IN
cleavage
site is spaced two nucleotides away from the DNA end. In thewild-type situation, the HIV-1 INcleavage
site is prob-ably also located two nucleotides away from the DNAterminus. This was inferred fromthe sequencesof
junction
fragments (so-called circlejunctions)
that aregenerated
invivo after circularization of linear double-stranded viral DNA molecules (17, 22, 23, 33, 39). We found that when
only one nucleotide was present atthe 3' side ofthe CA-3'
sequence, the level of
cleavage
was less than with thewild-type substrate. The additionofmore than two nucleo-tides to the 3' side of the CA-3' sequence resulted in a
gradual decrease of
site-specific cleavage.
Thefinding
thatall oligonucleotides with more (or
less)
thantworesidues 3'of the conserved CA-3' were indeed cleaved at the
phos-phodiester bond 3'adjacent
to the CA-3'implies
that thecleavagesite isdetermined
by
sequencesinternaltothe LTRtermini. This view is consistent with the observation that correct3' recessed terminiareformed invivoand in vitro
by
MoMLV mutants that contain additional nucleotides 3' of the CA-3' dinucleotide (8,
26).
It wasdemonstrated,
how-ever, thatcleavage atthe INcleavage
site did not occurinvivo when this site was located 12 bases internal to the terminus oftheMoMLV DNA
(26).
This result is in agree-ment with our finding that HIV-1 IN is not able tospecifi-cally cleave retroviralDNAsubstrates when the IN
cleavage
site is locatedmorethanfive nucleotidesinternaltothe DNA ends.Ourresultsarefurther consistentwith the observation
thatASLV circle
junction
sequences,in which the ASLVINrecognition
sites are present at internalpositions,
are notefficient
substrates forcleavage
and,
as a consequence, for integration (18, 19). Our observations couldprovide
anexplanation
forwhy
integrated
proviruses
arenotsubstrates for IN and thereforecannotbe excised from the host DNA.Site-specific cleavage
by
IN was enhanced when theresidues 3'
adjacent
to the conserved CA-3' dinucleotidewere present as
single-stranded
DNA. A stretch ofsingle-strandedDNA
might
be moreaccessible forspecific
cleav-age by IN than is double-strandedDNA,
because a single-stranded extension mayprovide
less steric hindrance than does a double-stranded extension.Single-stranded
exten-sionsatthe 5' endnearthe IN
cleavage
site didnothave anyeffecton
cutting
andintegration.
The terminal 15
bp
of HIV-1 DNA are sufficient forspecific cleavage
andintegration by
IN.Although
a28-bp
oligonucleotide
thatmimics the endsofHIV-1 DNAwas a better substrate forcutting
andintegration
thanwas a15-bp
substrate,
it was demonstrated that this difference was not the result of additionalspecific
recognition
ofsequences in theregion
that is located more than 15bp
away from the terminus. The additional stretch ofsingle-
ordouble-stranded DNA that was present in the substrates that are
longer
than 15 nucleotidespossibly plays
inrole inhelping
the IN
protein
toholdontoits viral DNA substrate. Similar observations have been made for many restriction endonu-cleases thatrequire
that theirrecognition
sitebeafewbasepairs
removed fromthe ends ofDNA.Itshouldbe
emphasized
that the assayin whichcleavage
and
integration
were testedrepresents
anartificialsystem.
For
example,
therequirement
forMn2+
ratherthanMg2+
as the divalentcation for efficientspecific cleavage
andintegra-tion
(8, 30)
is not consistent with the observations that in vitrointegration
ofendogenous
viralDNA,
which ispresent
in
nucleoprotein complexes,
takesplace
efficiently
withMg2+
asthedivalent cation(1, 10, 15).
The resultsconcern-ing
the substratespecificity
ofpurified
IN are,however,
inagreement with whatwas observed in in vivo
systems.
The limitedrequirement
forspecific
sequences at the retroviralDNA termini that we demonstrated for
purified
HIV-1 IN haspreviously
been shownin vivo for MoMLVIN(26).
Theinability
ofHIV-1INtoefficiently
cleaveits viraltarget
sitewhenthistarget site is locatedataninternal
position
hasalso been demonstrated in the in vivo MoMLVsystem
(26).
Therefore,
it isprobable
that theresults thatweobtaineddo indeed reflect trueproperties
of the INprotein.
Furtheranalysis
oftheproperties
ofthe INprotein
might
therefore contributeconsiderably
totheknowledge
of retroviralinte-gration.
ACKNOWLEDGMENTS
Wethank E.Wientjensand H.vanderGuldenfor
synthesis
oftheoligonucleotides,
and we thank M. Bolk for the construction ofplasmid pRP177.
This researchwas
supported by
grant900-502-119 fromtheDutchOrganizationforScientificResearch,
by
grant88033fromtheDutchMinistry
of Health, andby
a PIONIER grant from the DutchOrganization
for Scientific ResearchtoR.H.A.P. REFERENCES1. Brown, P.O., B.Bowerman,H.E.Varmus,andJ. M.Bishop.
1987. Correct
integration
of retroviral DNA in vitro. Cell 49:347-356.2. Brown,P.O., B.Bowerman,H. E. Varmus,andJ. M.Bishop.
1989. Retroviral
integration:
structure of the initial covalentproduct andits precursor, and a rolefor the viral IN
protein.
Proc. Natl. Acad. Sci. USA 86:2525-2529.3. Bushman,F. D.,and R.
Craigie.
1990.Sequence requirements
for
integration
ofMoloney
murineleukemia virus DNA in vitro. J. Virol.64:5645-5648.4. Bushman, F. D., and R.
Craigie.
1991. Activities of humanimmunodeficiency
virus(HIV)integration protein
in vitro: spe-cificcleavage
andintegration
ofHIV DNA. Proc. Natl. Acad. Sci. USA88:1339-1343.5. Bushman, F. D.,T.
Fujiwara,
and R.Craigie.
1990. Retroviral DNAintegration
directedby
HIVintegration
protein
invitro. Science 249:1555-1558.6. Colicelli,J.,and S. P.Goff. 1985. Mutantsand
pseudorevertants
of
Moloney
murine leukemia virus with alterations at theintegration
site. Cell42:573-580.on November 10, 2019 by guest
http://jvi.asm.org/
7. Colicelli, J., and S. P. Goff.1988. Sequence and spacing
require-mentsofaretrovirus integration site. J. Mol. Biol. 199:47-59.
8. Craigie, R., T. Fujiwara, and F. Bushman. 1990. The IN protein of Moloney murine leukemia virus processes the viral DNA ends and accomplishes their integration in vitro. Cell 62:829-837.
9. Donehower, L. A., and H. E. Varmus. 1984. A mutantmurine leukemia virus withasingle missense codon inpol is defective inafunction affectingintegration. Proc. Natl. Acad. Sci. USA
81:6461-6465.
10. Ellison, V., H. Abrams, T. Roe,J. Lifson, and P. Brown.1990. Human immunodeficiency virus integration in a cell-free
sys-tem. J. Virol. 64:2711-2715.
11. Farnet, C. M., and W. A. Haseltine. 1990. Integration of human immunodeficiency virustype1DNA in vitro. Proc. Natl. Acad. Sci. USA87:4164 4168.
12. Farnet, C. M., and W. A. Haseltine.1991. Determination of viral proteins present in the human immunodeficiency virus type 1 preintegration complex. J. Virol. 65:1910-1915.
13. Fisher, A. G., E. Colialti, L. Ratner, R. C. Gallo, and F. Wong-Staal. 1985. A molecular cloneofHTLV-IIIwith biolog-ical activity. Science316:262-265.
14. Fujiwara, T., and R. Craigie. 1989. Integration of mini-retroviral DNA:acell-free reaction for biochemical analysis of retroviral integration. Proc.Natl. Acad. Sci. USA 86:3065-3069. 15. Fujiwara, T.,and K. Mizuuchi. 1988. Retroviral DNA
integra-tion:structure ofanintegration intermediate. Cell 54:497-504.
16. Guyader, M., M. Emerman, P. Sonigo, F. Clavel, L. Montagnier, and M. Alizon. 1987. Genome organization and transactivation of the humanimmunodeficiency virustype2. Nature(London) 326:662-669.
17. Hong, T., K. Drlica, A. Pinter, and E. Murphy. 1991. Circular DNA of human immunodeficiency virus: analysis of circle junction nucleotide sequences. J. Virol. 65:551-555.
18. Katz, R. A., G. Merkel, J. Kulkosky, J. Leis, and A. M. Skalka. 1990. The avian retroviral IN protein is both necessary and sufficient forintegrative recombination in vitro.Cell63:87-95. 19. Katzman, M., R. A. Katz, A. M. Skalka, and J. Leis. 1989. The
avian retroviral integration protein cleaves the terminal se-quencesoflinear viral DNAatthe in vivo sites ofintegration. J. Virol.63:5319-5327.
20. Laemmli, U. K.1970. Cleavage of structuralproteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
21. Muesing, M. A., D. H.Smith, C. D. Cabradilla, C. V. Benton, L.A.Lasky, and D. J.Capon. 1985. Nucleic acidstructureand expression of the human AIDS/lymphadenopathy retrovirus. Nature(London) 313:450-458.
22. Pauza, C. D. 1990. Two basesaredeleted from the termini of
HIV-1 linear DNAduring integrative recombination. Virology
179:886-889.
23. Pullen, K. A., and J.J. Champoux. 1990. Plus-strandorigin for
humanimmunodeficiency virustype 1:implications for integra-tion. J. Virol.64:6274-6277.
24. Ratner, L., W. Haseltine, R.Patarca, K.J. Livak, B.Starcich, S. F.Josephs, E. R. Doran, J. A. Rafalski, E. A. Whitehorn,K.
Baumeister, L.Ivanoff, S. R.Petteway, Jr., M. L. Pearson, J. A.
Lautenberger, T. S. Papas, J. Ghrayeb, N. T. Chang, R. C.
Gallo, and F.Wong-Staal. 1985.Complete nucleotide sequence oftheAIDSvirus, HTLV-III.Nature(London) 313:277-284. 25. Rosenberg, A. H., B. N. Lade, D. Chui, S. Lin, J. J. Dunn, and
F. W. Studier. 1987. Vectorsfor selectiveexpression of cloned DNAsbyT7RNApolymerase. Gene 56:125-135.
26. Roth, M. J., P. L.Schwartzberg, and S. P. Goff.1989. Structure of the termini of DNA intermediates in the integration of retroviralDNA:dependenceonINfunction and terminalDNA sequence.Cell 58:47-54.
27. Sambrook, J., E. F.Fritsch,and T.Maniatis. 1989. Molecular cloning:alaboratory manual. ColdSpring Harbor Laboratory, ColdSpring Harbor,N.Y.
28. Schwartzberg, P., J.Colicelli,andS.P.Goff.1984.Construction andanalysisofdeletion mutationsin the polgene ofMoloney murine leukemia virus: a new viralfunction required for pro-ductive infection. Cell 37:1043-1052.
29. Seed, B. 1983.Purification of genomicsequencesfrom bacterio-phage librariesby recombination andselection in vivo. Nucleic AcidsRes. 11:2427-2447.
30. Sherman, P. A., andJ.A.Fyfe.1990. Humanimmunodeficiency virusintegration protein expressed in Escherichia coli possesses selective DNA cleaving activity. Proc. Nati. Acad. Sci. USA 87:5119-5123.
31. Shinnick, T. M., R. A. Lerner, and J. G. Sutcliffe. 1981. Nucleotide sequenceofMoloney murine leukaemia virus. Na-ture(London)293:543-548.
32. Shoemaker,C., S.Goff,E.Gilboa,M.Paskind,S. W.Mitra,and D. Baltimore. 1980. Structure of a cloned circular Moloney murine leukemia virus DNA molecule containing an inverted segment: implications for retrovirus integration. Proc. Natl. Acad. Sci. USA77:3932-3936.
33. Smith, J. S., S. Kim,and M. J. Roth. 1990. Analysis of long terminal repeat circle junctions of human immunodeficiency virustype 1. J. Virol. 64:6286-6290.
34. Stevenson, M., T. L. Stanwick, M. P. Dempsey, and C. A. Lamonica. 1990. HIV-1replication is controlledatthelevelofT cellactivation and proviral integration. EMBO J. 9:1551-1560. 34a.vanGent, D., Y. Elgersma, C. Vink, and R. H. A. Plasterk.
Submitted forpublication.
35. Varmus, H. E. 1983. Retroviruses, p. 411-503. In J. Shapiro (ed.), Mobile genetic elements. Academic Press, Inc., New York.
36. Vink, C., M. Groenink, Y. Elgersma, R. A. M. Fouchier, M. Tersmette, and R. H. A. Plasterk.1990.Analysis of thejunctions betweenhuman immunodeficiency virus type 1 proviral DNA andhuman DNA. J.Virol. 64:5626-5627.
37. Vink, C., D. C. van Gent, and R. H. A. Plasterk. 1990. Integration of human immunodeficiency virus types 1 and 2 DNA in vitro by cytoplasmic extracts of Moloney murine leukemia virus-infectedmouse NIH3T3cells.J.Virol. 64:5219-5222.
38. Wain-Hobson,S., P.Sonigo,0. Danos, S. Cole, and M. Alizon. 1985. Nucleotide sequence of the AIDS virus, LAV. Cell 40:9-17.
39. Whitcomb, J.M., R. Kumar, and S. H. Hughes. 1990.Sequence of thecirclejunction of human immunodeficiency virustype 1: implications forreversetranscriptionandintegration.J. Virol. 64:4903-4906.