0022-538X/92/095248-08$02.00/0
Copyright© 1992, American Societyfor MicrobiologY
Inhibition of Structural
Changes
in the Simian Virus 40
Core
Origin
of
Replication by Mutation of Essential
Origin
Sequences
JAMES A. BOROWIEC
Departmentof Biochemistry, New York University MedicalCenter,
New
York,
New York10016Received 15May 1992/Accepted 16 June 1992
Mutation of the simian virus 40 (SV40) originofreplication (ori)has revealed the presence of three critical domains needed for DNAreplication.Theoutertwo
domains,
the AT tract andearly
palindrome
element (EP), colocalize with DNAregionsthat becomestructurally
altered in the presence of theSV40largetumor antigen (Tantigen)and ATP. Mutations within each domainwereexamined for their effectonthe distortion ofori DNA by Tantigen,asassayed bythesensitivity
of DNA toKMnO4oxidation. We have found that mutations in the AT tractthat inhibitSV40DNAreplicationalso inhibit the distortion of the ATtract.Similarly,
mutations in the EP inhibited the generation of structural changes in this element by Tantigen.
Although
AT-tract mutations or mutationson the late side of ori affected structuralchanges only
in the ATtract,
certain EP mutations ormutations ontheearly
side ofoni also inhibited AT-tract distortion. Mutation of the flanking regions did notsignificantly
affect either theaffinity
of Tantigen
foroni ortherate ofbinding
tooni. We conclude from these results that the primaryfunction of theflanking
ori domains is toundergo
structural changes required during the initiation ofSV40 DNAreplication.
Moreover,
our resultssuggest
that theefficiencyofreplicationinitiation is
significantly
affectedby
thedegree
towhich theflanking
elementsundergo a structuraltransition.The initiation of DNAreplication from internal sites on chromosomespresentsa
challenging
obstacle forthecellularreplication
machinery.
Sufficientlengths
of double-strandedDNAmustbe rendered
single
strandedto serve astemplatesfor DNApolymerasestosynthesize thenascentstrands. The mannerby which
replication
factorsinteract withthe DNAhelixto convert it tosingle-stranded DNA remains poorly understood.The useofrelativelysimple modelsystemsfor eukaryoticDNA replication has made this problem tracta-ble.Examination ofsimian virus40(SV40) indicatesthat the
presynthetic changes in DNA structure take place in an elaborate series of steps(2).
An essential first step in SV40 DNA replication is the
binding of the SV40 largetumor antigen (T antigen) to the viral origin of replication (on). At elevated temperatures
(37°C),
stablebindingof Tantigento coresequenceswithin onrequiresATPandresults intheformationof amultimericcomplex containingup to12 monomersof protein (4, 14, 24).
This ATP-dependent complex is notable for its bifurcated structurewith each halfcontaining a hexamer of T antigen
(10, 24, 28). In the presence of human single-stranded
DNA-bindingprotein (human SSB, RP-A, RF-A [19, 35, 36]) or certain heterologous SSB proteins, the intrinsic DNA
helicase activity of T antigen can catalyze the large-scale
unwinding of DNA molecules fromon (8, 22, 37). Mecha-nistic parallels to these events were observed in prokaryotic model systems such as
oriC
ofEscherichia coli andoriA
ofbacteriophage X(1, 17).
The nucleation of DNA melting in the SV40 system appears to take place within the ATP-dependent complex.
Chemical probing of the on DNA within this complex reveals two regions of altered DNA structure. In one arm of
an
imperfect
inverted repeat, termed the early palindromeelement(EP)orinvertedrepeat,approximately 8 bp of DNA
are denatured. On the opposite flank of the on situated toward the late gene side of SV40, nearly 20 bp of an adenine-thymine (AT)tractundergoaconformational distor-tion whileremaining primarily double stranded(3, 5, 26,28).
These observed structuralchangesappear tolead,atleast in part, to topological untwisting of the on-containing DNA
(11, 29).
That each of these conformationalchanges in onstructure isacritical stepduring the initiation of SV40 DNA
replica-tion is suggestedby high-resolution genetic mapping of on. Tegtmeyer and colleagues have described three discrete elements within the65-bpcoreon that are essentialforSV40 DNA replication in vivo or in vitro (7, 12, 13, 15). Twoof thesefunctional elementsclosely overlap the two conforma-tionally flexible regions inthe EP and AT-tract regions. The thirdcentral region, containing four GAGGCelements in a perfect mirror repeat, acts as a recognition element for T-antigen binding. Point mutations in any of these three elements can reduce DNAreplication activity by 2 orders of magnitude in vivo (12, 13, 15).
Therelationship between the induction of conformational changes within on and theinitiation of replication was tested byexamining the effect of replication-defective mutations on thedistortion of the two flanking regions of on by T antigen.
Single-base-pair
mutations in critical late side elementssignificantly inhibited the generation of structural changes within the AT tract while having only minor effects on structural changes in the opposite flanking region. Con-versely, while all early side mutants had significantly lower levels ofstructural changes in the EP, two of these mutants were also defective in the generation of conformational changes in the AT tract. These results provide direct evi-dence that theimportance of the two flanking regions lies in
5248
on November 9, 2019 by guest
http://jvi.asm.org/
during the initiation ofSV40DNAreplication. 5230
5243M
1l l1i 31 lategenes -_
MATERIALSAND METHODS
Preparation of protein and DNA reagents. T antigen was purified from Sf9 insect cells infected with recombinant baculovirus vEV55SVT (25) or Ac941SVT. Ac941SVT, a kind gift of Monika Lusky, Cornell University Medical
College, New York, N.Y., wasprepared by initially cloning a T-antigen cDNA into the transfer vector pVL941 (23).
Ac941SVT, although producing T antigen identical in all
respects to thatisolated from
vEV55SVT,
yields three to five times more T antigen per liter of infected Sf9 cells. Prepara-tion of infected-cell lysates and immunoaffinity purificationofTantigenweredone by the method of Borowiec et al. (3). The DNA constructs used in these experiments were a kind gift of Peter Tegtmeyer, State University of New York,
Stony Brook. The nomenclature used by Tegtmeyer and
colleaguesin reference 7 ismaintained in this paper, with the
additional change that each pointmutation is termed bs (for
base). For example, the on mutated at position 5212 is
termed bs5212. Each DNA was prepared from DHSa cells
(21) by alkali lysisandCsClbanding (30).
KMnO4 footprinting of DNA. Standard reactionmixtures
(30
RI)
contained 40 mM creatinephosphate (di-Trissalt[pH 7.8]), 7 mM MgCl2, 0.5 mM dithiothreitol, 0.5 ,ug of theappropriateplasmidon DNA, 4 mM ATP, 25 ,g of creatine
phosphokinase per ml, andvarious quantities of T antigen.
Reactionswereincubated at 37°C for 60 min before
modifi-cationwith KMnO4. Although Tantigen can form a stable
complexwithon DNAaftershortertimes(e.g.,seeFig. 7),
the induction ofstructural changes that are detectable by
KMnO4 requires incubation times in excess of 30 min (3). KMnO4was added from a freshly prepared stock solution (200 mM) to give a final concentration of 20mM, and the reaction was incubated for an additional 4 min. Primer
extension and gel electrophoresis conditions were
essen-tiallyasdescribedby Borowiec and Hurwitz (5).
The level of KMnO4oxidationin eachregionwas
quanti-tatedby densitometric scanningofautoradiographswith an LKB Ultrascan XL laser densitometer. Autoradiographs
with various exposure times were used to ensure that the bandintensitywaswithinthe linear rangeoffilmsensitivity.
The extentof KMnO4 oxidationof theearlypalindromewas determinedby
quantitating
theintensity
of theregion
corre-sponding totop-strand
thymine
residues 5217 and 5218.Although thethymine atbs5214
produces
aKMnO4
signal,
thissignal constitutesonly
approximately
15%of thesignal
derivedfrom the
major
reactive nucleotidesatpositions
5217and 5218 andcanbe
easily distinguished.
For the ATtract, bandscorresponding
tothymine
residues16, 20, 29,
and30 of the top strandwere used. The overallintensity
of eachregionwas
separately
normalizedby
comparison
with fourcontrol
thymine
residues located outside theSV40coreon. The mutant originswere alwaysprobed inparallel
with thewild-type
(wt) on,
which servedas an internal control. The maximumnormalizedvaluefor theearly
palindrome
andthe AT tract of wt ori for each titration was set at 1.0. Thefraction of maximum oxidation for the mutant
origins
was then calculatedforeach datumpoint.
Gel
mobility
shiftanalysis
of Tantigen-oi complexes.
The32P-end-labeledori DNAwas
prepared by
firstdigesting
theappropriatemutant
plasmid
DNAwith EcoRI andHindIIIto release theoni-containing
DNAfragment
(approximately
100 bp inlength).
After treatment with calf intestinal alkaline> (18,41)
L4bslS
34)
--(8,31) bs521 ' j(6,27) bs26
bs5214 Ins5243 (1,22)
(8,16) (<1, 5)
Inv Pent1 InvPent 4 (<1, 1) (<1, 1)
FIG. 1. Mutations in theSV40 coreonthatwere tested for their effect on the distortion ofon DNA by T antigen. The general location of eachmutant is shown. bs5211, bs5212, and bs5214 are located in the EP; Inv Pent 1,Ins5243, bs4, and Inv Pent 4 are located in the central GAGGCelement (GAGGC); andbsl8,bs22, and bs26 are located in the AT tract. The numbers below each mutation indicate the replication efficiency relative to wt on (in percentages). The first number indicates the activity in vivo, while the second number indicates the activity in vitro, obtained by using crude cytoplasmic extracts of HeLa cells (data from reference 7).
phosphatase (USBiochemical), the DNA was end labeled in thepresence of
[_y-32P]ATP
andpolynucleotide kinase, and the labeled on DNA was purified after electrophoresis through an 8% polyacrylamide gel (acrylamide/bisacryl-amide ratio, 29:1). Each gel shiftassay used approximately 50 fmol ofDNA(at 1 x106
to2 x106
cpm/pmol). For gelmobility shift assays, standard reaction mixtures (20 ,ul)
contained 40 mMcreatinephosphate (di-Tris salt[pH7.8]),
7 mM MgCl2, 0.5 mM dithiothreitol, 4 mM ATP, 25
F±g
of creatinephosphokinaseperml,onDNA, and variousquan-tities of T antigen. Standard binding reactions were incu-bated at 37°C for 60 min and then cross-linked by the
additionofglutaraldehydeto0.1%andincubationfor 15 min at 37°C. Kinetic measurements used shorter binding times
(see Fig. 8), followed by the identical cross-linking proce-dure. The reaction mixtures were then loaded onto a 4% polyacrylamide gel (acrylamide/bisacrylamide ratio, 29:1) andelectrophoresed in thepresence of 25 mMTris, 190 mM
glycine,and 1 mM EDTA(pH 8.5). Gelslicescorresponding
to the shiftedT-antigen-DNAcomplexwere removed, and theradioactivitywasdeterminedbyscintillation counting.
RESULTS
Thebindingof Tantigen tothe SV40
on,
inthepresence of ATP, results in the induction ofsignificant
structural changes at two flanking regions within on. Each of these deformable DNA elements resides inregions
foundprevi-ouslytobecritical foronfunction. Wethereforewishedto understandthe
relationship
between thestructural deforma-tion of these elementsby
Tantigen
and theimportance
of these elements for DNAreplication.
We used a series of mutations located within on thatwereinitially
constructedby Tegtmeyer andcolleagues
(7,
12, 13,15).
Themutations werechanges of one to three basepairs
orsingle-base-pair
insertions andwere located withinone of the three criticalregionsof
on,
i.e.,theflanking
EPorAT-tractregions
orthe central GAGGC element(Fig. 1).
Each mutant on waspresentas a coreon
lacking
theflanking
T-antigen-binding
site Iand SV40
transcriptional
control elements.The induction ofstructural
changes
in each mutant on,contained in
plasmid DNA,
wasprobed by using KMnO4
oxidation. WhileKMnO4
reactspoorly
with DNA in the-.
I
on November 9, 2019 by guest
http://jvi.asm.org/
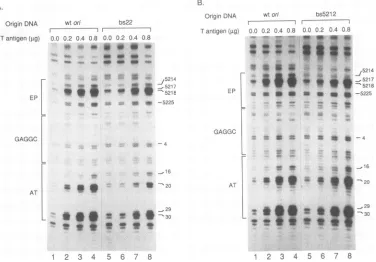
[image:2.612.321.560.81.198.2]A.
OriginDNA wtori bs22
I I
Tantigen(gLg) 0.0 0.2 0.4 0.8 0.0
.. 4.
.... j,:44
' I
0.2 0.4 0.8
. 'S
40 g
4 _m
B.
OriginDNA wtori bs5212
Tantigenhig) 0.0 0.2 0.4 0.8 0.0 0.2 0.4 0.8 P.
* ; ^ ^
J5214
jji _ 521
V * -521e
w w# -5225
EP
GAGGC
-4
a a
- _16
- 9 -20 AT
_29
* * * * -30
_ t_ * I e v
. S It
. _!
a. u l
.q
-m...
** 5214
* _5217
* .-5218 -5225
-_
.2!l _ 16
U,.~~~~~~~~~~~~~~~~.
-zoL.*X.t
ft-3g~_2
12 3 4 5 6 7 8 1 2 3 45 6 7 8
FIG. 2. Effect ofT-antigenlevelsonthe induction of structuralchangesin thewild-typeonandinonDNA moleculesmutated in theAT
tractorEP. Arepresentative autoradiograph showingthechangesinDNAreactivitytoKMnO4as afunctionof Tantigenis shown. Towt
on(AandB;lanes 1to4),bs22(A;lanes5to8),orbs5212(B;lanes5to8)wasadded 0.0,ug (lanes1 and5),0.2jig(lanes2 and6),0.4,ug
(lanes3 and7),or0.8pug (lanes4 and8)of Tantigen.After incubation for60min,the DNAwasmodified with 20 mMKMnO4.Theoxidation reactionwasthenquenchedand the sites of oxidationweredeterminedasdescribed in Materialsand Methods. Varioussequencepositions
areindicatedontherightsideof eachpanel.The locations of theEP,centralGAGGC,and ATtractareshownonthe leftside of eachpanel.
B-form, DNA that is meltedor apparently bent sharplyor
significantly untwisted (of altered helical pitch) becomes hyperreactive to KMnO4 oxidation, predominantly at thy-mine residues(5, 6).These residuescanbepreciselylocated by first extending a 32P-labeled DNA primer across the region of interestwith the Klenow fragment of DNA
poly-merase I. Because the oxidized nucleotides can stall the polymerase, analysis of the extensionproducts by denatur-ing gel electrophoresis and autoradiography indicates the sites and relative extentofKMnO4oxidation. With asingle
exception (bs5214; seeMaterials andMethods), noneof the
mutations affected thymines producing the KMnO4 oxida-tionsignal.
The inductionofstructuralchangesinthewtonDNAwas
first compared with an on DNA mutated in the AT-tract
element at nucleotide 22 (bs22). The bs22 mutation was
foundpreviouslytoinhibit SV40DNAreplicationto
approx-imately 40% ofwt levels when tested in vitro with crude extracts,and under 20% ofwtlevels whenexamined in vivo (7, 13, 15). These DNA molecules were incubated with
increasing levels of T antigen, and the DNA was then
modified with KMnO4. A representative autoradiograph of theprimer extension productscomparesthe KMnO4
oxida-tionpatternsforwtonand bs22 (Fig.2A). As theamountof Tantigen addedtowtonwasincreasedfrom 0.0to0.8 ,ug, thelevel of KMnO4oxidationalso increasedat tworegions withinon(Fig. 2A, lanes1to4).These regions,asindicated
onthe left side ofthe figure,correspond to the EP andAT tractofon.Titrationof similaramountsof T antigentobs22 induced identical nucleotides withinthis mutant tobecome hypersensitiveto KMnO4 oxidation (Fig. 2A, lanes5 to8).
Visual comparison of the amount of KMnO4 oxidation in thesetwoon DNAmolecules indicatedthatmodificationof the ATtract of bs22was significantlydecreased relative to wt on. Although oxidation of the EP element of bs22 appeared slightly less than that of wt on, quantitative
analysisof the data foundnosignificantdifference(see Fig. 3B). Thus,in this qualitative comparison, asinglebase pair mutation of the AT tract inhibited thegeneration of struc-turalchanges in the ATtractbyTantigen.
A similar experiment was performed comparing the
KMnO4oxidation patternsofwtonandan onDNAmutated
in the EP at nucleotide 5212 (bs5212; Fig. 2B). This point mutation inhibits SV40 replication tolevelsthat are31% in
vitro and8%in vivoofwtlevels (7, 12, 15). The EP mutation significantly decreased the extent ofKMnO4 oxidation in thisregionwhenintermediate levelsofTantigenwereused
(0.2and0.4 pg;comparelanes 2and 3 with lanes 6 and 7 in
Fig. 2B). Noobservableeffects of thebs5212 mutationwere
notedonmodificationoftheATtract.Thus, mutations in the EPorATtractthatareinhibitorytoSV40 DNA replication also inhibitdistortion ofthe mutated region by T antigen.
We performed a comprehensive analysis of the effectof on mutationon thedistortion ofon structureby T antigen. Eachmutation testedwas foundpreviouslyto significantly reduce SV40 replication both in vivo and in vitro (Fig. 1). Each ori mutant was incubated with various amounts of T antigen, in the presence of ATP, and the DNA was then
probedwithKMnO4. Theextentof KMnO4 oxidationfor the AT-tract and EPregionsineachon were separately
quanti-tatedbydensitometricscanning of theautoradiographs. The levels ofmodification in these twoon elementswere
sepa-EP
GAGGC
AT
on November 9, 2019 by guest
http://jvi.asm.org/
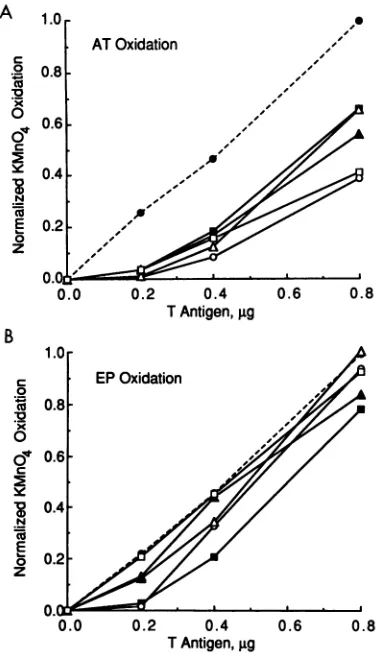
[image:3.612.131.507.80.342.2]1.0
ATOxidation
o0.8
Cu~~~~~~~~~~~~~~~~~~~~~~~4
~0 0.6
-C
0.4-E 0.2 -0 z
0.0
0.0 0.2 0.4 0.6 0.8
[image:4.612.90.278.76.403.2]TAntigen,gg
FIG. 4. Effect of T-antigen levelsonKMnO4 oxidation in theEP
and AT tract ofon DNA molecules mutated on the early side
(nucleotides 5211to5243). Quantitation ofoxidation in the EP (A) and ATtract(B) is shown. ThemutantonDNAs testedwerewton
(0),bs5211(D),bs5212(A), bs5214 (0),Ins5243 (A), andInv Pent 1(a). Quantitation of oxidationwasasdescribed in the legendto
Fig.3.
mutationreduced KMnO4 oxidation of the ATtracttolevels 18 to 40% of that of wton. High levels of T antigen (0.8 ,ug)
did not overcome the effect of the mutation, as KMnO4
modification within the ATtractremained 39 to66% ofwt levels. Incontrast,these identicalmutations had less signif-icant effectson the generationofstructural changes within
the EP(Fig. 3B).Inthepresenceof 0.4,ugof Tantigen,four of the five mutations reducedKMnO4modificationof the EP to levels 72 to 99% ofwt values. At 0.8 ,ugof T antigen, oxidation within the EP ofanyof thesemutantsdid notfall below 79% of the wt level. Therefore, late-side mutations that are deleterious to SV40 DNAreplication significantly inhibited AT-tract distortionbyTantigenwhilehaving only minor effectsondistortion of the EP.
Fiveearly-sideonmutantsweretested for theirabilityto
undergo T-antigen-mediated changes in DNA structure. Three of these mutations were located in the EP (bs5211,
bs5212, andbs5214).The fourth mutation was aninversion
of themostleftward(early) GAGGCsequence(InvPent 1), and the lastwas asingle-base insertion betweenthe middle two GAGGC pentamers (Ins5243). The Ins5243 mutation, because of itspositionin the center ofon,wouldpresumably separatethe two hexameric lobes ofT antigen by0.34 nm
and rotate one lobe with respect to the other by
approxi-C
~0
x
o
'a
.R
0
0.4 TAntigen, igg
0.8
0.0 0.2 0.4 0.6 0.8
TAntigen, igg
FIG. 3. Effect of T-antigen levelsonKMnO4 oxidation in the EP and AT tract of on DNA molecules mutated on the late side (nucleotides 1to31).Quantitation of oxidation in theATtract(A)
and EP(B) is shown. ThemutantonDNAs testedwerewton(0),
bsl8(-),bs22(A), bs26 (0),bs4 (A), and InvPent 4(El).Various amounts ofT antigen were incubated with the wt or mutant on
plasmid, and the sites of oxidationwerevisualizedasdescribed in thelegendtoFig. 2. The levels of KMnO4 oxidation for the EP and
ATtractwereindividuallydetermined for eachonbydensitometric
analysis of the autoradiograph. The data for each element were normalized with respectto themaximum level ofoxidation
deter-mined forwton(at0.8 jigofTantigen) by comparison with control
bands located outsideon.
ratelynormalizedby comparisonwith controlbands located outside the on region. The normalized levels of KMnO4
oxidation in the AT-tract and EPregionswere thenplotted
as a function of added T antigen for eachmutant on. The datawere grouped accordingtowhether the mutationwas
locatedon the late side (nucleotides 1 to26; Fig. 3) or the earlyside (nucleotides 5211 to 5243; Fig. 4)ofon. Control experiments examining on mutations that did not signifi-cantly affect SV40 DNA replication were also performed. These mutations (atnucleotides5222, 5228, and12)did not significantly affect the distortion of on structure (data not
shown).
Five late-side mutations were examined, and each had
severeeffectsonthe distortion of theAT tractbyTantigen
(Fig. 3A). Twowere located inthe central GAGGCregion (bs4andInvPent4),and the otherthreewerelocated within
theAT tract(bsl8, bs22,andbs26).TheInvPent 4 mutation
is an inversion of the most rightward (late) GAGGC
se-quence. At moderate levels of T antigen (0.4 ,ug), each
00 x
0 0
c
0
z
B
C
Y.
x
o
0
z
on November 9, 2019 by guest
http://jvi.asm.org/
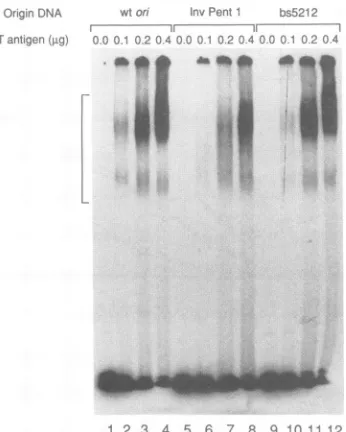
[image:4.612.353.541.81.411.2]OrgginrDNA vtIorl !nvPeflnt bs52 1 2
- ----r---
--Tantigen (LP, 0001 02025.4 r0 220. .2
1 2 3 4 5 6 7 8 9
11
Ii 12FIG. 5. Effect of on mutation on the formation of the ATP-dependentcomplexbyTantigen.Variousamountsof Tantigen,as
indicated, were incubated with 32P-end-labeled DNA fragments containing eitherwton (lanes1to4), InvPent1 (lanes5 to8),or
bs5212(lanes 9 to 12), in thepresence of ATP. After 60min, the T-antigen-DNA complexes were cross-linked by the addition of glutaraldehyde. Thecomplexeswere then separated from freeon DNAbyelectrophoresis through anondenaturing gel, and thegel was dried and autoradiographed. TheT-antigen-DNA complexis indicatedbyabracketonthe left sideof thegel.
mately 34°. Each mutation
significantly
inhibited the induc-tion of structuralchanges
within the EP(Fig.
4A).
Themutations locatedintheEP
region
reduced themodificationofthis region 28 to 65% compared with thewton at
high
levelsof Tantigen(0.8
,ug).
The Inv Pent 1 mutation had the greatesteffecton thegenerationofstructuralchanges, with theextentofKMnO4 modification reducedtoless than2%of wtlevels.Early-side mutationshad differentialeffects onthe induc-tion of structural changes inthe ATtract (Fig. 4B). At the
highestlevel ofT antigen (0.8 ,ug),AT-tract distortion was significantly inhibitedfortwomutants,bs5214and Inv Pent 1,to 28 and39%, respectively, ofwtlevels. The Inv Pent 1
mutationisparticularly informativesincethecorresponding mutationonthe lateside,InvPent4, hadinsignificanteffects on KMnO4 modification ofthe EP. Two other early-side
mutations
(bs5211
andbs5212)
had only minor effects onKMnO4 modificationof the ATtract.The last mutant tested,
Ins5243, had moderately reduced levels of AT-tract oxida-tion compared with the wton. Weconclude that although
late-sidemutations have little effect on structural changes in theEP, certain early-side mutations can affect the generation ofstructural changes in both the EP and AT tract.
Theinhibitory effect ofonmutation onondistortion could be either an indirect result of the mutation decreasing
T-antigen bindingor adirect effect of the mutation
decreas-ing the intrinsic ability of the flanking
on
elements toundergo a conformational change. To distinguish between these twopossibilities,weused a gel shift assay to examine the binding of T antigen to a
32P-labeled
DNA fragmentcontaining either a wt or mutant
on.
Increasing levels of Tantigen
were added to the on DNA molecules, and the4000
0..
E0
~o
C
0
z a
0.0 0.1 0.2
TAntigen,
9g
0.3 0.4
FIG. 6. Quantitationof the effect ofon mutationonthe forma-tion oftheATP-dependentcomplex by T antigen.Increasinglevels of Tantigenwereincubated with 32P-end-labeled DNA fragments containingwt or a mutanton. The T-antigen-on complexeswere thencross-linkedby usingglutaraldehyde and separated fromfree on DNAbynondenaturing gel electrophoresis asshown inFig. 5. The portions of the gel corresponding to the T-antigen-ori com-plexes,afterdetectionbyautoradiography,werethenremoved, and theradioactivitypresent wasdeterminedby scintillation counting. Theon DNAconstructstestedwere wton(0), bs5212(U),bs5214 (A),InvPent1(0), Ins5243(O),bs4(A),InvPent 4
(+),
bsl8(El), bs22(x),andbs26(*).T-antigen-DNA complexes were then cross-linked with glu-taraldehyde. TheT-antigen-DNA complexeswereseparated by nondenaturing gel electrophoresis and detected by auto-radiography.
Thebinding of Tantigen to wt on, Inv Pent 1, andbs5212
was examined first(Fig.5). The addition ofasmall amount of T antigen (0.1 jig) to wt on led to the production of two distinct complexes (lane 2). Comparison with the work of Parsons et al. (28) indicates that the faster and slower complexesareoni molecules boundbyahexamerordouble hexamer of Tantigen,respectively. Increasingthe levelof T antigen to0.4 jig increased the amountof DNAboundbya double hexamer of Tantigen (lane 4). Qualitativeinspection of theautoradiographshowsthat the Inv Pent 1mutation,as
expected,decreased theabilityof Tantigentobindtotheon (lanes 5 to 8), whereas the bs5212 mutation had little observable effect (lanes 9 to 12).
Similar conditionswereusedtoexamine thebindingof T antigentothemutantonDNAs. Thebindingof Tantigento each DNAwasquantifiedbydeterminingthe sum of radio-activity present in both bands of the complex (Fig. 6). Certain mutations located in the central GAGGC region significantly reduced thebindingof T antigen. At 0.4jigof T
antigen, the InvPent 1,bs4,and Inv Pent 4 mutations each decreased the amountofT-antigen-ori complex to
approxi-mately two-thirds of the wt level. On the other
hand,
T-antigen binding to DNA molecules containing mutations in the ATtract or EP(bs5212,bs5214,bsl8, bs22, and bs26) or to the Ins5243 mutant was not significantly different frombindingto wton, and any small differences were within the normalvariation of this assay. Thus, mutations in the EP and ATtractdidnotinhibit the induction of the conformational
changesin theseregions by preventing T-antigen binding to on. Ourresults suggest instead that these mutations directly affect the abilityof the AT-tract and EP regions to undergo structural changes mediated by T antigen.
on November 9, 2019 by guest
http://jvi.asm.org/
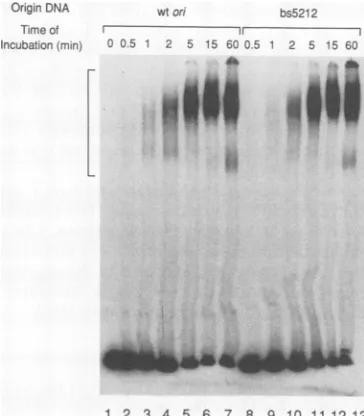
[image:5.612.339.545.75.245.2] [image:5.612.96.269.77.293.2]I
Incubation(min) 0 0.5
---I
1 2 5 15 600.5 1 2 5 15 60
.1611
!1|111
E 2000CL 0
m
z 1000
0 5 10 15
1 2 3 4 5 6 7 8 9 10 11 1213 FIG. 7. Effect ofon mutation on the rate of formation of the ATP-dependent complex. T antigen (0.2 ,ug) was incubated with 32P-end-labeledDNAfragments containing either wton(lanes 1 to 7)orbs5212(lanes 8 to 13). Aftervariousintervals (as indicated), the T-antigen-DNA complexes were cross-linked by the addition of glutaraldehyde. The complexes were then separated from freeon DNAby electrophoresis through a nondenaturing gel, and the gel was dried andautoradiographed. The T-antigen-DNA complex is indicatedby a bracket on the left side ofthe gel.
on mutation may in turn affect the rate of T-antigen
binding toon. To examine this possibility, we incubated T
antigen(0.2 ,ug)forvarioustimes with32P-end-labeled DNA
fragments containingamutant orwton. TheT-antigen-on complexes were cross-linked with glutaraldehyde, and the
relative amount ofcomplexformed was examined by a gel
shiftassay. AtimecourseofT-antigen bindingtothe wton
andtheEP mutantbs5212 indicated thatthe rates ofbinding tothewton (lanes 1 to7) and bs5212 (lanes 8 to 13)were verysimilar(Fig. 7).The amount of Tantigen boundtothese andother mutantoriginsateachtime pointwasquantitated
by determining the amount of radioactivity in gel slices
corresponding
to both bands of the T-antigen-DNA com-plex.Tantigen bound allmutanton DNAmoleculesat ratessimilartothatforbindingto wton DNA
(Fig. 8).
Althoughtheoverall amountof
T-antigen
bindingto DNAfragments
containing mutations intheGAGGCelement of on
(bs4,
Inv Pent1, and Inv Pent4)
was reduced, the rateofT-antigen
binding
tothesefragments
wassimilartothat forfragments
containingwton.
DISCUSSION
The
ATP-dependent
binding
of Tantigen
to the SV40 originofreplicationresults in the structural distortion oftwo flankingregions
of on. These tworegions
overlap
two onelements
required
fororigin
activity.
Wehave examined the effect ofreplication-defective
onmutationsonthe distortion ofonDNAbyTantigen.
Mutation of critical nucleotides ineither the EP or the AT tract decreased the
ability
of Tantigen to induce structural
changes
within thatregion.
Theseflankingmutations haveno
significant
effectoneither the amount of Tantigen
bound or the rate ofT-antigen
bindingtoon. We thereforeconcludethat the
importance
ofTime of Binding (min)
FIG. 8. Quantitation of the effectofonmutationonthe kinetics
offormation of the ATP-dependentT-antigen-on complex. T
anti-gen(0.2 ,ug)wasincubated withwtonor amutantonfor various
times. After cross-linking of the T-antigen-DNA complex, the
complex was separated from free on DNA by electrophoresis
throughanondenaturinggel (as shown in Fig. 7). The portionsof the
gel correspondingtothe T-antigen-ori complexes,after detection by
autoradiography,wereremoved, and the radioactivitypresentwas
determined by scintillationcounting. TheonDNAconstructstested
werewton(0), bs5212(U), bs5214 (A), Inv Pent1(0),Ins5243(l),
bs4(A),InvPent4(+), bsl8(E0),bs22 (x), and bs26 (*).
the two flanking regions lies in their ability to undergo structural changes required during the initiation of SV40 DNAreplication.
Of the 10mutations tested, 6werelocatedin the flanking
sequence elements. We found similar qualitative effects of
themutationson the generation of structural changes in the
mutated region andon SV40 DNA replication. The bs5214
and bs26 mutations had the greatest effect on SV40 DNA
replication in vitro andwerealso the mostinhibitory tothe generation of structural changes in the EP and AT tract, respectively. These results suggest that the efficiency of replication initiation issignificantly affected by the degreeto
which the flanking elements undergoastructuraltransition.
Mutation of the central GAGGC element inhibited the ability ofT antigen to bind toon. Thisresult is consistent
with various studies demonstrating close contacts of T antigen with 5'-GAGGC-3' sequences in both the core on
and theadjacent site1(5, 16, 32) andagreeswith the results
of Parsons et al. (26). Somewhat surprisingly, mutation of
the core sequences inhibited only the level of T antigen
bindingtotheonbut didnotsignificantlyinhibit theoverall rate of T-antigen-on complex formation. In other words, normalization of each set ofbinding data to the maximum amount of DNA bound indicated that the binding of the mutantonDNAwasneverless than 68% of thewton level ateach time point tested. Although the GAGGC elements
are presumably requiredto properly positionT antigen on on, the rate-limiting step of binding, under our reaction conditions, does not appear to be the recognition of these
sequences. Wehavenotperformedextensive kinetic
analy-sis of thebindingof Tantigen towt on, but it wouldseem
plausiblethata "slow" stepoccursafterthe initial interac-tionofTantigenwith the DNA. Dean et al. (9)haveshown
thatpreformedhexamers ofTantigencannotproperlybind to on and that therefore T-antigen hexamers are probably
formedontheDNAbysequentialaddition ofmonomers(see
60
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.86.268.75.283.2] [image:6.612.333.541.76.249.2]also reference 28). Apossible interpretation ofour data is that theformation of the hexameric Tantigenonon afterthe initial GAGGC recognition is the rate-limiting step in the formationof theATP-dependent complex. The useofother assays(e.g., dimethyl sulfatefootprinting)mayallow
detec-tion of intermediate complexes during the initial stages of binding.
Why does mutation of the flanking regions inhibit their
distortion by T antigen? The minimal effect of such
muta-tions on T-antigen binding demonstrates that the reduced
structural distortion is notaconsequenceofpoorT-antigen binding to the origin. However, the workbyParsons et al.
indicates that Tantigencanweakly recognizethe EPregion
in the absence of the central GAGGC and AT-tract regions (26). Thus,onepossibilityis that Tantigenstill bindswithwt efficiency to the mutant templates because of the intact
GAGGC elements, yet forms weaker contacts with the mutatedflanks. Tantigenwould therefore be unable toform
contacts sufficiently stable to fully distort the flanking
se-quence.Analternativemodel is that eachflankingelementis predisposed to undergo a structural transition and that
mutation lessens their ability to become distorted. In this
model, Tantigen can interactwith the flanking regionsbut
these contactsareinefficientlyconverted intoanobservable structural change. These two models are not mutually
ex-clusive, andour data cannotyet distinguishbetween them.
Our data bearuponthe interaction ofT-antigenmolecules
bound toon in thepresenceof ATP. Mutation of eitherthe EP or the AT tract affected primarily the generation of
structural changes within the mutant element whilehaving lesssignificanteffectsontheopposite flanking region. Thus,
these datasupport the hypothesis that the ATP-dependent complex is composed of two somewhat independent do-mains,each containingahexamer ofTantigen (3, 28). It is importanttonote that certainearly side mutations(bs5214, Inv Pent 1) significantly inhibited conformational changes
within the AT tract. Similarly, the Ins5243 mutation, a
single-base-pairinsertionbetween theearly and latehalves
ofon, reduced thelevel of structuralchangesinboth flanks
ofon. Our datatherefore suggestthat the twohexamers do
havesignificantinteractions andthat the hexamerboundto
theearlyhalf ofonappearstobedominant to the Tantigen
bound to the late half. This hypothesis has also been independently suggested by Parsons and Tegtmeyer (27),
whoexamined thespacing requirementsbetweenconserved
elements inon.
The events occurring prior to the synthesis of nascent
DNA duringthe initiation of SV40 DNA replication have beenthe focus ofsignificant effort (for areview, see
refer-ence 2). These studies suggest that T antigen, binding as
monomericunits toon, forms ahexamer over each half of on (24, 28). In the early half of on, the EP acts as the
nucleation site forDNAmeltingwithintheon (5, 26).Both
human SSB protein (RF-A, RP-A) and some noncognate SSB proteins can allow the unwinding of on-containing DNAmolecules by Tantigen (8, 22, 37). Because specific interactionsbetween T antigen and certain nonhuman SSB proteins (e.g.,E. coli SSB)seemunlikely,the major role for
SSB is presumably to bind to the single-stranded DNA
within the EP and thus further destabilize the
double-strandedstructure ofon. Wenote, however, thatT antigen canspecificallyinteract withhumanSSB (18)and that these
contacts mayfacilitatethisevent. TheATtract, relativeto
the EP, does not appear to be greatly melted within the ATP-dependent complex (5, 26). The structural changes
within theATtractwouldbothfurtherdecreasethe stability
of the double helixand allow release of theT-antigen DNA helicaseonbinding of SSB to the EP. The T antigen, inan as yetunidentified step,would release from on astwo hexam-ers,orpossiblyas adoublehexamer(33), with each T-anti-gen hexamerbinding to opposite strands so that T antigen can move outwardfrom on in the 3'-*5' direction (20, 31,
34). Each hexamer then individually unwinds the DNA primarily by usingcontactswiththe sugar-phosphate back-bone(31).
Furtherworkisnecessarytounderstandhow Tantigen is able to convertfromasequence-specific DNA-binding pro-tein to a sequence-independent DNA helicase. Our current understanding indicates that theprocess isan intricateand subtle interactionbetweenTantigenand theon, leading to the complete denaturation of the origin structure, which allows the subsequentsynthesisof nascentDNA.
ACKNOWLEDGMENTS
We thank PeterTegtmeyer for his kindgiftof the SV40 origin constructs,MonikaLuskyfor thegenerousgiftofAc941SVT,and WarrenJelinek foruseof the LKB Ultrascan XL laser
densitome-ter. We also thank Frank Dean and Jerry Hurwitz for critical
commentsonthemanuscriptand DhrubaSenGupta,Len Blackwell, and Tom Gillette forhelpfuldiscussions. Weappreciatethe techni-cal assistanceprovided byRichard Tomko and Max Kallet.
Researchwassupported byNIHgrantAI29963,the Pew Biomed-ical ScholarsProgram (T88-00457-063), andKaplan Cancer Center Developmental Funding and Cancer Center Support core grant (fromNCIP30CA16087).
REFERENCES
1. Baker, T. A., K. Sekimizu, B. E. Funnell, andA. Kornberg. 1986. Extensive unwinding of the plasmid template during staged enzymaticinitiation of DNAreplicationfrom theorigin of the Escherichia coli chromosome. Cell 45:53-64.
2. Borowiec, J. A.,F. B.Dean,P. A.Bullock,andJ.Hurwitz.1990. Bindingandunwinding-howTantigenengagesthe SV40origin ofDNAreplication.Cell 60:181-184.
3. Borowiec, J. A.,F. B.Dean,andJ.Hurwitz.1991. Differential induction ofstructuralchanges in the simian virus40originof replication byTantigen. J.Virol.65:1228-1235.
4. Borowiec, J. A., and J. Hurwitz. 1988. ATP stimulates the binding ofsimian virus 40 (SV40) large tumor antigen to the SV40originofreplication. Proc. Natl.Acad. Sci. USA 85:64-68.
5. Borowiec, J. A., andJ. Hurwitz. 1988. Localized melting and structural changes in the SV40 origin of DNA replication inducedbyTantigen. EMBOJ.7:3149-3158.
6. Borowiec,J. A., L. Zhang, S. Sasse-Dwight, and J. D.Gralla. 1987. DNA supercoilingpromotes formation ofa bent repres-sionloopinlacDNA.J. Mol. Biol. 196:101-111.
7. Dean,F., J. Borowiec,Y.Ishimi,S. Deb, P. Tegtmeyer, and J. Hurwitz. 1987. Simian virus 40 large tumor antigen requires threecore replication origin domainsfor DNA unwinding and replicationinvitro.Proc. Natl. Acad. Sci. USA84:8267-8271. 8. Dean,F.,P.Bullock,Y.Murakami, C. R. Wobbe, L. Weissbach, andJ.Hurwitz. 1987.Simian virus40(SV40)DNA replication: SV40largeTantigenunwindsDNAcontaining the SV40 origin ofreplication.Proc. Natl. Acad. Sci. USA 84:16-20.
9. Dean,F.B., J.A.Borowiec, T. Eki, and J. Hurwitz. 1992. The simian virus40 Tantigen doublehexamer assembles around the DNAatthereplication origin.J. Biol. Chem. 267:14129-14137. 10. Dean, F. B., M. Dodson, H. Echols, and J. Hurwitz. 1987. ATP-dependentformationof a specialized nucleoprotein
struc-turebysimian virus40(SV40) largetumor antigen at the SV40 replicationorigin.Proc.NatI.Acad. Sci. USA84:8981-8985. 11. Dean, F. B., and J. Hurwitz. 1991. Simian virus 40 large T
antigenuntwistsDNAattheorigin of DNA replication. J. Biol. Chem. 266:5062-5071.
12. Deb, S.,A. L.DeLucia,C.-P. Baur, A. Koff, and P. Tegtmeyer.
on November 9, 2019 by guest
http://jvi.asm.org/
13. Deb, S., A. L. DeLucia, A.Koff, S. Tsui, and P. Tegtmeyer. 1986. The adenine-thymine domain of the simian virus 40 core origin directs DNAbending and coordinately regulates DNA replica-tion. Mol. Cell. Biol. 6:4578-4584.
14. Deb, S., and P. Tegtmeyer. 1987. ATP enhances the binding of simian virus 40 large T antigen to the origin of replication. J. Virol. 61:3649-3654.
15. Deb, S., S. Tsui, A. Koff, A. DeLucia, R. Parsons, and P. Tegtmeyer. 1987. The T-antigen-binding domain of the simian virus 40 core origin of replication. J. Virol. 61:2143-2149. 16. DeLucia, A. L., B. A.Lewton, R. Tjian, and P. Tegtmeyer. 1983.
Topography of simian virus 40 A protein-DNA complexes: arrangement of pentanucleotide interaction sites at the origin of replication. J. Virol. 46:143-150.
17. Dodson, M., H. Echols, S. Wickner, C. Alfano, K. Mensa-Wilmot, B. Gomes, J. LeBowitz, J. D. Roberts, and R. Mc-Macken.1986. Specialized nucleoprotein structures at the origin ofreplication of bacteriophage X: localized unwinding of duplex DNAby a six-protein reaction. Proc. Natl. Acad. Sci. USA 83:7638-7642.
18. Dornreiter, I., L.F.Erdile,I.U.Gilbert,D. vonWinkler,T.J. Kelly, and E.Fanning. 1992. Interaction ofDNA polymerase alpha-primase with cellularreplication protein A andSV40 T antigen. EMBOJ. 11:769-776.
19. Fairman,M.P.,and B.Stillman.1988.Cellular factorsrequired formultiple stages of SV40DNAreplicationin vitro.EMBO J. 7:1211-1218.
20. Goetz, G. S.,F. B.Dean, J.Hurwitz,and S. W. Matson. 1988. Theunwinding ofduplexregionsinDNAbythesimian virus 40 large tumor antigen-associatedDNAhelicaseactivity. J. Biol. Chem. 263:383-392.
21. Hanahan,D.1983. Studiesonthetransformation of Escherichia coli withplasmids. J. Mol. Biol. 166:557-580.
22. Kenny, M. K., S.-H. Lee, and J. Hurwitz. 1989. Multiple functions of human single-stranded-DNA binding protein in simian virus 40DNAreplication:single-strandstabilization and stimulation of DNApolymerasesax and b. Proc. Natl. Acad. Sci. USA 86:9757-9761.
23. Luckow,V.A.,and M. D. Summers.1989.Highlevelexpression of nonfusedforeign genes withAutographacalifornica nuclear polyhedrosisvirusexpressionvectors. Virology170:31-39. 24. Mastrangelo,I.A.,P. V. C.Hough, J.S.Wall,M.Dodson,F. B.
Dean,andJ.Hurwitz.1989.ATP-dependent assemblyof double hexamersofSV40Tantigenatthe viraloriginof DNA replica-tion.Nature(London)338:658-662.
insect cells. J. Virol. 62:3109-3119.
26. Parsons, R., M. E. Anderson, and P. Tegtmeyer. 1990. Three domains in the simian virus 40 core origin orchestrate the binding, melting, and DNA helicase activities of T antigen. J. Virol. 64:509-518.
27. Parsons, R., and P. Tegtmeyer. 1992. Spacing is crucial for coordinationof domain functions within the simian virus 40 core origin of replication. J. Virol. 66:1933-1942.
28. Parsons, R. E., J. E. Stenger, S. Ray, R. Welker, M. E. Anderson, and P. Tegtmeyer. 1991. Cooperative assembly of simian virus 40 T-antigen hexamers on functional halves of the replication origin. J. Virol. 65:2798-2806.
29. Roberts, J. 1989. Simian virus 40 (SV40) large tumor antigen causes stepwise changes in SV40 origin structure during initia-tion ofDNAreplication. Proc. Natl.Acad. Sci. USA 86:3939-3943.
30. Sambrook, J.,E. F.Fritsch,and T.Maniatis. 1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
31. SenGupta, D., and J.A.Borowiec. 1992.Strand-specific recog-nition ofa syntheticDNAreplication fork by the SV40 large tumorantigen. Science 256:1656-1661.
32. Tjian,R.1979. Protein-DNA interactionsattheoriginofsimian virus 40DNA replication. Cold Spring Harbor Symp. Quant. Biol. 43:655-661.
33. Wessel, R., J. Schweizer,and H. Stahl. 1992. Simian virus 40 T-antigen DNA helicase is a hexamerwhich forms a binary complexduring bidirectionalunwinding from the viraloriginof DNAreplication. J. Virol. 66:804-815.
34. Wiekowski, M., M. W. Schwarz, and H. Stahl. 1988. Simian virus 40large T antigenDNAhelicase. J. Biol. Chem. 263:436-442.
35. Wobbe, C. R., L.Weissbach, J.A. Borowiec, F. B. Dean,Y. Murakami, P. Bullock, and J. Hurwitz. 1987. Replication of simian virus 40 origin-containing DNA in vitro with purified proteins. Proc.Natl.Acad. Sci. USA 84:1834-1838.
36. Wold,M.S.,andT.Kelly.1988. Purification and characteriza-tion ofreplication protein A, acellularprotein requiredfor in vitroreplicationofsimian virus 40DNA.Proc. Natl. Acad. Sci. USA85:2523-2527.
37. Wold,M.S.,J. J.IA,and T.J.Kelly.1987. Initiationof simian virus 40 DNA replication in vitro: large-tumor-antigen- and origin-dependentunwindingofthetemplate.Proc.Natl. Acad. Sci. USA 84:3643-3647.