0022-538X/89/083345-08$02.00/0
Copyright X31989, AmericanSocietyforMicrobiology
In
Vitro Processing of
Dengue
Virus Structural Proteins:
Cleavage of
the Pre-Membrane
Protein
LEWISMARKOFF
Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases, Bethesda, Maryland 20892
Received 15 February 1989/Accepted 19April 1989
Processing of dengue virusstructuralproteinswasassessed in vitro. RNAtranscripts forcell-freetranslation wereprepared fromclonedDNA (dengue virustype4, strain814669genome) encodingcapsid, pre-membrane
(prM), and the first 23 amino acids of envelope (E). Processing of a 33-kilodalton precursor polypeptide encodedbywild-typeRNAtranscriptsoccurredonlyin thepresenceofaddedmicrosomalmembranes. Under theseconditions, cleavageatthecapsid-prM and prM-E sitesandglycosylation ofprM occurredinassociation with translocation. Amino acid sequence analysis confirmed that translation initiated at the predicted N
terminus ofthecapsid and that capsid-prMcleavage occurred atthe predicted site forthe action of signal
peptidase following a candidate signal sequence (hydrophobic residues 100 to 113) in the dengue virus precursor. Mutations were introduced into the dengue virus DNA template by site-directed mutagenesis, altering nucleotide sequences encoding the capsid and the candidate signal for prM. The phenotypes ofthe mutantswerededuced by analysis oftheproducts of cell-free translationofthe respectiveRNA transcripts. The resultingobservationsconfirmedthatcleavageatthe capsid-prM and prM-Esites iseffected entirely by signal
peptidaseand that the candidate signalis requiredfor translocation.
The four serotypes of dengue virus are members of the
flavivirus family, which now includes more than 70 different viruses. Flavivirus particles are composed of three structural
proteins: agenome-associatedcapsid protein, a
membrane-associated (M) protein that is derived during virus
matura-tion by internal cleavage from a glycosylated precursor protein (prM), and a membrane-anchored hemagglutinating
envelope glycoprotein(E). E is themajor antigenic
determi-nantof freevirus responsible forserotype specificity(18-20,
31,32).
FlavivirusgenomicRNAs are10,500 to 11,000nucleotides
long and include a single open reading frame encoding a
precursorpolyproteinapproximately 3,400aminoacidslong (5-7, 10, 14-16, 18, 22, 26, 33). Structural proteins are
derivedbycleavage from the N terminus of the precursor in thefollowingorder: capsid,prM, and E (2-4, 18,22, 24, 28).
Since some viral proteins are glycosylated and virus
parti-cles are assembled in the endoplasmic reticulum (11, 19),
processing of the flavivirus precursor polyprotein must
in-clude not only cleavage but also translocation ofcleavage
products. An association betweencleavage ofcapsidand E
proteins and cotranslational translocation has been
sug-gested bythe resultsofanin vitrostudy(25). Inaddition,the
dengue virus capsid, prM, E, and NS1 were apparently normally cleaved and glycosylated when expressed in a
vaccinia virus recombinant containing a 4-kilobase (kb) fragmentof cloned dengue virus type 4 DNA encoding the
capsid, prM, E, NS1, and NS2a (34). Therefore,
nonstruc-tural proteins (NS2b through NS5 [14]) not encoded in the vaccinia virus recombinant are not required to process the
structuralproteins and NS1.
As expected, the locations ofpotential signal and
trans-membrane segments in the flavivirus precursor are
con-served. Within N-terminal sequences encoding structural
proteins,the precursor containsan uninterrupted 14-amino-acid hydrophobic segment(residues 100 to 113 in the
pre-cursorsequence).This sequenceprecedestheNterminus of prM determined by amino acid sequencing of labeled pro-teins isolated from virus-infected cells (2, 4) and is the
candidate signal sequence fortranslocation ofprM(18, 19, 33). Thecandidatesignal is preceded by a conserved pair or triplet ofbasic amino acids (Arg-Lys-Arg; residues 97 to 99 in the dengue virus type 4 sequence [33]). It has been
suggested that these conserved residues might constitute a proteolyticsite(18, 33). Inaddition, thepredicted C termini
of both prMand E arehydrophobic segments(residues246
to 279 and 735 to 773, respectively) asymmetrically inter-rupted in each case by a single Arg. These are candidate transmembrane segments for prM and E and signals for translocation of therespectivedownstreamglycoproteins, E and NS1 (14, 18, 22, 33).
To establish the association between cleavage of struc-turalproteinsfrom the precursor andtranslocation,
process-ing of a truncated dengue virus precursor protein was
studied in vitro. RNA transcripts were prepared from a 911-base-pair fragment of denguevirus type4DNA, includ-ingthecomplete capsid and prM nucleotide sequences and sequencesencoding thefirst23 amino acids of E. Cleavage ofthe encoded precursoratthecapsid-prMand prM-E sites
was dependent upon the presence of microsomes during cell-free translation and appeared to mimic the in vivo
events.Todeterminethemechanism of cleavage, mutations
of nucleotide sequencesencodingthe capsidand the candi-dateprMsignalwere introducedintodenguevirus DNAby site-directedmutagenesis.The resultsof cell-free translation of mutant RNA transcripts are also described. An intact
signalat residues 100to 113 wasrequiredfor translocation and cleavage ofboth prM and E. Mutations of the
capsid
sequence had no effect on processing, and cleavage ap-pearedto beentirely mediatedby the signal peptidase.
MATERIALSANDMETHODS
Preparation of recombinant pTZ/Sal. A 4,040-base-pair
(4-kb)fragment of cloned dengue virus type4 strain 814669 DNA bounded by BglII restriction endonuclease
cleavage
sitesatpositions88and 4128wassubcloned into theplasmid vector pTZ18U (Pharmacia, Inc.,
Piscataway,
N.J.) at theBamHI site within the pTZ18U polylinker in plus-sense
3345
on November 10, 2019 by guest
http://jvi.asm.org/
Bglllfragment, dengueDNA
r1'
NS1Sal I
IN
SalI .
Salfragment, dengue DNA
"I.
pTZ18U NI- -C jprM \ E
f Hind III linearpTZ/SalDNA
polylinker
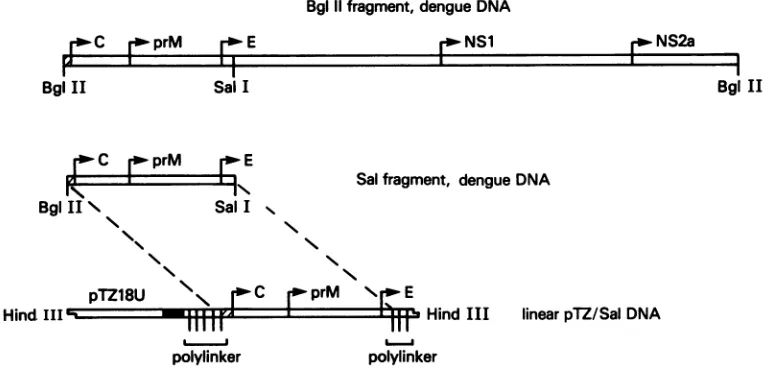
FIG. 1. Derivation of Sal DNA in the pTZ/Salconstruct. A4-kbfragmentof cloneddenguevirus(type 4,strain814669)DNApreviously expressed in a vaccinia virus recombinant (34) was cloned into the plasmid vector PTZ18U at the BamHI site. pTZ18U-dengue virus
recombinant DNAwasdigested withSall, excising 3'-terminalsequencesfrom thedenguevirus DNA. The resultantpTZ/Salrecombinant
DNAlinearized by HindIII digestion, usedasthetemplate for synthesis of RNA transcripts,isdepicted. X, Noncodingsequencesindengue virus DNA; T7 RNA polymerase promoterinpTZ18U.
relationshiptothe T7RNApolymerase promoter sequence
contained in thevector.3'-Terminal nucleotide sequencesin
the dengue virus DNA were excised by digestion of the
recombinant vectorDNA with the restrictionendonuclease
Sall,whichcleaves thedenguevirus DNAatnucleotide 999 andpTZ18UDNAatauniqueSallsite in thepolylinker3'to dengue virus DNAsequences. The Sall-digestedDNA was religated to construct pTZ18U-dengue virus recombinant DNAwhichincluded only the5' 911nucleotides (88to999)
of the4-kbdenguevirus DNAfragment. These 911
nucleo-tidesdefine the Sal fragment of dengue virusDNA.pTZ18U recombinant DNA containing the Sal fragment of dengue virus DNA isdesignated pTZ/Sal (Fig. 1).
Preparation of pTZ/SalssDNA and site-directed mutagene-sis of dengue virus DNA sequences. The vector pTZ18U
contains the fl origin of replication in opposite
transcrip-tional orientationto the T7 RNApolymerase promoterand totheampicillinresistancegene.Toproduce pTZ/Sal single-stranded DNA (ssDNA), we followed a standard protocol. Toproducemutations in dengue virusssDNA,weemployed the site-directed method of Zollerand Smith (35). Klenow
fragment was obtained from Promega Biotec, Madison, Wis., andT4 DNAligasewas obtained from New England BioLabs, Inc., Beverly,Mass.Oligonucleotide primerswere preparedon anApplied Biosystems380A DNASynthesizer.
The presence of a desired mutation was confirmed by primer-directed DNA sequencingofthepertinent region in
dengue virus DNA (21).
Synthesis of RNA transcripts from recombinant vector DNA. pTZ/Sal DNA and clonedmutantrecombinant DNAs were linearizedby digestion with the restriction endonucle-ase HindlIl (Fig. 1). Synthesis of RNA on 2 to 5 ,ug of linearized DNA was carried out by a standard procedure
using20to 40U of T7RNApolymerase (Promega Biotec).
Cell-freetranslation of RNA transcripts and polyacrylamide
gel analysis ofproducts. Cell-free translationwasconducted in a 25- or 50-,ul final volume using a supplemented rabbit
reticulocyte lysate pretreated with micrococcal nuclease
(Promega Biotec). A typical 25-pAl reaction mixture
con-tained17.5,ul of rabbit reticulocyte lysate, 0.5 ,ul ofa1 mM amino acid mixture minus methionine, 0.5 p.l of RNase
inhibitor(RNasin; Promega Biotec), 2.5 ,ul of
[35S]methio-nine(15 mCi/ml, >1,000Ci/mmol;Amersham Corp.,
Arling-tonHeights, Ill.), and 0.2to0.5 ,ug of RNA inwater.Protein synthesiswasconducted in thepresence orabsence of added caninepancreaticmicrosomes (1.0to1.5
RI
replacingwaterin a 25-pu reaction mixture; Promega Biotec). To prepare
proteins for sequencing, 1 mM amino acids minus
methio-nine and leucinewas addedtothe reactionmixture inplace of amino acids minus methionine, and the reaction was
carried out in the presence of 25 ,uCi of [3H]leucine (1
mCi/ml, 150 Ci/mmol; Amersham) and [35S]methionine. Syn-thesized proteinswere analyzed bysodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). Gels
con-tained 0.2% SDS and 15 to 17% acrylamide. The 17% gels wereelectrophoresed at140constantvoltsfor 16 h atroom
temperature, dried, and exposed to XAR-2 film (Eastman Kodak Co., Rochester, N.Y.).
Protease and endo F digestion of proteins. To digest
pro-teins with thermolysin (Boehringer Mannheim Biochemi-cals, Indianapolis,Ind.),5[lofacell-free synthetic reaction
mixture was added to 5 1ld of TC buffer (10 mm Tris hydrochloride [pH 7.5], 10 mMCaCl2), 1 ,ulofthermolysin (1-mglml solution in TC buffer) wasadded, and the reaction mixture was incubated for 60 min at 4°C. Thermolysin activitywasterminatedby the additionofa10-foldexcessof EDTA.
Endoglycosidase F (endo-,B-N-acetylglucosaminidase F [endo F])wasobtainedfromBoehringerMannheim. Twoto five microliters ofa cell-free synthetic reaction mixture or >5,000cpmofaneluted[35S]methionine-labeled proteinwas
incubatedin 25 ofendoFbuffer(Boehringer Mannheim) for 1to3 hat37°C in thepresenceof0.1to0.2 U ofenzyme.
Immuneprecipitation ofproteins. Antibody to peptide 30 (a gift of R.Houghten),a15-amino-acidpeptiderepresenting thepredicted N terminus of the dengue virus type 4, strain
814669 Eprotein(MRCVGVGNRDFVEGV; residues 280to 294 [33]), was prepared in rabbits after conjugation to
keyholelimpet hemocyanin. This antiserum hadan antipep-tide titerof1:1,280 by enzyme-linkedimmunosorbentassay. Proteinssynthesizedinvitrowere incubated for 16 hat 4°C
in thepresenceofantipeptide antibodyatdilutions of 1:10to BglII
Ix
BglII N N1
r.
NS2aBgl
IIHind III'-L-igI
11111 polylinker
---Tlll.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.119.509.79.263.2]1:50. The antigen-antibody complexes were then collected on staphylococcal protein A-Sepharose CL4B beads
(Phar-macia). Thebeads were washedin RIPA buffer (0.1 M Tris [pH 7.5], 0.15 M NaCl, 1% sodium deoxycholate, 1% Triton X-100,0.1% SDS) and boiled in Laemmli sample buffer (0.1
MTris [pH 6.8], 10%glycerol, 1% SDS, 0.2 M 2-mercapto-ethanol,0.001% bromphenol blue) prior to SDS-PAGE.
Amino-terminal amino acid sequencing of proteins. Pro-teins doubly labeled ([3H]leucine and [35S]methionine) as
describedabove were separated by SDS-PAGE on a gel that had been preelectrophoresed in the presence of running bufferplus 1 mMmercaptoacetic acid. A2%agarose
stack-ing gel was used instead of acrylamide, and 0.1 mM mercap-toacetic acid was added to the running buffer. Proteinswere eluted from polyacrylamide and sequenced in a Beckman
Model 890MProtein Sequencer.
RESULTS
Cell-free translation of wild-type RNA transcripts. Sal DNA in the pTZ/Sal construct includes the 13 3'-terminal nucleotides of the dengue virus 5' noncoding region and sequences encoding the capsid (amino acids 1 to 113), prM (amino acids 114 to 279), and the first 23 amino acids of E
(amino acids 280 to 302) in the dengue virus serotype 4 precursorpolyprotein (33). RNA transcripts prepared from
HindlIl-linearized pTZ/Sal DNA were expected to include 21 nucleotides of the pTZ polylinker 5' to dengue
virus-specificsequences as wellas 5'-terminal GGG appended to
all transcripts by T7 RNA polymerase (8). In addition, transcripts included polylinker sequences 3' to the dengue
virus insert which encode a common C terminus
(Gln-Ala-Cys-Lys) in cell-free translation products (Fig. 1). The products of cell-free translation of wild-type (wt) RNA transcripts are shown in Fig. 2A.
[35S]methionine-labeled proteins were directly applied to the gel for SDS-PAGE under reducing conditions. In the absence of added microsomal membranes (Fig. 2A, lane 0), a single major
productwasevident, with a molecular size of approximately 33kilodaltons(kDa). This is in agreement with the predicted
size ofthe uncleavedtranslationproductof full-lengthpTZ/ Sal RNA (306 amino acids; estimated molecular mass,
33,660 daltons). Minor lower- and higher-molecular-mass species were also detected. Higher-molecular-mass species are likely to be the product of RNA transcribed from
contaminating uncleaved circular pTZ/Sal DNA.
Lower-molecular-mass proteins could have resulted from internal initiation of translation or from internal cleavage of the
33-kDa polypeptide. Evidence favored internal initiation;
separate experiments failed to demonstrate a
precursor-product relationship between the 33-kDa and lower-molec-ular-mass proteins, suggesting that cleavage of the
full-length polypeptide did not occur in the absence of
microsomes (datanot shown).
When RNAtranscriptswere translatedin the presenceof canine pancreatic microsomal membranes, seven proteins were detected. Four high-molecular-mass products (A, B, and CinFig.2A, lanes 0.5to2., and C' inFig. 3A, left lane) ranged insize from 32 to 38 kDa. Protein C appeared to be
identical in size to the unglycosylated 33-kDa full-length product. Three low-molecular-mass proteins varied in size
fromabout 24to28kDa (a,b,andcinFig. 2A, lanes 0.5, 1., and 2.). Increasing portions of membranes added to the
reaction mixture were associated with both enhanced pro-duction ofproteinsa, b,andc relativetothe precursor and
diminished total synthesis.
A. B.
piMembranes
M
o75i1.
230
-30
Kd-+ :Mm M - + - +:Pr
A B
i...'h-.C
6
1.s" NW a
c c
21Kd-12
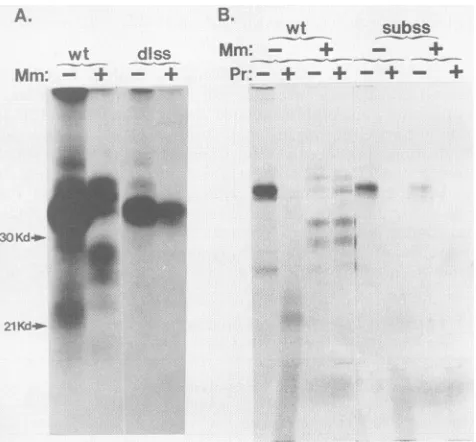
Kd-.-FIG. 2. (A) Cell-free translation ofSalRNA inthe absence(lane 0)orpresenceofincreasing portions (in microliters) of microsomal membranes (lanes 0.5 to 2.). One-half microgram of RNA tran-scribed in vitrowasaddedto17.5 ,ulof micrococcal nuclease-treated rabbit reticulocyte lysate plus 0.5 p.l ofRNasin, 0.5 p.l of 1 mM amino acids minus methionine, 2.5 ,u1 of [35S]methionine, and microsomes asindicated, to afinal volumeof25 ,ul. The cell-free translation reaction mixture was incubated for 60 min at 30°C. Three-microliter samples of each reaction mixture were addedto Laemmli sample buffer, heated to 90°C for 3 min, and directly applied to a 17% gel for SDS-PAGE. 14C-labeled low-molecular-mass marker proteins were co-electrophoresed (lane M). The gel wasdriedandexposedtoKodakXAR-2film.Kd, Kilodaltons. (B) Proteasesensitivity ofinvitro translationproducts.Cell-free trans-lation was carried out as described above in the absence (-) or presence (+) of microsomes (Mm). After incubation, aliquots of each reactionmixture weresubjectedtodigestionwiththermolysin for60min at4°CinTCbuffer(Pr+).Protease-digested aliquotswere coelectrophoresed withaliquots of undigested product (Pr-). Elec-trophoresis was done as described in the legend to panel A. M, 14C-labeledmarkerproteins.
Translocation of cell-free translation products. Resistance
toprotease digestion ofcell-freetranslation productsunder
conditions that maintained the structural integrity of the
endoplasmic reticulum demonstrated that they had been translocated (Fig. 2B). As acontrol, the 33-kDafull-length polypeptide synthesized in the absence of membranes was completely protease sensitive. Under the same conditions, membrane-dependent products B, C, a, b, and c were protected from protease digestion. Protein C' was not
de-tected in this experiment. Protein A appeared partially
protease sensitive. Protease digestion removed protein A
and introduced a new high-molecular-mass product that migrated slightlyslower thanproteinC. This resultcould be
explained by a transmembrane orientation of protein A which rendersanextralumenalpiecesusceptibletoprotease. Digestion of glycoproteinswith endo F. Todetect N-linked
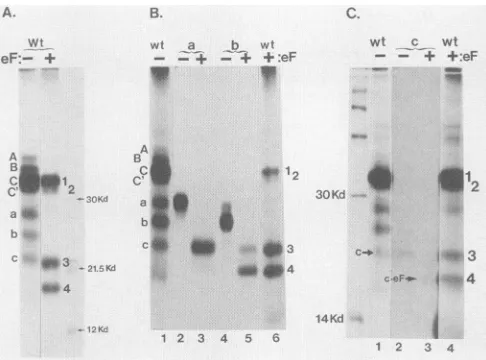
glycosylation of translocated proteins, the mixtureofA,B, C, and C' with a, b, and c was digested with the enzyme endo F (Fig. 3A). The products of
digestion
were fourpolypeptides designated 1, 2, 3,and4indescendingorder of
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.315.561.73.319.2]B.
wt
eF:- +
A
b
c 9443
wt a b wt
- -+ - + +:eF
_
C' 4w1
a
.-
50Kd
af
b
C.
4 403-21.5KtY _ 44
4
3OKd
14Kd
1 2 3 4 5 6
FIG. 3. (A)Digestion ofwtcell-freetranslati C, and C' plus a, b, and c) with endo F tc
carbohydrate.Invitro translation ofwtRNAtrar outin thepresenceof1.5,ulof added microsome,
legendtoFig. 2A.Total productswere subjecte 0.2 U of endo F for 2 hat37°C. Aliquotsofun
endo F-digested (eF+) proteins were coelectro labeled marker proteins as in Fig. 2. (B)Identi translation products a and b in relation to en(
cell-free translationproducts preparedasdescril Fig. 2Awere separated on a preparative gel. I productsaandbwerelocatedbydirectexposur film, excised, and electroeluted. Products a
separately digested by endo F. The electroph proteinsaand b(lanes2and4)and their endo and 5) were compared with total proteins (lai digestedtotalproteins (lane6)duringSDS-PAG] ofcell-free translation product c in relation tc
[35S]methionine-labeled protein c was prepare
other cell-free translation products, digested by paredwith total wt and endoF-digested wt pr above. Lane 1,wtproducts (cisindicated);lan
3,endoF-digestedc;lane4,totalwtproteins di~ Theidentityof endo Fpolypeptide 4 with endo
c(c eF)is indicated. 14C-labeled markerprote phoresed (left lane). kd, Kilodaltons.
molecular size(33, 31, 22, and19kDa,res were consistent with the expected sizes c
polypeptides representingthefull-lengthpr
prM, prM + E, and prM. A polypeptide ( with that of the cleaved capsid (12 to ]
detected. Translation products were next fromapreparative gelanddigestedwithen B, C, and C' collectively yielded polypo
Proteins AandBwerereduced insizebyi
showing that they were glycosylated. Pr appearedidenticaltopolypeptides 1 and 2
Glycosylation of proteinsa,b, and c was a (Fig. 3B and C). Glycoprotein a yielde Glycoproteinsb andcbothyielded
polypep
teins a and b were each approximately 6 polypeptides 3 and 4, respectively. Glyc4kDalargerthan its endoFdigestion produ(
ItappearedthatendoFremovedtwocarb( (GlcNAc2-Man9-Glc3, where GlcNAc
cosamine, Manis mannose, andGlc isglu
eachfromproteinsaand b andonefromc
precursor,potential glycosylation sites liee
prMsequence (discussed elsewhere).
c. Sequencing and immune precipitationof endo F digestion
wt _ wt products. The N and C terminiof endo F digestion products
+ +:eF 1 through 4 were subsequently identified. Initially, the N-terminal 30 residues ofpolypeptides that had been labeled with both [35S]methionineand [3H]leucinewere sequenced. By alignment of the labeled residues, the N-terminal se-quences ofpolypeptides 1 and 2 were demonstrated to be
identical to thatpredictedfor thedengueviruscapsid protein
*
(data
notshown).Sequence
data forpolypeptide
3 is shownin Fig. 4A. Within the first 30 residues, methionine was
detected inposition12 and leucinewasdetected inpositions 3, 11, 23, and 24. The results of sequencing the first 30
cb~ residues ofpolypeptide 4 were identical. The locations of ceF- * 54 methionine and leucine residues within the N termini of
polypeptides3 and 4wereinagreementwith thosepredicted for the N-terminalsequenceofthedenguevirusprM (2, 4, 5, 18, 33), showingthatspecific cleavageat thecapsid-prMsite 1 2 3 4 occurred when microsomes were present during cell-free ionproducts(A, B, translation.
remove N-linked To detect
cleavage
ofthe first23amino acidsof Efromthenscriptswas carried C terminus of the truncated precursor, we prepared rabbit
nas
describedin the antibody to a peptide representing the 15 N-terminalresi-dtodigestionwith dues ofEand demonstrated thatantipeptide antibody
pre-digested (eF-)and cipitatedE fromalysateofdenguevirus-infectedcells. The
phoresed with 14C- same antibody preparation specifically immune precipitated
ification of cell-free polypeptides 1 and 3 (Fig. 4B, lane3), indicating thateach
do F digestion. wt retained the C-terminal E sequences. Precipitation by
pre-bedinthelegendto immune serum is shown as a negative control (lane 4). In
retoKodakXAR-2 contrast, polypeptides 2 and 4 were not immuneprecipitated
and
b were thenby
theantipeptide
antibody andappeartorepresent speciesaretic mobilities of cleaved at the prM-E site. The endo F polypeptides were
Fproducts (lanes 3 therefore identifiableby apparentmolecularsize, N-terminal
ne 1) and endo F- sequence, and C-terminus-specific immune precipitation
E.(C)Identification (Table 1):polypeptide 1= capsid+ prM+ E(the full-length
endo F products. precursor);polypeptide 2 = capsid+ prM;polypeptide 3 =
-d, separated from prM + E; andpolypeptide4 =prM. Consequently,proteins
tendo Fs
and corn- a, b, and c were verified as specific glycosylated cleavagee2,isolatedc; lane products of the 33-kDa precursor. Protein a was identified as gestedwith endo F. prM + E bearing two carbohydrate residues. Proteins b and F-digested product c were identified as alternate glycosylated forms of prM. inswerecoelectro- Cleavagewas membranedependent at both thecapsid-prM andprM-E sites, and cleavage products weretranslocated.
These results confirmand extend data whichsuggestedthat
pectively). These thesignal peptidaseisresponsiblefor cotranslational
cleav-)f unglycosylated ageof the flavivirusstructuralproteins (25); however, they
ecursor,capsid+ do not clarify the significance of the conserved potential consistent in size proteolytic site in the capsid sequence or localize signal 14 kDa) was not function in the truncatedprecursor. Experimentsthat follow
separately eluted address thesequestions.
do F. ProteinsA, Cell-freetranslation oftranscripts bearingmutations in the
eptides 1 and 2. capsid sequence. Two in-frame deletion mutations were
endoFdigestion, introduced into the capsid nucleotide sequence in pTZ/Sal -oteins C and C' DNAbysite-directedmutagenesistotest thepossibilitythat
(datanotshown). cleavageofthecapsidatthe conservedpotential proteolytic lsodemonstrated site(Arg-Lys-Arg; residues 97 to99) isrequiredin associa-d polypeptide 3. tion with signal recognition. The mutant dIC polypeptide )tide 4.Glycopro- lacked an internal 74 amino acids ofthe capsid sequence kDa larger than (residues 13 to86), which should haveabrogated any
auto-oprotein c was 3 proteolytic activity of the capsid or any conformational ct,polypeptide4. requirement for normalprocessing. Mutant dlC-pM lacked ohydrateresidues thetrypsinlike site itself(residues 97 to 99; Fig. 5). In the
is N-acetylglu- absence of microsomes, the predominant product of dlC
icose; Mr- 3 kDa) transcripts was a protease-sensitive 25-kDa polypeptide, :. Inthe truncated consistent in size with that expected for the full-length -ntirelywithin the unglycosylated product of the nucleotide sequence (Fig. 6A).Also notedasaminorspecieswasa22-kDaprotein,the A.
MYt
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.62.305.77.257.2]B.
anti-ERIP./+4
-Ii
eF: - + a 4
_r _m _
1.2 11 1 i i I II I7
1.1
II
O3H-DPM
0 35S-DPM
A-
B- C-
C,-
a-
b-
c-a.
I.
-30kd
-21.5kd
-12kd 0.1 SIL S T1 I I DG111lII I Ii,,I RGI II I I Ii TI G
F SLST RDG E PLM VA KHE RG R PL L F KTTEG*
[image:5.612.64.552.77.357.2]*Predlc*ted N-terminal secuence of orM 1 2 3 4
FIG. 4. (A) Sequencing of polypeptide 3. Cell-free translation was carried out in the presence of membranes and [3H]leucine and [35S]methionine labels. The reaction products were digested with endo F and separated on a modified 17% SDS-polyacrylamide gel.
Polypeptide 3waslocated byexposureof the geltoKodak XAR-2 film andexcised. The proteinwaselectroeluted, andasample containing 2.7x 104dpm of 3H and 4.9x 104dpm of 35Swasfractionatedon aBeckmanModel 890M Protein Sequencer. Thirty residuesweresequenced
from the Nterminus(radiolabel in fractions 1to30is shown, lefttoright). The resultsarecompared with the predictedsequenceof the N terminus ofthe dengue virustype4strain 814669 prM (33). The first fraction is aligned with the N-terminal F(single-letter code), and labeled Mand Lresiduesareindicated in boldface. (B)Immuneprecipitation of endo F-digested cell-free translation products by antibody specific
forC-terminal Esequences. [35S]methionine-labeled cell-free products (lane 1)weredigested with endo F (lane 2). A samplewasincubated
for 16hat4°C with rabbit antibodytoa15-amino-acid peptide representing the predicted N terminus of E (lane 3)orwith preimmuneserum
(lane 4). The immune precipitates werecollected by using staphylococcal protein A-Sepharose CL4B beads, eluted from the beads, and coelectrophoresed with undigested and digested total proteins. eF-, Not digested with endo F; eF+, digested with endo F; anti-E+, radioimmunoprecipitation (RIP) with rabbitantipeptide antibody; anti-E-,RIPwith preimmunerabbitserum.kd, Kilodaltons.
expected sizeof a productinitiating internally at Met-112 of
the wt capsid sequence. The Met-112 codon lies within a
consensus nucleotide sequence favoring initiation (12). In the presence of microsomes, translation of mutant diC transcripts produced three translocated proteins which
[image:5.612.54.298.633.714.2]comigratedwith theproducts ofwttranscripts (proteinsa,b, and c)already identified as glycosylated forms ofprM + E and prM. Similarly, the three low-molecular-mass cell-free translation products of mutant dIC-pM transcripts in the presence orabsenceof microsomes were not distinguishable from those of the wt by size or proteasesensitivity (Fig. 6B;
TABLE 1. endoF-digested productsof cell-free translation ofwtRNAtranscripts
endo F Molecular RP Translation eo size(kDa) N-terminal RIP"
product(s)
peptYde
bySDS- sequence antE IdentitypeptidePAGE anti-B
A, B,C,C' 1 33-34 Capsid + Cb+ prM + E
2 31-32 Capsid - C + prM
a 3 22 prM + prM+ E
b,c 4 19 prM - prM
aRIP,Radioimmunoprecipitation. bC,Capsid.
compare to Fig. 1B). To confirm theiridentity, processed mutant dlCand mutantdIC-pM products were subjected to endo F digestion and subsequent immune precipitation by
rabbit antibody to E sequences at the C terminus of the
precursor(datanotshown).Thisanalysisdemonstrated that dlC anddlC-pM products comigrating withwtglycoproteins
a,b,andc wereidenticaltothem. Inconclusion, cleavageof thecapsid andglycosylation andtranslocation ofcapsidless proteins were not detectably altered by the capsiddeletion mutations.
Cell-free translation oftranscripts bearingmutationsinthe
prM signal. Toidentifythe signalsequencein the truncated
precursor,two mutantDNAswerepreparedwithalterations inthecandidatesignal (residues 100to 113) (Fig. 5). Mutant dlss DNA had an in-frame internal deletion of sequences
encoding 10 amino acids of the hydrophobic segment
(resi-dues 102to 111). Mutantsubss bears a substitution ofthe codon for Ile at position 109 by a codon for Lys, which interruptsthehydrophobicsequence.The results of cell-free translation of dlsstranscriptscomparedwithwtareshownin Fig. 7A, and the results of protease digestion of subss products compared with wt are shown in Fig. 7B. The phenotypes of these two mutants were indistinguishable. The signal-minus precursor synthesized in the presence of microsomeswasnotprotectedfrom proteasedegradation,as
A.
-I
on November 10, 2019 by guest
http://jvi.asm.org/
DNA
wt X
Map
r-Capsid
Bgl II
Modification
. Env
r-' preM
Sal I
Deletecapsidsequences fromaa13throughaa86 Aaa1a-"-a86
RKRK
R _AaalO2-8a111
K
Delete -Arg-Lys-Arg preceding preM signal
sequence
Delete intemal 10aas(102-p 111) from preM signal
sequence
Substitutelle1i8within preM signal sequence
by Lys
FIG. 5. Diagrams of Sal DNA andmutationsproducedinSalDNAbysite-directedmutagenesisofsingle-strandedpTZ/SalvectorDNA.
aa,Amino acid. _, Consecutive hydrophobic residues; 3, noncoding region.
shown for subss. Failure ofglycosylation and cleavage is
shown for both dlss and subss. The 33-kDa truncated pre-cursor wasunaltered, despitethepresenceof microsomes in thetranslation reaction mixture. Interruptionordeletion of
A.
dIC
:Mm
+ -+: Pr
w s1p-iw,
+ ;
MP,
the hydrophobic segment at once prevented translocation, glycosylation,andcleavage of theprecursorateither poten-tial site. These results confirmed a signal function for the hydrophobic segment atthe capsid-prM juncture.
DISCUSSION
In vitro translation of RNA transcripts prepared from cloneddenguevirus DNAencodingthecapsid, prM,and the
A.
wt Mm: - +
qW...-'
dissl
Bwt subss
Mm: - + - +
Pr:- + - + + +
a,
.. _.
.,. it
2 1Ka ..:.. .:::.:::..
*.,... 30
Kd-21Kd
[image:6.612.123.499.72.315.2]21 Kd
FIG. 6. Cell-free translation of dlC (A)and dlC-pM (B) mutant RNAtranscripts and proteasesensitivity of the products.
Transla-tion wascarried out as described in the legend to Fig. 2A in the
presence (+) or absence (-) of 1.5 1.l of microsomes (Mm).
Thermolysin digestionwascarriedoutasdescribed in the legendto
[image:6.612.91.286.419.655.2]Fig. 2B. Pr-, Undigested products; Pr+, thermolysin-digested products. Productswere coelectrophoresed with "C-labeled stan-dards(lane M), showninpanelB. Kd, Kilodaltons.
FIG. 7. Cell-free translation of dlss (A) and isubss (B) mutant RNA transcripts and protease sensitivity of the subss product.
Conditions were as described in the legend to Fig. 6. Mm-,
Microsomes absent; Mm+, 1.5 of microsomes present; Pr-, productsnotdigestedwiththermolysin; Pr+, products digestedwith
thermolysin. Kd,Kilodaltons.
dlC
dl C pM
diss
subss
r
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.326.564.454.675.2]N-terminal23amino acids of E resulted in the
synthesis
ofaprecursor polypeptide which was cleaved. Cleavage
prod-ucts
representing
thecapsid
+prM, prM+ E,andprMwereidentified,
and glycosylated forms of prM + E and prMbearing
twocarbohydrate
residues weredetected(proteinsaand
b).
The33-kDain vitroprecursorcontainedoneconsen-sus site
(Asn-X-Ser/Thr)
fortheaddition ofN-linkedcarbo-hydrate,
at Asn-182 in prM. However, prM contains anadditional
site, Asn-X-Cys (in
the conserved sequenceAsn-Lys-Cys-Thr;
residues145to148),
whichhasbeen showntohave
carbohydrate
acceptorfunctioninvitro (1)and in vivo(23).
We presume that both Asn residues 145 and 182 areglycosylated
inproteins
a and b. A third glycosylatedcleavage product,
prMbearing
asingle carbohydrate
resi-due,
wasalso detected(protein
c).Cleavage
ofthe precursorwaswholly dependentonthe presenceofmicrosomesduring
translation. Evidence that in vitro cleavage events are a
model of in vivo processing ofthe dengue virus precursor
polyprotein
wasprovided
by sequence analysis of the NterminusofprMproduced invitro; its sequencewas
identi-caltothe
predicted
Nterminus ofprM deducedbycompar-ing
thedengue virus sequencetothose ofotherflaviviruses for which the N-terminal sequenceofprMhas beendirectly
determined (2, 4, 33). Theapproximate
accuracy of thecleavage
event atthe prM-Esitewasestablishedbyimmuneprecipitation
ofendoF-digested
products with an antibodydirected
against
thepredicted
first 15 amino acids ofE(33).The
capsid-prM
andprM-E cleavage sites inthe contextofhydrophobic
sequences thatprecedeeach (5, 18, 19, 33)arehighly
favored sites for the action of signal peptidase (27),the
presumed cleavage
enzyme.To test the
hypothesis
thatsignal peptidase
isuniquely
responsible
forcleavage
of thecapsid
and prM and toidentify
thesignal,
the phenotype ofmutant dengue virusDNA
containing
eitheran altered capsidsequence orsignalsequence
(residues
100 to 113) was determined in cell-freetranslation
experiments.
Thecapsid
mutationsweregener-ated to rule out the
possibility
that the capsid is initiallycleaved at or upstream from a
pair
of basic amino acids(residues
98 and 99) which precede the signal for prM, inassociationwith
recognition
ofthatsignal.
TheSindbisviruscore
protein,
which lies N terminal topE2
and El in theSindbis virusstructural
polyprotein,
initiatesprocessing by
a similarcleavage
event which isautocatalyzed
(9). Mutantswith a
large
internal deletion of capsid sequences (mutantdIC)
or a deletion of theputative cleavage
site (mutantdlC-pM)
wereanalyzed.
Themutationin the dlCpolypeptide
shouldhave abolished any
enzymatic activity
ofthecapsid
and removed alternate
trypsinlike cleavage
sites upstreamfrom residues 98 and 99. However, neithermutation of the
dengue
viruscapsid
hadadetectableeffectonprocessing
ofthe precursor
polypeptide, suggesting
thatcleavage
of thecapsid by
anactivity
other than thatofsignal peptidase
isnota
prerequisite
forprocessing.
Asaconsequence,it wouldbepredicted
that thesignal
segment is the C terminus of themature
capsid protein. Signallike
function ofthe candidatesegment
(residues
100to113)
wasestablishedby analysis
of mutantpolypeptides
withalarge
internal deletion ofhydro-phobic
residues(mutantdlss)
orwith aninterruption
in thehydrophobic
sequenceby
aninternalsubstitutionofasingle
hydrophilic
amino acid forahydrophobic
amino acid(Ile
-*Lys;
mutantsubss).
Both mutations of theputative signal
prevented
translocationofthepolypeptide.
Failureoftrans-locationwas associated with failure of
processing,
suggest-ing
that access of intralumenalsignal peptidase
to the precursorwasrequired
forcleavage.
Both mutantpolypep-tides contained the hydrophobic segmentatthe C terminus ofprM (residues 246 to 279) thought to be the signal for E (14, 18, 22, 33). This segment did notmediate translocation of the truncated precursorpolypeptide encoded bypTZ/Sal transcripts, perhaps because the sequences downstream (residues 279 to 302) are insufficient in length to drive translocation (17) or because, in fact, translocation of E is dependent on the signal for prM. These findings together
suggestthatprocessing of theflavivirus precursor is initiated by cotranslational recognition of the prM signal segment. Translocation affords access of intralumenal signal peptidase to hydrophobic segments preceding the N termini of both prMand E. Signal peptidase effects cleavage at thepredicted sites.
The cleaved capsid protein was not detected in these experiments. The capsid should have been labeled at a reduced specific activity with respect to prM, since sequence datapredict that it contains only 5 methionines among 113 residues, whereas prM contains 10 methionines among 166 residues. This disparity alone would probably not account for our failure to locate the capsid among labeled translation
products. Once cleaved, the capsid may be unstable due to
proteolysisor maybe poorly resolved under the electropho-retic conditions employed. Chimeric glycoproteins contain-ing the capsid were identified, and these appeared to be translocated (proteins A and B). The finding of uncleaved productscontainingthe capsid and prM may be an artifact of
inefficiency ofcleavage duringtranslocation in vitro. How-ever,achimera of the capsid and prM, the 30-kDa glycopro-tein NVX, has been detected among flavivirus proglycopro-teins
synthesized during infection (13, 29, 30), suggesting that
cleavage ofthecapsid may alsobe inefficient in vivo.
ACKNOWLEDGMENTS
I thank Bangti Zhao and Ching-Juh Lai forthe generousgift of cloneddengue virusDNA,Suzanne Emerson andPeter L. Collins for manyhelpful discussions, John Coligan and MichaelRaumfor protein sequencing,MyronHill for preparation ofoligonucleotides, and ChristinaFonseca andLindaJordan for editorial assistance.
LITERATURE CITED
1. Bause, E., and G.Legler. 1981. Therole ofthehydroxy amino acid in the triplet sequence Asn-Xaa-Thr (Ser) for the N-glycosylation step during glycoprotein biosynthesis. J. Bio-chem. 195:639-644.
2. Bell, J. R., R. M. Kinney, D. W. Trent, E. M. Lenches, L. Dalgarno,andJ. H. Strauss. 1985. Amino-terminal aminoacid sequencesof structuralproteins of three flaviviruses. Virology 143:224-229.
3. Boulton, R. W., and E. G. Westaway. 1972. Comparison of togaviruses: Sindbis virus (group A) andKunjin virus(group B). Virology 49:283-289.
4. Castle,E. T., U.Leidner,G. Wengler, and G. Wengler. 1985. Sequence analysisof the viral coreprotein and the membrane-associated proteins Vl and NV2 of the flavivirus West Nile virus and the genome sequence for these proteins. Virology 145:227-236.
5. Coia, G., M. D. Parker, G. Speight, M. E. Byrne, and E. G. Westaway. 1988. Nucleotide and complete amino acid
se-quences ofKunjin virus: definitive gene order and characteris-ticsofthevirus-specified proteins. J.Gen.Virol. 69:1-21. 6. Dalgarno,L., D. W. Trent, J. H.Strauss,andC. M. Rice. 1986.
Partial nucleotide sequence ofMurrayValleyencephalitisvirus: comparisonof the encodedpolypeptideswithyellowfevervirus structural and nonstructuralproteins.J.Mol.Biol. 187:309-323. 7. Deubel, V.,R.M. Kinney,andD.W. Trent. 1986. Nucleotide
sequence and deduced amino acid sequence ofthe structural proteins ofdengue type 2 virus, Jamaicagenotype. Virology 155:365-377.
on November 10, 2019 by guest
http://jvi.asm.org/
8. Dunn, J. J., and F. W. Studier. 1983. Complete nucleotide sequence of bacteriophage T7 DNA and the locations of T7 genetic elements. J. Mol.Biol. 166:477-535.
9. Hahn, C. S., E. G. Strauss, and J. H. Strauss. 1985. Sequence analysis ofthreeSindbis virusmutantstemperature-sensitive in the capsid protein autoprotease. Proc. Natl. Acad. Sci. USA 82:4648-4652.
10. Hahn, Y. S., R.Galler,T. Hunkapiller, J. M. Dalrymple, J. H. Strauss, and E. G. Strauss. 1988.Nucleotidesequenceof dengue 2 RNA andcomparison of the encoded proteins with those of other flaviviruses. Virology 162:167-180.
11. Hase, T., P. L. Summers, K. H. Eckels, and W. B. Baze. 1987. Anelectron and immunoelectron microscopic study of dengue 2 virus infection of cultured mosquito cells: maturation events. Arch. Virol. 92:273-291.
12. Kozak, M. 1986. Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell44:283-292.
13. Lyapustin, V. N., Y. V. Svitkin, T. Y. Ugarova, V. A. Lash-kevich, and V. A. Agol. 1986. Atentative model of formation of structural proteins of tick-borne encephalitis virus (flavivirus). FEBSLett. 200:314-316.
14. Mackow, E., Y. Makino, B. Zhao, Y.-M. Zhang, L.Markoff,A. Buckler-White, M. Guiler, R. Chanock, and C.-J. Lai.1987. The nucleotidesequence of dengue type 4virus: analysis ofgenes coding for nonstructural proteins. Virology159:217-228. 15. Mason, P. W., P. C. McAda, T. L. Mason, and M. J. Fournier.
1987. Sequence of the dengue-1 virus genome in the region encoding the three structural proteins and the major nonstruc-tural protein NS1. Virology161:262-267.
16. McAda, P. C., P. W. Mason, C. S.Schmaljohn,J. M. Dalrymple, T. L. Mason, and M. J. Fournier. 1987. Partial nucleotide sequenceoftheJapanese encephalitis virusgenome. Virology 158:348-360.
17. Rappoport, T. A., R. Heinrich, P.Walter, and T. Schulmeister. 1987. Mathematical modeling oftheeffects of the signal recog-nition particle on translation and translocation of proteins across the endoplasmic reticulum membrane. J. Mol. Biol. 195:621-636.
18. Rice, C. M., E. M. Lenches, S. R. Eddy, S. J. Shin, R. L. Sheets, andJ. H. Strauss. 1985. Nucleotide sequence ofyellow fever virus:implications for flavivirusgeneexpressionandevolution. Science229:726-733.
19. Rice, C. M., E. G. Strauss, and J. H. Strauss. 1986.Structure of theflavivirusgenome, p. 279-326. InS. Schlesingerand M. J. Schlesinger (ed.), The Togaviridae and Flaviviridae. Plenum Publishing Corp.,New York.
20. Russell, P. K., and A. Nisalak. 1967.Denguevirus identification by the plaque reduction neutralization test. J. Immunol. 99:
291-296.
21. Sanger, R., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
22. Speight, G., G. Coia, M. D. Parker, and E. G. Westaway. 1988. Genemapping and positive identification of the non-structural proteins NS2A, NS2B, NS3, NS4B and NS5 of the flavivirus Kunjin and their cleavage sites. J. Gen. Virol. 69:23-34. 23. Stenflo, J., and P. Fernlund. 1982. Amino acid sequence of the
heavy chain of bovine protein C. J. Biol. Chem. 257:12180-12190.
24. Strauss, E. G., and J. H. Strauss. 1985. Assembly of enveloped animalviruses,p. 205-234.InS.Casjens (ed.), Virus structure andassembly. JonesandBarlett, Boston.
25. Svitkin, Y. V., T. Y. Ugawara, T. V. Chemovskaya, V. N. Lyapustin, V. N. Lashkevich, and V. L. Agol. 1984.Translation of tick-borne encephalitis virus (Flavivirus) genome in vitro: synthesis of two structural polypeptides. Virology 110:26-34. 26. Trent, D. W., R. M.Kinney, B. J. B. Johnson, A. V. Vorndam,
J. A. Grant, V. Deubel, C. M. Rice, and C. Hahn. 1987. Partial nucleotide sequence of St. Louis encephalitis virus RNA: structuralproteins, NS1, NS2a, and NS2b. Virology 156:293-304.
27. von Heijne, G. 1986. A new method for predicting signal sequencecleavage sites. NucleicAcidsRes. 14:4683-4690. 28. Wengler, G., G. Wengler, and H. J. Gross. 1978. Studies on
virus-specific nucleic acidssynthesized in vertebrate and mos-quito cells infectedwithflaviviruses. Virology 89:423-427. 29. Westaway, E. G. 1973. Proteinsspecified bygroup Btogaviruses
in mammalian cells during productive infections. Virology51: 454-465.
30. Westaway, E. G. 1987. Flavivirus replication strategy. Adv. Virus. Res. 33:45-90.
31. Winkler, G., F. X. Heinz, and C. Kunz. 1987. Studies on glycosylation of flavivirus E proteins and the role of carbohy-drateinantigenicstructure. Virology 159:237-243.
32. Wright, P. J., H. M. Warr, and E. G. Westaway. 1981.Synthesis of glycoproteins in cells infected by the flavivirus Kunjin. Virology 109:418-427.
33. Zhao, B., E. Mackow, A. Buckler-White, L. Markoff, R. M. Chanock, C.-J. Lai, and Y. Makino. 1986. Cloning full-length
denguetype 4 viralDNAsequences: analysis ofgenes coding for structuralproteins. Virology155:77-88.
34. Zhao, B., G.Prince, R. Horswood, K. Eckels, P. Summers,R. Chanock, and C.-J. Lai. 1987. Expression of dengue virus structuralproteins and nonstructural protein NS1 by a recom-binant vaccinia virus. J. Virol. 61:4019-4022.
35. Zoller, M. J., and M. Smith. 1983. Oligonucleotide-directed mutagenesis ofDNAfragments cloned into M13-derived vec-tors. MethodsEnzymol. 100:468-500.

![FIG. 4.fromforfor2.7terminus(lanecoelectrophoresedPolypeptideMradioimmunoprecipitation[35S]methionine and (A) Sequencing of polypeptide 3](https://thumb-us.123doks.com/thumbv2/123dok_us/1325004.86298/5.612.64.552.77.357/fig-fromforfor-terminus-lanecoelectrophoresedpolypeptidemradioimmunoprecipitation-methionine-and-sequencing-polypeptide.webp)