Copyright © 1991, American Society for Microbiology
Elements in the
Transcriptional Regulatory Region Flanking
Herpes
Simplex Virus Type
1oriS
Stimulate Origin Function
SCOTT W. WONGAND PRISCILLA A. SCHAFFER*
Laboratory of Tumor Virus Genetics, Dana-FarberCancer Institute, andDepartmentof Microbiology and Molecular Genetics, HarvardMedical School, Boston, Massachusetts 02115
Received 12 December 1990/Accepted 7 February 1991
Like other DNA-containing viruses,the threeorigins ofherpessimplexvirustype1(HSV-1)DNA replication are flanked by sequences containing transcriptional regulatory elements. In a transient plasmid replication assay,deletion ofsequences comprisingthetranscriptional regulatory elementsofICP4andICP22/47, which flankoriS, resulted inagreater than 80-fold decreaseinorigin function comparedwith aplasmid, pOS-822, whichretains thesesequences. Inanefforttoidentify specific cis-acting elements responsible forthiseffect,we
conductedsystematic deletion analysis of the flanking region withplasmid pOS-822 and tested the resulting mutantplasmids for origin function. Stimulation by cis-acting elements was shown tobe both distance and orientation dependent, as changes in either parameter resulted in a decrease in oriS function. Additional evidencefor thestimulatory effect of flankingsequences onorigin functionwasdemonstrated by replacement of thesesequenceswith thecytomegalovirus immediate-earlypromoter,resultinginnearly wild-type levels of oriS function. In competition experiments, cotransfection of cells with the test plasmid, pOS-822, and
increasingmolarconcentrations ofacompetitor plasmid which contained theICP4and ICP22/47 transcrip-tionalregulatory regionsbutlackedcore origin sequencesresultedinasignificantreductionin thereplication
efficiency ofpOS-822, demonstrating thatfactors which bind specifically tothe oriS-flanking sequences are
likely involved as auxiliary proteins in oriSfunction. Together, these studies demonstrate thattrans-acting
factorsandthesitestowhichthey bind playacritical rolein theefficiency of HSV-1 DNA replication from oriS intransient-replication assays.
Initiation ofherpes simplex virus type 1 (HSV-1) DNA
replication beginsatdiscretesequenceelementsororigins of DNAsynthesis and requires theexpression ofatleastseven
viralgenes(Fig.1A) (55). The HSV-1genomecontains three
origins of DNA synthesis: a single copy of oriL, located
within theunique longsegmentof thegenome(51),andtwo copies oforiS, located in the invertedrepeats flanking the
uniqueshortsegment(Fig. 1A) (49). oriLisnotessentialfor virus growth, since deletion mutants lacking a functional copy of oriL have been isolated and are capable of nearly
wild-type levels of replication (39). At least onefunctional copy of oriS appears to be required, since attempts to constructviralmutantslacking both copies of oriShave been unsuccessful (7).
Comparison ofthe DNAsequences of oriL and oriS has revealed similar structural organization and extensive se-quence homology (51). oriL is contained within a perfect
144-bp palindrome, while oriS is locatedwithinanimperfect
palindrome of 45 bp (49, 51). Both origins can support plasmid replication in transfected cells whenessential
repli-cation proteinsareprovidedintransby superinfection with
HSV-1. Furthercharacterization of the HSV-1 origins has focused on oriS rather than oriL because the large oriL
palindrome is deletion-prone upon cloning in bacteria(39, 51).
Thecore origin ofHSV-1 oriSconsists ofa 67- to 90-bp
sequence thatcontains an imperfect 45-bp palindrome(Fig.
2) (11, 31, 49). Included within the palindrome are two binding sites for the origin-binding protein, the product of the UL9gene, that flank a central 18-bp A+T-rich region
(37). It has been postulated that short inverted repeats are
*Correspondingauthor.
capableofformingcruciformstructuresinDNA, suggesting that the palindrome in oriS may exist functionally in this form. Further delineation of the structural requirement for oriSwasprovided byDeb andDoelberg,who used deletion
analysis to demonstrate that the arm of the palindrome
adjacent to the short flanking sequence is not required for DNA replication (11). This observation, together with the
properties ofa seriesofdinucleotide insertion mutations in the center of the A+T-rich region that maintained the
integrityof thepalindromeandproducedanoscillating effect onorigin function,arguethat theorigindoesnotexist in the
expectedcruciformstructureasfirstpredicted, but exists in a linear configuration (31). The role ofsequences in
main-tainingoriS inafunctional configurationisnotknown. Enhancement oforiginfunctionby transcriptional regula-tory elements is a common phenomenon among DNA vi-ruses. Firstobservedinpolyomavirus (17),thisphenomenon has also beenshown for simian virus40(SV40) (8, 15, 24, 26, 28, 30), bovinepapillomavirus (32, 48),adenovirus(45, 53),
and Epstein-Barrvirus (23, 44, 56). Thus,for example, the core origin of SV40 DNA replication consists of a 65-bp segment that contains numerous binding sites for large T
antigenand a 17-bp A+T-rich region. The core origin has been demonstrated to be sufficient to support initiation of DNA replication both in vivo and in vitro. Replication efficiencyinvivo, however, hasbeen showntobe
substan-tiallyincreasedby sequenceelements whichflank the core
origin (8, 15, 24, 26, 28, 30, 36).Thesesequencesinclude GC
boxesaswellasthetranscriptionalactivatorelements of the SV40 enhancer.
Based on findings from these systems, DePamphilis (16)
has suggested that eukaryotic origins of DNA replication
contain two primary components: a core component
re-quired specificallyfor the initiation ofDNAreplication and 2601
on November 10, 2019 by guest
http://jvi.asm.org/
A.
HSV-1 Genome
Mapunits
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
5 89 2930 42 52
a b b' a' c' ca
C3 *_ _ -
C-O1'iL OriS Ori
UL
us__- ~~~~~~~~~~~~~I
__- ~~~~~~~~~~~~~~~~~~~~~~~~~~~I
B.
ICP4
Ba
A.
base pairs 100 200 300 400 S00 600 700 800
ICP44* 1* a
oris 1-ICP22/47
B.
Bs SS Ba
I orICP247
H N
FIG. 1. HSV genome. (A) Beneath the scale of map units is
shownadiagram of the HSV-1 genome, indicating the locations of
reiteratedsequences (a, b, and c) and theirinvertedcounterparts (a',
b', and c') as well as the positions of the three origins of DNA
replication (oriL and two copies of oriS). Also shown are the locations of the seven genes that specify proteins required for
HSV-1 DNAreplication, UL5, 8, 9, 29, 30, 42, and52 (55). (B)
Diagram ofthe 822-bpBamHI fragment from plasmid pIE3CAT,
containing theintergenic region betweenICP4 andICP22/47. The arrowsdenotethetranscriptional start sites for ICP4 andICP22/47; the box represents the core origin oforiS. Restriction sites: Ba, BamHI;S,SmaI;Bs, BssHII; H,HpaIl;and N, Narl.
ICP4-GATCOGTGTCGGCCAG30 CTCX3TG153 ACT3AT300G COTCTOl GCTCATATAGT 60
COCaGG= GG03GGAAGAGGA}CGCGGAGC G OG-C 18D
GOCcACCccnGAACW~ ~crroco ISOOGCAGczOG 8
CCOWGWCG
CGG(~ ~ ~ ~
A C2TCCG= ATCTGCATG 2401300500GAGGGC0GT G0GrGC000W ATGG0C~~ X(~ 300
Sma
GATCCGGCM3G T -
C~
Aa3WCA1GM380Tooccorn GoG
3~
UcfA&
42DSma BssHII
(T,30s=
CjTGX=C
C 540Hpa 11 UL9site1
GCCGGTMAAGAGTGAGAAGGCAdG TrCACITT GTCOCAATATATATATATTA 600
UL9 site 2 Smat
TrAG~GGOG GrGOQ4AWXT=S3f300T =OmCTIr= O=GW=i3 ... 800
SinalSma NorINar ICP22/47
CCC53GC)GG(53~G1-CTCTCIGCsATAAMG C X OACG 720
c00GAOGG 03GOcA0GAoG_o0)3AG0_GC5o5GS3_A4Co_5(5000_33770
an auxiliarycomponent composed of eitheranenhanceror
promoterelementthat functions in transcriptionaswellasin DNAreplication. The auxiliarycomponentinsomesystems isabsolutely essential for DNA replication, while itappears
tobe dispensableinothers (16). Although the precise man-nerby which DNA replication is activated by transcription
factors is notknown, several hypotheses have been postu-lated. Recent studies by Cheng and Kelly support the hypothesis that transcription factors inhibitnucleosome
for-mationthroughout theSV40 origin andpromoterregulatory region (9). Incell-freeorigin-function assaysinwhichSV40 origin-containing plasmids were associated with nucleo-somes and origin function was diminished, these authors reported that the introduction ofa unique nuclear factor I
(NF-I) binding site adjacent to the core origin enabled the
origintoreplicateasefficientlyasthewild-type origin. These
experiments, the authors suggest, are consistent with the concept that transcription factors generate a nucleosome-free region, making DNA more accessible to replication machinery.
IntheHSV-1genome, theorigins ofDNAreplication are located in contexts similar to those ofother DNAviruses.
oriLispositioned between the divergent transcriptionalstart
sitesof thegenesencoding the major DNA-binding protein ICP8and the DNA polymerase (39, 51). The transcriptional
regulatory elements surrounding oriL are not well defined. oriS, like oriL, is also located between divergently
tran-scribed genes. These genes encode the immediate-early
proteins ICP4 and ICP22/47 (49) (Fig. 1 and 2). The
tran-scriptional regulatory elements within whichoriSresidesare
wellcharacterized and collectively exhibit thepropertiesof anenhancer element (40-42). (Fig. 2A and B). A variety of
recognized transcription factors have been reportedtobind
specifically to oriS-flanking sequences. These include the potent HSV-1 transactivator VP16(4)and cellular
transcrip-FIG. 2. Intergenic region between immediate-early genes ICP4 and ICP22/47. (A) Beneath the scale (inbase pairs)is shownthe 822-bp BamHI fragment containing the intergenic region between ICP4and ICP22/47. Thediagram shows the locationsof the
tran-scriptional start sites forICP4 and ICP22/47(arrows), aswell the binding sites for severalrecognized transcriptional regulatory
ele-ments.TAATGARATmotifs that bind VP16areshownashatched boxes;Spl binding sitesareshownasstippled ovals; TATA boxes areshownasblackrectangles;theGCGGAAC motifisshownasan open box; an NF-III-binding site is shown as a checkered box; CCAATbox-likesequencesareshownasopencircles;and thecore origin, oriS, is shown as a large rectangular box. (B) Nucleotide
sequence of the 822-bp fragment shown in panel A. Again, the locations of thetranscriptionalstartsitesareshownas arrows.The TAATGARATmotifs, Spl binding sites, TATA boxes, GCGGAAC motifs, NF-III-binding site, and CCAAT box-like sequences are boxed, underlined by dashed lines, underlined by bold lines, boxed withdashed lines, underlined by double lines, and enclosed with roundedboxes,respectively. Thetworecognized UL9 binding sites are denoted by dark lines, and the oriS palindrome is shown as
convergentarrows.
tion factors Spl, nuclear factor III (NF-III) (1), and the factor recognizing the GCGGAAC sequence shown by Triezenberget al. tobe requiredfor efficient expression of ICP4 (50). Collectively, the transcriptional regulatory
ele-ments located on both the right and left sides of the core
origin promote transcription from both the ICP4 and
ICP22/47 promoters early in infection, resultingin the syn-thesisofthree of the five HSV-1regulatory proteins (40, 47).
ICP4 is essential for virus replication and acts to repress
transcription of immediate-early genes and activate tran-scriptionofearlyandlategenes(13, 14). The roles ofICP22
andICP47,bothimmediate-early proteins,havenotyetbeen
determined; however, neither is essential for virus
replica-tion incell culture (47).
GTCGCAGAGGGCCGTCGGAGCGGACGGOGT CGGCATCGGATC
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.327.552.75.365.2] [image:2.612.62.299.83.245.2]Early studies designed toidentify the cis-acting elements involved in oriS function included analysis of the intergenic region between the transcriptional start sites of ICP4 and
ICP22/47 (49). These studies demonstrated that plasmids
containing this region could replicate in transfected cells when infected with HSV-1. Systematic deletion of se-quences inthis region ledto our presentunderstanding of the
limits of oriS. Although not demonstrated in these early
studies,Stow et al. postulated that elements in the
transcrip-tional regulatoryregions flanking oriS might have an effect onoriSfunction(49). In this report, we presentevidencethat
these elements have asignificant stimulatory effect on oriS
function intransient DNAreplicationassays. MATERIALS AND METHODS
Viruses and cells. The KOS strain of HSV-1was used as the wild-type virus, and all viral DNA fragments were
derived fromKOS DNA. Virus was grown and assayed in Vero cells (13), which were propagated as described previ-ously (13).
Plasmids. Plasmid pOS-822, containing the intergenic
re-gionbetweenthetranscriptionalstartsitesof the
immediate-early genes ICP4 and ICP22/47 (Fig. 2Aand B), was
con-structedby digesting pIE3CAT(13)withBamHIandcloning the resulting 822-bp fragment into pGEM7Zf(+). Plasmid
pOS-230 contains the 227-bp SmaI fragment of pOS-822
insertedinto pGEM7Zf(+) (Fig.1B and 2B).PlasmidpOS-80
wasconstructed by digesting the 227-bp SmaI fragment with
HpaII and NarI andinserting the resulting 80-bp fragment containing the minimum recognized oriS sequence into
pGEM7Zf(+) (Fig. 1B, 2B, and3A). The referenceplasmid
pBR325-ori-230 contains the 227-bp SmaI fragment inserted
into pBR325. Plasmids in which sequences were deleted
starting from the ICP4 transcriptional start site ofpOS-822
(Fig. 4) were generated by digestion with HindIIl at the
single HindIII site located in the polylinker flanking the
intergenic region. The linearized plasmids were then incu-bated with Bal-31 nuclease for various lengths of time. Bal-31nuclease-treated DNAs were blunt ended with mung bean nuclease, and Hindlll linkers were added to
recircu-larize theplasmids. Fragments containing the desired
dele-tions were cloned into M13mpl9 and sequenced by the
dideoxynucleotide method to determine the deletion
end-points.
Plasmids containing internal deletions in theinter-genic region (Fig. 5A) were constructed by cleavage with BssHII(Fig. 1B), digestion with Bal-31, and incubation with
mung bean nuclease to generate blunt ends. BssHII linkers
were added to the deletion endpoints, and the limits of the
deletionwere determinedby DNA sequencing inM13mpl9
asdescribedabove.
Thetranscriptional regulatory element between the ICP4 start site and core origin was inverted as follows. The
HindIIl
site ofdeletion clone pOS-H302 (Fig. 4) wasre-placed withauniqueSpeI sitetogenerateplasmid pOS-S302 (Fig. 6A). PlasmidpOS-822 wasdigested with SacI, which
cleaves the DNA at a unique site within the polylinker
downstream ofthetranscription start siteofICP22/47. The
linearizedDNA was thenincubatedfor asufficienttime with
Bal-31 nuclease to delete both the ICP22/47 transcription
start site and oriS. The nuclease-treated DNA was then treated with mung bean nuclease, and SpeI linkers were added. Theresulting plasmid, containing onlythe transcrip-tional regulatory elements located between the ICP4
tran-scription start site and the core origin, was cloned in both
A.
SaseCalrs 1C0 200. 300 400 5cc cc ci 8
--C.P4
uOS 23
cOS-230
B. ;, oijr5 4 -r -5
0
o < 1>1 o 0 el
g rs - C
r ,
0 4n0 @n bs0 0
2. .~c.-rs
00
_ qw
C.
30
)- 20 N N N
25
z 1o
0)2
< 10
0.
FIG. 3. Replication of oriS-containing plasmids. (A) The 822-bp BamHI fragment containing the intergenic region between ICP4 and ICP22/47 as described in Fig. 2A is shown beneath the scale (in base pairs). The limits of the 80-, 227-, and 822-bp oriS-containing fragments that were cloned into pGEM7Zf(+) to generate plasmids pOS-80, pOS-230, and pOS-822, respectively, are shown beneath the diagram of the BamHI fragment. (B) Southern blots of replicated plasmid DNA. Total cellular DNA was isolated from Vero cells transfected with either vector DNAororiS-containing plasmidDNA and superinfected with HSV-1 strain KOS at 8, 14, and 20 h postinfection. DNA wasdigestedwith EcoRI and DpnI,run ona1% agarose gel, and hybridized to a32P-labeled pUC8 probe. The arrow denotes the position of the replicating internal standard plasmid
pBR325-ori-230. (C) Replication efficiencies of oriS-containing
plas-mids. Each lane from the autoradiograph shown in panel B was
subjectedtoscanningdensitometry,and curves for the test plasmid and internal standard plasmid at the various times postinfection were established. The values for replication efficiency were derived bycalculatingthe ratio of the area under the scanning curve of each plasmid tothat of thereplicatinginternal standard as 1. Under these conditions, pOS-80replicated only approximately 0.25-fold as
effi-cientlyas the interal standardin all tests.
orientations into plasmid pOS-S302toyield plasmids pOS-822(s) and
pOS-822(inv) (Fig.
6A).Plasmids containing heterologous DNA inserts between the intergenic transcriptional regulatory sequence and the coreoriginwereconstructedby
inserting
blunt-ended DNAfragmentsfromMspI-digested pBR322DNA towhichwere
attached SpeI linkers into the SpeI site of
plasmid
pOS-822(s).
All plasmidswerepropagatedinEscherichia coliHB101, purified bythe standardalkalimethod,and banded twice
by
CsCl
equilibrium
centrifugation.
oriSreplication assay. Vero cells(5 x 105) in60-mmdishes
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.322.554.79.388.2]A.
::jseDa s !Z0 200 300 400 500 6CO 72 57
ICP4' a A a
;97n. -. 0 --0 -
n---DOS- H 7.75
p0SH60B
D)OS-"5 pOS-H42 p0S-H443 p0S-H389
pOS H302
B.
C.
'4
0
.0
a:
IN
CDu) o'4 _ r- '4r s007°
O I =: =' I...m.... ...= = = =
C.
A.
se z)a.,s OC00 200 300 400 500 600 700 800
ISI
or,s ICP2'247
B d45 B d75
B3d D5
B.
U w
U z
0
'4
lj
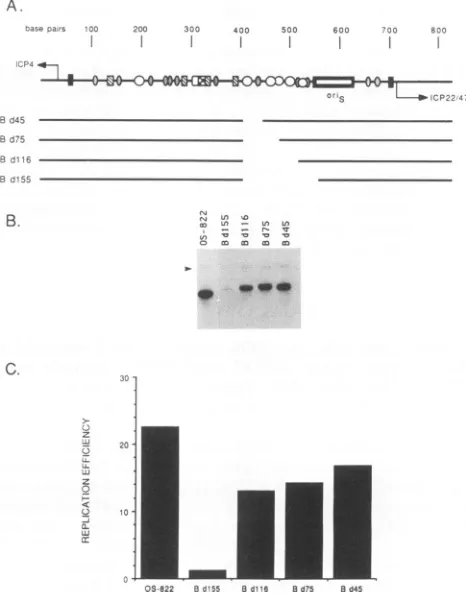
Q-FIG. 4. Replication oforiS-containing plasmids lacking various
amounts of left-hand flanking sequences. (A) Diagram of deletion
mutant genotypes. The intergenic region between ICP4 and ICP22/47 shown is described inthelegend to Fig. 2A.The number ofeachplasmid describes the number ofnucleotidesremaining after Bal-31 nuclease treatment. (B)Southern blots ofreplicated Bal-31 deletionplasmid DNAs. TransientDNAreplication assays of mu-tant plasmids and of the replicating internal standard plasmid pBR325-ori-230 (arrow)wereperformedas described inthelegend
toFig. 3B.(C)Replication efficiencies of deletionmutantplasmids werecalculated as described in the legendtoFig. 3C.
were transfected by the calciumphosphate method with a
mixture containing test plasmid DNA, internal reference
plasmidpBR325-ori-230 DNA, and salmon testis DNA (39).
After 4 h of incubation at 37°C, cells were shocked with glycerol and incubated for an additional 12 to 16 h. Cells weretheninfected with HSV-1, strain KOS, at a multiplicity
of infection of10PFU/cell. Attheindicatedtime postinfec-tion, monolayerswererinsedtwice withphosphate-buffered
saline and lysedby the addition of0.5 ml of0.6% sodium
dodecyl sulfate-10 mM EDTA-10 mM Tris hydrochloride (pH 7.5)-0.4 mg of proteinase K per mlat37°Cfor 10 to 12 h. Total cellular DNA was then isolated and quantified
spectrophotometrically. Approximately 5 ,ug of DNA (38) wasdigested withDpnI to removeunreplicated input DNA andwithEcoRItolinearize both test and reference plasmids. Thedigested DNAs werefractionated on a 1% agarose gel,
transferredto nitrocellulose, and hybridized to
32P-labeled,
nick-translatedpUC8. Southern blots were exposed to pre-flashed X-ray film, and the autoradiographs were scanned withan LKB scanning densitometer. Replication efficiency
is definedastheratio of the area under the peak of the test
[image:4.612.61.298.75.387.2]plasmid
totheareaunder thepeakof thereferenceplasmid,FIG. 5. Replication of pOS-822 deletion mutant plasmicts. tA)
The 822-bp BamHI fragment containing the intergenic region be-tween ICP4 and ICP22/47 is described in the legend to Fig. 2A. Beneath thediagram of the BamHIfragment are shown the limits of the deletionsin plasmids derivedfrom Bal-31 enzymedigestionat theunique BssHII site in the intergenic region of plasmidpOS-822
(Fig. 1B). The numbers in the plasmid designations indicate the number ofnucleotides deleted. (B) Southern blots of replicated deletionplasmid DNAs. Transient DNA replication assays of
mu-tant plasmids and of the replicating internal standard plasmid pBR325-ori-230 (arrow) were performed as described in thelegend toFig. 3B. (C)Replication efficiencies of deletionmutantplasmids were calculated as describedin the legendtoFig. 3C.
pBR325-ori-230. Although the reference plasmid pBR325-ori-230 carries the same oriS-containing fragment as
pOS-230, it replicates less efficiently than pOS-230 because it contains poison sequences that retard its replication in eukaryotic cells. The presence of these sequences did not affect theutility of thisplasmidasacontrol fortestplasmid replication. Theslight but significantcompetitionof the test plasmids with the reference plasmidfor replication factors
wastaken intoaccount in our calculations.
Toensurereproducibility of thetransient oriSreplication assays, all assays were performed three to five times. The results ofonerepresentativeassayareshown in each figure.
Althoughtheoverallefficiencyofplasmidreplication varied fromtest to test, the replication efficiencyoftest plasmids
relative to one another andto the internal control plasmid varied less than10% from assay toassay.
RESULTS
DNA sequences flanking oriSstimulatereplication of
oriS-containing plasmids. To determine whether replication of
oriS-containing plasmidswasaffected by the presence
oriS-" Lt) 10
m LI) - LO in
z z r- V
0 m m m m
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.322.555.76.375.2]A
DaIs Ds0 2C0 3 0C 400 500 6 Z.
60S0ON . ICP4
PT4' A a
1I - S < .:
up
217 bp 320bp
434bp
640bp
B.
C.
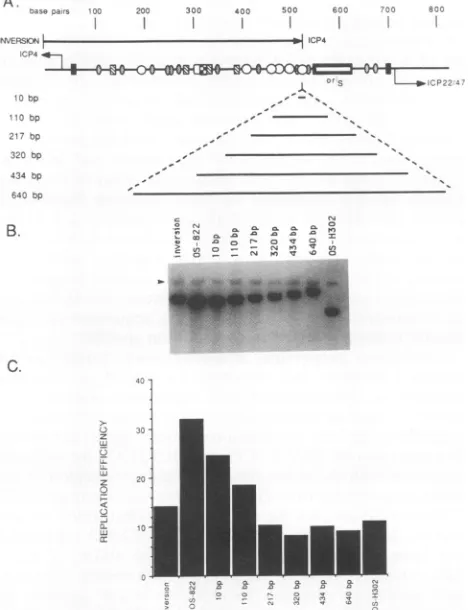
FIG. 6. Replication of pOS-822 insertion mutant plasmids. (A)
The intergenic region between ICP4 and ICP22/47 shown is
de-scribed in the legendtoFig.2A. Beneath thediagram ofthe BamHI
fragment are shown the DNA content of the mutant plasmids
derived from insertion of defined lengths of pBR322 DNAbetween
theICP4 promoter/enhancerand oriS. The numberadjacenttoeach
plasmid definesthe lengthofheterologous DNA inserted. A diagram
of the plasmid containing the HSV-1 immediate-early regulatory
element inthe inverted orientationis also shown. (B)Southemblots
of replicated insertion plasmid DNAs. Transient-replication assays
ofmutantplasmids andofthe replicating internal standard plasmid
pBR325-ori-230(arrows)wereperformedasdescribed in the legend
toFig.3B. (C)Replication efficiencies of each insertionmutantwere
calculatedas describedin the legend toFig. 3C.
flankingsequences, we cloned HSV-1 DNA fragments
con-tainingoriSand all (pOS-822)orportions(pOS-230, pOS-80)
of the intergenic region between the 5' transcriptional start sites of ICP4 and ICP22/47 into a common genetic
back-ground, pGEM7Zf(+) (Fig.1B,2B, and 3A). Vero cells were
cotransfected with each of the oriS-containing plasmids together with a reference plasmid, pBR325-ori-230, which
contains a 227-bp fragment of oriS. The reference plasmid wasincluded tocontrol for efficiency of transfection and to
serve as aninternal standardinthe calculation ofreplication
efficiencies. Although the referenceandtestplasmids would
be expected to compete for HSV-1 replication machinery, competition between the test and reference plasmids at the molarconcentrations of thetwoplasmids usedinthesetests
was minimal.
The results of replication assays with the three test
plas-midsare shownin Fig. 3BandC.Whenreplicationefficiency
wasplotted as afunction of time, the replicationrates of all
three plasmids were linear through 20 h postinfection (data
notshown). Of the three plasmids,pOS-822, which contains the entire intergenic region between thetranscriptional start sites of ICP4 and ICP22/47, replicated 16- to 22-fold more
efficiently than the reference plasmid at all three times tested, 8, 14, and 20 hpostinfection. (Note: plasmidpOS-822 migrates to approximately the same position as plasmids pOS-80 and pOS-230, due to the presence of an additional EcoRI site in the intergenic region that releases a 710-bp fragment from the vector.) PlasmidpOS-230,which contains the same 227-bp fragment as the reference plasmid,
repli-cated less efficiently than pOS-822 but four- to sevenfold more efficiently than the reference plasmid at all times tested. The level of replication of plasmid pOS-80, which contains only the coreorigin, aspreviously definedbyothers (11, 31),replicated fourfold lessefficiently than the reference plasmid through 20 hpostinfection. From theseresults, we
conclude that the presence of elements in theHSV-1 imme-diate-early transcriptional regulatory regions flanking oriS stimulate replication fromoriS by at least 80-fold relativeto
an oriS-containing plasmid (pOS-80) lackingthese elements. Because of the consistency and reproducibility of these tests, and because of thelinearity of the response overtime,
all subsequent tests were conducted at 8 h postinfection,
when the rate of viral DNA replication is high and when DNA synthesis is most synchronous in infected cells.
To further assess the role ofsequences to the left of the core origin in mediating the stimulatory effect on oriS,
systematic deletions were made in this region starting from
the ICP4 transcriptional start site (Fig. 4). Deletions were
made in these sequences because the great majority ofthe regulatory elements controlling the expression of both ICP4
and ICP22/47 are located between the core origin and the ICP4transcriptional start site (40, 47). Figure 4A shows the HSV DNAsequence content of the deletion mutants gener-ated, and the results oforiS replication assays areshownfor
each mutant in Fig. 4B and C. Progressive deletion of
sequences in the intergenic region beginning with the ICP4 transcriptional start site (pOS-H775) and the TATA element (pOS-H704) resulted in minorbutreproducible reductionsin replication efficiency, whereas plasmid pOS-H608, which
also lacks sequences downstream of the first TAATGARAT and CCAATbox-likebinding site nearest the ICP4
transcrip-tional start site (Fig. 2B), reproducibly replicated to nearly
wild-type levels. Deletion ofsequences from the ICP4
tran-scriptional start site through the second TAATGARAT element in plasmid pOS-H517 produced a minor but repro-ducible decrease in replication efficiency. Plasmids
pOS-H472 and pOS-H443, which lack more than half of the sequences between the core origin and the ICP4
transcrip-tional start site, replicated only half as efficiently as the wild-type plasmid. Notably, nearlywild-type levelsof
repli-cation were reproducibly demonstrated with plasmid pOS-H389, whichlacks allelements except 110bpimmediatelyto
the left of the core origin. This result was obtained in five independent tests. Plasmid pOS-H302, which retains only
the core origin, right-hand flanking sequences, and the
ICP22/47 start site, exhibited a fivefold reductionin
replica-tion efficiency relative to the wild-type plasmidpOS-822.
The results of these tests demonstrate the
stimulatory
effect of origin-flanking sequences on oriS function and indicate that the moreoriS-flanking sequenceswereincluded in the test plasmid, the more efficiently the plasmid
repli-cated. Thisobservation is contrary tothegenerally
accepted
axiom that the longer the replicating sequence, the less efficient is plasmid replication. Thus, specific elements in oriS-flanking sequences must have a stimulatory effect on
r,
, - 0
;; " - a -- -- -- pn
.
-0
0 r- o 'T o 7> "4 ?n 17 0
.:- 0 2 z ;. 'o v 'D o
4'
...W
,I I
I , I
I I,
I
I
I
I ,I
I I I
z
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.61.295.83.388.2]replication. The periodicity in replication efficiency ob-served withthesemutantsmaycorrespondwith thebinding
of individual transcription factors, differences in the
effi-ciency
oftranscription from theICP22/47 promoter, orthefactthat thedeletionmutations placesequencesnormallyat adistance from oriSadjacenttothe coreorigin.
Becauseplasmid pOS-H389 yielded nearly wild-type lev-els of origin function whereas pOS-H302 yielded greatly reduced levels, we wished to examine the sequences
be-tween positions 389 and 302 in greaterdetail. Progressive deletion of sequences from the convenient BssHII site towards the origin are represented by mutant plasmids pOS-Bd45, pOS-Bd75, and pOS-Bdll6 (Fig. 5A). This
re-gion containsthree copies ofthe sequence element CCCGT TGG and two copies ofa closely related sequence, CCCT
TGGG (Fig. 2). The former element, first described by Whitton and Clements (52), is conserved in the
intergenic
region between ICP4 and ICP22/47 in both HSV-1 and
HSV-2. This element resembles but is not identical to the
binding
siterecognized by CCAAT box transcription factors(5, 10, 18, 43). As shown in Fig. 5, deletion of a single CCCGTTGG element
(pOS-Bd45)
resulted inaslight
reduc-tion in oriSreplication
efficiency relative to pOS-822,whereas deletion of two elements (pOS-Bd75) decreased
replication efficiency by at least one-third. Deletion ofall three elements resultedin nearly 50% reductionin
replica-tion
efficiency
comparedwiththewild-type plasmid.
The deletion in plasmid pOS-Bdl55 removes some
se-quences in thecoreorigin, and therefore itreplicated
ineffi-ciently
as expected. Progressive deletion of CAAT-likeelements decreased replication
efficiency
significantly
butnot to the level shown in Fig. 4 (pOS-H302 versus pOS-H389). This differencecanonlybe duetothecontributionof sequences to the left ofthe BssHII restriction site. In the
experiments
shown inFig.
5,these sequenceswerebrought
progressively closer tothe core origin. The numerous
rec-ognized
cis-acting elements in these sequences may wellhave an enhancing effect on origin function as they are
brought
nearer the coreorigin.
Together, the resultspre-sented in
Fig.
4 and 5 suggest that CCCGTTGG sequenceelements aswellas sequencesto the leftofthe BssHII site maybe involved in the stimulation oforiS replication. To
determine the precise roles ofthese elements in oriS
func-tion,
substitution mutations inthese elements arecurrently
being
generated.Distance between the core origin and left-hand flanking sequences affects the efficiency oforiS function. In order to
evaluate the contribution of distance between oriS and left-hand flanking sequences on the efficiency of oriS
func-tion,
heterologous DNA from plasmid pBR322 wasintro-duced into the wild-type plasmid (pOS-822) atposition 302
(plasmid pOS-H302, Fig. 4A). Mutant plasmids containing insertsof10, 110, 217, 320, 434, and 640 bp ofheterologous
DNA were assayed for oriS replication efficiency (Fig. 6).
Although the insertion of 10 bp had only a minor effect on
oriS function, insertion of 110 bp resulted in a greater reduction inoriSfunction.The efficienciesof originfunction ofplasmids containing heterologousinsertsranging from 217
to 640bp were approximately equal, exhibiting only
-30%
of
wild-type
plasmid replication efficiency. These resultsshow that theinsertionof sequences of217bporgreaterhad
equally deleterious effects on oriS function. There was no
differencein thereplicationefficienciesof pOS-H302(which lacksallleft-handflanking sequences)andplasmids
contain-ing
insertionsrangingfrom 217to640bp,indicating
thatatadistance of at least 217 bp, the stimulatory effect of the flanking sequences onoriSfunctionis lost.
Thereduction inoriginfunctionobservedfollowing inser-tion of nonspecific DNA between the core origin and the
left-hand flanking sequences demonstrates that the distance between the left-hand transcriptional regulatory regionand the coreoriginaffects oriSfunctionsignificantly.Inaddition toillustrating the effects on oriSfunction of the insertion of
heterologous DNA between the core origin and left-hand flanking sequences, Fig. 6illustrates the effect ofinversion ofthese flanking sequences on originfunction. Relative to
plasmid pOS-822, in which flanking sequences are in the wild-type orientation, pOS-822(inv) exhibited a 50% reduc-tion in origin function. This result, together with those obtained with the insertion mutants, further supports a
stimulatoryrole fororiS-flankingsequences onoriSfunction anddemonstrate that the effect of these sequences onorigin function is both distance and orientation specific.
Heterologous herpesvirus immediate-early transcriptional regulatory elements can substitute for HSV-1 oriS-flanking sequences in stimulation of oriS replication efficiency. To determine whetherstimulation of DNA replication from oriS is specific to the transcriptionalregulatory region controlling the expression of HSV-1 ICP4 and ICP22/47, we replaced this region with the transcriptional regulatory region of the humancytomegalovirus (HCMV)immediate-early gene IEl (3).Figure 7 illustrates the effects ofthis substitution on oriS function. Because not allHCMV originsofDNA replication have been mapped, we first assayed the ability of
pUC-CMV, which contains the HCMV immediate-early transcrip-tional regulatory element, to replicate in our system. As shown inFig.7BandC, plasmid pUC-CMV didnot replicate in this system, indicating that it contains no origin that is responsive to superinfection with HSV-1. By contrast,
plas-mid pOS-CMV4, which contains the HCMV immediate-early regulatory element in the same orientation as the
HSV-1 immediate-early regulatoryelement inplasmid
pOS-822, replicatedto 90% ofwild-type (pOS-822) levels.
Inver-sion oftheHCMVregulatoryelement(pOS-CMV3)resulted in only a minor change in the efficiency of oriS function relative to pOS-CMV4. Notably, bothpOS-CMV3and
pOS-CMV4 exhibited significantlygreaterreplicationefficiencies thandidpOS-H302, which lacksnearlyallleft-handflanking sequences. Thus, the HCMV immediate-early regulatory
element can substitute for theHSV-1 immediate-early regu-latory element in the enhancement of oriS replication
effi-ciency. Whether the tworegulatory elements stimulate oriS function bythe same ordifferent mechanisms is notknown;
however, the HCMV immediate-early regulatory element,
like its HSV-1 counterpart, contains numerousbindingsites forrecognized cellular transcription factors. If indeed
tran-scriptionalcomplexes playarole instimulation of oriS,it is not surprising that the HCMV element can substitute
func-tionally for the HSV-1 element in origin replication tests.
Unlike the HSV-1 element, however, the HCMV element was capable of enhancing replication in either orientation.
This suggests the possibility that sequences critical for
stimulationof oriSactivity are evenlydistributed within the HCMV immediate-early regulatory element, whereas those inthe HSV-1 element are not.
Factors thatbindto
oriS-flanking
sequencesenhanceorigin function. Theresults of the studies described above indicate that theefficiency oforiS-containingplasmids is affectedby transcriptional regulatory elements adjacent to the coreorigin. Additionally,thefact that substitution ofthiselement
by a heterologous herpesvirus regulatory element, the
on November 10, 2019 by guest
http://jvi.asm.org/
basspaws 100 200 300 400 500 600
i:CP4*,ImI
OM_
g_~~~~~~~~~~~~~~~~1
o Cn M
orls ,^;P22
4-Os-822
oOS0-H302
,OS-CMAV3 = 3CSCSMV4 =
E N D
>-r> N
-1 V) U) Xn c
OL O O O
wm
C.
z CU
LUL
z
0
Q
C-C'
CC
10
8
6
4.
2
0'
I
[image:7.612.160.460.81.429.2]pUC CMV OS822 OS-H302 OS-CMV3 OS-CMV4
FIG. 7. Replication of plasmids containing the HCMV immediate-early promoter. (A) The 822-bp BamHI fragment containing the intergenic region between ICP4 andICP22/47 is described in the legendtoFig. 2A. Beneath the diagram of the BamHI fragmentarediagrams ofpOS-822, pOS-H302, and the plasmids containing the HCMV immediate-earlypromoter(pUC-CMV, pOS-CMV3, andpOS-CMV4). (B) Southern blots of replicated insertionplasmid DNAs. Transient-replicationassaysofmutantplasmids and of the replicating internal standard plasmid pBR325-ori-230 (arrows) were performed as described in the legend to Fig. 3B. (C) Replication efficiencies ofplasmids were calculatedasdescribed in thelegendtoFig. 3C.
HCMV IElregulatory element, resulted innearly wild-type
levels of oriS function demonstrates that a homologous
regulatory element is not required for efficient plasmid replication. Rather, these results suggest thatprotein-DNA complexes associated with origin-flanking sequences likely
affectoriginfunction.
To examine the role that trans-acting factors playinoriS function,weselectively deleted thecoreorigin from plasmid
pOS-822 to yield plasmid poriS(-), which lacks oriS func-tion yet retains nearly all cis-acting regulatory elements flankingoriS (Fig. 8A). Thisplasmid was used to compete
with plasmid pOS-822 for factors that bind to flanking
sequences in transient DNAreplication assays. Figures 8B and C illustrate the results of such tests when the plasmid poriS(-) wascotransfected in 25-, 50-, and 100-fold molar excess relative to the test plasmid, pOS-822. The internal control plasmid pBR325-ori-230 was not included in this
experiment because cis-acting elements in this plasmid would alsocompetewithpOS-822 for specific DNA-binding
factors. Toensure sequence specificity of the HSV-1 DNA in the competitor plasmid poriS(-), vector DNA was in-cluded in each transfection.
Infourindependent experiments, the addition ofa25-fold molarexcessof thecompetitor plasmidreducedreplication of plasmid pOS-822 to approximately 30% of the level observed withoutcompetitor,and addition of thecompetitor to a 100-fold molar excess reduced replication of the test plasmid to approximately 10% of the control level. These results demonstrate that the binding oftrans-acting factors
tocis-actingelements within theintergenic regulatory region
is essential for maximum replication of an HSV-1
oriS-containingplasmid.
DISCUSSION
HSV-1 oriS: theorigin functiontest. Byanalogy toSV40, adenovirus, andotherDNA-containing viruses, the proxim-ity ofHSV-1 oriSto avariety oftranscriptional regulatory complexes containingboth viral and cellular proteins
sug-geststhepossibilitythatcomplexesaffectoriginfunction. In
orderto test this possibility, we measured the effects ofa
varietyof mutations withinsequences flankingoriSon oriS replication efficiency.Toensureanidenticalgeneticcontext
for theseanalyses, origin-containingHSV-1 DNAfragments
A.
B.
on November 10, 2019 by guest
http://jvi.asm.org/
A.
base pa,rs 100 200 300 400 500 600 0 -D 8000 ICP441
IO ,.P22 4
Ol --- "P224.
B.
C.
.J EC
z ICL
401
4j]
[image:8.612.63.296.77.380.2]FO -,.ri-MPrIORtmoa,!1
FIG. 8. Effect ofcompetition oforiS-flanking sequences onthe replication of theoriS-containing plasmidpOS-822. (A) The 822-bp
BamHIfragmentcontainingtheintergenic region between ICP4and
ICP22/47 is shown in Fig. 2A. Also shown is the wild-typeHSV-1 oriS plasmid pOS-822. The oriS deletion mutant poriS(-) was constructed by partial digestion of pOS-822 DNA with SmaI,
followedby religation.(B) Transient-replicationassayofpOS-822in
the presence of increasing molar concentrations of competitor plasmid poriS(-). To exclude the possible effects of vector
se-quencesinthecompetition,the totalamountofvectorDNA in each transfectionwasthesame. Assayswereperformedas described in thelegendtoFig. 3B,and cellswereharvestedat8 hpostinfection. The replicating internal standard plasmid pBR325-ori-230 was not
included in these assays. (C) Replication efficiency of plasmid pOS-822 in thepresence of different concentrations ofcompetitor plasmid. Percentageswerecalculatedby scanningdensitometryand comparisonof thearea under thepeak produced by each
concen-trationofcompetitortothe controlpeakwithout thecompetitor.
bearingmutationsadjacenttothecoreoriginweresubcloned into the sameplasmid vector. Toovercome inherent varia-tions in transient DNAreplicationassaysthatmight compli-cate accurate quantitation ofmutant plasmid replication, a
replicating internal control plasmid consisting of oriS se-quencesinadifferent plasmidvectorwasincluded fordirect comparisoninalltransfections.
Our data clearly indicate that the existence ofcis-acting
elements adjacent to oriS affects the overall efficiency of origin-dependent DNA replication. Moreover replication efficiency was shown to be affected by the protein factors which bindtothese cis-acting elements.Theserequirements
forreplicationfrom HSV-1oriSareconsistent with those of otherDNA-containing animal viruses, such as SV40,
poly-omavirus,bovine papillomavirus, adenovirus, and
Epstein-Barr virus(8, 15, 17, 23, 24, 26, 27, 32, 34, 36, 44, 48, 53).
Basedon ourknowledge of originfunction in SV40 and other DNA-containing viruses and on the demonstration herein that HSV-1origin-flanking sequences stimulate oriS function in transient assays, one must ask whether oriS-flanking sequences affect origin function in the context of the viral genome and, if so, how these sequences contribute to the underlying mechanism of initiation of HSV-1 DNA replica-tion. Efforts are under way to transfer selected mutations into the HSV-1 genome. Given that oriS is both diploid and essential for virus replication, construction of the desired viral mutants is not straightforward. Preliminary evidence, however, indicates that replication-competent mutant vi-ruses with mutations in oriS-flanking sequences can be isolated.
Inthe absence of such mutant viruses, our studies were conducted with an approach used previously to define the limits of HSV-1 oriS. Thus, Stow et al. demonstrated by deletion analysis of the intergenic region between ICP4 and ICP22/47 that the minimal origin was contained within a
90-bp DNA fragment (49). Although these authors did not observe the stimulatory effect of flanking sequences on oriS function, they postulated that thismight in fact be the case. One may well ask why we observed the stimulatory effect whereas Stow et al. (49) did not. Aside fromdifferences in the cell types used in the two studies, three possible
expla-nations come tomind. First, in the present study but not in the earlier study, an internal replicating plasmid was in-cluded to aid in the precise measurement of replication
efficiency and to serve as a control for transfection
effi-ciency. Second, this study used aquantitative approach to
measuring originfunction, whereas the study of Stow etal. used a qualitative approach to identify the minimal origin
(49). Lastly,differences in assay procedures may account for this discrepancy. Inthe study of Stow etal., DNA
replica-tionwas assessed at 18 hpostinfection,whereas inthisstudy
replication was assessed at 8 h postinfection. It is possible that the replication rate of pOS-822 is reduced by 18 h postinfection and/or that plasmids that replicate less
effi-ciently eventually attain the same level of replication as
efficiently replicating plasmids at 18 h. Differences in the
replication rates of SV40 origin-containing plasmids as a
function of time postinfection have been observed (21). In
our hands, however, the replication rates of pOS-822 and
pOS-230 werelinearthrough 20 hpostinfection.
The differences in the results obtained in this study and thatofStow et al. may also reflect differences in the relative timesof transfection and infection. If thiswerethecase, the
discrepancy in the outcomes could be explained by nucleo-some exclusion. Previous studies of SV40 have demon-strated that once in the nucleus, transfectedDNA is associ-ated with nucleosomes asit is assembledinto chromatin(6,
27). However, specific regions of transfected DNA were found to be devoid of nucleosomes. These regions
corre-sponded to the SV40 promoter/regulatory elements and
originof DNAreplication.Whentransferredtoothersites in theSV40 genome, theseelements remainednucleosome-free
atthese sites (27). In their study, Stow et al. infected baby
hamsterkidney(BHK) cells4hfollowing dimethyl sulfoxide
(DMSO) shock, whereas in this study we infected Vero cells 16 h afterglycerol shock. At 4h post-DMSO shock,
trans-fected plasmid DNA may still be devoid ofnucleosomes,a
situation in which noorigin-containing plasmidwould beat a
competitiveadvantageoverany otherorigin-containing
plas-mid. By contrast, at 16 h posttransfection, association of specific plasmid sequences with nucleosomes could have
far-reaching effects on the replication efficiency of
origin-x x x
004 Ln
on November 10, 2019 by guest
http://jvi.asm.org/
containing plasmids. In order to evaluate differences in the time between transfection and infection asa possible expla-nation for the disparity in the two studies, Vero cells were transfected with plasmids pOS-822 and pOS-230. At 4 h post-glycerol shock, monolayers were infected, and 8 h later the DNA from the infected cells was harvested. This proce-dure was identical to that of Stow et al. except that Vero cells instead of BHK cells and glycerol instead ofDMSO were used. The results of this test were consistent with the findings reported above, that flanking sequences increased the efficiency of DNA replication by -20-fold (unpublished results). At present, therefore, the reasons for the discrep-ancy between the findings reported here and those of Stow et al. remain unclear.
How dooriS-flanking sequences affect origin function? The results presented in this report demonstrate that, like other DNA-containing viruses, the promoter-regulatory elements adjacent to the core origin act to increase the overall replication efficiency of oriS-containing plasmids. In addi-tion to the CCCGTTGG elements immediately adjacent to the core origin, other sequences far upstream of these elements also have stimulatory effects on origin function. Consequently, a multiplicity of elements is likely involved in the enhancement process, and multiple elements may well work together to provide the wild-type level of replication.
The precise roles of protein-DNA complexes adjacent to origins of replication have not been determined; however, three possible functions for these complexes have been proposed. One is that they prevent nucleosome formation and hence enhance access of DNAreplication proteins to the origin. Another is that complexes involved in transcription facilitate melting of the DNA duplex at the origin. The third proposes that proteins involved with transcription interact with replication proteins. Whether the proteins involved in these complexes can rightly be considered associated with transcription or DNA replication or both will be difficult to determine.
Do transcription initiation complexes enhance accessibility of DNA replication proteins to oriS? The trans-acting factors that bind to the cis-acting regulatory elements may serve specifically to makeoriS more accessible to proteins of the initiation complex. As mentioned above, nucleosome-free regions on the SV40 genome occur where promoters, en-hancers, and the origin of DNA replication are found. A similar mechanism may also be operative in the transient assays reported herein. Thebinding oftrans-acting factors to the transcriptional regulatory region surrounding HSV-1 oriS may affect the distribution of nucleosomes, thereby increasing the accessibility of theorigin to viral and cellular initiation factors. Although this remains a distinct possibil-ity, other mechanisms may also be operative in oriS activa-tion. Like the numerouscellulartrans-acting factors that can bind to the transcriptional regulatory element, the TATA box-binding protein TFIID can prevent nucleosome-medi-ated repression of a promoter (54). If inhibition of nucleo-some-mediated repression by TFIID were the sole mecha-nism involved in the activation of HSV-1 oriS in these studies, then inversion of the ICP4 andICP22/47 transcrip-tional regulatory element, whichplaces the ICP4 TATA box adjacent to the core origin, should have resulted in substan-tially higher levels oforiS activity (Fig. 6B and C) (54). As shown in Fig. 6, inversion of the transcriptional regulatory region resulted in areplication efficiencyapproaching that of pOS-H302, which lacks the ICP4 and ICP22/47 transcrip-tional regulatory region. Thisresult suggests that inhibition of nucleosome-mediated repression may not be the sole
mechanismresponsible for the stimulation of replication at
HSV-1oriS observedinthis study.
Doestranscriptionexert asecondary effectonorigin
config-uration? With regard to the putative role of flanking se-quencesin theinitiationofDNAsynthesis, onehypothesis is that they act primarily to regulate transcription of the
adjacent immediate-early genes and that they exert a
sec-ondary effect on oriS function because oftheir proximity.
Theimmediate-earlygenesspecifyingICP4andICP22/47are
transcribed shortlyafter infection (13). ICP4 represses
tran-scription of immediate-early genes and activates
transcrip-tionofearlyand late genes
(13).
Thus,theICP4proteinitself is thought to downregulate transcription from the promotersof ICP4 and ICP22/47 by binding to sequences at the transcriptional start sites of these genes (13). Although transcription from the ICP4 and ICP22/47 promoters is
substantially reduced when viral DNA replication begins at
oriS,this region of the genome is in a transcriptionally active conformation, since transcription from these promoters can
still be detected. This low levelof transcription couldmodify
the localconfiguration oftheDNA,facilitatingmeltingof the duplex at oriS. The enhancement of origin function by
activation of transcription from origin-flanking sequences has been shown to be required in vivo for replication of bacteriophage X DNA (19) and has been observed during
initiation of replication of the E. coli chromosomal origin,
oriC (2).
Do proteins involved in transcription and DNA replication
interact? If a relationship exists between oriS-dependent
DNA replication and transcription of flanking genes, it is
possiblethat theproteins which bindto andregulatespecific
promoters and enhancers interact directly with DNA repli-cation proteins. Studies of both SV40 and adenovirus sug-gest that such a direct interaction may well assistinitiation from the core origins of these two viruses (21, 22, 45, 53). Such interactions may assist in promoting localized strand separation, as observed for SV40 (21, 22). In HSV-1, the proximity of the CCCGTTGG sequence elements to oriS suggests that the trans-actingfactor(s) that forms a complex with this element may function in a manneranalogous to the GC boxes ofSV40 to stimulate oriS function. Consistent with this hypothesis is the observation that a protein(s) present in mock-infected cells binds to this element (unpub-lished results). This hypothesis is not without support, since comparison of the replication efficiencies of plasmids pOS-H389 and pOS-H302 (Fig. 4B and C) demonstrates that the presenceof these elements resultsin a significant stimulation oforiS function.
Evidence to support the involvement of the CCAAT box-like transcription factors was presented in Fig. 7. The HCMV immediate-early regulatory element, which contains numerous binding sites for NF-I, can substitute in either orientation for the ICP4 and ICP22/47 transcriptional regu-latory region. The uniform distribution ofNF-I-binding sites and CCAAT box sequences within the HCMV immediate-early regulatory element (20, 35) may explain why this element can stimulateoriS function in the inverted orienta-tion but the HSV-1 immediate-early regulatory element cannot. As shown in Fig. 2A and B, the majority of the CCAAT box-like elements within the ICP4 and ICP22/47 transcriptional regulatory region are located adjacent tothe core origin. By inverting the ICP4 and ICP22/47 transcrip-tional regulatory region, the closest CCAAT box-like ele-ment is located approximately 200 bp from the origin, and the cluster of these elements (normally adjacentto the core origin) is located at a distance ofapproximately 400 bp. At
on November 10, 2019 by guest
http://jvi.asm.org/
these distances, thestimulatory effects of the CCCGTTGG elements may no longer be effective(Fig. 6).
The potential involvement of CCAAT box transcription
factor(s)in HSV-1oriS function is intriguing. Nuclear factor I, amember of the CCAAT box transcription factor family (46), is essential for adenovirus DNA replication (45, 46).
Althoughthe precisemechanism of activation of adenovirus DNA replication is not known, the binding of NF-I to its
recognition site is required (45). Furthermore,
structure-functionanalysis ofNF-Ihas shownthata domainrequired
for DNA binding distinct from the domain involved in transcription is absolutely required for adenovirus DNA replication (33).
Does transcription across oriS affect oriS function? An additional mechanism to explain how oriS may be activated inthe viral genomehas recently been proposed. Hubenthal-Voss and Roizman (25) have identified transcripts which traverse oriS sequences in both HSV-1 and HSV-2. It has been postulated that these transcripts function to regulate DNA replication by inducing changes in the superhelical density of the template or by serving as primers for DNA replication. Although this hypothesis cannot be discounted
as a potential mechanism for enhancing DNA replication from oriS in the context of the viral genome, it does not explain the observations reported in this article because the presumed transcriptional start sites for the oriStranscripts
are not present in the plasmids used in this study.
The possible association between transcription ofadjacent
viralgenes and oriS function clearly warrants further inves-tigation. The use of viral mutants should prove especially valuable for this purpose. For example, in one study of mutations in this region of the HSV-2 genome, Smith et al. (47) reported the constructionof viable HSV-2 mutantswith deletions inthe intergenic region between ICP4 andICP22/
47. These mutants induced the synthesis of substantially reducedlevels ofICP4, ICP22, andICP47 mRNA and were growthimpairedrelative to thewild-typevirus. Whether the growth impairment was a consequence of the reduced
tran-scription of ICP4andICP22/47orofreduced oriS function or both was not determined. The introduction of selected
mutations generated in this study into the HSV-1 genome together with the simultaneous assessment oftranscription ofICP4,ICP22, andICP47and theefficiency oforiSfunction
mayprovide a means of addressing this question.
ACKNOWLEDGMENTS
Wethank Neal DeLucaandChristine E. Dabrowski forhelpful discussions and Meg Kaveny and Marybeth Pilat for manuscript preparation.
Thisinvestigation wassupportedby Public Health Servicegrants R01AI28537from the National Institute ofAllergy and Infectious Diseases and R37CA20260 from the National Cancer Institute. S.W.W. has been supported by Public Health Service grant 5T32CA09031 and a postdoctoral fellowship (PF-3403) from the AmericanCancerSociety.
REFERENCES
1. apRhys, C. M. J., D. M. Ciufo, E. A. O'Neill, T. J. Kelly, and G. S.Hayward. 1989. Overlapping octamer and TAATGARAT
motifs inthe VF65-response elements in herpes simplexvirus
immediate-earlypromoters represent independent binding sites
for cellular nuclear factorIII. J.Virol. 63:2798-2812.
2. Baker, T. A., and A. Kornberg. 1988. Transcriptional activation ofinitiation ofreplication from the E.colichromosomalorigin:
anRNA-DNA hybrid near oriC.Cell 55:113-123.
3. Boshart, M., F. Weber, G. Jahn, K. Dorsch-Hasler, B.
Flecken-stein,andW.Schaffner. 1985. A verystrong enhancer is located
upstreamofanimmediate earlygene ofhuman cytomegalovi-rus.Cell41:521-530.
4. Bzik, D. J., and C. M. Preston. 1986. Analysis of DNA se-quences which regulate thetranscriptionofherpes simplexvirus immediate early gene 3: DNA sequencesrequired for enhancer-like activity andresponse to trans-activation by a virion poly-peptide. Nucleic Acids Res. 14:929-943.
5. Celada, A., and R. A. Maki. 1989. DNAbindingof themouse class IImajorhistocompatibilityCCAAT factordependsontwo components. Mol. Cell. Biol. 9:3097-3100.
6. Cereghini, S., and M. Yaniv. 1984. Assembly of transfected DNA into chromatin: structuralchanges in the origin-promoter-enhancerregionuponreplication. EMBO J. 3:1243-1253. 7. Challberg, M. D., and T. J. Kelly. 1989. Animal virus DNA
replication. Annu. Rev. Biochem. 58:671-717.
8. Chandrasekharappa, S. C., and K. N. Subramanian. 1987. Effects ofposition and orientation of the 72-base-pair-repeat transcriptionalenhancer onreplicationfrom the simian virus 40 coreorigin. J.Virol. 61:2973-2980.
9. Cheng, L., and T. J. Kelly. 1989. Transcriptional activator nuclear factor I stimulates thereplicationofSV40 minichromo-somesinvivo andinvitro. Cell59:541-551.
10. Chodosh, L. A., A. S.Baldwin,R. W.Carthew,and P.A.Sharp. 1988.HumanCCAAT-binding proteins have heterologous
sub-units. Cell53:11-24.
11. Deb, S., and M. Doelberg. 1988. A67-base-pairsegmentfrom theOri-Sregionofherpes simplex virustype 1encodesorigin function. J. Virol.62:2516-2519.
12. Deb, S., and S. P. Deb. 1989. Analysis of Ori-S sequence of
HSV-1: identification of one functional DNA bindingdomain.
NucleicAcids Res. 17:2733-2752.
13. DeLuca, N. A., and P. A. Schaffer.1985. Activation of immedi-ate-early, early, and late promoters by temperature-sensitive
and wild-type forms of herpes simplex virus type 1 protein
ICP4.Mol. Cell. Biol. 5:1997-2008.
14. DeLuca, N.A.,and P. A.Schaffer.1988.Physicalandfunctional
domains of the herpes simplex virustranscriptional regulatory proteinICP4. J. Virol. 62:732-743.
15. DeLucia, A. L., S. Deb, K. Partin, and P. Tegtmeyer. 1986.
Functional interactions ofthe simian virus 40 core origin of
replicationwithflanking regulatorysequences. J. Virol. 57:138-144.
16. DePamphilis,M. L. 1988. Transcriptional elementsas compo-nentsofeukaryotic originsof DNAreplication.Cell 52:635-638. 17. de Villiers, J., W. Schaffner, C. Tyndall, S. Lupton, and R. Kamen. 1984. Polyoma virus DNA replication requires an
enhancer. Nature(London) 312:242-246.
18. Dorn, A., J.Bollekens, A. Staub, C. Benoist, and D. Mathis. 1987. A multiplicity of CCAAT box-binding proteins. Cell
50:863-872.
19. Furth, M. E., W. F. Doves, and B.J. Meyer. 1982. Specificity
determinants forbacteriophageA DNAreplication. III. Activa-tion ofreplicationin Aricmutantsbytranscriptionoutsideof ori. J. Mol. Biol. 154:65-83.
20. Ghazal, P., H. Lubon, B. Fleckenstein, and L. Hennighausen.
1987. Binding oftranscription factors and creation ofa large
nucleoprotein complex on the human cytomegalovirus
en-hancer.Proc. Natl. Acad. Sci.USA 84:3658-3662.
21. Guo, Z.-S., C. Gutierrez, U. Heine, J. M. Sogo, and M. L.
DePamphilis. 1989. Origin auxiliary sequences can facilitate initiation ofsimian virus 40 DNAreplication in vitroastheydo
in vivo. Mol. Cell. Biol. 9:3593-3602.
22. Gutierrez, C., Z.-S. Guo, J. Roberts, and M. L. DePamphilis.
1990. Simianvirus 40originauxiliaryfacilitates DNA unwind-ing.Mol. Cell. Biol. 10:1719-1728.
23. Hammerschmidt, W., and B. Sugden. 1988. Identification and
characterization oforilyt, alytic origin ofDNA replicationof
Epstein-Barrvirus. Cell55:427-433.
24. Hertz, G. Z., and J. E. Mertz. 1986. Bidirectional promoter elements of simian virus 40arerequiredfor efficientreplication of the viral DNA. Mol. Cell. Biol. 6:3513-3522.
25. Hubenthal Voss, J., and B. Roizman. 1988. Properties oftwo
5'-coterminal RNAs transcribed part way and across the S
on November 10, 2019 by guest
http://jvi.asm.org/
componentorigin of DNA synthesis of the herpes simplex virus
1 genome. Proc.Natl. Acad.Sci. USA85:8454-8458.
26. Innis, J. W., and W. A. Scott. 1984. DNA replication and
chromatin structureof simian virus40insertionmutants. Mol.
Cell. Biol.4:1499-1507.
27. Jongstra, J., T. L. Reudelhuber, P. Oudet, C. Benoist, C.-B. Chae, J.-M. Jeltsch, D. J. Mathis, and P. Chambon. 1984.
Induction of altered chromatin structures by simian virus 40
enhancer and promoter elements. Nature (London) 307:708-714.
28. Lee-Chen, G.-J., and M. Woodworth-Gutai. 1986. Simianvirus 40 DNA replication: functional organization of regulatory
ele-ments. Mol. Cell. Biol.6:3086-3093.
29. Leinbach, S. S., and W. C. Summers. 1980. The structure of herpes simplex virus type 1 DNA as probed by micrococcal nuclease digestion. J. Gen. Virol. 51:45-59.
30. Li, J. J., K. W. C. Peden, R. A. F. Dixon, and T. Kelly. 1986.
Functional organization ofthe simian virus 40 origin of DNA
replication. Mol. Cell. Biol.6:1117-1128.
31. Lockshon, D., and D. A. Galloway. 1988. Sequence and struc-tural requirements of a herpes simplex viral DNA replication
origin. Mol. Cell. Biol.8:4018-4027.
32. Lusky, M., and M. R. Botchan. 1986. Transient replication of
bovinepapillomavirustype 1 plasmids: cisand trans
require-ments. Proc. Natl.Acad. Sci. USA83:3609-3613.
33. Mermod, N., E. A. O'Neill, T. J. Kelly, and R.Tjian. 1989. The proline-rich transcriptional activator ofCTF/NF-I is distinct
fromthereplicationand DNAbinding domain. Cell58:741-753. 34. Muller, W. J., D. Dufort, and J. A. Hassell. 1988. Multiple
subelements within the polyomavirusenhancerfunction
syner-gisticallytoactivateDNAreplication. Mol. Cell. Biol.
8:5000-5015.
35. Nelson, J. A., C. Reynolds-Kohler, and B. A. Smith. 1987.
Negative and positive regulation by a short segment in the
5'-flanking region ofthe humancytomegalovirusmajor
immedi-ate-earlygene. Mol.Cell. Biol. 7:4125-4129.
36. O'Connor, D. T., and S. Subramani. 1988. Do transcriptional enhancers alsoaugment DNAreplication? Nucleic Acids Res.
16:11207-11222.
37. Olivo,P.D., N. J. Nelson, and M. D. Challberg. 1988. Herpes
simplex virus DNA replication: the UL9 gene encodes an
origin-binding protein. Proc. Natl. Acad. Sci. USA
85:5414-5418.
38. Peden, K. W.C., J. M. Pipas, S. Pearson-White, and D. Nathans. 1980.Isolation ofmutantsofananimal virusinbacteria. Science
209:1392-1396.
39. Polvino-Bodnar, M., P. K. Orberg, and P. A. Schaffer. 1987.
Herpes simplex virus type 1 oriL is not required for virus
replication or for the establishment ofreactivation of latent
infectioninmice.J. Virol. 61:3528-3535.
40. Preston, C. M., M. G. Cordingley, and N. D. Stow. 1984.
Analysis ofDNA sequenceswhichregulate the transcription of
aherpes simplex virus immediate-earlygene. J. Virol. 50:708-716.
41. Preston, C. M., and D. Tannahill. 1984. Effects of orientation
andpositionontheactivity ofaherpes simplex virus immediate earlygenefar-upstream region. Virology 137:439-444.
42. Preston, C. M., M. C. Frame, and M. E. M. Campbell. 1988. A
complexformed between cellcomponents andan HSV struc-turalpolypeptide bindsto aviral immediateearly gene regula-tory DNA sequence.Cell52:425-434.
43. Raymondjean, M., S. Cereghini, and M. Yaniv. 1988. Several
distinct "CCAAT" boxbinding proteins coexist ineukaryotic cells. Proc. Natl. Acad. Sci. USA85:757-761.
44. Reisman, D., J. Yates, and B.Sugden.1985.Aputativeorigin of replication of plasmids derived from Epstein-Barr virus is composed of two cis-acting components. Mol. Cell. Biol.
5:1822-1832.
45. Rosenfeld, P. J., E. A. O'Neill, R. J. Wides, and T. J. Kelly. 1987. Sequence-specific interactions between cellular
DNA-binding proteins and the adenovirus origin ofDNAreplication. Mol. Cell. Biol. 7:875-886.
46. Santoro, C., N. Mermod, P. C. Andrews, and R. Tjian.1988.A
family ofhuman CCAAT-box-binding proteins active in
tran-scriptionand DNAreplication: cloning and expression of mul-tiple cDNAs. Nature(London) 334:218-224.
47. Smith, C. A., M. E. Marchetti, P. Edmonson, and P. A. Schaffer. 1989. Herpessimplexvirus type 2 mutants withdeletionsin the
intergenic region between ICP4andICP22/47: identification of nonessential cis-acting elements in the context of the viral
genome. J. Virol.63:2036-2047.
48. Stenlund, A., G. L. Bream, and M. R. Botchan. 1987. A promoterwithaninternal regulatory domain ispartoftheorigin
ofreplication inBPV-1. Science236:1666-1671.
49. Stow, N. D., and E. C. McMonagle. 1983. Characterization of theTRS/IRSorigin ofDNAreplication of herpes simplex virus
type 1.Virology130:427-438.
50. Triezenberg, S.J.,K. L. LaMarco, and S. L. McKnight. 1988.
Evidence of DNA:protein interactions that mediate HSV-1 immediate early gene activation byVP16. Genes Dev.
2:730-742.
51. Weller, S. K.,A.Spadaro,J. E.Schaffer,A. W.Murray, A. M. Maxam, and P. A. Schaffer. 1985. Cloning, sequencing, and
functionalanalysis of oriL,aherpes simplex virustype 1origin ofDNAsynthesis. Mol. Cell. Biol. 5:930-942.
52. Whitton,J.L., andJ.B.Clements. 1984.Replicationorigins and
asequence involved incoordinate induction oftheimmediate earlygenefamilyareconserved inanintergenicregion of herpes simplex virus. Nucleic AcidsRes. 12:2061-2079.
53. Wides, R. J., M. D.Challberg,D. R.Rawlins,and T.J. Kelly.
1987.Adenovirusorigin ofDNAreplication:sequence
require-mentsforreplicationin vitro. Mol. Cell. Biol. 7:864-874.
54. Workman, J. L., and R. G. Roeder. 1987.Binding of transcrip-tion factor TFIID to the major late promoterduring in vitro nucleosomeassembly potentiates subsequent initiationbyRNA
polymerase II.Cell51:613-622.
55. Wu,C.A.,N.J. Nelson,D.J.McGeoch,and M. D. Challberg.
1988. Identification ofherpes simplex virus type 1 genes
re-quired for origin-dependentDNA synthesis. J. Virol. 62:435-443.
56. Wysokenski, D. A., and J. L. Yates. 1989. Multiple EBNA1-binding sites are required to form an EBNA1-dependent en-hancer andtoactivateaminimalreplicative originwithin oriPof Epstein-Barr virus. J.Virol. 63:2657-2666.