0022-538X/09/$08.00⫹0 doi:10.1128/JVI.01185-09
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Analysis of Human Alphaherpesvirus MicroRNA Expression in
Latently Infected Human Trigeminal Ganglia
䌤
Jennifer L. Umbach,
1Maria A. Nagel,
2Randall J. Cohrs,
2Donald H. Gilden,
2,3and Bryan R. Cullen
1*
Department of Molecular Genetics and Microbiology and Center for Virology, Duke University Medical Center, Box 3025, Durham,
North Carolina 27710,1and Departments of Neurology2and Microbiology,3University of Colorado—Denver,
School of Medicine, 12700 East 19th Ave., Aurora, Colorado 80045
Received 9 June 2009/Accepted 26 July 2009
Analysis of cells infected by a wide range of herpesviruses has identified numerous virally encoded micro-RNAs (mimicro-RNAs), and several reports suggest that these viral mimicro-RNAs are likely to play key roles in several aspects of the herpesvirus life cycle. Here we report the first analysis of human ganglia for the presence of virally encoded miRNAs. Deep sequencing of human trigeminal ganglia latently infected with two pathogenic alphaherpesviruses, herpes simplex virus 1 (HSV-1) and varicella-zoster virus (VZV), confirmed the expression of five HSV-1 miRNAs, miR-H2 through miR-H6, which had previously been observed in mice latently infected with HSV-1. In addition, two novel HSV-1 miRNAs, termed miR-H7 and miR-H8, were also identified. Like four of the previously reported HSV-1 miRNAs, miR-H7 and miR-H8 are encoded within the second exon of the HSV-1 latency-associated transcript. Although VZV genomic DNA was readily detectable in the three human trigeminal ganglia analyzed, we failed to detect any VZV miRNAs, suggesting that VZV, unlike other herpes-viruses examined so far, may not express viral miRNAs in latently infected cells.
MicroRNAs (miRNAs) are a family of⬃22-nucleotide (nt) noncoding RNAs that are capable of binding to specific target mRNAs and inhibiting their expression (reviewed in reference 1). They are typically derived from one arm of RNA stem-loops found within noncoding regions of capped and polyade-nylated transcripts (4, 26). Successive cleavage of these hairpin structures by the RNase III enzymes Drosha in the nucleus (25) and Dicer in the cytoplasm (7, 20) generates a miRNA duplex structure of⬃20 bp with 2-nt 3⬘overhangs. One arm of this duplex is then loaded into the RNA-induced silencing complex (RISC), where it is used as a guide to target comple-mentary transcripts for inhibition (19, 28). In mammalian cells, miRNAs usually guide the RISC to imperfectly complemen-tary target sites, resulting in the translational arrest of bound mRNAs and a modest but detectable mRNA destabilization (12, 31, 43).
Due to their small size and nonimmunogenic nature, miRNAs appear ideally suited for use as regulatory molecules by viruses, and indeed, a number of human DNA viruses, including many herpesviruses, have now been reported to encode miRNAs (39). Herpesviruses can be divided into three subfamilies, the alpha-, beta-, and gammaherpesviruses, based on replication characteristics, genomic organization, and preferred latency sites. Members of all three subfamilies have been found to encode miRNAs, ranging from a low of 3 in the alphaherpes-virus herpes simplex alphaherpes-virus 2 (HSV-2) (37, 38) to a high of 25 in Epstein-Barr virus (EBV) (5, 17, 33, 46). The fact that all herpesviruses examined to date express miRNAs suggests that miRNAs play important roles in the herpesvirus life cycle, and several studies have in fact demonstrated the downregulation
of cellular and/or viral mRNA targets by herpesvirus miRNAs (reviewed in reference 16).
HSV-1 and varicella-zoster virus (VZV) are pathogenic hu-man viruses both of which belong to the alphaherpesvirus subfamily. HSV-1, the prototypic alphaherpesvirus, typically initiates productive replication in the mucosal epithelia of the face and establishes latency in neurons of multiple cranial nerve ganglia, including the trigeminal ganglia (TG) (35, 41). VZV replicates in the mucosal epithelia of the respiratory tract and establishes latency not only in the TG but also in the dorsal root and autonomic ganglia (15). During latency, transcription of the HSV-1 genome is largely restricted to a single RNA: the latency-associated transcript (LAT) (2, 36). Although the LAT is capped and polyadenylated, it does not appear to encode a protein. The LAT is unusual in that the spliced⬃6.3-kb tran-script is highly unstable, while the single⬃2-kb intron accu-mulates to significant levels within latently infected cells, al-though its function remains unknown (13, 21).
Previously, we used deep sequencing of RNA harvested from murine TG latently infected with HSV-1 to demonstrate that HSV-1 expresses at least five miRNAs, four of which (miR-H2, miR-H3, miR-H4, and miR-H5) are derived from the unstable exonic regions of the LAT (40). A fifth miRNA, miR-H6, was found to lie in the opposite transcriptional ori-entation, just upstream of the LAT and antisense to another HSV-1 miRNA, miR-H1, that is expressed exclusively during productive replication (11). Two of the HSV-1 LAT-derived miRNAs, miR-H2 and miR-H6, have been found to downregu-late the expression of the viral proteins ICP0 and ICP4, re-spectively (40). ICP0 and IPC4 are viral immediate-early pro-teins that function as potent activators of productive HSV-1 replication (3, 18, 34), and it has been hypothesized that their repression by miR-H2 and miR-H6 might facilitate the estab-lishment and/or maintenance of the latent state in HSV-1-infected neurons (40).
* Corresponding author. Mailing address: Department of Molecular Genetics and Microbiology, Box 3025, Durham, NC 27710. Phone: (919) 684-3369. Fax: (919) 681-8979. E-mail: bryan.cullen@duke.edu.
䌤Published ahead of print on 5 August 2009.
10677
on November 8, 2019 by guest
http://jvi.asm.org/
Humans are the only natural hosts of HSV-1 and VZV, although cell lines of various origins will support productive HSV-1 or VZV replication in culture. Importantly, neither HSV-1 nor VZV latency can be established in vitro using currently available cell culture systems. Although various ani-mal models can be latently infected with either HSV-1 or VZV by artificial means (30, 35, 45), these animal models do not fully recapitulate all the hallmarks of true latent infection in humans, suggesting that aspects of the human neuronal repli-cation environment may be unique.
In order to identify viral miRNAs that are expressed by HSV-1 or VZV in latently infected human neurons in vivo, we used Solexa/Illumina technology for deep sequencing of cDNA libraries prepared from postmortem human TG samples nat-urally infected with latent HSV-1 and/or VZV. Based on these data, we demonstrate the in vivo expression of the five previ-ously reported HSV-1 miRNAs, miR-H2 to miR-H6, and we identify two novel HSV-1 LAT-derived miRNAs, miR-H7 and miR-H8. Surprisingly, despite the successful recovery of large numbers of HSV-1 miRNAs from the same samples, no la-tently expressed VZV miRNAs were identified.
MATERIALS AND METHODS
Human tissues.Both TG were removed within 61 h after death from three subjects (clinical data provided in Table 1) who, at autopsy, did not show cutaneous signs of recent herpesvirus infection. The dura, nerve roots, and connective tissue were trimmed aseptically, and the ganglia were quick-frozen in liquid nitrogen.
Nucleic acid extraction and PCR analysis.Ganglionic tissue was powdered under liquid nitrogen, and⬃30 mg was used for DNA extraction (DNeasy; Qiagen, Germantown, MD), of which 100 ng was analyzed for the presence of HSV-1 and VZV DNA by simultaneous fluorescence-based amplification and product detection (real-time PCR). Based on the complete sequences of HSV-1 (NC_001806) and VZV (NC_001348), forward primer TGGTATTGCCCAAC ACTTTCC, reverse primer GCGCCAGGCACACACAT, and probe CGTGTC GCGTGTGGT were used to assay for HSV-1, while forward primer CGAAC ACGTTCCCCATCAA, reverse primer CCCGGCTTTGTTAGTTTTGG, and probe TCCAGGTTTTAGTTGATACCA were used to assay for VZV. Repli-cate DNA samples were amplified using the 7500-Fast real-time PCR system (Applied Biosystems, Foster City, CA) as described previously (9). The remain-ing powdered ganglionic tissue was dissolved in 7 ml Tri reagent (Molecular Research Center, Inc., Cincinnati, OH) and stored at⫺80°C. RNA was extracted according to the manufacturer’s directions.
Solexa/Illumina sequencing and data analysis.cDNA libraries for deep se-quencing were prepared using the protocol outlined by Umbach et al. in 2008 (40), adapted for the Solexa/Illumina sequencing platform. Twenty-five micro-grams of total RNA from each right and left TG were pooled and used to prepare the cDNA libraries, using the linkers described in the Solexa Digital gene expression kit, but chemically modified as described by Lau et al. in 2001 (24). In particular, the custom 3⬘linker obtained from IDT (Coralville, IA) was 5⬘adenylated and 3⬘blocked. The 5⬘DNA/RNA hybrid linker was also obtained from IDT. Raw sequence data were returned in FASTA format, and 3⬘linker sequences were removed. Final sequences were collapsed, indexed, and size filtered for reads of 18 to 24 nt in order to generate a final list for MegaBLAST analysis. Sequences were analyzed against the HSV-1 genome (NC_001806), the VZV genome (NC_001348), and the miRBASE database (release 9.2) using the
formatdb, megablast, blastoutparse, and filter_alignment scripts found in the miRDeep software package (14).
Cell culture, virus infection, and RNA extraction.Vero and HeLa cells were maintained in Dulbecco’s modified Eagle medium with 10% fetal bovine serum (FBS). SY5Y cells were maintained in RPMI medium with 10% FBS. SY5Y, HeLa, and Vero cells were infected with HSV-1 strain KOS at a multiplicity of infection of 10. Cells were washed with serum-free medium before being infected with a minimal volume of virus for 1.5 h at 37°C with occasional rocking. Virus was removed and cells were washed before the medium with 10% FBS was replenished. Samples were harvested at various hours postinfection (hpi), and total RNA was isolated using Trizol (Invitrogen, Carlsbad, CA) according to the manufacturer’s directions.
Stem-loop real-time PCR (RT-PCR).HSV-1 miRNA expression levels were analyzed with the TaqMan microRNA assay system (Applied Biosystems, Foster City, CA) according to the manufacturer’s directions, using 10 ng of total RNA per RT reaction. All miRNA levels are given as the level of expression compared to that of an uninfected control sample (27). In these experiments, RNA from subject 3, which was VZV positive but HSV-1 negative, was used as the non-HSV-1-infected control. All values were normalized against levels of the ubiq-uitous cellular miRNA miR-16, and all samples were run in triplicate. Custom primers for detecting each miRNA were designed by Applied Biosystems against the most common isoform of each miRNA recovered during deep sequencing.
RESULTS
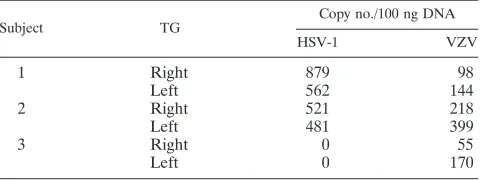
Deep sequencing of small RNAs from human TG latently infected with HSV-1 and/or VZV. TG were harvested from three recently deceased humans who did not present any symp-toms of viral reactivation at the time of death (Table 1). The left and right TG of each subject were individually assayed for the presence of both the HSV-1 and the VZV genome by real-time PCR; both TG of subjects 1 and 2 were found to harbor HSV-1 and VZV genomic DNA, while those of subject 3 contained only VZV DNA (Table 2).
Deep sequencing returned 3,086,881 usable reads from sub-ject 1, 9,152,800 reads from subsub-ject 2, and 4,063,968 reads from subject 3. Because neurons make up only⬃10% of all cells in the TG, and only a small percentage of neurons are actually latently infected by either HSV-1 or VZV (10), the vast ma-jority of sequences recovered proved, as expected, to be human cellular miRNAs. Members of the let-7 family were the
pre-TABLE 1. Clinical descriptions of tissue donors
Subject Time (h) from
death to autopsy Age (yr) Gender Cause of death Rash
Immunosuppressive therapy in the past
30 days
1 61 74 Male Sepsis with PCPa No No
2 22.5 57 Male Pancreatic cancer No Unknown
3 22.7 45 Male Myocardial infarction No No
a
[image:2.585.42.543.82.146.2]PCP,Pneumocystis jiroveciipneumonia.
TABLE 2. Analysis of viral loads in human TG by real-time PCR
Subject TG
Copy no./100 ng DNA
HSV-1 VZV
1 Right 879 98
Left 562 144
2 Right 521 218
Left 481 399
3 Right 0 55
Left 0 170
on November 8, 2019 by guest
http://jvi.asm.org/
[image:2.585.302.543.635.725.2]dominant species detected, in particular isoforms let-7a, let-7b, let-7f, and let-7c (data not shown).
Identification of HSV-1 miRNAs expressed in vivo. Deep sequencing of TG from subject 1 recovered all five HSV-1 miRNAs previously reported to be expressed in latently
in-fected murine neurons (40) except miR-H5, while all five la-tency-associated miRNAs, including miR-H5 (albeit only one copy), were recovered from subject 2 (Table 3). The HSV-1 miR-H1 miRNA, which has previously been detected only in cells undergoing productive HSV-1 replication (11, 39), was not recovered in this analysis of latently infected human TG.
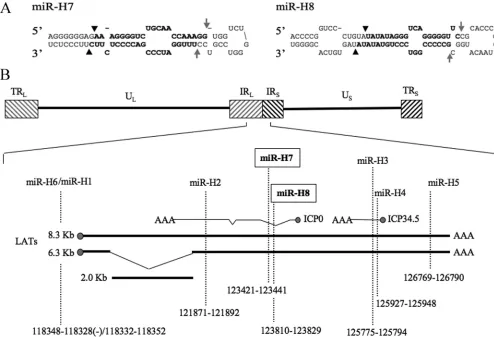
[image:3.585.43.285.88.203.2]In addition to finding all HSV-1 latency-associated miRNAs previously reported (40), we also recovered two new LAT-derived miRNAs, designated miR-H7 and miR-H8 here (Ta-ble 3). miR-H7 was recovered from both subjects 1 and 2, while miR-H8 was recovered only from subject 2. The loci encom-passing both miRNAs can be folded into the stem-loop struc-tures typical of miRNA precursors (25, 44), and recovery of mature miRNAs derived from both the 5⬘ (5p) and 3⬘ (3p) arms confirms that, when annealed, these miRNAs display the 2-nt 3⬘ overhangs characteristic of miRNA duplex intermedi-ates (Fig. 1A) (25), suggesting that they are indeed genuine miRNAs. The genomic locations of these miRNAs place them within the LAT, antisense to intron 1 ofICP0(Fig. 1B). No HSV-1 miRNAs were recovered from subject 3, whose TG did not contain detectable HSV-1 genomic DNA (Table 2).
TABLE 3. HSV-1 miRNA expression profile in human TG latently infected with HSV-1
HSV-1 miRNA
Frequencya
Subject 1 Subject 2
5p 3p 5p 3p
miR-H2 0 15 4 127
miR-H3 0 117 53 1,099
miR-H4 107 0 1,401 200
miR-H5 0 0 1 0
miR-H6 0 2 0 11
miR-H7 22 1 40 23
miR-H8 0 0 2 1
aNumber of copies of each miRNA recovered from deep sequencing. 5p and
3p indicate derivation from either the 5⬘or the 3⬘arm of the predicted miRNA stem-loop precursor, respectively.
FIG. 1. Two novel HSV-1 miRNAs recovered from latently infected humans. (A) Predicted secondary structures of the primary miRNA stem-loops of the novel HSV-1 miRNAs miR-H7 and miR-H8. Arrowheads indicate sites of Drosha cleavage, and arrows indicate sites of Dicer cleavage. The recovered, mature 5p and 3p miRNA strands are shown in boldface. (B) Schematic of the HSV-1 genome expanded to show details of the LAT locus. Relative sizes, locations, and orientations of other viral transcripts are also indicated. Sequence coordinates of viral miRNAs are given according to the HSV-1 strain 17 syn⫹genome (NC_001806). IR, internal repeat; TR, terminal repeat; UL, unique long; US, unique short.
The novel HSV-1 miRNAs miR-H7 and miR-H8 both map antisense to an intronic region in the viralICP0gene.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.585.43.537.331.668.2]Expression of HSV-1 miRNAs in latently and productively infected cells. The presence of all HSV-1 miRNAs was ana-lyzed by quantitative RT-PCR (qRT-PCR) in the original HSV-1-positive TG samples used for deep sequencing (Fig. 2A). miR-H2 through miR-H7 were found to be present at readily detectable levels; miR-H2 was the most abundant miRNA in subject 1, and miR-H4-3p was the most abundant miRNA in subject 2 (Fig. 2A). Despite our ability to recover miR-H8, albeit infrequently, from subject 2, this miRNA was not detectable by qRT-PCR using RNA recovered from either subject 1 or subject 2. Surprisingly, miR-H1 was detected, although at a low level, in subject 1, suggesting either that some neurons were undergoing viral reactivation at the time of post-mortem sample collection, despite the absence of a visible rash (Table 1), or that low levels of miR-H1 are expressed in la-tently HSV-1 infected cells.
HSV-1 miRNA expression was also assessed by qRT-PCR in productively infected SY5Y cells, a neuronal cell line of human origin. At both 6 and 18 hpi, all nine HSV-1 miRNAs, miR-H1 through miR-H8, were detectable, with miR-H1 being the most abundant, as expected (11, 39), and miR-H8 being the least abundant (Fig. 2B). miR-H6, which is induced during productive replication (40), was also detected at elevated lev-els. Concordant with LAT expression, which is low at the start of the productive replication cycle but increases through the course of infection (23), the amount of each HSV-1 miRNA increased significantly as infection progressed from 6 to 18 hpi. Examination of infected HeLa and Vero cells demonstrated that all nine HSV-1 miRNAs were also expressed in these cell lines during productive replication (Fig. 2C and D).
No VZV miRNAs were identified in latently infected human TG.Despite our ability to recover large numbers of latency-associated HSV-1 miRNAs (Table 3), no VZV-derived small RNAs were identified in samples from any of the three human subjects analyzed here. In agreement with these findings, Sol-exa/Illumina deep sequencing of small RNAs derived from TG recovered from rhesus macaques latently infected with simian varicella virus (SVV), the primate equivalent of VZV, yielded 1,420,064 usable reads, but, again, did not yield any small RNAs of SVV origin (J. L. Umbach, I. Messaoudi, and B. R. Cullen, unpublished data). Together, these results suggest that both VZV and SVV fail to express viral miRNAs during la-tency in vivo.
DISCUSSION
Although HSV-1 and VZV both readily establish productive infections in culture, these viruses are able to establish latency only in primary neurons, an environment that has so far proven impossible to fully recapitulate in culture. While HSV-1, but not VZV, can also establish latency in vivo in the sensory ganglia of laboratory animals, such as mice and rabbits, these model systems do not faithfully mimic the latent infections seen in humans, the only natural host for HSV-1. We therefore felt that it was important to assess the miRNA profiles of these viruses in naturally infected human neurons by using deep sequencing to analyze RNA samples derived from human TG latently infected with HSV-1 and/or VZV.
[image:4.585.91.499.69.292.2]Using latently infected human TG obtained postmortem, we were able to identify two new HSV-1 miRNAs (Fig. 1A and
FIG. 2. HSV-1 miRNA expression determined by qRT-PCR in latently and productively infected cells and tissues. All values are given as changes in expression relative to that of a non-HSV-1-infected control sample and were normalized to values for a cellular miRNA, miR-16. (A) HSV-1 miRNA expression levels in latently infected human TG from subject 1 (filled bars) and subject 2 (shaded bars). Despite its recovery during deep sequencing, miR-H8 was not detected in either TG sample. Subject 3 served as the negative control. (B) HSV-1 miRNA expression profiles, at 6 hpi (solid bars) and 18 hpi (shaded bars), of SY5Y cells productively infected with HSV-1 strain KOS at a multiplicity of infection of 10. (C and D) Similar to panel B, except that results for HeLa cells at 18 hpi are shown. (D) Similar to panel B, except that Vero cells at 24 hpi were analyzed.
on November 8, 2019 by guest
http://jvi.asm.org/
Table 3). The existence of one of these novel HSV-1 miRNAs, miR-H7, was verified in both latently and productively infected cells by qRT-PCR (Fig. 2), while the other miRNA, miR-H8, was detected only in productively infected cells, even though it was initially sequenced from latently infected TG (Table 3; Fig. 2). Overall, there was only a modest level of correlation in miRNA expression levels as determined by deep sequencing (Table 3) or qRT-PCR (Fig. 2A). This may reflect differences in linker ligation efficiency during cDNA synthesis, differential PCR amplification of the cDNA library during sample prepa-ration and sequencing, and/or different annealing efficiencies of the primers used for the qRT-PCR analysis.
The genomic locations of miR-H7 and miR-H8 place both antisense to the first intron ofICP0 (Fig. 1B). This location suggests that they are unlikely to downregulate ICP0 expres-sion as reported for miR-H2, which lies antisense to an exonic region ofICP0(Fig. 1B), since RNA interference is thought to operate exclusively in the cytoplasm (42). However, there is a report of alternative splicing ofICP0intron 1, which generates as many as four different introns of variable size, although the significance and function, if any, of these alternatively spliced
ICP0mRNAs are unknown (6). It is possible that miR-H7 and miR-H8 either act directly on these splice variants or, con-versely, act on currently unknown cellular mRNA targets, as previously reported for several other herpesvirus miRNAs (16).
Analysis of the mature HSV-1 miRNA sequences recovered revealed a significant level of sequence heterogeneity at both the 5⬘ends and (especially) the 3⬘ends of some of the mature HSV-1 miRNAs (Table 4). The 5⬘region of miRNAs, espe-cially nt 2 to 8, has been shown to be particularly important for target mRNA binding, and full sequence complementarity to this so-called “seed” region is usually, but not invariably, es-sential for mRNA translational inhibition (1). Therefore, even small sequence differences at the 5⬘end of the miRNA, as seen
with miR-H2-3p, miR-H6-3p, and miR-H7-5p (Table 4), have the potential to affect the identity of the mRNAs targeted by these viral miRNAs. A comparison to cellular miRNA se-quences obtained from the same RNA samples in the same sequencing run showed very little sequence variation at the 5⬘ end, with 96.0% of the 3,634,951 individual reads of let-7a and 96.3% of the 1,489,388 reads of let-7b recovered from these two HSV-1-positive small-RNA samples showing the same, predicted 5⬘end (data not shown). Therefore, it is unlikely that the sequence variation observed at the 5⬘ends of miR-H2-3p and miR-H6-3p, but not at the 5⬘end of H3-3p or miR-H4-5p (Table 4), is due to degradation during RNA isolation or some other sequencing artifact. It is also noteworthy that the minor 5⬘sequence variants observed with miR-H6-3p and miR-H7-5p are actually 1 nt longer than the consensus se-quence (Table 4), which is clearly inconsistent with exonucleo-lytic RNA degradation. Of note, because miR-H2-3p lies an-tisense to its only known target,ICP0(Fig. 1B), a 1-nt change at the 5⬘end would not affect the level of miRNA complemen-tarity to this transcript. In contrast, a 1-nt addition at the 5⬘end of miR-H6, which has been reported to target ICP4 mRNAs via the seed region (40), would be predicted to disrupt this interaction and hence may attenuate ICP4 downregulation.
The lack of detectable VZV-derived miRNAs in any of the three VZV-infected TG samples was unexpected, given that all members of the herpesviruses superfamily examined thus far, including primate, murine, and avian herpesviruses, have been found to encode multiple viral miRNAs (39). It is possible that, despite analyzing more than 1.6⫻107cDNA sequence reads,
this method was still not sensitive enough to detect VZV miRNAs, even though⬃3,000 HSV-1 miRNAs were recovered from the same samples (Table 3). As is commonly the case in coinfected TG, the HSV-1 genomic DNA load was modestly higher than that of VZV (Table 2) (10), but this minor differ-ence seems unlikely to explain the lack of VZV miRNAs
re-TABLE 4. Major species of mature HSV-1 miRNAs
HSV-1 miRNAa Sequence Length (nt) Prevalenceb
miR-H2-3p CTGAGCCAGGGACGAGTGCGA 21 18/142 (13)
CTGAGCCAGGGACGAGTGCGACT 23 46/142 (32)
TGAGCCAGGGACGAGTGCGACT 22 16/142 (11)
miR-H3-3p CTGGGACTGTGCGGTTGG 18 491/1,216 (40)
CTGGGACTGTGCGGTTGGG 19 138/1,216 (11)
CTGGGACTGTGCGGTTGGGA 20 244/1,216 (20)
CTGGGACTGTGCGGTTGGGAC 21 301/1,216 (25)
miR-H4-5p GGTAGAGTTTGACAGGCAAG 20 217/1,508 (14)
GGTAGAGTTTGACAGGCAAGCA 22 1,054/1,508 (70)
miR-H5 CAGAGATCCAAACCCTCCG 19 1/1 (100)
miR-H6-3p CCACTTCCCGTCCTTCCATCCC 22 2/13 (15)
CACTTCCCGTCCTTCCATCC 20 3/13 (23)
CACTTCCCGTCCTTCCATCCC 21 3/13 (23)
miR-H7-5p GAAAGGGGTCTGCAACCAAAGG 22 7/62 (11)
AAAGGGGTCTGCAACCAAAGG 21 39/62 (63)
miR-H8-5p TATATAGGGTCAGGGGGTTC 20 2/2 (100)
aMajor species of each HSV-1 miRNA comprisingⱖ10% of the total mature miRNA population are included. Data were pooled from subjects 1 and 2. bExpressed as the number of copies of each particular sequence isoform/number of copies of all sequences in the miRNA species (percentage).
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.585.44.546.80.300.2]ported here. Interestingly, a previously reported computational analysis of the genomes of HSV-1, HSV-2, and VZV, as well as of those of several other herpesviruses, predicted that HSV-1 and HSV-2 would encode multiple viral miRNAs but that VZV was unlikely to produce viral miRNAs in infected cells (32).
It should also be noted that the mechanism underlying VZV latency is thought to differ significantly from that observed for HSV-1. During HSV-1 latency, the only transcript expressed at significant levels is the LAT, which is contained within the repeat regions of the viral genome (Fig. 1B). Although the VZV genome shares significant homology with HSV-1, VZV lacks sequences complementary to much of the HSV-1 long-internal-repeat and long-terminal-repeat regions (Fig. 1B) (29) and is therefore not predicted to encode a LAT (10). In addi-tion, VZV differs from HSV-1 in that it expresses at least five viral protein-coding mRNAs during latency, albeit at low levels (8, 9, 22). Since most of the HSV-1 miRNAs are LAT derived, it is perhaps not surprising that VZV does not appear to express any latency-specific viral miRNAs. The apparent lack of VZV latency-associated miRNAs, however, does not pre-clude the possibility of expression of VZV miRNAs during productive replication, as is indeed observed with HSV-1 miR-H1 (11) (Fig. 2).
ACKNOWLEDGMENTS
This work was supported in part by Public Health Service grants AI067968 (to B.R.C.), AG032958 (to D.H.G. and R.J.C.), and AG006127 (to D.H.G.) from the National Institutes of Health. M.A.N. was supported by Public Health Service grant NS007321 from the National Institutes of Health. J.L.U. was supported by NIH training grant T32-CA009111.
We thank Ilhem Messaoudi for providing SVV-infected simian TG and IDT, Inc., for the 3⬘linker used during miRNA cloning.
REFERENCES
1.Bartel, D. P.2009. MicroRNAs: target recognition and regulatory functions. Cell136:215–233.
2.Bloom, D. C.2004. HSV LAT and neuronal survival. Int. Rev. Immunol.
23:187–198.
3.Cai, W., T. L. Astor, L. M. Liptak, C. Cho, D. M. Coen, and P. A. Schaffer.
1993. The herpes simplex virus type 1 regulatory protein ICP0 enhances virus replication during acute infection and reactivation from latency. J. Virol.
67:7501–7512.
4.Cai, X., C. H. Hagedorn, and B. R. Cullen.2004. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA10:1957–1966.
5.Cai, X., A. Scha¨fer, S. Lu, J. P. Bilello, R. C. Desrosiers, R. Edwards, N. Raab-Traub, and B. R. Cullen.2006. Epstein-Barr virus microRNAs are evolutionarily conserved and differentially expressed. PLoS Pathog.2:e23. 6.Carter, K. L., and B. Roizman.1996. Alternatively spliced mRNAs predicted
to yield frame-shift proteins and stable intron 1 RNAs of the herpes simplex virus 1 regulatory gene alpha 0 accumulate in the cytoplasm of infected cells. Proc. Natl. Acad. Sci. USA93:12535–12540.
7.Chendrimada, T. P., R. I. Gregory, E. Kumaraswamy, J. Norman, N. Cooch, K. Nishikura, and R. Shiekhattar.2005. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature436:740–744. 8.Cohrs, R. J., M. Barbour, and D. H. Gilden.1996. Varicella-zoster virus
(VZV) transcription during latency in human ganglia: detection of tran-scripts mapping to genes 21, 29, 62, and 63 in a cDNA library enriched for VZV RNA. J. Virol.70:2789–2796.
9.Cohrs, R. J., and D. H. Gilden.2007. Prevalence and abundance of latently transcribed varicella-zoster virus genes in human ganglia. J. Virol.81:2950– 2956.
10.Cohrs, R. J., J. Randall, J. Smith, D. H. Gilden, C. Dabrowski, H. van Der Keyl, and R. Tal-Singer. 2000. Analysis of individual human trigeminal ganglia for latent herpes simplex virus type 1 and varicella-zoster virus nucleic acids using real-time PCR. J. Virol.74:11464–11471.
11.Cui, C., A. Griffiths, G. Li, L. M. Silva, M. F. Kramer, T. Gaasterland, X. J. Wang, and D. M. Coen.2006. Prediction and identification of herpes simplex virus 1-encoded microRNAs. J. Virol.80:5499–5508.
12.Doench, J. G., C. P. Petersen, and P. A. Sharp.2003. siRNAs can function as miRNAs. Genes Dev.17:438–442.
13.Farrell, M. J., A. T. Dobson, and L. T. Feldman.1991. Herpes simplex virus latency-associated transcript is a stable intron. Proc. Natl. Acad. Sci. USA
88:790–794.
14.Friedla¨nder, M. R., W. Chen, C. Adamidi, J. Maaskola, R. Einspanier, S. Knespel, and N. Rajewsky.2008. Discovering microRNAs from deep se-quencing data using miRDeep. Nat. Biotechnol.26:407–415.
15.Gilden, D. H., M. A. Nagel, R. Mahalingam, N. H. Mueller, E. A. Brazeau, S. Pugazhenthi, and R. J. Cohrs.2009. Clinical and molecular aspects of varicella zoster virus infection. Future Neurol.4:103–117.
16.Gottwein, E., and B. R. Cullen.2008. Viral and cellular microRNAs as determinants of viral pathogenesis and immunity. Cell Host Microbe3:375– 387.
17.Grundhoff, A., C. S. Sullivan, and D. Ganem.2006. A combined computa-tional and microarray-based approach identifies novel microRNAs encoded by human gamma-herpesviruses. RNA12:733–750.
18.Halford, W. P., C. D. Kemp, J. A. Isler, D. J. Davido, and P. A. Schaffer.2001. ICP0, ICP4, or VP16 expressed from adenovirus vectors induces reactivation of latent herpes simplex virus type 1 in primary cultures of latently infected trigeminal ganglion cells. J. Virol.75:6143–6153.
19.Hammond, S. M., E. Bernstein, D. Beach, and G. J. Hannon.2000. An RNA-directed nuclease mediates post-transcriptional gene silencing in Dro-sophilacells. Nature404:293–296.
20.Hutva´gner, G., J. McLachlan, A. E. Pasquinelli, E. Balint, T. Tuschl, and P. D. Zamore.2001. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science293:834– 838.
21.Kang, W., R. Mukerjee, J. J. Gartner, A. G. Hatzigeorgiou, R. M. Sandri-Goldin, and N. W. Fraser.2006. Characterization of a spliced exon product of herpes simplex type-1 latency-associated transcript in productively in-fected cells. Virology356:106–114.
22.Kennedy, P. G., E. Grinfeld, and J. E. Bell.2000. Varicella-zoster virus gene expression in latently infected and explanted human ganglia. J. Virol.74:
11893–11898.
23.Krause, P. R., K. D. Croen, J. M. Ostrove, and S. E. Straus.1990. Structural and kinetic analyses of herpes simplex virus type 1 latency-associated tran-scripts in human trigeminal ganglia and in cell culture. J. Clin. Investig.
86:235–241.
24.Lau, N. C., L. P. Lim, E. G. Weinstein, and D. P. Bartel.2001. An abundant class of tiny RNAs with probable regulatory roles inCaenorhabditis elegans. Science294:858–862.
25.Lee, Y., C. Ahn, J. Han, H. Choi, J. Kim, J. Yim, J. Lee, P. Provost, O. Radmark, S. Kim, and V. N. Kim.2003. The nuclear RNase III Drosha initiates microRNA processing. Nature425:415–419.
26.Lee, Y., M. Kim, J. Han, K. H. Yeom, S. Lee, S. H. Baek, and V. N. Kim.2004. MicroRNA genes are transcribed by RNA polymerase II. EMBO J.23:4051– 4060.
27.Livak, K. J., and T. D. Schmittgen.2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2⫺⌬⌬CTmethod. Methods
25:402–408.
28.Maniataki, E., and Z. Mourelatos.2005. A human, ATP-independent, RISC assembly machine fueled by pre-miRNA. Genes Dev.19:2979–2990. 29.McGeoch, D. J.1989. The genomes of the human herpesviruses: contents,
relationships, and evolution. Annu. Rev. Microbiol.43:235–265.
30.Mueller, N. H., D. H. Gilden, R. J. Cohrs, R. Mahalingam, and M. A. Nagel.
2008. Varicella zoster virus infection: clinical features, molecular pathogen-esis of disease, and latency. Neurol. Clin.26:675–697.
31.Olsen, P. H., and V. Ambros. 1999. The lin-4 regulatory RNA controls developmental timing inCaenorhabditis elegansby blocking LIN-14 protein synthesis after the initiation of translation. Dev. Biol.216:671–680. 32.Pfeffer, S., A. Sewer, M. Lagos-Quintana, R. Sheridan, C. Sander, F. A.
Grasser, L. F. van Dyk, C. K. Ho, S. Shuman, M. Chien, J. J. Russo, J. Ju, G. Randall, B. D. Lindenbach, C. M. Rice, V. Simon, D. D. Ho, M. Zavolan, and T. Tuschl.2005. Identification of microRNAs of the herpesvirus family. Nat. Methods2:269–276.
33.Pfeffer, S., M. Zavolan, F. A. Grasser, M. Chien, J. J. Russo, J. Ju, B. John, A. J. Enright, D. Marks, C. Sander, and T. Tuschl.2004. Identification of virus-encoded microRNAs. Science304:734–736.
34.Preston, C. M.1979. Control of herpes simplex virus type 1 mRNA synthesis in cells infected with wild-type virus or the temperature-sensitive mutant tsK. J. Virol.29:275–284.
35.Roizman, B., D. M. Knipe, and R. J. Whitley.2007. Herpes simplex viruses, p. 2501–2601.InD. M. Knipe, P. M. Howley, D. E. Griffin, R. A. Lamb, M. A. Martin, B. Roizman, and S. E. Straus (ed.), Fields virology, 5th ed., vol. 2. Lippincott Williams & Wilkins, Philadelphia, PA.
36.Stevens, J. G., E. K. Wagner, G. B. Devi-Rao, M. L. Cook, and L. T. Feldman.
1987. RNA complementary to a herpesvirus alpha gene mRNA is prominent in latently infected neurons. Science235:1056–1059.
37.Tang, S., A. S. Bertke, A. Patel, K. Wang, J. I. Cohen, and P. R. Krause.2008. An acutely and latently expressed herpes simplex virus 2 viral microRNA
on November 8, 2019 by guest
http://jvi.asm.org/
inhibits expression of ICP34.5, a viral neurovirulence factor. Proc. Natl. Acad. Sci. USA105:10931–10936.
38.Tang, S., A. Patel, and P. R. Krause.2009. Novel less-abundant viral microRNAs encoded by herpes simplex virus 2 latency-associated transcript and their roles in regulating ICP34.5 and ICP0 mRNAs. J. Virol.83:1433–1442. 39.Umbach, J. L., and B. R. Cullen.2009. The role of RNAi and microRNAs in
animal virus replication and antiviral immunity. Genes Dev.23:1151–1164. 40.Umbach, J. L., M. F. Kramer, I. Jurak, H. W. Karnowski, D. M. Coen, and B. R. Cullen.2008. MicroRNAs expressed by herpes simplex virus 1 during latent infection regulate viral mRNAs. Nature454:780–783.
41.Warren, K. G., S. M. Brown, Z. Wroblewska, D. Gilden, H. Koprowski, and J. Subak-Sharpe.1978. Isolation of latent herpes simplex virus from the superior cervical and vagus ganglions of human beings. N. Engl. J. Med.
298:1068–1069.
42.Zeng, Y., and B. R. Cullen. 2002. RNA interference in human cells is restricted to the cytoplasm. RNA8:855–860.
43.Zeng, Y., E. J. Wagner, and B. R. Cullen.2002. Both natural and designed micro RNAs can inhibit the expression of cognate mRNAs when expressed in human cells. Mol. Cell9:1327–1333.
44.Zeng, Y., R. Yi, and B. R. Cullen.2005. Recognition and cleavage of primary microRNA precursors by the nuclear processing enzyme Drosha. EMBO J.
24:138–148.
45.Zerboni, L., C. C. Ku, C. D. Jones, J. L. Zehnder, and A. M. Arvin.2005. Varicella-zoster virus infection of human dorsal root ganglia in vivo. Proc. Natl. Acad. Sci. USA102:6490–6495.
46.Zhu, J. Y., T. Pfuhl, N. Motsch, S. Barth, J. Nicholls, F. Grasser, and G. Meister.2009. Identification of novel Epstein-Barr virus microRNA genes from nasopharyngeal carcinomas. J. Virol.83:3333–3341.