Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Molecular Characterization of Proteolytic Processing of the
Pol Proteins of Human Foamy Virus Reveals
Novel Features of the Viral Protease
KLAUS-INGMAR PFREPPER,1HANS-RICHARD RACKWITZ,2MARTINA SCHNO¨ LZER,3 HANS HEID,2MARTIN LO¨ CHELT,1ANDROLF M. FLU¨ GEL1*
Abteilungen Retroviral Gene Expression, Research Program Applied Tumorvirology,1Cell Biology,
Research Program Cell Differentiation and Carcinogenesis,2and Central Protein Analysis
Group,3German Cancer Research Center, 69009 Heidelberg,
Federal Republic of Germany
Received 6 February 1998/Accepted 5 May 1998
Spumaviruses, or foamy viruses, express a pol-specific transcript that codes for a Pol polyprotein that con-sists of the protease, reverse transcriptase, ribonuclease H, and the integrase domains. To delineate the pro-teolytic cleavage sites between the Pol subdomains, recombinant human foamy virus (HFV) Pol proteins were expressed, purified by affinity chromatography, and subjected to either HFV protease assays or autocatalytic processing. In control experiments, HFV protease-deficient mutant proteins in which the active site Asp was replaced by an Ala residue were used to rule out unspecific processing by nonviral proteases. Specific proteo-lytic cleavage products were isolated, and the cleavage sites were analyzed by amino acid sequencing. Peptides spanning the resulting cleavage sites were chemically synthesized and assayed with HFV protease, and the cleaved peptides were subjected to mass spectrometry. The cleavage site sequences obtained were in complete agreement with the amino-terminal sequences from amino acid sequencing of authentic cleavage products of the HFV Pol proteins. Analysis by fast-protein liquid chromatography of a short version of the active HFV protease revealed that the enzyme predominantly formed dimeric molecules.
Human foamy virus (HFV) is the prototypic spumavirus that has distinct features of gene expression different from those of other known retroviruses. Foamy viruses (FVs) as complex retroviruses express genes from two different promoters (3, 13, 15), and unlike other retroviruses, HFV and feline FV (FeFV) express subgenomic pol-specific transcripts (1, 4, 16, 27). Al-though recombinant forms of the HFV Pol proteins have been shown previously to express active HFV reverse transcriptase (RT), RNase H (RH), and integrase (IN), to date, proteolytic processing of HFV Pol proteins has not been analyzed, and the precise interdomain cleavage sites of HFV Pol are unknown (2, 8, 14, 19, 21). Genetic analysis has shown that the HFV pro-tease (PR) is absolutely required for infectivity and processing, since the PR(D/A) mutant, in which the Asp residue was re-placed by Ala, resulted in a noninfectious HFV provirus (9). In HFV-infected cells, the integrase has been shown to exist as a protein with a size of about 40 kDa, whereas the RT protein migrates as an 85-kDa protein (p85) under denaturing condi-tions (16, 19). Both proteins were shown to be derived from the pre127Pro-Polpolyprotein (16).
To map the precise residues that flank a given cleavage site, the following approaches were employed: (i) autocatalytic pro-cessing of His-tagged HFV Pol proteins shortened at various defined positions, (ii) in vitro PR assays with incubation of an appropriate HFV Pro-Pol protein that contained a suspected virus-specific cleavage site and subsequent isolation of suffi-cient amounts of the resulting cleavage products for amino acid microsequencing, and (iii) in vitro PR assays with syn-thetic peptides that were chosen from the regions flanking the
putative HFV-specific cleavage sites. To rule out any unspecific proteolytic cleavages, PR-deficient mutant proteins in which the Asp of the active center of the viral PR was replaced by an Ala residue were generated. In these control experiments, the PR(D/A) mutant proteins were expressed in parallel, and the cleavage pattern was directly compared to that of the authentic HFV Pol proteins. Three His-tagged Pol polyproteins differing only in the COOH-terminal regions, namely viral inserts 2, 3, and 6 (Fig. 1), were cloned into the pET22b plasmid vector by PCR, expressed in Escherichia coli BL21(DE3) cells, purified by affinity chromatography on Ni21-chelate columns, and re-acted with a polyclonal antiserum directed against HFV PR (20). The immunoblot showed that the full-length proteins were expressed and affinity purified, and the NH2-terminal PR domain reacted with the PR antiserum (not shown). The ap-parent molecular masses of the PR-RT-RH-DIN and PR-DRT Pol proteins were in close agreement with the calculated val-ues. As expected, the PR-RT-DRH protein migrated slightly faster, consistent with previous reports (8). Remarkably, the PR subdomain was retained in the three Pol proteins, as shown by the positive immunoreaction with a polyclonal antiserum directed against PR.
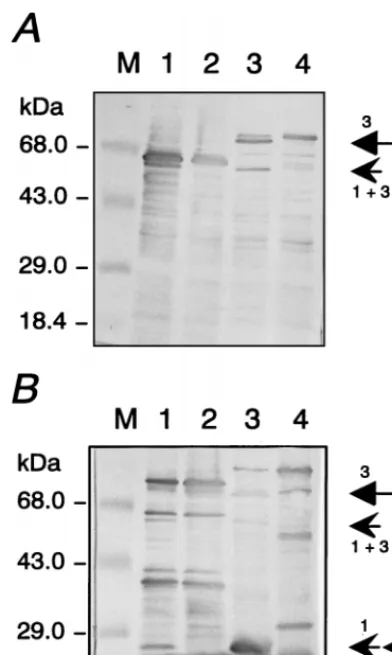
Upon close inspection, however, it was observed that in total bacterial lysates, various expressed HFV Pol proteins were invariably accompanied by autocatalytic processing (Fig. 2). To show autocatalytically proteolytic processing, the pET22b plas-mids containing the HFV pol inserts were expressed in E. coli BL21(DE3) cells, and whole bacterial lysates analyzed by im-munoblotting. The 72-kDa HFV PR-RT-DRH protein was partially cut to a smaller Pol protein of about 67 kDa (Fig. 2A, lane 1), which was a specific cleavage product, since it was not detected in the control reaction with the PR-inactive D/A mutant protein (lane 2). The same specific cleavage product as that in lane 1 was identified upon autocatalytic processing of * Corresponding author. Mailing address: Abteilung Retroviral Gene
Expression, Applied Tumorvirology, DKFZ, INF 242, 69009 Heidel-berg, Germany. Phone: 49-6221-424611. Fax: 49-6221-424865. E-mail: [email protected].
7648
on November 9, 2019 by guest
http://jvi.asm.org/
PR-RT-RH-DIN (arrowhead 113 in Fig. 2A, lane 3). During expression of the HFV PR-RT-RH-DIN protein, another cleavage event must have occurred, since a novel band with a size of about 85 kDa was observed in lane 3 (arrow 3). The additional cleavage products were not observed because of their small molecular sizes of about 3 to 4 kDa. Again, the cleavage products were not detectable in the PR(D/A) mutant protein (lane 4). In order to obtain protein bands suitable for amino acid sequencing, Pol proteins were expressed with longer COOH-terminal extensions that contain another PR(D/A) sequence (Fig. 1) reactive with antiserum against HFV PR (20). The viral inserts are schematically shown in Fig. 1 as proteins 4 and 5. Bacterial expression and autocatalytic processing of the PR-RT-DRH-TH-PRO(D/A) and of PR-RT-RH-DIN-TH-PRO(D/A) proteins is illustrated in Fig. 2B in parallel with the double PR-deficient mutants. Both recombi-nant proteins were autocatalytically processed to the cleavage product of about 67 kDa observed previously (arrowhead 113) that corresponds to the PR-RT domains shared by both pro-teins (lanes 1 and 3). In addition, a relative large protein with a size of about 85 kDa was detectable in lane 3 (labeled with arrow 3), as expected for a cleavage between the RH and the
IN domains of PR-RT-RH-DIN-TH-PRO(D/A) with one of the PR domains active.
It is noteworthy that two protein bands with sizes of about 27 kDa were additionally identified (double arrowheads 1 and 3). These sizes are consistent with the calculated values for the COOH-terminal extensions of inserts 4 and 5. Importantly, a cleavage product with a relatively high intensity of about 27 kDa was observed after autoprocessing of insert 5. The PR-inactive D/A mutant proteins showed unspecific bands that did not comigrate with the actual cleavage products in lanes 1 and 3. To determine the cleavage sites, the reaction products of 27 kDa were affinity purified and subjected to amino acid micro-sequencing (data not shown). The results revealed that the site where the amino terminus of the integrase was cleaved from the RH domain consisted of the sequence (NH2)-CNTKKPN LDA. The amino-terminal part of the RT-RH cleavage site obtained by amino acid sequence analysis was (NH2)-YTDG SAIKS (data not shown). Both sequences are unique and occur at the appropriate locations in the HFV Pol protein sequence deduced from nucleotide sequencing of the infectious HFV DNA (12).
[image:2.612.112.486.70.278.2]To independently confirm and prove the authenticity of the
FIG. 1. Schematic diagram of the different forms of viral and recombinant HFV Pol polyproteins. The top line represents the structure of the HFV pre127Pro-Pol protein. Structures of viral inserts cloned into pET22b vector plasmids are schematically shown in lines 2, 3, and 6. Inserts 4 and 5 contain longer carboxy-terminal extensions [TH(His)6-PRO(D/A)] of about 25 kDa to facilitate solubility, immunodetection, and amino acid sequencing. M, all inserts start with the authentic Met; TH, His-tagged thioredoxin (20).
TABLE 1. Locations of proteolytic cleavage sites of FV proteins
Virus Location of cleavage site in FV protein:
PR-RT RH-IN RT-RH p70-p3
HFV HWEN2QVGH YVVN2CNTK EGVF2YTDG RAVN2TVTQa
SFV-1b HWEN2QVGH YVVH2CNTT AMVF2YTDG RSVN2TVTA
SFV-3b HWEN2QVGH YVVN2INTT SMVF2YTDG RNVD2TVTA
BFVc CWEN2QVGH YKVF2SNEL EAIF2YTDG SAVH2SVRL
FeFV SWEN2QVGH FKVH2MTKN QHIF2YTDG AAVH2TVKA
Consensus XWEN2QVGH Y/FXVX2XN/TXX XXV/IF2YTDG XXVX2T/SVXX
aReported previously by Pfrepper et al. (20). bSFV-1 and SFV-3, simian foamy virus types 1 and 3. cBFV, bovine foamy virus.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.50.553.601.701.2]cleavage sites, peptides that span the processing site were syn-thesized and assayed in vitro by either the HFV PR-DRT-His or TH-PRO in the presence of EDTA (20). Synthetic peptides that correspond to the RH-IN and the RT-RH sites were sub-jected to PR assays, and the cleavage products were analyzed by matrix-assisted laser desorption ionization (MALDI) mass spectrometry (20). This analysis revealed that pro-teolytic cleavage of the two peptides occurred at the sites marked by the vertical arrows TQGSYVVN2CNTKK and PEGVF2YTDGSR, respectively, in agreement with the known HFV sequences. The resulting cleavage sites are compiled in Table 1. In parallel, apparent reaction products after incuba-tion with HFV PR-deficient D/A mutant proteins were also subjected to analysis by MALDI mass spectrometry; mass
peaks with sizes that corresponded to specific reaction prod-ucts were not found.
So far, two processing sites of the HFV Pol protein have been identified, namely those between the RT and RH and the RH and IN domains. To determine which of the different proteolytically active forms of PR-Pol was the shortest version, a 179-amino-acid-long recombinant Pol protein, PR-DRT-His (protein 6 in Fig. 1) that starts from the first Met residue of Pol and extends to residue 159, followed by a 14-amino-acid-long stretch of a vector-derived peptide sequence and a hexa-His tag was subjected to analysis. The PR-DRT-His protein was bacterially expressed and purified in parallel with the
corre-FIG. 2. Bacterial expression and concomitant autocatalytic processing of au-thentic and deficient mutant Pol polyproteins. (A) Immunoblots of the PR-RT-DRH (lane 1) and the corresponding PR-deficient D/A mutant proteins (lane 2); PR-RT-RH-DIN and the corresponding PR(D/A) mutant proteins (lanes 3 and 4) reacted with antiserum against HFV PR. Arrowhead 113 marks the proteolytic cleavage products with sizes of about 67 kDa in lanes 1 and 3; arrow 3 indicates the reaction product with a size of 85 kDa in lane 3; M, marker proteins with apparent molecular masses shown to the left. For the structures of the HFV inserts, see Fig. 1. (B) Immunoblot of PR-RT-DRH-TH-PRO(D/A) (lane 1) and the corresponding PR-deficient double D/A mutant proteins (lane 2) reacted with antibody against HFV PR. Arrowhead 113 marks the PR-RT cleavage product p67, and arrowhead 1 marks a product with a size of about 27 kDa derived from the carboxy-terminal part of insert 4 (Fig. 1). Panel B also represents an immunoblot of processed PR-RT-RH-DIN-TH-PRO(D/A) and the corresponding PR(D/A) double mutant proteins (lanes 3 and 4). Arrowhead 113 points to the p67 PR-RT cleavage product, arrow 3 points to the p85 reaction product, and arrowhead 3 marks a product with a size of about 27 kDa derived from the COOH-terminal regions of HFV insert 5. Double PR(D/A) mutant proteins served as controls (lanes 2 and 4).
FIG. 3. Autocatalytic processing of PR-DRT-His proteins (viral insert 6). (A) Gel electrophoretic analysis and staining with Coomassie blue of PR-deficient D/A mutant PR-DRT-His. Lane 1, PR-DRT-His protein; lane 2, band of the PR-DRT-His substrate with a size of about 21 kDa (arrow) and HFV PR as cleavage product of 17 kDa (arrowhead). The product cleaved at residue 166 (from within the vector backbone protein sequence) with a size of 19.6 kDa is marked with an open circle) (see panel B); the double arrowhead marks the two comigrating faint cleavage products of low molecular mass used for N-terminal sequencing. M, low-molecular-mass markers. (B) Schematic drawing of the pro-teolytically processed PR-DRT-His protein cleaved at sites 143 and 166 resulting in four cleavage products; the two smaller peptides comigrated under the con-ditions used. Both the 3.5- and 1.5-kDa bands were microsequenced. N- and C-terminal residues are in the one-letter amino acid code.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.314.540.218.614.2]sponding PR-deficient D/A mutant in order to rule out unspe-cific proteolytic cleavages. Surprisingly, autocatalytic process-ing of the purified PR-DRT-His (21 kDa) protein exhibited three additional bands that were identified after sodium do-decyl sulfate-polyacrylamide gel electrophoresis (Fig. 3) and characterized. N-terminal amino acid sequencing of the fast-est-moving protein band (marked by two arrowheads in Fig. 3) resulted in the sequence (NH2)-QVGHRKIR and another protein described below. This sequence is located at position 144 of HFV Pol. Thus, the cleavage between the PR and the RT domains occurred at HWEN2QVGH (Table 1). The band moving at about 17 kDa was characterized as 143-resi-due-long HFV PR that was proteolytically active, since it cleaved the p3Gagfrom the pre74Gag(reference 20 and data not shown).
An additional cleavage product derived from processing of the HFV PR-DRT-His protein of 179 amino acids was ob-served. Figure 3B schematically shows the processing patterns. The analysis revealed that a cleavage occurred within the vec-tor-derived part of the recombinant PR-DRT-His protein. Mi-crosequencing identified this site as KLAAALE(H), which is actually contained within the flanking sequencing of the hexa-His tag of pET22b as SSSVD2KLAAALE(H)6. MALDI mass spectrometry of the cleaved HFV PR-DRT-His protein con-firmed this result independently.
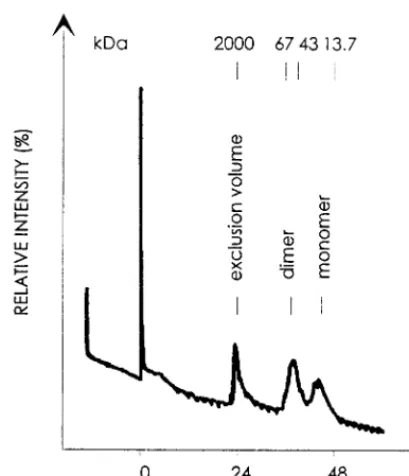
To analyze the molecular structure of the enzymatically ac-tive HFV PR, PR-DRT-His protein was purified by Ni21 -che-late affinity chromatography and subsequently subjected to fast-protein liquid chromatography (FPLC) in PR reaction buffer on a Superose 12 HR10/30 column (Pharmacia) calibrated with molecular size markers as shown in Fig. 4. The resulting elution profile clearly showed two peaks in a region, as ex-pected. The first eluted at approximately 43 kDa, consistent with the dimeric form of the HFV PR-DRT-His protein (Fig. 4). The second peak was eluted at about 20 kDa, consistent with the molecular size of the monomeric HFV PR-DRT-His protein. Subsequent analysis by sodium dodecyl
sulfate-poly-acrylamide gel electrophoresis of both protein peaks eluted showed that not only the monomeric but also the dimeric PR-DRT-His peak comigrated as a protein band of 21 kDa, as expected. This result is in agreement with the properties of other retroviral enzymes of the aspartic PR family that were unambiguously shown to exist as active homodimers (23, 25, 26). We identified three cleavage sites of the HFV Pol polypro-tein by means of immunoblots with HFV PR-specific antisera, isolation of defined cleavage products from soluble, affinity-purified recombinant proteins that contained authentic HFV Pol sequences, and subsequent amino acid sequencing. Resi-dues spanning the cleavage sites were chemically synthesized; the resulting peptides were subjected to proteolysis by the HFV PR (10) and analyzed by MALDI mass spectrometry. The PR-deficient D/A mutant proteins were consistently em-ployed to control the specificity of the proteolytic cleavages observed. Our data are consistent with the molecular sizes of virtually all HFV Pol proteins reported to occur in HFV-infected cells (5, 8, 16, 18). The mature HFV Pol proteins encompass the integrase, the p85RTthat also contains the RH domain, and the p67RT. Since an RH with the calculated value of 17 kDa was not detectable in HFV-infected cells, it is likely that the active HFV RT consists of a heterodimer of p85/p67 comparable to those of other retroviruses (7). Our results open the way to prove this hypothesis. Since only N-terminal and no C-terminal sequencing was carried out, we cannot rule out whether additional spacer peptides exist.
As to the HFV PR, our data show that distinct forms of PR were proteolytically active. One short form of HFV PR, PR143, was capable of cleaving the HFV Gag precursor into p70 and p3 and the two peptides that link the RT-RH and the RH-IN domains (data not shown). One of the open questions is which form of the proteolytically active HFV PR is respon-sible for the individual steps of Pol processing in vivo.
[image:4.612.68.273.70.308.2]It is noteworthy that forms of the HFV Pol proteins in which the PR domain was removed were not consistently observed in infected cells. The cleavage between the RH domain and the IN domain was faster and more efficient than those of the two other sites that were cleaved with suboptimal efficiency. This was also observed during proteolysis of the corresponding junction peptides. It is worth mentioning that processing of the site spanning the PR and RT was even less efficient. The poor cleavage efficiency of this site might be one reason that only minute amounts of PR143 are available for processing of FV Gag proteins that in virus-infected cells has been reported to be invariantly incomplete (14, 27). It seems possible that the PR-RT cleavage might not be required for virus infectivity. The inefficient cleavage between residues 143 and 144 may also be reflected by the fact that the PR143 did not cleave the corresponding peptides of various lengths that span this site (data not shown). Close examination of the predicted second-ary structures of HFV PR by the EMBL phd program (22) showed that FV PR-specific residues from 121 through 144 form a stable alpha helix. A search program in the data banks revealed that part of this HFV PR sequence from residues 128 to 139, KTLFVKYDNLWQ, was highly homologous to an alpha-helical sequence, KKLLTKYDNLFE (identical residues are underlined), of the galactose-1-phosphate-uridyltransfer-ase as determined by X-ray crystallography (11, 24). The three-dimensional structure of an FV PR will be required to solve this question and related issues. Table 1 shows that FV PRs seem to prefer Val at the P2 or P29position, the scissile bond being P1 and P19(23, 26). A comparison of the flap regions of well-studied retroviral PRs with those of the HFV PR illus-trates large differences and relatively few common features (6, 17, 23, 26).
FIG. 4. Identification of purified HFV PR-DRT-His dimers by FPLC sepa-ration. A Superose 12 HR10/30 column was calibrated with the marker proteins as indicated at the top. HFV PR reaction buffer was used to load and elute the affinity-purified HFV PR-DRT-His protein monomer and dimer.
on November 9, 2019 by guest
http://jvi.asm.org/
We thank Helmut Bannert for excellent technical assistance, Jen-nifer Reed for critically reading the manuscript, and Harald zur Hausen for support.
REFERENCES
1. Bodem, J., M. Lo¨chelt, I. Winkler, R. P. Flower, H. Delius, and R. M. Flu¨gel. 1996. Characterization of the spliced pol transcript of feline foamy virus: the splice acceptor site of the pol transcript is located in gag of foamy viruses. J. Virol. 70:9024–9027.
2. Coffin, J. M. 1996. Retroviridae: the viruses and their replication, p. 1767– 1847. In B. N. Fields, D. M. Knipe, and P. M. Howley (ed.), Fields’ virology. Raven Press, New York, N.Y.
3. Cullen, B. R. 1991. Human immunodeficiency virus as a prototypic complex retrovirus. J. Virol. 65:1053–1056.
4. Enssle, J., I. Jordan, B. Mauer, and A. Rethwilm. 1996. Foamy virus reverse transcriptase is expressed independently from the Gag protein. Proc. Natl. Acad. Sci. USA 93:4137–4141.
5. Giron, M.-L., F. Rozain, M. C. Debons-Guillemin, M. Canivet, J. Peries, and
R. Emanoil-Ravier.1993. Human foamy virus polypeptides: identification of
env and bel gene products. J. Virol. 67:3596–3600.
6. Hayakawa, T., Y. Misumi, M. Kobayashi, Y. Yamamoto, and Y. Fujisawa. 1992. Requirement of N- and C-terminal regions for enzymatic activity of human T-cell leukemia virus type I protease. Eur. J. Biochem. 206:919–925. 7. Katz, R. A., and A. M. Skalka. 1994. The retroviral enzymes. Annu. Rev.
Biochem. 63:133–173.
8. Ko¨gel, D., M. Aboud, and R. M. Flu¨gel. 1995. Molecular biological charac-terization of the human foamy virus reverse transcriptase and ribonuclease H domains. Virology 213:97–108.
9. Konvalinka, J., M. Lo¨chelt, H. Zentgraf, R. M. Flu¨gel, and H.-G. Kra¨usslich. 1995. Active foamy virus proteinase is essential for virus infectivity but not for formation of a Pol polyprotein. J. Virol. 69:7264–7268.
10. Kotler, M., R. A. Katz, W. Danho, J. Leis, and A. M. Skalka. 1988. Synthetic peptides as substrates and inhibitors of a retroviral protease. Proc. Natl. Acad. Sci. USA 85:4185–4189.
11. Leslie, N. D., E. B. Immerman, J. E. Flach, M. Florez, J. L. Fridovich-Keil,
and L. J. Elsas.1992. The human galactose1-phosphate uridyltransferase gene. Genomics 14:474–480.
12. Lo¨chelt, M., H. Zentgraf, and R. M. Flu¨gel. 1991. Construction of an infec-tious DNA clone of the full-length human spumaretrovirus genome and mutagenesis of the bel 1 gene. Virology 184:43–54.
13. Lo¨chelt, M., W. Muranyi, and R. M. Flu¨gel. 1993. Human foamy virus
genome possesses an internal, Bel 1-dependent and functional promoter. Proc. Natl. Acad. Sci. USA 90:7317–7321.
14. Lo¨chelt, M., and R. M. Flu¨gel. 1995. The molecular biology of primate spumaviruses, p. 239–292. In J. A. Levy (ed.), The Retroviridae, vol. 4, Plenum Press, New York, N.Y.
15. Lo¨chelt, M., S. F. Yu, M. L. Linial, and R. M. Flu¨gel. 1995. The human foamy virus internal promoter is required for efficient gene expression and infec-tivity. Virology 206:601–610.
16. Lo¨chelt, M., and R. M. Flu¨gel. 1996. The human foamy virus pol gene is expressed as a Pro-Pol polyprotein and not as a Gag-Pol fusion protein. J. Virol. 70:1033–1040.
17. Miller, M., M. Jaskolski, J. K. M. Rao, J. Leis, and A. Wlodawer. 1989. Crystal structure of a retroviral protease proves relationship to aspartic protease family. Nature 337:576–579.
18. Netzer, K.-O., A. Schliephake, B. Maurer, R. Watanabe, A. Aguzzi, and A.
Rethwilm.1993. Identification of pol-related gene products of human foamy virus. Virology 192:336–338.
19. Pahl, A., and R. M. Flu¨gel. 1993. Endonucleolytic cleavages and DNA-joining activities of the integration protein of human foamy virus. J. Virol.
67:5426–5434.
20. Pfrepper, K.-I., M. Lo¨chelt, M. Schno¨lzer, and R. M. Flu¨gel. 1997. Expres-sion and molecular characterization of an enzymatically active recombinant human spumaretrovirus protease. Biochem. Biophys. Res. Commun. 237: 548–553.
21. Rethwilm, A. 1995. Regulation of foamy virus gene expression. Curr. Top. Microbiol. Immunol. 193:1–24.
22. Rost, B., and C. Sander. 1994. Combining evolutionary information and neural networks to predict protein secondary structure. Proteins 9:55–72. 23. Skalka, A. M. 1989. Retroviral proteases: first glimpses at the anatomy of a
processing machine. Cell 56:911–913.
24. Wedekind, J. E., P. A. Frey, and I. Rayment. 1995. Three-dimensional struc-ture of galactose-1-phosphate uridyltransferase from E. coli at 1.8 A reso-lution. Biochemistry 34:11049–11061.
25. Wlodawer, A., M. Miller, M. Jaskolski, B. K. Sathyanarayana, E. Baldwin,
I. T. Weber, L. M. Selk, L. Clawson, J. Schneider, and S. B. H. Kent.1989. Conserved folding in retroviral proteases: crystal structure of a synthetic HIV-1 protease. Science 245:616–621.
26. Wlodawer, A., and J. Erickson. 1993. Structure-based inhibitors of HIV-1. Annu. Rev. Biochem. 62:543–585.
27. Yu, S. F., D. N. Baldwin, S. R. Gwynn, S. Yendapalli, and M. L. Linial. 1996. Human foamy virus replication—a pathway distinct from that of retroviruses and hepadnaviruses. Science 271:1579–1582.