0022-538X/93/095617-06$02.00/0
Copyright© 1993, AmericanSocietyforMicrobiology
Functional Analysis of Interactions between
Tat and the
trans-Activation
Response
Element of Human
Immunodeficiency Virus Type
1 in
Cells
YINGLUO,1 STEVEN J. MADORE,2 TRISTRAM G. PARSLOW,3 BRYAN R. CULLEN,2
ANDB. MATIJAPETERLINl*
HowardHughesMedical Institute andDepartments of Medicine, MicrobiologyandImmunology'and
Pathology,3 University of California, San Francisco, California 94143, andHowardHughesMedical Institute
and Sectionof Genetics, Duke University Medical Center, Durham, North Carolina 277072
Received 12 March1993/Accepted 15 June 1993
Transcriptionaltrans-activationofthe humanimmunodeficiencyvirustype 1long terminalrepeatrequires
that the virally encoded Tat effector interacts with its targettrans-activation response element (TAR) RNA stem-loop. Althoughthearginine-rich regionof Tatfrom amino acids49to59issufficienttobindtoTAR RNA invitro,theRNA-binding domain ofTat hasnotbeendefinedinvivo. Human immunodeficiency virustype1 also encodes the Rev protein, which acts through an RNA stem-loop called the Rev-response element to transport unspliced and singly spliced viral RNA species from the nucleus to the cytoplasm. To map the RNA-binding domain of Tat, we performed assays that relied on Rev function using the heterologous RNA-tethering mechanism of Tatand theTAR.Byexaminingtheeffects of selectedtargetedmutations of Tat on the abilities ofhybrid Tat/Rev proteins to rescuethe expression of unspliced mRNA via the TAR, we demonstrated thatresiduesthroughouttheN-terminal 59 amino acids of Tatare requiredforbindingof Tat and TAR RNA in vivo.
Tat interacts with the trans-activation response element (TAR) RNAstem-loop, which is locatedatthe 5' end of all viral transcripts, to greatly increase human immunodefi-ciency virus type 1 (HIV-1)geneexpression and replication (3, 9, 15, 24). Tat is essential for high levels of virion production, cellular cytopathology, and expression of
hu-mandisease (4,8, 13).From studiestodate,Tat appears to contain two functional domains. Whereas the N-terminal domain, from amino acids 1 to48, comprises the activation domain ofTat, theadjacentarginine-rich regionfrom amino acids 49 to58 is necessary and sufficient forits binding to
TAR RNAin vitro (2, 6,31) and isrequired for its function in vivo (10, 16, 26). Furthermore, it is the number of basic amino acids inthis domain rather than theprecisesequence of arginines that is important for trans-activation (1, 5). However, if thearginine-richsequenceswereindeed the sole determinant of RNA binding in vivo, then any cellular protein withasimilar basic amino acid motif should interact with the TAR and compete for trans-activation by Tat. Clearly this does not occur. For example, whereas the arginine-rich regions of Revandbacteriophage A Nprotein
can functionally replace the basic domain ofTat, no inhibi-tion by Rev of trans-activation by Tat has been observed (28). Thus, other sequences in Tat must also contribute to
specific interactions between Tat and the TAR.
Revbindsto the Rev-response element (RRE), which is located in the middle of theenvgene,totransportunspliced and singly spliced HIV-1 RNAspecies from thenucleusto
thecytoplasm(3, 7, 19, 24). Althoughtheprecisemechanism of action of neither trans-activator is understood,the func-tions of theTARandRRE are solelytobringTat andRev, respectively, to HIV-1 RNAs. Thus,both Tatand Revcan
be targetedtoviral transcripts via heterologous
RNA-teth-* Correspondingauthor.
ering mechanisms, for example, by the coat protein of bacteriophage MS2 and its operatorRNAstem-loop(22,26, 30).Functionalequalitiesbetweenprotein-RNAinteractions of these trans-activators suggested that Rev could be
tar-getedtoHIV-1 RNAsvia Tat and the TAR and that assays of Rev function in cells could be usedtomapRNA-binding domains of Tat in vivo. To thisend,we constructed several hybrid Tat/Rev proteins and replaced the RRE with the TAR. Whereas Rev by itself could not interact with this modified target, Tat in the hybrid Tat/Rev protein efficiently targeted Revtothe TAR, which resulted in the transport of unsplicedviral RNAs from the nucleustothecytoplasm. By testing several mutants of Tat in this context, we demon-strated thatnotonlybasic but alsoactivation domains of Tat
arerequiredforbindingof Tat toTAR RNAinvivo.
MATERIALSANDMETHODS
Plasmid constructions.Plasmidswhich direct thesynthesis ofwild-type andmutanthybridTat/Revproteinswere
con-structedasfollows. pcTat and pcRev have beenpreviously described (17, 19, 29). In pcTat, the transcription termina-tion signal AATAGA was replaced by CCATGG, which formsanNcoI site, byusing synthetic oligonucleotides. In pcRev, the translation initiation site CTATGGwasmutated
toCCATGGto create anNcoI site. TheSalI-NcoI fragment, which contains the entiretatgene,wasthenligated into the SalI-NcoI sites inpcRevtoformpcTat/Rev (29).
pcTat/M1O was created in the same way as pcTat/Rev, exceptthat mutatedpcRevwasused in this construction. In pcTat/M10,LandE, whichare amino acidsatpositions78 and 79 in theactivation domain ofRev,were changedtoD and L, respectively. This mutantof Rev has no biological activity asRev(17).
pARK/Rev, pAN/Rev, pC22S/Rev, pC37S/Rev, and pK41A/Revwerecreatedby polymerasechainreaction with
5617
on November 9, 2019 by guest
http://jvi.asm.org/
1 TAT 861 REV 116
/////////zzzzzzzp
B.
TARGETS
TAR
pHIVSCAT
-HIV-1 LTR CTRRE
_lll
SD
SV4 CAT
pDM12TAR
pDM257
|SD 12TAR SA
SD 4MS2 SA
[image:2.612.117.496.71.372.2]C
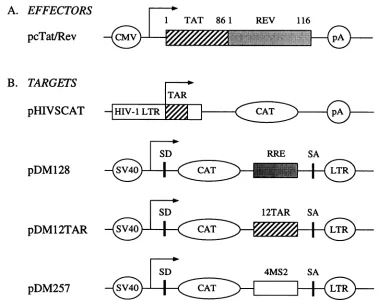
FIG. 1. Effector and targetplasmidconstructions. (A) Diagrammatic representationofplasmidswhich code forhybrid Tat/Rev proteins (pcTat/Rev). The immediate-early promoter of cytomegalovirus directs the synthesis offull-length Tat (amino acids 1 to 86) linked to full-length Rev(amino acids 1to 116). Allmutations in Tat and Revwere introduced into thisplasmid construction. (B) Diagrammatic representationof the four plasmids which codefor RNAeffectors.AlldirectthesynthesisofCAT.pHIVSCATcontains the TAR of HIV-1 inits nativepositionnext tothe site of initiationoftranscription. pDM128containssplicedonors(SD)and acceptors(SA)of HIV-1 and the RRE3'tothe CATreportergene.pDM12TARandpDM257werecreated frompDM128and contain 12 TARs(12TAR)orfour operators of
bacteriophage MS2 (4MS2)inplace of the RRE. TranscriptionofpDM128,pDM12TAR,andpDM257intitiates from the simian virus 40early
promoter(SV40) and terminates attheSV40 polyadenylationsite(pA). SeeMaterials and Methods formoredetails.
overlapping primerswhich containedappropriatenucleotide substitutions. In pARK/Rev, KKRRQRR sequences in the
arginine-richdomain ofTatwerereplaced by YVQILLY.In pAN/Rev, the N-terminal proline-richdomain of Tat (amino acids 2 to 12) was deleted. In pC22SRev, the cysteine at position 22 inTatwasreplaced bya serine. InpC37S/Rev, the cysteine at position 33 was replaced by a serine. In pK41A/Rev, the lysine at position 41 was replaced by an alanine.
pDM128,which contains thechloramphenicol
acetyltrans-ferase (CAT) reporter gene and the RRE, has been previ-ouslydescribed (12). pDM257wascreatedby replacing the RRE in pDM128 with four tandem MS2 operators (22). pDM12TARwasconstructed byreplacing theRREwith 12 tandem TARs fromp12TAR (11).
Transientexpressionassays.CV1 cellsweretransfectedby theCaPO4methodasdescribedpreviously (22).Inassaysof Tatfunction,5,ugofpHIVSCAT and 5,ug of various hybrid Tat/Rev plasmids were used in each transfection (12). pCMVA, aderivative ofpcTat/Revwith adeletion of both
the tat andrevgenes,was used as a controlplasmidin all
experiments. In assays of Rev function, 20 ,ug of hybrid
Tat/Reveffectorplasmidand 1,ugoftargetplasmid (pDM257 or pDM12TAR) were used in each transfection. For pDM128, only 1 ,ugofhybrid Tat/Rev plasmidwasusedin cotransfectionexperiments. Cellswereharvested 40to48h
after transfection. Aftertransfection, mediumwaschanged after 18 h.Cellswereharvested 40to48 h later andlysedin Triton lysis buffer. CAT assays were performed as previ-ously described (12). Protein concentrations were used to normalizeCATassaysby usingaBio-Rad kit.Experiments were performed several times in triplicate when standard errorsof the mean wereless than 30%.
RESULTS
Wild-type and mutant Tat and Revinthehybrid Tat/Rev proteinsareexpressedasfunctionalproteins.To characterize interactions between Tat and the TAR in vivo, we deter-mined RNA-binding properties of Tat independentlyof its abilitytotrans-activate transcription. For this purpose,we constructed hybrid Tat/Rev proteins and examined Rev activities in the contextof theheterologous RNA-tethering
mechanismprovided byTat and the TAR. SinceRev hasto
bind to and multimerize on the RRE, which consists of several RNA stem-loops, to transportunspliced and singly splicedviral RNAspeciesfrom the nucleustothecytoplasm (7, 19),wereasoned that several TARs inplaceof the RRE should allow Tatin hybrid Tat/Rev proteinsto performthe same function. Thus, we only had to examine the Rev activitytodeterminethebindingof Tat to TAR RNA in vivo. However,wefirst hadto demonstrate thatwild-type and
A. EFFECTORS
pcTat/Rev
pDM128
SA
LTR
on November 9, 2019 by guest
http://jvi.asm.org/
A.
Tat
En~ V)-!
Ci4 r. "4~
CI- C# t P
A
Rev
lI
Activation
l
J
Basic
Basic
l
I
Activation
FOLD-TRANS-ACTIVATION
FOLD-TRANS-ACTIVATION
n 2 3 4 5 0 10 20 30
pcTat/Rev
pTat/MIO
pAN/Rev
pC22S/Rev
pC37S/Rev
pK41A/Rev
pARK/Rev
pfHVSCAT
pDM128
FIG. 2. Activities ofhybrid Tat/Rev proteinsonthe HIV-1LTR and RRE RNAtargets.(A)Diagrammatic representation of deletions and
mutations in TatorRev, whichwereexpressedashybrid Tat/Rev proteins. Deletions and mutations in Tat (hatched bar) and Rev (shaded
bar)aredepictedabove thefull-length fusion protein. Locations of activation and basic domains of Tat and Revaregiven below the bar
diagram. (B) Inthe left and right panels are CATactivities from cotransfections with target plasmids, which contained wild-type TAR (pHIVSCAT) andRRE(pDM128)sequencesintheir nativeconfigurations,respectively,withplasmids that coded forsevenhybrideffectors,
oneofwhich containedamutantof Rev(pcTat/MlO)and theothers of which containedmutantsofTat(pAN/Rev, pC22S/Rev, pC37S/Rev, pK41AlRev, and pARK/Rev). pcTat/Rev contained wild-type Tat and Revsequences.Shownarewild-typeTat and Revsequences(solid
bars),wild-type Tatsequenceswith mutations inRev (hatched bars), andwild-type Revsequenceswith mutations inTat (shaded bars).
Resultsarerepresentativeofseveral experimentsperformedintriplicate. Standarderrorsof themeans wereless than 30%.
mutated hybrid Tat/Rev proteinswere expressed and func-tioned in cells. To this end, effector plasmids, which ex-pressed mutants of Tat or Rev in the context of hybrid Tat/Revproteins, were cotransfected withtarget plasmids, which contained either HIV-1 long terminal repeat (LTR) targets (pHIVSCAT) for Tat assays or RRE targets (pDM128)for Revassays,intoCV1cells(Fig. 1).In
pHIVS-CAT, the HIV-1 LTR directs the synthesis of the CAT reporter gene. In pDM128, the CAT reporter gene was inserted into the env intron of HIV-1 such that increased CAT activities were observed only ifunspliced RNAwas transported from the nucleus to the cytoplasm by Rev.
Representative plasmid constructions are diagrammed in Fig. 1.
Results of these cotransfectionsarepresentedinFig.2B. Consistent with previous observations and in contrast to pcTat/Rev, pAN/Rev, pARK/Rev, pC22S/Rev, pC37S/Rev,
andpK41A/Rev,whichcontained deletionsormutations in activationand basicdomains of Tat(Fig. 2A),alleliminated trans-activationbyTat (Fig. 2B,leftpanel). pcTat/Revand
pcTat/M10,which contained wild-type Tat, trans-activated
pHIVSCATfour-tofivefold (Fig. 2B, leftpanel). It should be noted that muchhigherlevels oftrans-activation, which
were equivalent to those with Tat expressed as a single protein, were observed with these plasmids in HeLa cells
(datanotpresented). However, Revfunctionednormallyin thesemutantsofthe hybrid Tat/Rev protein (Fig. 2B, right panel). The sole exceptionwaspcTat/MlO, which lacked a functional activation domain of Rev andwas inactive (Fig. 2B, right panel). Thus, pcTat/Rev, pAN/Rev, pARK/Rev, pC22S/Rev, pC37S/Rev, and pK41A/Rev increased CAT activities frompDM128from15-to30-fold inCV1 cells(Fig. 2B, right panel). These results confirmed that all hybrid Tat/Rev proteins were expressed and functioned in CV1
cells.
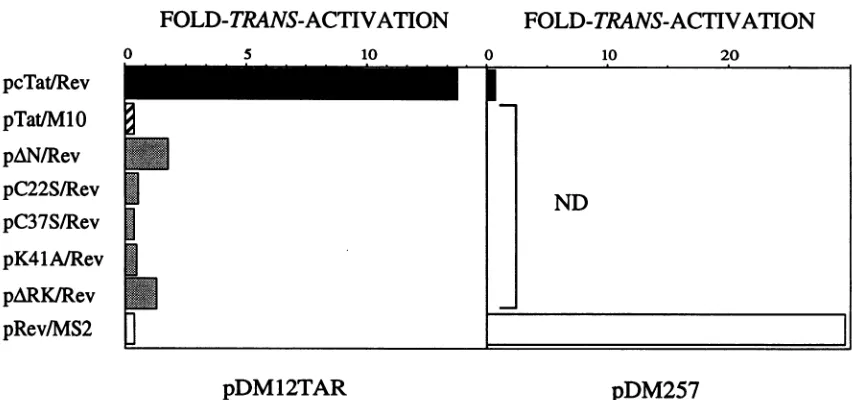
Hybrid Tat/Revproteins functionasRev viathebindingof Tat to TAR RNA. To determine whether interactions be-tween Tat and the TAR could replace those between Rev and theRRE,wecotransfectedpcTat/RevwithpDM12TAR
into CV1 cells. InpDM12TAR, 12tandemly repeatedTARs
replacedthe RRE inpDM128(Fig. 1B). Asdemonstratedin the left panelofFig.3, pcTat/Revincreased CAT activities frompDM12TAR14.5-fold. To furtherprovethat thiseffect wasdue tothe activation domain ofRev, a mutantofRev, pcTat/M10, was also cotransfected with pDM12TAR into CV1 cells. TheactivityofRev via the RREwasabolishedin
B.
00,A
L
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.104.537.69.390.2]FOLD-TRANS-ACTIVATION
pcTatlRev
pTat/M1O
pAN/Rev
pC22S/Rev
pC37S/Rev
pK41A/Rev
pARK/Rev
pRev/MS2
FOLD-TRANS-ACTIVATION
10 20
pDM12TAR
pDM257
FIG. 3. Activities of hybrid Tat/RevproteinsonTAR andoperatorRNAtargets.Inthe left panelareCATactivities from cotransfections
of thetargetplasmid,which contained 12 TARs(pDM12TAR), with plasmids which coded for hybrid effectors. pRev/MS2 containedthe full-lengthRev linkedtothecoatproteinofbacteriophageMS2. In theright panelareCAT activitiesfrom cotransfections ofacontroltarget
plasmid, which contained fouroperatorsof MS2 (pDM257), with thesame effector plasmids. pRev/MS2 servedasthe positivecontrol for transfections withpDM257 (open bars).ND(not done) referstoexperiments whichwerenotperformed. Other designationsare asinFig.2. Resultsarerepresentative of several experiments performed in triplicate. Standarderrorsof themeans wereless than 30%.
this mutant (Fig. 3, left panel). This result confirmed that
increased CAT activitieswereduesolelytothe mechanism
of action of Rev. However, Tat, by interacting with the
TAR, broughtRev in these hybrid Tat/Rev proteins to the
unspliced RNA.
Totestthespecificityof thebindingofTattoTARRNA, multiple TARswere also replaced by four operators from
bacteriophageMS2 (pDM257) (Fig. 1B).Cotransfections of
pcTat/Revwith pDM257 resulted in no trans-activation in CV1 cells(Fig. 3, right panel).We conclude that thebinding
of TattoTAR RNA inpDM12TARisspecificand isrequired
for the effects of Rev.
The activationdomain of Tat isrequiredfor thebindingof
TattoTAR RNA in vivo. Invitro, the basic nine amino acids from positions 49 to58 are sufficient forbinding of Tat to TARRNA(1). This basic domain is also absolutely required
for trans-activation in vivo (10, 16, 26). However, the N-terminal 48 amino acids represent theactivation domain of Tat thatcan functionindependentlyoftheRNA-binding
domain when presented to the transcription complex via
heterologousRNA- andDNA-tetheringmechanisms(14, 26, 29).To determine whether thesearginine-richsequencesare also sufficient for binding of Tat to TAR RNA in vivo,
various deletions and mutations of Tat in hybrid Tat/Rev proteinswere examined. In pAN/Rev, pARK/Rev, pC22S/ Rev, pC37S/Rev, andpK41A/Rev,N-terminalsequencesof Tat were deleted, the basic domain ofTat was replaced, cysteines at positions 22 and 37 in Tat were replaced by serines,and thelysineatposition41 inTatwasreplaced by an alanine. Except in the case ofpARK/Rev, all of these mutationswerein the activation domainof Tat.
As presented in the left panel of Fig. 3, none of these mutants of Tat trans-activated pDM12TAR. Among these plasmid effectors, pARK/Rev and pAN/Rev increased CAT
activities only 1.29- and 2-fold, or 9 and 14% of levels observed withpcTat/Rev, respectively, and other mutations
inthecoreand cysteine-rich domainsofTatresultedinno detectable trans-activation (Fig. 3, leftpanel, pK41A/Rev,
pC37S/Rev,andpC22S/Rev).These dataareconsistent with therequirementof ninebasic residues forbindingof Tatto TAR RNA in vitro. Moreover, theyindicate that coreand
cysteine-richdomainsarealsoabsolutely requiredfor inter-actions between Tat and the TAR in cells. Given thegreatly
reducedactivityof Tatwitheventhe deletion of N-terminal aminoacids(pAN/Rev),weconclude that the entire activa-tion domain of Tat is required forbinding of Tat to TAR RNA in vivo.
DISCUSSION
By using hybrid Tat/Rev proteins,westudied the require-ments forbinding of Tat toTAR RNA in vivo. Since the
originaltargetplasmid pDM128contained the CAT reporter gene in the intron ofthe envgene, CATactivity increased
only if unspliced transcripts were transported from the nucleustothecytoplasm byRev(12). However, the modi-fiedtargetplasmidpDM12TARcontained the TAR inplace
of the RRE. To increase the CAT activities of thisplasmid, unspliced transcriptshadtobetransportedfrom the nucleus to the cytoplasm via the binding of the hybrid Tat/Rev protein toTAR RNA. Thus, interactions between Tat and the TARreplaced thosebetween Rev and the RRE in our
assay. However, the activation domain of Rev still
per-formed the function of RNA transport.By mutatingTat and Rev in these hybrid proteins, we demonstrated that basic andactivationdomains of Tatarerequiredfor thebindingof TattoTAR RNA invivo.
In this study, Revcould not bindto TAR RNA(Fig. 3, pRev/MS2andpDM12TAR). Previously,basic amino acids wereexchanged betweenRev andTat, and this substituted Tat still trans-activated the HIV-1LTR,whichmightsuggest that the full-length Rev could interact with the TAR (28). However, hybrid Tat/Rev proteins, which contained mu-tants ofTat, did notfunction via the TAR in pDM12TAR (Fig. 2),nordid Rev inthehybrid Tat/Rev proteininterfere with trans-activation of the HIV-1 LTR(Fig. 2). Moreover,
ND
~~~~~~~~I
0 5 10 0
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.85.514.68.268.2],.
0000000
NS
'.CO
,CRIPTION
.IPLEX
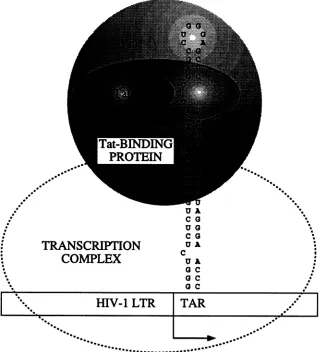
FIG. 4. Model for interactionsbetweenTat andtheTAR in vivo. Although the basic nine amino acids from positions 49to58aresufficient
forbindingof TattoTAR RNA invitro,theactivation domain of Tat from amino acids 1to48 isalsorequiredforproductiveinteractions betweenTat and the TAR in vivo.Thus,the ninebasic residuescontactthe5'bulgeinthestem-loop,whereas theN-terminalhalfofTat interacts withacellularproteinthatnotonlyincreases thebindingtoRNAbutconveysactivationby Tattothe transcription complex. This single proteinorcomplexofproteinsmostlikelyinteracts with the centralloopinthe TAR and withacomponentof RNApolymeraseII.The
activation domain of Tat(48aminoacids)andthe minimal functional Tat of HIV-1(60 aminoacids)arerepresentedbytwoellipses,thelarger ofwhichnotonlycontains thesmalleractivationdomain of Tat but also encircles the 3'bulged nucleotides.Tat-binding proteinisrepresented byalargegrayspherethatcontactsthetranscription complex (dashed ellipse).
Tat did not block the activity of Rev, and neither trans-activator aloneortogether hadanyeffect via theoperatorof MS2 in the absence of thecoat protein (Fig. 3, pcTat/Rev and pDM257). Together, these data demonstrate that inter-actions between allthreeproteins and their RNAtargetsare veryprecise and specific in vivo.
However, toobserve the effects of Rev with the hybrid Tat/Rev protein, multiple TARs in place of the RREwere required. Reducing the number of TARs from 12 to 2 completely eliminated the Rev effect (data not presented).
Since thismirrors therequirementfor Revtomultimerizeon
theRRE(18),wepostulate thatmultiple TARsarenecessary toachieve sufficientdensity of Rev for their transportfrom
thenucleustothecytoplasm. Similar resultswereobtained
when the coat protein ofbacteriophage MS2 was used to target Rev to the RNA, i.e., up to four operators were neededforhigh levels of trans-activation (22). However,we cannotabsolutely exclude the possibility that multiple TARs
areonly requiredfor theproperfoldingofoneor afew TARs in the middle ofa long primary transcript, especially since
the TAR(AG.= -35kcal) (1 cal=4.184J)(25) hasahigher
predicted freeenergythan the RRE(AG = -115 kcal) (19).
These datalead to two possible models for interactions between Tat and the TAR. The first is that Tat is a very compact protein and that any changes in the activation
domaindisrupt the secondarystructureof the entireprotein (23). In this scenario, basic residues would assume new
conformations with different mutations in the activation domain, which could interfere with the binding of Tat to TAR RNA. However, given all the in vitro binding studies with different lengths and compositions of these basic amino acids, this possibility appears unlikely (1, 2, 6, 31, 32). Alternatively, acellularproteinorcomplexofproteinsthat
interacts with the activation domainof Tat is also required for the specific binding of Tat toTAR RNA (Fig. 4). This
"coactivator" couldperformtwofunctions, activating tran-scriptionandbinding toTAR RNA. It isveryattractiveto consider thepossibilitythat thisproteinbindstotheloopof the TAR,which is absolutely required for trans-activation
by Tat in vivo but isdispensable for bindingof Tat toTAR U A
CG UG IJ A C
U A GC GC
610000040
****
00
000
a
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.155.476.76.428.2]RNA invitro. This protein isunlikely to beTRP-1/TRP-185, whichby binding to theloop displaces Tat from the TAR in vitro (27, 33). Furthermore, there is precedence for such a
protein in Escherichia coli.NusA, which is a bacterial protein that is required forantitermination ofbacteriophageX tran-scriptionby the N protein,notonly increases thebindingto
the Nutilization site (Nut B), which forms an RNA
stem-loop, but also conveys the activation by N to core RNA polymerase (20,21). Thus, these studies support the existence ofaprotein or acomplexofproteins thatinteractwith Tat, the TAR, and thetranscription complex (Fig. 4). The identifica-tion,cloning,characterization,andexpression of this protein
arenecessary next stepsin the research ofTat.
ACKNOWLEDGMENTS
We thank Michael Armanini for expert secretarial assistance,
David McDonald forgiftsofplasmids, and members ofour labora-tories forinstructive criticism.
Ying Luo issupportedbyafellowshipgrantfrom theCalifornia
UniversitywideAIDS ResearchProgram.
REFERENCES
1. Calnan, B.J., S. Biancalana, D. Hudson, and A. D. Frankel. 1991. Analysis of arginine-rich peptides from the HIV Tat protein reveals unusual features of RNA-protein recognition. GenesDev. 5:201-210.
2. Calnan, B.J.,B. Tidor, S.Biancalana, D.Hudson, andA. D. Frankel. 1991. Arginine-mediated RNArecognition: the argi-nine fork. Science 252:1167-1171.
3. Cullen, B. R. 1991. Regulation of human immunodeficiency virus replication.Annu. Rev. Microbiol.45:219-250.
4. Dayton, A. I., J. G. Sodroski, C. A. Rosen, W.C. Goh, and W. A. Haseltine. 1986. The trans-activatorgeneofthehumanT celllymphotropicvirustypeIIIisrequiredforreplication.Cell 44:941-947.
5. Delling, U., S. Roy,S. M. Sumner, R.Barnett,L. Reid,C. A. Rosen, and N. Sonenberg. 1991. The number of positively charged amino acids in the basic domain ofTatis critical for trans-activation andcomplex formation withTARRNA. Proc. Natl. Acad.Sci.USA 88:6234-6238.
6. Dingwall, C.,I.Ernberg,M.J.Gait,S. M.Green,S.Heaphy, J. Karn, A. D. Lowe, M.Singh, M. A. Skinner, and R. Valerio. 1989.Humanimmunodeficiencyvirus 1 Tatprotein binds
trans-activation-responsive region (TAR)RNA in vitro. Proc. Natl. Acad. Sci.USA86:6925-6929.
7. Felber, B. K.,C. M. Hadzopoulou,C.Cladaras, T. Copeland, and G. N. Pavlakis. 1989. Revprotein ofhuman
immunodefi-ciencyvirustype1affects thestabilityandtransportof the viral mRNA.Proc. Natl.Acad. Sci. USA 86:1495-1499.
8. Fisher,A. G., M. B. Feinberg, S. F. Josephs, M. E. Harper,
L.M.Marselle, G. Reyes,M.A.Gonda,A.Aldovini,C.Debouk, R.C.Gallo,and F.Wong-Staal.1986. The trans-activator gene ofHTLV-III is essential forvirusreplicationNature(London)
320:367-371.
9. Frankel,A. D. 1992. Activation of HIVtranscription by Tat. Curr.Opin. Genet. Dev.2:293-298.
10. Frankel,A. D., S.Biancalana,and D.Hudson.1989.Activity of
syntheticpeptides from theTatproteinof human
immunodefi-ciencyvirustype 1. Proc.Natl. Acad.Sci.USA 86:7397-7401. 11. Graham, G. J., andJ. J. Maio. 1990. RNAtranscripts of the human immunodeficiency virus trans-activation response
ele-ment caninhibitaction of the viral trans-activator. Proc. Natl. Acad. Sci. USA87:5817-5821.
12. Hope, T.J., X. J. Huang, D. McDonald, and T. G. Parslow. 1990. Steroid-receptorfusion of the human immunodeficiency
virustype 1Revtransactivator:mapping crypticfunctions of the arginine-rich motif. Proc. Natl.Acad. Sci. USA87:7787-7791. 13. Huet, T., M. C. Dazza,V. F. Brun, G. E. Roelants, and S.
Wain-Hobson. 1989. A highly defective HIV-1 strain isolated from a healthy Gabonese individual presenting an atypical
Western blot. AIDS 3:707-715.
14. Kamine, J.,T. Subramanian, and G. Chinnadurai. 1991. Spl-dependentactivation ofasynthetic promoter byhuman immu-nodeficiency virustype 1 Tatprotein. Proc. Natl. Acad. Sci. USA 88:8510-8514.
15. Karn, J.1991. Control ofhumanimmunodeficiencyvirus repli-cation by the tat, rev, nefand protease genes. Curr. Opin. Immunol.3:526-536.
16. Kuppuswamy, M., T. Subramanian, A. Srinivasan, and G. Chinnadurai. 1989. Multiple functional domains of Tat, the trans-activator of HIV-1, defined by mutationalanalysis. Nu-cleic AcidsRes. 17:3551-3561.
17. Malim, M. H., S. Bohnlein, J. Hauber, and B. R. Cullen. 1989. Functional dissection of the HIV-1 Rev trans-activator--deri-vation of a trans-dominant repressor of Rev function. Cell 58:205-214.
18. Malim, M. H., and B. R. Cullen. 1991. HIV-1 structural gene expression requires thebinding ofmultiple Rev monomers to theviral RRE:implicationsfor HIV-1latency.Cell 65:241-248. 19. Malim, M. H., J. Hauber, S.-Y. Le, J. V. Maizel, and B. R. Cullen. 1989. The HIV-1 Rev trans-activator acts through a structured target sequence to activate nuclear export of un-splicedviralmRNA. Nature(London)338:254-257.
20. Mason, S. W., and J. Greenblatt. 1991.Assemblyof transcrip-tion elongatranscrip-tion complexes containing the N protein of phage lambda and the Escherichia coli elongation factors NusA, NusB, NusG, andS10. Genes Dev.5:1504-1512.
21. Mason, S. W., J. Li, and J. Greenblatt. 1992. Host factor requirements for processiveantitermination of transcription and suppression of pausing by the N protein of bacteriophage lambda. J. Biol. Chem. 267:19418-19426.
22. McDonald, D., T. J. Hope, and T. G. Parslow. 1992. Posttran-scriptionalregulation by thehumanimmunodeficiency virustype 1 Rev and human T-cell leukemia virus type I Rex proteins throughaheterologousRNAbinding site. J. Virol. 66:7232-7238. 23. Rice, A. P., and F. Carlotti. 1990. Structural analysis of wild-type and mutant human immunodeficiency virus type 1 Tat proteins. J. Virol.64:6018-6026.
24. Rosen, C. A., and G. N.Paviakis. 1990.TatandRev: positive regulatorsofHIVgeneexpression. AIDS 4:499-509.
25. Selby, M. J., E. S. Bain, P. A. Luciw, and B. M. Peterlin. 1989. Structure, sequence, and position of the stem-loop in TAR determinetranscriptional elongation byTatthrough theHIV-1 longterminal repeat. Genes Dev.3:547-558.
26. Selby, M. J., and B. M.Peterlin.1990.trans-activationby HIV-1 Tat via aheterologousRNAbinding protein. Cell62:769-776. 27. Sheline, C. T., L. H. Milocco, and K. A. Jones. 1991. Two
distinct nuclear transcription factors recognize loopandbulge residuesof the HIV-1 TAR RNAhairpin. GenesDev. 5:2508-2520.
28. Subramanian, T., R. Govindarajan,andG. Chinnadurai. 1991. Heterologous basic domain substitutions in the HIV-1 Tat protein reveal and arginine-rich motif required for transactiva-tion. EMBO J. 10:2311-2318.
29. Tiley, L.S., S. J.Madore, M. H. Malim, and B. R. Cullen. 1992. The VP16 transcription activation domain is functional when targeted to a promoter-proximal RNA sequence. Genes Dev. 6:2077-2087.
30. Venkatesan, S., S. M.Gerstberger, H.Park,S. M.Holland, and Y.-S. Nam. 1992. Human immunodeficiency virus type 1 Rev activation can be achieved without Rev-responsive element RNAif Revisdirectedtothetarget as aRev/MS2fusionprotein which tethers theMS2 operator RNA. J. Virol.66:7469-7480. 31. Weeks, K. M., C. Ampe, S. C.Schultz,T.A. Steitz,and D.M.
Crothers. 1990.Fragmentsof the HIV-1 Tatproteinspecifically bind TAR RNA. Science249:1281-1285.
32. Weeks, K. M., and D. M. Crothers. 1991. RNArecognitionby Tat-derived peptides: interaction in the major groove? Cell 66:577-588.
33. Wu, F., J. Garcia, D. Sigman, and R. Gaynor. 1991. Tat regulates binding of the human immunodeficiency virus
trans-activating region RNA loop-binding protein TRP-185. Genes Dev. 5:2128-2140.