0022-538X/94/$04.00+0
CopyrightC) 1994,AmericanSocietyforMicrobiology
Differences
in
CD4
Dependence
for
Infectivity
of
Laboratory-Adapted
and
Primary
Patient Isolates of
Human
Immunodeficiency
Virus
Type
1
DAVIDKABAT,I* SUSAN L.
KOZAK,'
KATHYWEHRLY,2 ANDBRUCE CHESEBRO2 Departmentof Biochemistry and MolecularBiology, Oregon Health SciencesUniversity, Portland, Oregon97201-3098,1 andLaboratoryofPersistent ViralDiseases, National InstitutesofAllergyand InfectiousDiseases, Rocky MountainLaboratories, Hamilton, Montana598402
Received 15 October1993/Accepted 12January 1994
CD4 is known tobeanimportant receptor forhumanimmunodeficiencyvirustype1 (HIV-1)infectionof T
lymphocytesand macrophages.However,thelimiting stepsinCD4-dependentHIV-1 infectionsinvivoand in vitro are poorlyunderstood. To address thisissue,weproducedapanel of HeLa-CD4cellclones that express widelydifferent amounts of CD4 andquantitatively analyzedtheir infectionbylaboratory-adaptedandprimary patientHIV-1isolates.ForallHIV-1isolates,adsorptionfrom themedium ontoHeLa-CD4 cellswasinefficient and appeared to be limiting for infection in the conditions of our assays. Adsorption of HIV-1 onto CD4-positive peripheralblood mononuclear cells was alsoinefficient.Moreover,therewas astrikingdifference between laboratory-adapted and primary T-cell-tropic HIV-1 isolates in the infectivity titers detected on different HeLa-CD4 cells. Laboratory-adapted HIV-1 isolates infected all HeLa-CD4 cell clones with equal efficiencies regardless of the levels ofCD4,whereas primary HIV-1 isolates infected these clones in direct proportion tocellularCD4expression.Our interpretationis that forlaboratory-adapted isolates,abarrier step that preceedsCD4encounter waslimitingandthesubsequentCD4-dependentvirus capture process washighly efficient, even at very low cell surface concentrations. In contrast, for primary HIV-1 isolates, the CD4-dependent steps appeared to be much less efficient. We conclude that
primary
isolates of HIV-1 infectinefficiently
followingcontact with surfaces ofCD4-positive cells,and we proposethat thisconfersaselectivedisadvantage during passage in rapidly dividing leukemia cell lines. Conversely, in vivo selective pressure appears to favor HIV-1 strainsthatrequire largeamounts ofCD4 for infection.
Although CD4 is the principal cell surface receptor for humanimmunodeficiency virus type1(HIV-1),thepathwayby which it mediates infectionisnotfully understood (8,26,51). It is believed that HIV-1 binding to CD4 initiatesa series of steps that culminate in the fusion of viral and plasma
mem-branes,resulting in viral core transfer into the cytosol(28,30,
47). Bindingonto CD4 weakens associationbetween the viral gpl20 surface glycoprotein and itsgp4l transmembrane sub-unit(33-35,51), therebyexposingpreviouslyburiedregionsof gpl20 and gp4l (34, 42, 51) and enabling the gp4l amino-terminal fusion peptide to contact the plasma membrane.
CD4-dependent loosening ofgpl2O-gp4l subunitassociations apparently enables other specific cellular factors to interact with the virus-CD4 complexin amanner that isconducive to themembrane fusionprocess (2, 4, 9, 13, 15, 23).
A difficulty in understanding CD4-mediated infection de-rives from the enormous diversity of HIV-1 isolates and from differences in their abilities to infect different CD4-positive cells suchasmacrophages and Tlymphocytes (12, 16, 41,45, 51). Primary patient isolates ofHIV-1 infect normal
prolifer-ating CD4-positive T lymphocytes but not certain leukemic CD4-positive T-cell lines such as H9 orCEM, whereas labo-ratory-adapted (LA) viruses replicate in these immortalized leukemiccells(8,12,51). LAvirusisolatesarealso much more
easilyinactivated by soluble CD4 (sCD4) (3, 7, 14,21,22, 33,
34,37, 48, 49).This evidence suggests that different isolatesof
*Correspondingauthor. Mailing address: Department of Biochem-istryand Molecular Biology L224, Oregon Health Sciences University,
Portland,OR97201-3098. Phone: (503) 494-8442. Fax: (503) 494-8393.
Electronicmailaddress:[email protected].
HIV-1 interactdifferently withCD4 andpossiblywithcellular accessoryfactors.
Amajorgap inunderstanding CD4-mediated HIV-1 infec-tions and viraladaptationsandtropismsderives from a lackof informationconcerning the steps of the infectionpathway that are limiting in cell culture conditions. Knowledge of the limiting stepsof infectionmightlead tomore-focusedefforts at drugdevelopment, because inhibitingthese steps would have relatively large effects on viral replication. For the present experiments, we produced a panel of HeLa-CD4 cell clones that expresswidelydifferent amounts ofCD4,and we quanti-tatively analyzed their infection by different HIV-1 isolates. CD4-positiveHeLa cells were used for this work becausethey aresusceptibletobothprimaryandLAT-lymphotropic HIV-1 isolates (10-12). Our results suggest that for both types of viruses, adsorptionontothe cells isinefficientinpartbecause of a limitation that precedes encounter of virus with CD4. After thatstep, LAviruses efficiently infectcells even at very low cell surface CD4 concentrations, whereas infectivity of primary isolates is strongly dependent on the level of CD4 expression.
MATERUILSAND METHODS
Cells and viruses. Cell lines were grown in Dulbecco's modified Eagle's medium with 10% fetal bovine serum. HeLa cells(20)wereobtainedfromRobinWeiss(Institute of Cancer Research, London, England). Retroviral packaging cell lines t2 (31, 32) were used for retroviral vector amplification as previously described (6, 50). To construct the pSFF-CD4 vector,the 3.0-kbp EcoRI-EcoRI fragment of T4-pMV7 (30) 2570
on November 9, 2019 by guest
http://jvi.asm.org/
containing the entire CD4 cDNA was subcloned into the EcoRI site of pUC 1813 (25). The 1.8-kbp BamHI-BamHI
CD4-encoding fragment was then cloned into the BamHI site of pSFF (6) to obtain pSFF-CD4. This construction removed the transcription termination [poly(A) addition] signal from the CD4 cDNA, thus ensuring relatively efficient function of pSFF-CD4 as a retroviral vector. The resulting CD4-encoding virions (6, 50) were used to infect HeLa cells, and HeLa-CD4 clones were then isolated by limiting dilution cloning. Other HeLa-CD4 cell clones were previously described (9, 10). HIV-gpt virions were prepared by cotransfecting Cos-7 cells with pHIV-gpt and pHXB2-env in the presence of 10
,uM
chloroquine as described elsewhere (38, 39). Unless otherwise stated, assays of HIV-gpt infection were done on cells plated at 2 x 105 cells per 25-cm2 dish 24 h previously which were treated with 8 ,ug of Polybrene (Sigma, St. Louis, Mo.) per ml for 30 min before infection. A 2-ml volume of virus-containing culture medium was incubated with the cells at 37°Cfor 2 h, and the cultures were maintained for 48 to 72 h before trypsinizing and replating at 6 x 105 cells per 100-mm dish. The medium was replaced 24 h later with growth medium supplemented with 250 ,ug of xanthine, 3 ,ug of hypoxanthine, 4,ugof thymidine, 10 ,ug of glycine, and 150,ug of glutamine per ml, 10 mM HEPES (N-2-hydroxyethylpiperazine-N'-2 ethanesulfonic acid; pH 7.4), and 40 ,ug of mycophenolic acid perml. Drug-resistant colonies were stained with 0.1% tolui-dine blue in 30% ethanol 10 to 14 days later. Efficiencies of infection were increased approximately twofold by Polybrene pretreatment of the cells, but the Polybrene effect was the same for all HeLa-CD4 cell clones (results not shown). Rep-lication-competent HIV-1 LA isolates NL4-3 and Ala (9) and primary patient isolates (11) were assayed on the HeLa-CD4 cell clones by the focal infectivity assay method (10). Primary HIV-1 isolates were made as previously described (11). Virus was amplified by passage of undiluted culture supernatant in fresh phytohemagglutinin (PHA)-stimulated peripheral blood mononuclear cells. Stocks were made from supernatant fluid taken at days 7 to 10 at the time of peak reverse transcriptase activity. Stocks used in these experiments were made at the following passages: 208K10 and 208K2 2nd passage; 13956 and 208K4, 3rd passage; 13231, 5thpassage; and 14558, 7th passage
(two of these passages were done at end point dilutions). CD4 expression assays. CD4 expression on cells was de-tected by incubating the cells with a CD4-specific monoclonal antibody, ADP318 (kindly provided by Paul Clapham and RobinWeiss, Institute of Cancer Research) (4 ,ug/ml), at 37°C for 1 h. The cells were washed with medium, incubated with a 1:500dilution of rabbit anti-mouseimmunoglobulin G-fluores-ceinisothiocyanate(Tago, Inc., Burlingame, Calif.) at 37°C for 1 h, and then they were washed with medium, fixed in cold 100% methanol, and mounted for observation by immunoflu-orescence microscopy. Quantitative CD4 analysis employed
the same CD4 monoclonal antibody diluted 1:500 and incu-bated with confluent 9-cm2 monolayers for 1 h at 37°C. The cells were rinsed with culture medium several times and were
then incubated inmedium with
[1251I]protein
A (0.2 ,uCi/ml, 2to 10
jiCi/jig;
DuPont NEN Research Products, Boston,Mass.) for 1 h. The cells were then washed for 5 min three times with fresh medium. The cells were dissolved in 1 ml of 0.1 NNaOH andwere counted inagamma counter. Aliquots wereassayed forprotein concentrations by the Coomassie dye method (Bio-Rad Laboratories, Richmond, Calif.).
Cell clones were also analyzed for CD4 by quantitative
fluorescence-activated cell sorting (FACS) as previously de-scribed (9). SIM.4 monoclonal anti-CD4 antibody was ob-tained from the National Institute of Allergy and Infectious
Diseases AIDS Research and Reference Program, Rockville, Md., and was used instead of OKT4A as the first antibody for indirect immunofluorescence. The second antibody was fluo-rescein isothiocyanate-conjugated goat anti-mouse immuno-globulin (Organon Teknika Corp, West Chester, Pa.).
RESULTS
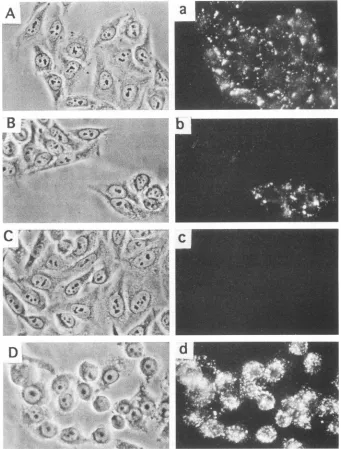
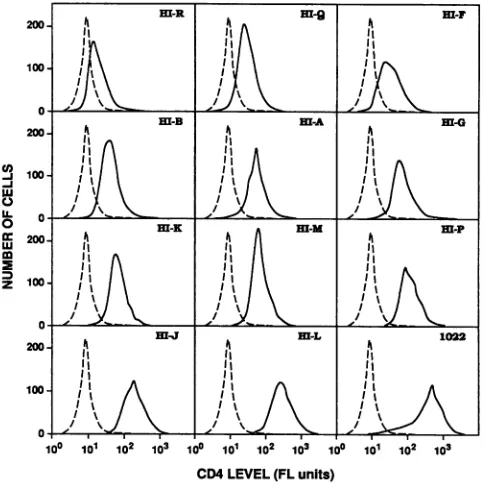
HeLa-CD4 cell clones with different quantities of CD4. Retroviral vector pSFF-CD4 was amplified and expressed in packaging cells (6,50), resulting in high titers of CD4-encoding amphotropic host range virions. Infections of HeLa cells resulted in CD4 synthesis by nearly all cells in the cultures. As seen by two-stage immunofluorescence microscopy of viable cells (see Materials and Methods), certain clones had abun-dant cell surface CD4, whereas others had trace amounts (Fig. 1). For the clones that expressed the least amount, the immunofluorescent clusters of CD4 were tiny and few, and they occupied only a minute proportion of the cell surfaces (Fig. lb, upper clone). The quantity of cell surface CD4 expression in each clone was analyzed by FACS analysis (Fig. 2). Mean fluorescence intensity results demonstrated a wide range of CD4 expression (Table 1). Similar relative estimates of CD4 quantities were obtained by the
[125I]protein
A binding method (50) (results not shown).Infection of HeLa-CD4 cells by a molecularly cloned LA
HIV-1
(HIV-gpt).
CD4 expression was more homogeneous in each clonal cell population produced by the pSFF-CD4 vector (Fig. 2) than in HeLa-CD4 clones previously prepared with the T4 pMV7 retroviral vector (9, 30). Therefore, the new clones were used for infectivity studies. Initially, we studied infection by the HIV-gpt method (38, 39). Replication-defective HIV-gpt virions contain an RNA genome encoding the gpt gene for resistance to mycophenolic acid in virions that contain gpl2O-gp4l complexes encoded by the LA HIV-1 molecular clone HXB2 (18). Infected cells grow as colonies in a mycophenolic acid-supplemented selection medium. The virus was incubated with 2 x 105cells in standard conditions for 2 h at37°C.
At the low multiplicities of infection that occur by this assay (multi-plicity of infection of approximately 0.005) (38), the numbers of colonies were directly proportional to virus concentrations (Fig. 3A) and were the same, within experimental error, for HeLa-CD4 cells with widely differing amounts of CD4 (Fig. 3A and Table 1). Thus, cells with trace amounts of CD4 were infected as efficiently as cells with large amounts.If the virus was completely adsorbed even by cells with little CD4, equal infections of the high- and low-expresser cell clones would be anticipated. However, evidence indicated that adsorption of HIV-gpt onto HeLa-CD4 cells was inefficient under the conditions of our assays. First, the number of infections was directly proportional to the number of HeLa-CD4 cells plated in the cultures and was the same for high (clone HI-L)- and low (clone HI-B)-CD4 expressers (Fig.3B). If 2 x 105cells of either clone were able to completely adsorb the virus, addition of more cells would not give more infec-tions. Thus, the equal infection of clone HI-L and HI-B cells was not a consequence of using a limiting amount of HIV-gpt. Second,virus-containing culture medium that had been prein-cubated for 2 h with HeLa-CD4 cultures gave a titer that was approximately equal to 60% of the initial titer when assayed on a second culture (Table 2). Furthermore, the approximately 40%loss of infectious HIV-gpt during the first incubations was causedby spontaneous inactivation rather than byadsorption, because it occurred in the absence of cells or in the presence of CD4-negative control HeLa cells (Table 2). These results
imply that an undetectably small percentage of the HIV-gpt
on November 9, 2019 by guest
http://jvi.asm.org/
FIG. 1. CD4expressionbyHeLa-CD4cells. HeLa cellswereinfectedwith the amphotropic pseudotypeofthe SFF-CD4retrovirus,and the cells were thenpassaged into fresh cultures. Thephotographsshow small colonies ofthe cells inphasecontrast(leftpanels)andcorresponding CD4-specificimmunofluorescence (rightpanels). (Aanda)Cell clone with mediumexpressionofCD4;(Bandb)twocellclones,onewithmedium expressionof CD4 and theother with low butsignificant expressionofCD4;(Candc)control HeLa cellslackinganyCD4;(Dandd)clonedHI-L
cells,which express ahigh amountof CD4. CD4expression appeared uniform within cells of each clone.
was adsorbed onto HeLa-CD4 cells during our standard 2-h incubations at 37°C. Third, HIV-gpt titers reproducibly in-creasedwhen adsorption timeswereextendedbeyond2 h. For example, in oneexperiment, the titers ofHI-Lcellswere598, 1,360, and 1,620 colonies after2-, 4-,and8-hadsorption times, respectively.
The results described above suggest that HIV-gpt infected HeLa-CD4 cells by an inefficient adsorption-limited process under the conditions of our assays. Paradoxically, however, raising CD4 levels above a low threshold had no effect on infections. To stringently test this conclusion, we mixed low
(HI-B)- and high (HI-L)-expresser HeLa-CD4 cells in a 1:1 proportion, infected them with HIV-gpt under our standard
conditions, and obtained approximately 500infected colonies afterselection withmycophenolic acid-containingculture me-dium. The infected cellswerethentrypsinizedinapool,plated
on coverslips, and labeled for CD4-specific immunofluores-cence.The ratioof HI-BcellstoHI-Lcellswasthesamewithin experimental error for the infected and selected (0.87 + 0.085), uninfected (0.96 ± 0.10), and mock-infected (0.94 + 0.10) cell populations. The errors in these numbers are the standarddeviations of the ratios. Therefore,HeLa-CD4cells
expressing low or high levels of CD4 were equally infected within the same culture. All of these results imply that the limiting step in infection of our HeLa-CD4 cell clones by
HIV-gptwasindependentof the level of CD4 expression.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.132.473.81.530.2]100
0 r-K Hn-M rn-P
C2C200tft
2100-0
Wj , < , A Wr-L
200 lo 102 103 10° 10f102 103 10° lo, 103
CW4LEVEL(FL units)
FIG. 2. FACS analysis of CD4 expression in HeLa-CD4 clones. Solid lineswereobtained with anti-CD4 monoclonalantibodySim.4as
the firstantibody. Dashed lines Werenegative controls withnofirst
antibody. The second antibody in both cases was FITC-conj'ugated
goatanti-mouse immunoglobulin.FL, fluorescence.
Infection ofHeLa-CD4 cell clones
by
replication-competent
IA and
'primary
HIV-1 isolates. Apreviouis study
of three HeLa-CD4 cell clones hadsuggested
that HIV-1 titersmight
correlate
positively
withC1D4 expression
levels(9).
Toresolve differences thatmight
deperid
on HIV-1 strains or on assayconditions,
we used thequantitative
focalinfectivity
assay tostudy
infectionof all ofourHeLa-CD4 cell clonesby
variousprimary
and IA HIV-1 isolates(Fig.
4).
The LA HIV-1 isolatesNILA-3 and Ala behaved in a manneridenticalto that of theHIV-gpt
described above.Only
the lowest C1D4 ex-presser, cloneiC,
showed a reduced titer withNL4A-3,
as describedpreviously (9).
Above this low threshold of CD4expression,
titersdidnotsignificantly depend
onCD4levels in the HeLa-CD4 clones. Incont'rast,
titers of theprimary
HIV-1 isolates13956,
208K10,
14558,
208K2,
a'nd208K4 differedby
2 to3 orders ofmagnitude
thatwerehighly
correlated withC1D4expression
levels. The data with oheprimary
isolate(13231)
wereintermnediate betw'een thesetwo
patterns
andwerec'on-sequently
difficulttointerpret.
Itis conceivable that thisisolate containednmultiple
HIV-1 clones with both LA andprimary
isolate
phenotypes.
We also studied
adsorption
ofreplication-competent
ILA(NL4A-3)
andprimary
(208K2,
208K4, and208K10)
HIV-1 isolates onto cellsby
using
the methods described above in Table 2 forHIV-gpt.
The resultsclearly
indicated that virusadsorptions
in culture 'wereinefficient,
and noCD4-specific
depletion
of viralinfectivity
fromthesupernatant
fluid couldbe detected
(data
notshown). Lack of sufficientcellnumbers in theseabsorption experiments
might
explain
ourfailure toseesignificant depletion
of virus from the culture media.There-fore,
weused 10- and 100-foldmoreCD4-positive
clone 1022 [image:4.612.65.307.81.324.2]cellsa's wellasPHA-stimulated
CD4-positive peripheral
blood mononuclear cells(PBMC)
inanadditionalabsorption
exper-iment.Again,
however, nodepletion
ofinfectivity
could beTABLE 1. Equal susceptibilities of HeLa-CD4 clones toinfectionby HIV-gpt
Celllinea CD4level' HIV-gptinfections
(colonies/dish)c
Assay 1 Assay2
HeLa 0 0 0
HI-R 14 702 341
HI-Q 25 403 276
HI-F 30 936 503
HI-B 34 545 454
HI-A 52 838 318
HI-K 71 873 458
HI-G 71 852 592
HI-Md 75 492 295
HI-P 128 716 487
HI-J 234 662 371
HI-L 300 737, 598, ands51e 256
aThe cellswere either controlHeLa cells, which lack CD4, or clones of HeLa-CD4cells.
bMeanfluorescence intensity inarbitraryrelative units was determined by
FACS analyses (9) for each population.Abackgroundvalue of 11 was found for
normal HeLa cells, and thisnumberwassubtracted from thevalue foreach populationtogive the valuesshown. By aradioactivityassay for cell surface CD4
(50), theH9line of standard CD4-positive Tlymphoblasts (40),which have approximately1.7 x I04CD4 moleculesper cell(27),wasfound to have an amountofCD4 percell similartothat of our cloneHI-Qcells.
CVirusfor the HIV-gptassays waspreparedfromCos-7cells cotransfected
with pHIV-gpt and pHXB2-env in the presence of 10 R1M chloroquine as
described in Materials and Methods. The results are the numbers of drug-resistant coloniesonthe 100-mm-diameterdishes.
dHI-M grew moreslowlythan HeLa cells and the otherCD4-HeLaclones.
'The numbers listed inseries werereplicateassays.
demonstrated (Fig.5). These results indicated that as a result
of the inefficient
absorption
process, the amounts of HIV-1 infectivity detected on both HeLa-CD4 cells and PBMC were verylow percentagesof the totalinfectivity in the viral stock.DISCUSSION
Theresults of the presentexperimentsindicatethat primary and LA HIV-1isolates differ intheir interactionswithcellular CD4duringthe infection process. Infectivitytiters ofprimary
isolates weredirectly influenced by the level of CD4 expressed on HeLa cell clones (Fig. 4). This result implies that the
virus-CD4
interaction isacritical limiting step for infection ofthesecellsby primaryHIV-1 isolates,which is consistent with
the
important
role of CD4as areceptorfor HIV-1.Incontrast,infectivity titers of LA HIV-1 isolates were not significantly
influencedby the level of CD4 expressed on thesesametarget cell populations, and it was unclear from this result what
exactlywas the limiting step for infection by the LA HIV-1 isolates. However, several lines of evidence suggest that
ad-sorption ofLAviruses from the medium onto the cells is a
limitingprocess ininfection. Duringthe infection step under the conditions of our assays, infectious HIV-1 was removed very inefficientlyfromthe medium (Table 2andFig. 5). This
couldnotbe dueto asaturation of receptorsbyvirusor free
gpl20, because infections occurred at low multiplicities and virustitersincreaseddirectlyin
proportion
tovirus concentra-tions(e.g., Fig. 3).Themostlikely
meansfor theconcentrationof virus in the mediumtohavea
directly
proportional
influence oninfectionofHeLa-CD4cellsisduring
the step ofadsorption
onto the cells
(1,
5,24,29,
53).
Ifinfectionswere limitedby
saturation or depletion of factors atany
postadsorptive
step, increases in the virusconcentrations would not increase the titers obtainedinourassays.Thus,
weinterpret
ourresultsto suggestthatadsorption
of HIV-1 onto the cells increases inon November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.325.564.100.245.2]1200
a
0 0
IL
800
400
0 0.2 0.6 1
B
* HI-L
* --U--Hi-B ,
A HeLa
0,
A 2 3 eeAe A A
1 2 3 4 5 Relative Virus Concentratlon Cell Number (10 5)
FIG. 3. HIV-gpt infection is proportionaltovirusconcentration and cell concentration. (A) Virus-containingmediumwasseriallydiluted 1:2 with mock-transfected Cos-7-conditioned medium,andthe dilutionwasusedtoinfect cells inourHIV-gptassayasdescribedinMaterialsand Methods. The number of drug-resistant colonies ofhigh-expressing CD4-HeLa HI-L clone and oflow-expressing CD4-HeLa clonesHI-Band HI-Ris proportionaltothe relative virus concentrationand is 0intheCD4-negativeHeLa cells. Each datumpointis theaverageofduplicate
assays.(B) HeLa cells and CD4-HeLa clonesHI-Land HI-Bwereplatedatvarious concentrationsin25-cm2 dishesandwereinfectedwithour
HIV-gpt. The infected culturesweretrypsinized48 hlater,andcellsequaltothe number of cellsatinfectionwere platedin100-mm-diameter dishes andweresubjectedtoourmycophenolicacid-supplementedselection medium(seeMaterials andMethods). Drug-resistantcolonieswere
stainedand counted after 10days.
direct proportiontotheconcentration of virus in the medium and that this results in a corresponding increase in virus titers in our infectivity assays. In this circumstance of
adsorption-limitedinfection, we anticipated that infection woulddepend
on the concentration ofadsorptive sites on the cell surfaces. For primary HIV-1 isolates, these limiting adsorptive sites appeared to be CD4. However, for LA HIV-1 isolates,
infec-tionrequired CD4-positivetarget cells butwas not limitedby
CD4.
These results areconsistent withamultistagemodelof virus attachment and entry into cells. First, virusmustdiffusetothe cellsurfaces prior topotential encounter with CD4. Diffusion oflarge substances suchasviruses is slow andcanbelimiting
for some infections (1, 5, 24, 29, 43). Other steps preceding
potential interaction with CD4 could involve barrier or pre-requisite reactions such as passage through the exoskeletal meshwork, release ofhydration, and the overcoming of elec-trostaticrepulsion(53).Inaddition,theremightbe features of thecell surfaces such as weakbindinginteractions that entrap HIV-1 andhold it long enough for subsequent efficient asso-ciation with CD4. Analogously, attachment onto accessory cell surface components facilitates adsorption of herpes simplex
TABLE 2. Inefficientadsorptionof HIV-gpt onto HeLa-CD4cellsat37°C
Cells used for Firstinfection Cells used for Second infection first infectiona colonies/dish infection colonies/dish
None HI-L 887
HeLa 0 HI-L 812
HI-B 1,401 HI-L 860
HI-R 2,210 HI-L 1,000
HI-L 1,770 HI-L 923
aA2-mlvolumeofvirus-containing mediumwas incubatedwiththecellsfor
2h at37°CasdescribedinMaterialsand Methods.Afterthefirst infections,the samemediumwastransferred for2 hontoculturesthat containedHI-Lcellsthat
hadbeenplated and treated in a manner identicalto thatused forthefirst
infections.The same medium was usedforeachround.Eachrow indicatesthe samemedium.Thus,themedium preincubatedinanemptyflaskandlaterused toinfectHI-Lcellsresultedin887drug-resistantcolonies afterselection.
virustype1 (19, 44). Second, the virusmust encounterCD4in
asterically appropriatemannerand formareversibleunivalent encounter complex (29, 43). Third, other CD4 molecules presumably interact with the virus to form a multivalent
irreversiblecapturecomplex (29,43) that is committedto enter postadsorptionsteps ofthe infectionpathway.
From theperspective ofourresults withLA HIV-1isolates,
the stepspreceding CD4 encounterwould obviously be inde-pendent of CD4 expression levels. Moreover, if the CD4 receptordensitywereaboveacertainlowthreshold, formation ofencountercomplexes and their conversion into irreversible capture complexes could also behighly efficient, as has been demonstrated experimentally for phage lambda adsorption ontoEscherichia coli (5, 43). Efficient bindingto CD4 could alsoensue ifthevirus were held evenbriefly byan accessory
factoron the cell surfaces. Thus,our resultswith LA HIV-1 isolates suggest that the preencounter reactions are slowbut
that capturecomplexes thenform efficientlyeven atvery low
cell surface densities of CD4. In contrast, our results with
primary isolates of HIV-1 suggest that their attachment to CD4must be relatively inefficient. Consequently,we propose
thatprimary HIV-1 isolates formencountercomplexes with a
lowefficiency that is highly dependenton CD4concentration
and that these encounter complexes might also frequently dissociatewithout conversiontocapturecomplexes.
Thebiochemicalexplanationforthesedifferences is unclear; however, some clues maybe provided by previous studies of
interactions of HIV-1 with sCD4.Although sCD4 binds with approximately equal affinitiesto gpl20 purified fromLAand primary virus isolates, recent evidence suggests that sCD4 binds to gpl20 of intact virions with lower affinity in primary isolatesthanin LAisolates(22, 34). Furthermore, LAHIV-1 isolatesareinactivated byexposuretoconcentrations of sCD4 that are 100- to 1,000-fold lower than those required to inactivate primary isolates (14, 21, 33). The kinetics of virus bindingandentryintoCD4-positiveTcellsappearstobemore
rapid forLAthan forprimary isolates (17, 46). These
differ-ences cannot be ascribed to amounts of gpl20 per virion,
because LA viruses spontaneously shed gpl20 and have a
relatively low gpl20 content (52). These observationson the A
1200 a 0 0
o
800
400
0
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.132.485.80.233.2]5.0-
4.0-i
3.0-cm
-Cs
_ 2.0-0
O
--
5.0-> 4.0-Ic
3.0-2.0- 1 I
1.0
-1.0 2.0 3.0 1.0 2.0 3.0 1.0 2.0 3.0 1.0 2.0 3.0
CD4
Expression(log mean FL)FIG. 4. Comparison of HIV-1 titer versus CD4 expression using two LA HIV-1 isolates (NL4-3 and ALA) and six primary AIDS
patient-derivedHIV-1isolateson aseries of HeLa-CD4clones. The individual clones can be identified by their CD4 expression expressed as the
logof themeanfluorescence units(FL)onthe horizontal axis: clone 1C, 0.7; clone 6C, 0.9; HI-R, 1.15; HI-Q, 1.40; HI-F, 1.48; HI-B, 1.53; HI-A,
1.72;HI-G, 1.85; HI-K, 1.85; HI-M, 1.87; HI-P, 2.11; HI-J, 2.37; HI-L, 2.48; clone 1022, 2.68. Error bars indicate standard errors of geometric meansbasedonfoci counted at several different dilutions. The experiment was repeated three times with similar results. The solid lines are
least-squarebest fitstothe datum pointsfor all clones, except the two with the lowest CD4 contents; the latter two datum points are indicated by opencircles.This exclusionwasdone becausethesepoints deviatesomewhat from the curves in a manner consistent with the fact that infections are0for controlHeLacells, and because these cell clones haverelatively heterogeneous CD4 expression on cells of the clonal populations. The
correlation coefficientsfor these computer-drawn best-fit lineswere asfollows:Ala, 0.13; NL4-3,0.25; 208K2, 0.92; 208K4, 0.89; 14588, 0.84;
208K10, 0.81; 13956, 0.84; and 13231, 0.42. Thisstronglysupportsthe conclusionthat titers correlate with CD4 levels for virus strains 208K2, 208K4,
14588,208K10,and 13956.Theslopes ofthebest-fitlines are also close to 1 (0.74 to 1.48) for all of these last viruses, consistent with a direct
relationshipof titers with CD4. The results suggest no relationship of titers with CD4 levels for the LA viruses. The primary virus 13231 data are
scatteredand areof uncertainsignificance (see text).
interactions of sCD4 with LAversus primary HIV-1 virions
might reflect similar differences in the interactions of these viruses with cellsurfaceCD4. If so,thismight account for our
dataon the differences in CD4 dependencefor infectivity of
primaryand LAviruses.Forexample,LAviruses withahigher CD4bindingaffinity andmorerapidkinetics of entry might be lessdependent onthe level of cell surface CD4,andprimary
isolate virions with a lower CD4binding affinity and slower
kineticsof entrymightbemoredependentonthe level of cell
surface CD4.
The HIV-1envelopeproteinis knowntobeimportantin cell
binding and entry and probably is the main viral structure
influencing virus-CD4 interactions of LA andprimaryHIV-1 isolates. HIV-1 envelope is also known to influence cell tropism, leading to differences between
T-cell-tropic
virusstrains,whichreadily infect HeLa-CD4 cells, and macrophage-tropicvirus strains,which do notinfect HeLa-CD4 cells
(12).
Previous studies ofvirus-CD4 interactions using sCD4
com-pared
macrophage-tropic primary
isolates or clones withT-cell-tropicLAisolatesorclones
(14).
Intheseexperiments,
the V3 region of the envelope gene from amacrophage-tropic
clonewas shown to influence both macrophagetropism
and resistancetosCD4(36).
The present data withLAandprimary
HIV-1isolates,whichwereall T-cell
tropic,
demonstrate that thephenotypicdifferences between virus-CD4interactions ofLAand primary isolates are not solely due to differences in macrophage or T-cell tropism. Thus, it seems likely that the
envelope regions such as V3, which are involved in the
specificityof celltropism,maybedifferent fromthose involved in acquisition of the LA phenotype. For example, in our
previousstudy, the V3 region of primary isolate 14558 gave the LAphenotype ofhighinfectibility for leukemia T cells when it was inserted in the genome of an LA virus (clone NL4-3),
whereasthis same V3regionwasassociated with the
primary-isolate phenotype when in its original 14558 genome
(12).
Studies are in progress to determine whether these non-V3 differences alsocorrelatewithchangesinsensitivityto neutral-izationbysCD4.
LA isolates differ from primary isolates in their ability to
spread efficientlyinrapidly dividing leukemicT-cell cultures. HeLa-CD4 or T-cell-tropic primary isolates often appear to
enter leukemic T cellsinitially, but subsequently
they
fail tospread rapidly enough to keep pace with the
dividing
cells(1Oa).Asaresult,theseinfected cell cultures
usually
clear the infections byoutgrowing the virus. Our resultswouldpredict
that in mixed virus infections, more
rapidly
spreading
LA HIV-1variants would beselectedbygrowth
individing
human leukemic Tlymphocytes. In contrast, passage in PHA-stimu-latednormalPBMC,whichreplicate
moreslowly,
might apply
less pressure selecting for LA HIV-1 variants.
Thus,
PBMCon November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.129.492.82.336.2]4.00
-NL4-3 208K10
3.50
3.00-None 2x104 2xl05 2xl 06 None 2x104 2x105 2x1
Number ofAbsorbingCells
FIG. 5. Failure ofadsorptionwithlargenumbers ofCD4-positive
HeLacellsorPBMC todepleteinfectious HIV-1 fromcell-free virus
stocks. A 0.2-ml volume of a 1/10 dilution of each virus stock was incubated for 2 hat37°Cwith various numbers ofCD4-positiveclone
1022HeLa cells (0) orwith PHA-stimulated PBMC (A).The cells
were thenpelleted, andtitersweredeterminedforsupernatantfluid
forinfectivityon 1022cellsasusual.
cultures might not result in selection of the LA phenotype capable of efficient replicationin leukemic T cells.
Conversely, inseropositive patients,thereappears tobeno selection for the LA phenotype, at least prior to onset of
advanced disease. Inpart,thismaybebecause normal
nonma-lignant CD4-positive lymphocytes are not a rapidly dividing
population. One consequence is that the T-cell-tropicvirus
populationsinvivorequire highamounts ofCD4 forinfection
of cells. Because these viruses probably adsorb onto cells
inefficiently, they might also be prone to disseminate in vivo and to escape local immune responses. It isconceivable that
selection of these viruses might account for the increasing killingof CD4-positive Tlymphocytes duringtheprogression
ofAIDS. The isolatesthatwestudied herewereobtained from
bothsymptomaticandasymptomatic patients. Further studies
will be required to determine howthese viruses are selected
and tounderstandtheir roles in disease.
ACKNOWLEDGMENTS
Supportedinpart byNIHgrantsCA25810 andAI28572.This work
was initiated by D.K. during a sabbatical leave with Robin Weiss
(Instituteof CancerResearch)thatwassupported byaFogartySenior InternationalFellowshipfrom theNIH.
We aregrateful to RobinWeiss and his coworkers forgenerous
warmhospitality and forstimulatingdiscussions. PaulClapham
gen-erouslydonatedmonoclonalantibodyADP318. REFERENCES
1. Allison,A.C.,andR. C.Valentine. 1960. Virusparticle adsorption.
III.Adsorptionofvirusesbycellmonolayersandeffects ofsome variables onadsorption. Biochim.Biophys.Acta40:400-410.
2. Aoki,N.,T. Shioda,H. Satoh, and H.Shibuta. 1991. Syncytium
formationofhumanandnon-humancellsbyrecombinant vaccinia
virusescarryingtheHIVenvgeneandhumanCD4gene.AIDS
5:871-875.
3. Ashkenazi, A., D. H. Smith, S. A. Marsters, L. Riddle, T. J. Gregory,D. D.Ho,and D.J.Capsen.1991.Resistanceofprimary
HIV-1 isolates to soluble CD4 is independent of CD4-gpl20
binding affinity.Proc.Natl.Acad. Sci.USA88:7056-7060.
4. Ashorn,P.A.,E. A.Berger,and B.Moss.1990. Human immuno-deficiency virus envelope glycoprotein/CD4-mediated fusion of nonprimatecellswith humancells. J. Virol. 64:2149-2156.
5. Berg,H.C.,and E. M.Purcell. 1977.Physicsofchemoreception.
Biophys. J. 20:193-219.
6. Bestwick, R. K., S. L. Kozak, and D. Kabat. 1988. Overcoming interference toretroviral superinfection results in amplified
ex-pressionand transmission of cloned genes. Proc. Natl.Acad. Sci. USA85:5404-5408.
7. Brightly, D. W.,I.S.Y.Chen, M. Rosenberg, and M. Ivey-Hoyle.
1991. Neutralization-resistant primaryclinical isolates of HIV-1 possess gpl20 glycoproteins with high affinity for recombinant
sCD4. Proc.Natl. Acad. Sci. USA 88:7802-7805.
8. Capon, D. J., and R. H. R. Ward. 1991. The CD4-gpl20interaction
and AIDSpathogenesis. Annu. Rev. Immunol. 9:649-678.
9. Chesebro, B., R. Buller, J. Portlis, and K. Wehrly. 1990. Failure of
humanimmunodeficiency virusentryand infection in CD4-posi-tive human brain and skin. J. Virol.64:215-221.
10. Chesebro, B., andK.Wehrly. 1988. Development of a sensitive
quantitative focalassayfor humanimmunodeficiency virus
infec-tivity.J.Virol.62:3779-3788.
10a.Chesebro, B.,andK.Wehrly. Unpublished data.
11. Chesebro, B., K. Wehrly, J. Metcalf, and D. E.Griffin.1991. Use
ofa newCD4-positiveHeLa cell clone for directquantitationof
infectious human immunodeficiency virus from blood cells of
AIDSpatients. J. Infect. Dis. 163:64-70.
12. Chesebro, B., K. Wehrly, J. Nishio, and S. Perryman. 1992.
Macrophage-tropic human immunodeficiency virus isolates from
differentpatients exhibit unusual V3 envelopesequence homoge-neity in comparison with T-cell-tropic isolates: definition ofcritical
amino acids involved in celltropism. J. Virol. 66:6547-6554. 13. Clapham, P. R., D. Blanc, and R. A. Weiss. 1991. Specific cell
surfacerequirements for infection of CD4-positive cells byhuman
immunodeficiency virus type 1, type 2, and simian immunodefi-ciency virus. Virology 181:703-715.
14. Daar, E. S., X. L. Li, T. Moudgil, and D. D. Ho. 1990. High concentrations of recombinant soluble CD4arerequired to
neu-tralize primary human immunodeficiency virus type I isolates.
Proc.Natl. Acad. Sci.USA87:6574-6578.
15. Dragic, T.,P.Charneau,F.Clavel,andM. Alison.1992. Comple-mentation of murine cells for human immunodeficiency virus envelope/CD4-mediatedfusion in human/murine heterokaryons. J. Virol. 66:4794-4802.
16. Fenyo, E. M., L. Morfeldt-Manson, F. Chiodi, B. Lind, A. von
Gegerfelt, J. Albert,E.Olausson,and B.J. Asyo. 1988. Distinct replicative and cytopathic characteristics of human immunodefi-ciencyvirus isolates.Virology62:4414-4419.
17. Fernandez-Larsson, R., K. K. Srivastava, and H. L. Robinson. 1992.Replication ofpatient isolates of human immunodeficiency virustype1 in T cells:aspectrumofratesand efficiencies ofentry. Proc. Natl. Acad. Sci.USA 89:2223-2226.
18. Fisher,A.G.,M.B.Feinberg,S. F.Josephs,M. E.Harper, L. M.
Marselle,G.Reyes,M. A.Gonda,A.Aldovini, C. Debouk, R. C.
Gallo, and F. Wong-Staal. 1986. The trans-activator gene of
HTLV-III is essential for virus replication. Nature (London)
320:367-371.
19. Fuller,A.O., and W.C. Lee. 1992. Herpes simplex virustype 1 entrythroughacascade ofvirus-cell interactionsrequires different roles ofgOandgHinpenetration.J.Virol.66:5002-5012. 20. Gey, G. O., W. D. Coffman, and M. T. Kubicelk 1952. Tissue
culturestudies oftheproliferative capacity ofcervicalcarcinoma
and normalepithelium. Cancer Res. 12:264-265.
21. Gomatos,P.J.,N. M.Stamatos,H. E.Gendelman, A. Fowler, D. L.
Hoover, D. C.Kalter, D. S.Burke, E. C. Tramount, and M. S. Meltzer. 1990. Relativeinefficiency ofsoluble recombinant CD4 forinhibitionofinfectionby monocyte-tropic HIVin monocytes and Tcells. J. Immunol. 144:4183-4188.
22. Harrison,S.C.,J. Wang, Y. Yan, T. Garrett, J. Liu, U. Moebius, and E. Reinherz. 1992. Structureandinteractions of CD4. Cold SpringHarborSymp. Quant.Biol. 57:541-548.
23. Henderson,L.A., and M. N. Qureshi. 1993. A peptide inhibitor of human immunodeficiencyvirus infection binds to novel human cell surfacepolypeptides.J.Biol. Chem. 268:15291-15297. 24. Incardona, N. L. 1981. The chemical nature of virus-receptor
interactions. p. 157-168. In K. Lonberg-Holm and L. Philipson (ed.),Virusreceptors, part2.ChapmanandHall, London. 25. Kay, R.,andJ. McPherson.1987.Hybrid pUCvectorsfor addition
E
0
00)
0
-J
-j
a)
F
Ir
I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.62.303.78.241.2]ofnew restriction enzyme sites to the endsofDNAfragments.
NucleicAcids Res.15:2778.
26. Kieber-Emmons, T., and D. B. Weiner. 1992. Structural perspec-tivesofthe CD4 antigen. AIDS Res. Rev. 2:65-90.
27. Koenig, R., G. Ashwell, and J. A. Hanover. 1988. Glycosylation of CD4: tunicamycin inhibits surface expression. J. Biol. Chem. 263:9502-9507.
28. Lifson, J. D., G. R. Reyes, M. S. McGrath, B. S. Stein, and E. G. Engleman. 1986. AIDS retrovirus induced cytopathology: giant
cell formationand involvementof CD4 antigen. Science 232:1123-1127.
29. Lonberg-Holm, K. 1981.Attachment of animal viruses to cells: an introduction, p. 1-21. In K. Lonberg-Holm and L. Philipson (ed.), Virus receptors, part 2. Chapman and Hall, London.
30. Maddon, P. J., J. S. McDougal, P.R.Clapham, A. G. Dalgleish, S. Jamal, R. A. Weiss, and R. Axel.1988.The T4 gene encodes the AIDSvirusreceptorand is expressed in theimmune system and brain. Cell 54:865-874.
31. Mann, R., R.C. Mulligan, and D. Baltimore. 1983.Constructionof aretroviruspackagingmutantand itsuse toproducehelper-free defectiveretrovirus. Cell 33:153-159.
32. Miller, A. D., M.-F. Law, and I. M. Verma.1985. Generation of helper-free amphotropic retroviruses that transducea
dominant-acting, methotrexate-resistantdihydrofolate reductasegene.Mol.
Cell.Biol. 5:431-437.
33. Moore, J. P., and P. J. Klasse.1992.Thermodynamic and kinetic analysis ofsCD4bindingtoHIV-1virionsand ofgpl20
dissocia-tion. AIDS Res. Hum. Retroviruses8:443-450.
34. Moore, J. P., J.A. McKeating, Y. X.Huang, A.Ashkenazi, and D.D. Ho.1992. Virions ofprimary human immunodeficiency virus
type 1 isolates resistant to soluble CD4 (sCD4) neutralization differ in sCD4 binding and glycoprotein gpl20 retention from
sCD4-sensitive isolates.J.Virol. 66:235-243.
35. Moore, J. P., J. A. McKeating, R. A. Weiss, andQ.J. Satenau. 1990.Dissociationofgpl20 fromHIV-1virionsinducedby soluble
CD4. Science250:1139-1142.
36. O'Brien,W.A.,I.S. Y.Chen,D. D.Ho,and E.S. Daar.Mapping genetic determinants for human immunodeficiency virus type 1
resistancetosoluble CD4. J.Virol. 66:3125-3130.
37. Orloff,S.L.,M.S.Kennedy,A. B.Belperron,P.J. Maddon,and
J.S.McDougall.1993. Twomechanismsof soluble CD4 (sCD4)-mediated inhibition of human immunodeficiency virus type 1
(HIV-1) infectivity and their relation toprimary HIV-1 isolates with reducedsensitivitytosCD4. J. Virol. 67:1461-1471.
38. Page, K. A., N. R. Landau, and D. R. Littman. 1990.Construction
anduseofahumanimmunodeficiency virusvectorforanalysis of
virus infectivity.J.Virol. 64:5270-5276.
39. Page, K. A.,S.M.Stearns,and D. R.Littman. 1992.Analysis of mutations in the V3 domain of gpl60 that affect fusion and
infectivity.J.Virol.66:524-533.
40. Popovic, M., M. Sarngadharan, E. Read, and R. Gallo. 1984.
Detection, isolation and continuous production of cytopathic retroviruses(HTLV-III)from patients with AIDS and pre-AIDS. Science 224:497-500.
41. Sakai, K., S.Dewhurst, X. Ma, and D. J.Volsky.1988.Differences in cytopathogenicity and host cell range among infectious molec-ular clones of human immunodeficiency virus simultaneously isolated from an individual. J. Virol. 62:4078-4085.
42. Sattentau, Q. J., and J. P. Moore. 1991.Conformational changes
inducedinthehuman immunodeficiency virus envelope glycopro-teinby soluble CD4 binding.J. Exp.Med. 174:407-415. 43. Schwartz, M. 1976.The adsorptionofcoliphage lambda to its host:
effect of variations in the cell surface density of receptor and in phage-receptoraffinity.J. Mol.Biol. 103:521-536.
44. Shieh, M.-T., D. WuDunn, R.I.Montgomery, J. Esko, and P. G. Spear. 1992. Cell surface receptors for herpes simplex virus are
heparin sulfate proteoglycans.J. CellBiol. 116:1273-1281. 45. Shioda, T., J. A. Levy, and C. Cheng-Mayer. 1992. Small amino
acidchanges inthe V3hypervariable region of gpl20canaffectthe
T-cell-lineandmacrophage tropism of humanimmunodeficiency virustype 1.Proc. Natl. Acad.Sci. USA89:9434-9438.
46. Srivastava, K. K., R. Fernandez-Larsson, D. Zinkus,and H. L. Robinson. 1991. Human immunodeficiency virus type 1 NL4-3
replicationinfourTcelllines: rate andefficiencyof entry,amajor determinant ofpermissiveness.J. Virol.65:3900-3902.
47. Stein, B. S., S. D.Gowda, J. D.Lifson, R. C.Penhallow, K. G. Bensch, and E.G. Engleman. 1987. pH-independent HIVentry intoCD4-positiveTcellsvia virusenvelopefusion to theplasma
membrane. Cell49:659-668.
48. Thali, M., U.Olshevsky, C.Furman, D. Gabuzda, J. Li, and J. Sodroski. 1991. Effectsofchangesingpl2O-CD4 binding affinity
on human immunodeficiency virustype 1 envelopeglycoprotein function and solubleCD4sensitivity.J.Virol.65:5007-5012.
49. Turner,S., R. Tizard, J. DeMarinis, R. B.Pepinsky,J.Zullo,R.
Schooley,andR. Fisher. 1992. Resistance ofprimaryisolatesof humanimmunodeficiency virustype 1toneutralizationbysoluble
CD4 isnotduetoloweraffinity with theviralenvelope glycopro-teingpl20.Proc.Natl. Acad. Sci. USA 89:1335-1339.
50. Wang, H., R. Paul, R. E. Burgeson, D. R. Keene, and D. Kabat. 1991. Plasma membrane receptor for ecotropicmurine
retrovi-rusesrequiresalimitingaccessoryfactor. J.Virol.65:6468-6477. 51. Weiss, R. A.Cellularreceptorsand viralglycoproteins involvedin retrovirus entry. In J. A. Levy (ed.), The retroviruses, vol. 2. PlenumPress,in press.
52. Werner, A., and J. A.Levy.1993. Humanimmunodeficiencyvirus type 1 envelope gpl20is cleaved after incubation with recombi-nantsolubleCD4. J.Virol.67:2566-2574.
53. White, J.,M. Kielan, and A. Helenius. 1982. Membrane fusion proteins of enveloped animal viruses. Q. Rev. Biophys. 16:151-195.