0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.13.8422–8430.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
A Functional Histidine-Tagged Replication Initiator Protein:
Implications for the Study of Single-Stranded DNA
Virus Replication In Planta†
Julio C. Vega-Arreguı´n, Tatiana Timchenko, Bruno Gronenborn,*
and Bertha Cecilia Ramı´rez*
Institut des Sciences du Ve´ge´tal, CNRS, 91198 Gif sur Yvette Cedex, France
Received 19 November 2004/Accepted 6 March 2005
Replication initiation of nanoviruses, plant viruses with a multipartite circular single-stranded DNA ge-nome, is triggered by the master Rep (M-Rep) protein. To enable the study of interactions between M-Rep and viral or host factors involved in replication, we designed oligohistidine-tagged variants of the nanovirusFaba
bean necrotic yellows virus(FBNYV) M-Rep protein that allow affinity purification of enzymatically active M-Rep
from plant tissue. The tagged M-Rep protein was able to initiate replication of its cognate and other FBNYV DNAs in Nicotiana benthamiana leaf disks and plants. The replicon encoding the tagged M-Rep protein multiplied and moved systemically in FBNYV-infectedVicia fabaplants and was transmitted by the aphid vector of the virus. Using the tagged M-Rep protein, we demonstrated the in planta interaction between wild-type M-Rep and its tagged counterpart. Such a tagged and fully functional replication initiator protein will have bearings on the isolation of protein complexes from plants.
Nanoviruses are single-stranded DNA (ssDNA) plant viruses (41). Their multipartite genome consists of at least six to eight circular individually encapsidated ssDNA molecules of ca. 1 kb, with one virion sense open reading frame (ORF) each (5, 7, 23, 35). Nanovirus DNA components and their respective encoded proteins of known function comprise DNA-R encod-ing the master Rep (M-Rep) protein (39), DNA-S encodencod-ing the capsid protein (43), DNA-C encoding the cell cycle link protein Clink (4), DNA-M encoding a movement protein, and DNA-N encoding a nuclear shuttle protein (42). The functions of at least three more proteins, all encoded by additional in-dividual genome components, remain unknown (41).
Nanoviruses multiply in the nucleus of infected cells by roll-ing-circle replication (RCR). Upon infection of a host cell, short DNA molecules encapsidated together with the viral ssDNA serve as primers for host polymerase(s) to initiate synthesis of the complementary (minus) strand DNA, creating a double strand (16). The double strand serves as transcription template and for RCR, which is initiated and terminated by replication initiator (Rep) proteins. Nanoviruses encode dif-ferent Rep proteins; however, only the 33-kDa M-Rep protein is required and sufficient to catalyze replication initiation of its coding DNA and of the other virus genome components (22, 39, 40). Faba bean necrotic yellows virus (FBNYV) M-Rep, expressed inEscherichia coli, has origin-specific DNA cleavage and nucleotidyl transfer activities in vitro and is an ATPase, both essential functions for viral DNA replication in vivo (39). During RCR the M-Rep protein cleaves the consensus
nonamer sequence TAGTATT2AC located at the origin of replication (39), creating a 3⬘-OH terminus and, by analogy to geminivirus replication, is thought to prime viral (plus) strand DNA synthesis (26).
For geminivirus Rep proteins, interactions with several host proteins have been described (1, 25). All these different pro-teins interacting with Rep were identified by using the yeast two-hybrid system. However, due to difficulties encountered in purifying the respective protein complexes, very little is known about the in planta interaction of these proteins. For nanovi-ruses, nothing is known about the host proteins that interact with M-Rep during replication.
We have designed oligohistidine-tagged M-Rep variants of the nanovirus FBNYV that are proficient to catalyze viral DNA replication initiation and termination in Nicotiana benthamiana and report the affinity purification from plant tissue of enzymatically active histidine-tagged M-Rep protein. To the best of our knowledge, this is the first example of a tagged ssDNA virus replication initiator protein that is func-tional in vivo and that is readily purified from plant tissue. In addition, replicons encoding oligohistidine-tagged M-Rep multiplied and moved systemically along with wild-type FBNYV in its natural hostVicia faba.The replicon encoding the modified M-Rep protein could be transmitted by aphids in a mixed infection with wild-type virus. Finally, we show the in planta interaction of the tagged M-Rep with wild-type M-Rep, suggesting that this protein may be used to identify other protein partners.
MATERIALS AND METHODS
Construction of DNA-R-His replicons of FBNYV.Basic molecular biology protocols were as described previously (34). Cloning of FBNYV DNAs of the Egyptian isolate EVI-93 (FBNYV-EG) has also been described elsewhere (39). A BamHI site was introduced by PCR-based QuikChange mutagenesis (Strat-agene) at nucleotide position 54 into the noncoding sequence of FBNYV-EG DNA-R (previously designated C2) (23, 39) by using the primers C2BamHI(⫹)
* Corresponding author. Mailing address: Institut des Sciences du Ve´ge´tal, CNRS, Avenue de la Terrasse, Baˆt. 23, 91198 Gif sur Yvette Cedex, France. Phone: (33) 169823571. Fax: (33) 169823695. E-mail for B. C. Ramı´rez: [email protected]. E-mail for B. Gronenborn: [email protected].
† Supplemental material for this article may be found at http://jvi .asm.org/.
8422
on November 8, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 8, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 8, 2019 by guest
http://jvi.asm.org/
(5⬘-GAGTCATCACGTGGATCCCACGTGATATTAG-3⬘) and C2BamHI(⫺) (5⬘-CTAATATCACGTGGGATCCACGTGATGACTC-3⬘). The DNA-R thus modified was subsequently inserted into the BamHI site of pBluescript II KS(⫹) (Stratagene). This plasmid (pBKSC2-B⫹) was used to construct DNA-R deriv-atives (replicons) encoding the oligohistidine-tagged M-Rep proteins. An NheI site was introduced by site-directed mutagenesis at nucleotide position 124 of DNA-R in plasmid pBKSC2-B⫹by using the primers C2NheI(⫹) (5⬘-GAATA AAATATGGCTAGCCAAGTTATATGC-3⬘) and C2NheI(⫺) (5⬘-GCATATA ACTTGGCTAGCCATATTTTATTC-3⬘). Complementary oligonucleotides coding for the His6tag flanked by an NheI site, 6His-NheI(⫹) (5⬘-CTAGCCA
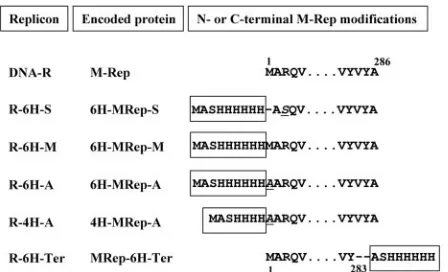
TCACCATCACCATCACG-3⬘) and 6His-NheI(⫺) (5⬘-CTAGCGTGATGGTG ATGGTGATGG-3⬘), were annealed and inserted into the previously generated NheI site of DNA-R, yielding replicon R-6H-S encoding protein 6H-MRep-S (Fig. 1).
The wild-type sequence at the N terminus of 6H-MRep-S was restored by reintroduction of methionine 1 (wild-type amino acid sequence) and substitution of serine 3 by arginine, leading to replicon R-M encoding protein 6H-MRep-M (Fig. 1). The primers used for site-directed mutagenesis of R-6H-S to create these amino acid changes were 6HisM-Rep2(⫹) (5⬘-CATCACCATCAC CATCACATGGCTCGGCAAGTTATATGCTGGTGC-3⬘) and 6HisM-R ep2(⫺) (5⬘-GCACCAGCATATAACTTGCCGAGCCATGTGATGGTGATG GTGATG-3⬘).
Replicon 6H-MRep-A was generated by substitution of the methionine of 6H-MRep-M by alanine (Fig. 1) by using primers 6HisA-Rep2(⫹) (5⬘-CATCA CCATCACGCGGCTCGGCAAGTT-3⬘) and 6HisA-Rep2(⫺) (5⬘-AACTTGC CGAGCCATGTGATGGTGATG-3⬘). The replicon containing only four histi-dines (4H-MRep-A) was obtained fortuitously in the same mutagenesis experiment (Fig. 1).
To generate the replicon R-6H-Ter encoding a M-Rep protein with a His6tag
at its C terminus (MRep-6H-Ter), an NheI site was introduced by site-directed mutagenesis at nucleotide position 970 of DNA-R by using the primers C2Nhe970(⫹) (5⬘ -GATAGGATTGTCTATGCTAGCGCGTGACGTCATGT-3⬘) and C2Nhe970(⫺) (5⬘-ACATGACGTCACGCGCTAGCATAGACAATCC TATC-3⬘), and the annealed oligonucleotides 6His-NheI(⫹) and 6His-NheI(⫺) (see above) were inserted into this NheI site (Fig. 1).
The correctness of all of the DNA-R modifications was verified by sequence analysis. Dimers of the respective molecules, excised by BamHI, were inserted into the BamHI site of plasmid pBin19. Subsequently, the pBin19 derivatives were transferred by electroporation (30) intoAgrobacterium tumefaciensstrains LBA4404 (31) and COR308 (17), provided by Cornell Research Foundation, Inc.
Agrobacterium-mediated inoculation.Agroinoculation, as well as agroinfec-tion, of plants by DNA virus genomes requires more than one complete unit of viral genetic information to be placed within the T-DNA, so that infectious viral DNA molecules can be produced by recombination or replication in the plant cell (13, 14, 37).N. benthamianaleaf disks were inoculated byA. tumefaciens
LBA4404 carrying dimers of the respective FBNYV DNAs in pBin19 and kept on culture media at 24°C with continuous light as described previously (21, 39). Leaves of 4- to 5-week-oldN. benthamianaplants were infiltrated at their abaxial
face withA. tumefaciensLBA4404 carrying the FBNYV DNAs in pBin19 by using a needleless syringe.
V. fabaplants were agroinoculated withA. tumefaciensCOR308 grown in YEB medium (0.5% nutrient broth, 0.5% peptone, 0.1% yeast extract, and 5 mM MgSO4, adjusted to pH 7.2) containing 50g of kanamycin/ml and 5g of
tetracycline/ml. An overnight culture was diluted 10-fold in YEB, 50g of kanamycin/ml, 5g of tetracycline/ml, 10 mM MES (morpholinoethanesulfonic acid; pH 6.0), and 50M acetosyringone (Sigma) and then cultured at 30°C until an optical density at 600 nm of ca. 1.5 was reached. The bacteria were pelleted by centrifugation (20 min at 3,500⫻g) and resuspended in 1/10 of the original volume of MS medium (Sigma), 10 mM MES, and 150M acetosyringone.V. fabaplants were injected with ca. 1 ml of the bacterial suspension by using a syringe with a needle.
All agroinoculated or virus-infectedN. benthamianaandV. fabaplants were cultivated at 25°C, in 50% humidity and with 16 h of light in growth chambers inside a restricted-access S3-confinement facility.
Aphid transmission assays.Virus and DNA-R-His replicon transmission as-says were performed by using the insect vectorAphis craccivora(kindly provided by L. Allala, INA, El Harrach, Algeria) and the hostV. faba. Viruliferous and nonviruliferous aphids were reared onV. fabaplants in cages inside growth chambers in S3 confinement as described above. Healthy plants, 7 to 10 days old, were infected with viruliferousA. craccivorafed on V. fabainfected by the FBNYV Algerian isolate (FBNYV-DZ [provided by L. Allala]). The plants were agroinoculated either immediately or 2 days later withA. tumefaciensCOR308 carrying dimers of the DNA-R-His replicons in pBin19. The presence of the rep-licons in the infected symptomatic plants was tested 10 to 15 days postagroin-oculation (dpa) by immunocapture-PCR (IC-PCR) with the DNA-R-His-specific oligonucleotides 6His-NheI(⫹) and C2Nhe970(⫺). For transmission, nonvirulif-erousA. craccivorawere fed on FBNYV-infected and agroinoculated plants for an acquisition access period of 3 days and subsequently transferred to healthy 7-to 10-day-oldV. fabaplants for an inoculation access period of 5 days. Insects were killed by treatment with an insecticide (0.2% Dedevap; Bayer). After 2 weeks, samples of newly developed leaves of these plants were tested for viral or replicon DNAs by PCR with purified DNA or by IC-PCR of crude extracts with the primers C2Nhe205(⫹) (5⬘-ATGAAGTATCTTGCTAGCCAAACTGAAC AA-3⬘) and C2Nhe970(⫺) for the virus and 6His-NheI(⫹) and C2Nhe970(⫺) for the replicons. The primers M13(⫹) (5⬘-GTAAAACGACGGCCAGT-3⬘) and M13(⫺) (5⬘-GGAAACAGCTATGACCATG-3⬘) were used to confirm the ab-sence of any contaminating pBin19 T-DNA in the samples.
Further transmission of the DNA-R-His replicon byA. craccivorawas per-formed by first agroinoculation with FBNYV-EG cloned DNAs (T. Timchenko et al., unpublished data) and subsequently infection with viruliferous (FBNYV-DZ) A. craccivora.V. faba plants were agroinoculated with A. tumefaciens
COR308 cultures, each one carrying a dimer of the eight FBNYV-EG genome components (DNA-R, DNA-C, DNA-M, DNA-N, DNA-S, DNA-U1, DNA-U2, and DNA-U4) (41), along with agrobacteria carrying a dimer of the DNA-R-His replicon. Agroinoculated plants that had developed symptoms at 21 dpa (4 of 34 plants) were superinfected with viruliferousA. craccivorafor 3 days, after which the insects were removed. After a further 3 days, nonviruliferous insects were allowed access to these primary plants (agroinoculated with eight FBNYV-EG DNAs plus the DNA-R-His replicon and subsequently exposed to viruliferous insects) for a 6-day acquisition period. The aphids were then transferred to healthyV. faba(secondary plants), and transmission of the DNA-R-His replicon to secondary plants was tested after 8 and 25 days by PCR with the DNA-R-His-specific primers 6His-NheI(⫹) and C2Nhe970(⫺).
IC-PCR.Detection of FBNYV DNA-R and DNA-R-His replicons was carried out by IC-PCR as described previously (12) with a monoclonal antibody raised against FBNYV (kindly provided by H.-J. Vetten, BBA, Braunschweig, Ger-many). Primers C2Nhe205(⫹) and C2Nhe970(⫺) were used to detect DNA-R (797-bp fragment), and primers 6His-NheI(⫹) and C2Nhe970(⫺) were used to detect the DNA-R-His replicons (891-bp fragment).
Analysis of DNA replication by Southern blot.Replication inN. benthamiana
leaf disks of the DNA-R-His replicons was analyzed as described previously (39, 40). DNA was extracted fromN. benthamianaleaf disks at 4 to 5 dpa, fraction-ated by electrophoresis in 1% agarose gels, transferred to nylon membranes, and detected by hybridization with component-specific32
[image:2.585.53.277.68.204.2]P-labeled probes. The probe used for DNA-R was the entire component (1003 bp) of R-6H-A linearized by BamHI. Probes for DNA-S (a fragment of 608 bp) and DNA-C (a fragment of 543 bp) were generated by PCR with the following sets of primers: C5Nhe328(⫹) (5⬘-AAAATGGCTAGCAAATGGAATTGGTCTGGTACGAA-3⬘) and FBIR2 B(⫺) (5⬘-TCCGCTGAACCTGGGGCGGGGGTAATACTAAGCCC-3⬘), fol-lowed by HincII digestion for DNA-S; C10BamHI(⫹) (5⬘-CGTTGTTCTTGG ATCCAAGATGGGTCTGAA-3⬘) and C10EcoRI(⫺) (5⬘-TTTAATTACGAAT
FIG. 1. DNA-R-His replicons. The designations of the replicons, their respective encoded proteins, and the N- and C-terminal amino acid sequences of the corresponding proteins (wild-type and oligohis-tidine-tagged M-Rep) are shown. Amino acid changes other than the oligohistidine tag, indicated inside a rectangle, are in italics and un-derlined or represented by a dash.
on November 8, 2019 by guest
http://jvi.asm.org/
TCTCAACTAATTACAATATCC-3⬘) for DNA-C. DNA fragments were fractionated by 1% agarose gel electrophoresis, purified by MiniElute columns (Qiagen), and32P labeled (Amersham Megaprime Kit).
For replication assays of other FBNYV DNAs, leaf disks were coagroinocu-lated with equal volumes of two cultures ofA. tumefaciensLBA4404: one car-rying in pBin19 dimers of one of the DNA-R-His replicons and another carcar-rying in pBin19 dimers of DNA-S or DNA-C, respectively. Quantification of the replicative DNA forms was done by using a Storm 840 PhosphorImager and ImageQuant software (Molecular Dynamics).
Purification of oligohistidine-tagged proteins.Since it had been shown that FBNYV Clink protein enhances replication of FBNYV DNA up to 7-fold (3), agrobacteria containing the Clink-encoding DNA-C was always included, along with the DNA-R or DNA-R-His replicons in transfection experiments for pro-tein expression. For analytical purposes, total propro-tein fromN. benthamianaleaf disks harvested 4 or 5 dpa was extracted (at 1:5 fresh wt/vol) in HEPES buffer (50 mM HEPES [pH 8.0], 150 mM NaCl, 50 mM EDTA, 0.05% NP-40, 10% glycerol, 1 mM phenylmethylsulfonyl fluoride [PMSF], 0.1 KI units of aproti-nin/ml [Roche], 10 mM-mercaptoethanol). The extract was centrifuged at 15,000⫻gfor 15 min at 4°C. The protein concentration of the supernatant was determined by the Bradford assay (6). Then, 10g of total protein was fraction-ated by sodium dodecyl sulfate–12% polyacrylamide gel electrophoresis (SDS– 12% PAGE). Proteins were electrotransferred for 1 h at 200 mA to a polyvi-nylidene difluoride membrane (Amersham), which then was blocked for 2 h at room temperature with 5% (wt/vol) skimmed milk in TBS buffer (20 mM Tris-HCl [pH 7.6], 150 mM NaCl) containing 0.1% Tween 20. The membranes were incubated with polyclonal anti-M-Rep antibodies (1:2,000 dilution in TBS buffer of a sixth bleeding serum, preadsorbed by overnight incubation at 4°C with total protein extracts ofN. benthamiana,Arabidopsis thaliana, andV. faba). Detection of antigen was performed with anti-rabbit immunoglobulin G coupled to alkaline phosphatase (Sigma-Aldrich). For antibody production, oligohistidine-tagged M-Rep protein purified fromEscherichia colicontaining plasmid pQE30-rep2 was used (39). Antisera were raised in rabbits and kindly provided by H.-J. Vetten.
Purification of oligohistidine-tagged M-Rep proteins by immobilized metal ion affinity chromatography (IMAC) was essentially as described previously (3) with slight modifications. AgroinfiltratedN. benthamianaleaves or agroinoculated leaf disks were harvested at 3 or 4 days postinfiltration. Total protein from leaves (10 g [fresh weight]) was extracted at a 1:5 (wt/vol) ratio in TN buffer (20 mM Tris-HCl [pH 8.0], 500 mM NaCl, 1 mM PMSF, 0.1 KI units of aprotinin/ml, 5 mM-mercaptoethanol, 5 mM imidazole, and one tablet of Complete Mini, EDTA-free, protease inhibitor cocktail [Roche] per 10 ml of buffer). The extract was centrifuged at 15,000⫻gfor 15 min. All extraction and purification steps were carried out at 4°C. The protein concentration was determined by the Bradford assay (6). Total protein (30 mg) was incubated with 500l of Talon metal (Co2⫹) affinity resin (Clontech) for 30 min. The resin was washed once
with 40 ml of TN buffer and once with 20 ml of TN buffer containing 20 mM imidazole. Oligohistidine-tagged M-Rep proteins were eluted with 500 mM im-idazole in TN buffer. Otherwise, the resin was stored at⫺20°C until proteins were analyzed by SDS-PAGE and Western blotting as described above.
For purification of enzymatically active tagged M-Rep, total protein was ex-tracted from 9 g of leaf disks (ca. 300 disks) at 5 dpa with 45 ml of modified TN buffer (without -mercaptoethanol and containing 10% glycerol and 0.5% Tween 20). The extract was incubated for 30 min on ice with DNase and RNase (10g/ml each) prior to centrifugation. For protein purification by IMAC, 500
l of Talon metal (Co2⫹) affinity resin were used. After protein binding, the
column was washed as described above, and the tagged M-Rep protein was eluted with 5 column volumes of modified TN buffer containing 400 mM imida-zole. Samples were kept on ice at 4°C for a maximum of 1 week.
Purification of M-Rep fromE. coli.The 6H-MRep-M coding sequence was excised from the replicon R-6H-M by digestion with NheI and BamHI. The fragment was purified from an agarose gel, digested with Sau3AI, and cloned into pET21a (Novagen) at the NheI and BamHI sites. The resulting plasmid pET21a-R-6H-M was introduced intoE. coliBL21(DE3)-recA(2, 38), harboring plasmid pRep4 (Qiagen) for tightly controlled expression of the recombinant protein. Bacteria were grown at 37°C in LB medium to an optical density at 600 nm of ca. 0.6 and transferred to 18°C for 1 h before induction at 18°C with 0.5 mM IPTG (isopropyl--D-thiogalactopyranoside) for 5 h. The bacteria were centrifuged, and the pellet was resuspended in buffer A (50 mM phosphate buffer [pH 8.0] containing 500 mM NaCl, 10% glycerol, 0.5% Tween 20, and 1 mM PMSF). Bacterial lysis and protein purification fromE. coliwas by IMAC as described previously (34), except that the washes were performed as follows: two washes with 40 column volumes of buffer A containing 10 mM imidazole and one wash with 20 column volumes of buffer A containing 40 mM imidazole. The
6H-MRep-M protein was eluted with 5 volumes of buffer A containing 400 mM imidazole. Samples for enzymatic activity were kept on ice at 4°C for 1 week.
ATPase assay.The ATPase activity of the tagged M-Rep purified from plant tissue and fromE. coliwas determined as described previously (39). Totals of 175 ng of tagged M-Rep fromN. benthamianaand 3.5g fromE. coliwere incubated either at 37°C for 45 min, at room temperature for 45 min, or at room temper-ature for 16 h with 5 nM [␥-33P]ATP and 5M nonlabeled ATP. The reaction
products were separated by thin-layer chromatography on polyethyleneimine cellulose F plastic sheets (PEI; Merck) using 0.5 M LiCl–1 M formic acid as a running buffer. The amount of [33
P]orthophosphate liberated by M-Rep was quantified by using a PhosphorImager (Molecular Dynamics).
RESULTS
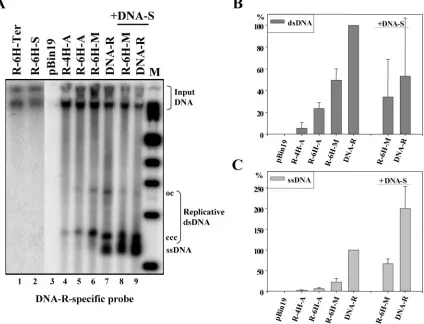
Oligohistidine-tagged FBNYV M-Rep proteins are func-tional in planta.To determine whether the addition of a series of histidines to the N or C terminus of FBNYV M-Rep inter-feres with protein function, the replication initiation activity of several oligohistidine-tagged protein variants was studied in planta. Agrobacteria containing FBNYV DNA-R or one of the replicons encoding an M-Rep protein with an N- or C-terminal oligohistidine tag were used (Fig. 1), and agrobacteria with the T-DNA vector pBin19 alone served as a control. Total DNA was extracted from leaf disks at 4 or 5 dpa and analyzed by Southern blot with DNA-R as the probe. Wild-type M-Rep protein initiated replication of its coding DNA (Fig. 2A, lanes 7 and 9). In contrast, the oligohistidine-tagged M-Rep protein lacking methionine 1 (of the wild-type protein) and whose arginine 3 was replaced by serine (6H-MRep-S, encoded by replicon R-6H-S; Fig. 1) was not able to catalyze replication initiation (Fig. 2A, lane 2). The same result was obtained when six histidines were added to the C terminus of M-Rep (MRep-6H-Ter, encoded by replicon R-6H-Ter; Fig. 2A, lane 1). Therefore, changing arginine 3 and deleting methionine 1 at the N terminus in conjunction with addition of a His6tag, as
well as deleting two amino acids at the C terminus of M-Rep in conjunction with addition of a His6 tag (Fig. 1), abolished
the capacity of the respective modified M-Rep proteins to initiate FBNYV DNA replication.
In contrast, replicon R-6H-M encoding the 6H-MRep-M protein with no changes in the M-Rep sequence other than the N-terminal addition of the tag (Fig. 1) replicated in leaf disks (Fig. 2A, lanes 6 and 8). Protein analysis suggested that only a single M-Rep species was produced from the R-6H-M replicon (see below). To rule out the possibility of any low-level trans-lation restart after the tag at methionine 1 of the wild-type sequence that might have been undetected, the replicon R-6H-A coding for 6H-MRep-A in which the wild-type methi-onine 1 of 6H-MRep-M was replaced by alanine was generated (Fig. 1). Replicon R-4H-A (coding for 4H-MRep-A) was ob-tained fortuitously. These two replicons also replicated in leaf disks (Fig. 2A, lanes 4 and 5), albeit less efficiently. This result excluded the possibility that the observed replication of R-6H-M was due to undetected amounts of wild-type Rep expressed by internal initiation. Replication experiments of DNA-R and replicon R-6H-M in the presence of DNA-S ex-pressing the virus capsid protein revealed a higher level of ssDNA, probably due to protection of the ssDNA by capsid protein (Fig. 2A, lanes 8 and 9, and Fig. 2C).
The amount of replicative DNA species in three indepen-dent experiments was quantified. Replicons R-6H-M, R-6H-A, and R-4H-A led to less replicative DNA than wild-type
on November 8, 2019 by guest
http://jvi.asm.org/
DNA-R. In the case of replicon R-6H-M compared to DNA-R the reduction was 2-fold for double-stranded DNA (dsDNA) (Fig. 2B) and 4-fold for ssDNA (Fig. 2C); it was about 5-fold for dsDNA and 14-fold for ssDNA in the case of R-6H-A compared to DNA-R (Fig. 2B and 2C). Since R-6H-M (Fig. 2A, lane 6) replicated more efficiently than R-6H-A and R-4H-A (Fig. 2A, lanes 4 and 5), methionine 1 of the wild-type protein may be important for M-Rep function. In the presence of DNA-S, the increase of ssDNA (Fig. 2C) seems to correlate with a decrease of dsDNA (Fig. 2B). Collectively, the results demonstrate that the addition of an oligohistidine tag to the amino terminus of FBNYV M-Rep (MRep-M, 6H-MRep-A, and 4H-MRep-A; Fig. 1) does not abolish replica-tion in planta.
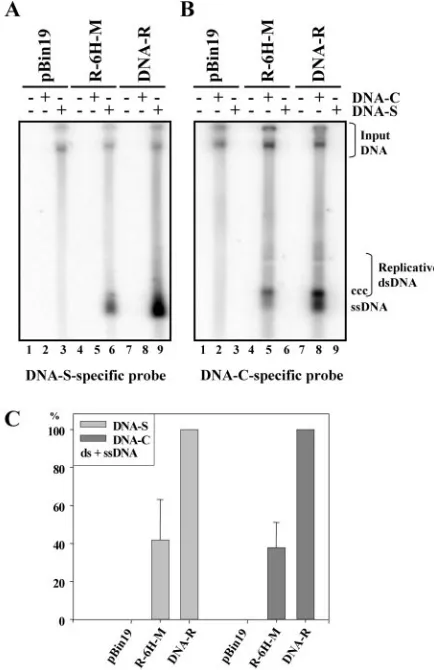
Initiation of replication of other FBNYV genome compo-nents by the oligohistidine-tagged M-Rep proteins.M-Rep is the only viral protein necessary to initiate replication of all the genomic DNAs of a nanovirus (39, 40). To determine whether the tagged M-Rep proteins also catalyzed replication initiation of other FBNYV DNAs, we tested replication of FBNYV DNA-S (capsid protein) and DNA-C (Clink protein) in the
presence of the DNA-R-His replicons. N. benthamiana leaf disks were coagroinoculated with different combinations of two cultures of agrobacteria: one carrying in pBin19 dimers of either DNA-R, R-6H-M, R-6H-A, or R-4H-A and the the other carrying dimers of FBNYV DNA-S or DNA-C, respec-tively (39). Wild-type M-Rep protein initiated replication of FBNYV DNA-S (Fig. 3A, lane 9) and DNA-C (Fig. 3B, lane 8). Oligohistidine-tagged M-Rep proteins MRep-M, 6H-MRep-A (not shown), and 4H-6H-MRep-A (not shown) proved capable of catalyzing replication of FBNYV DNA-S (Fig. 3A, lane 6) and DNA-C (Fig. 3B, lane 5). Quantification of the transreplication data of DNA-S and DNA-C from three inde-pendent experiments showed reduced replication levels with the modified M-Rep protein (Fig. 3C), probably due to the reduced levels of autonomous replication observed in the au-toreplication assays (Fig. 2). These results demonstrate that the oligohistidine-tagged M-Rep proteins act as master Rep proteins.
Systemic spread of the DNA-R-His replicons in FBNYV-infectedV. fabaplants.To determine whether the DNA-R-His replicons spread systemically within the host plant in the con-FIG. 2. Replication of FBNYV DNA-R-His replicons inN. benthamianaleaf disks. (A) Total DNA extracted from leaf disks agroinoculated with FBNYV DNA-R-His replicons was analyzed by Southern blotting with a DNA-R-specific probe. DNA from inoculation with pBin19 alone was used as a control (lane 3). Lanes 1 and 2 are from a different gel. Positions of replicative viral DNA forms and input DNA are indicated. ccc, covalently closed circular DNA; oc, open circular DNA. M, molecular marker. (B and C) Quantification of the ds- and ssDNA forms from three independent replication assays. The amount of the respective DNA forms is given as a percentage of wild-type DNA (DNA-R). Replication of R-6H-M and DNA-R in the presence of DNA-S (encoding the capsid protein) and its quantification are shown in panel A, lanes 8 and 9, and panels B and C as indicated.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.83.507.76.400.2]text of a virus infection, V. faba was infected with wild-type FBNYV byA. craccivoraand subsequently agroinoculated with A. tumefaciens strain COR308 carrying in pBin19 dimers of replicons R-6H-M or R-4H-A. Disease symptoms were moni-tored, and the presence of the DNA-R-His replicons was an-alyzed by IC-PCR of noninoculated symptomaticV. fabaleaves that had developed 2 weeks after agroinoculation. The R-6H-M replicon was detected in 100% (15 of 15) of the symptomatic plants (Fig. 4A shows four representative exam-ples). In two other experiments the R-6H-M replicon was detected in 87% (13 of 15) and 75% (6 of 8) of the symptom-atic plants. For R-4H-A the experiment was performed twice, and the replicon was detected in 67% (10 of 15) and 70% (7 of 10) of the symptomatic plants. T-DNA-specific primer se-quences (see Materials and Methods) were used to confirm the absence of pBin19 DNA in the leaves tested (not shown). These results indicate that the R-6H-M and R-4H-A replicons are able to spread systemically along with the wild-type
FBNYV DNAs. Systemic spread of the R-6H-M and R-4H-A replicons was not detected in control plants, which were agro-inoculated with R-6H-M or R-4H-A alone (Fig. 4A).
Transmission of FBNYV DNA-R-His replicons by aphids. To investigate whether the replicons are transmitted by aphids to healthy plants, FBNYV-infected symptomatic plants in which the R-6H-M or R-4H-A replicons had moved systemi-cally were used for transmission assays. For this purpose, non-viruliferous A. craccivora were starved for 5 h prior to an acquisition access period of 3 days on FBNYV-infected and R-6H-M or R-4H-A replicon-positive symptomatic plants (pri-mary plants). The aphids were then transferred to healthy plants (secondary plants) and maintained for an inoculation access period of 5 days. Secondary plants were monitored for symptom development and analyzed by IC-PCR 2 weeks after aphid access. Figure 4B shows a representative experiment in which only wild-type DNA-R was detected in four symptomatic secondary plants tested 2 weeks after inoculation access (Fig. FIG. 3. Trans-replication of FBNYV DNA-S and DNA-C. (A and
[image:5.585.55.272.70.405.2]B) Total DNA extracted fromN. benthamianaleaf disks agroinocu-lated with DNA-R and the R-6H-M replicon, along with DNA-S (A) or DNA-C (B) as indicated, was analyzed by Southern blotting with component-specific probes. Replicative viral DNA forms and input DNA are indicated as in Fig. 2. (C) Quantification of viral DNA forms. The amount of DNA-S and DNA-C (ds- and ssDNA) was quantified in three experiments, and the mean for each DNA-S and DNA-C replicated in the presence of the R-6H-M replicon is given as a percentage of that obtained in the presence of wild-type DNA-R.
FIG. 4. Systemic spread and transmission of FBNYV DNA-R-His replicons. (A) Detection in primary plants of the R-6H-M replicon by IC-PCR with DNA-R-His-specific primers in newly developed leaves ofV. fabainfected with FBNYV byA. craccivoraand agroinoculated with the R-6H-M replicon. Lanes 1 to 4, different individual aphid-infected and agroinoculated primary plants;non-inf, agroinoculated but not aphid-infected plant. (B) Detection in secondary plants of DNA-R and R-6H-M replicons by IC-PCR in newly developed leaves of V. fabainfected by aphids fed on primary FBNYV-infected and agroinoculated plants. DNA-R (left panel)- and DNA-R-His (right panel)-specific primers were used. Lanes 1 to 4, samples from different individual secondary plants. Numbers in the left and right panels cor-respond to samples from the same plants.inf, infected but not agro-inoculated primary plant;p, sample from an aphid-infected and agro-inoculated primary plant. (C) Detection in primary symptomatic plants of the DNA-R (lanes 1 and 2) and R-6H-M (lanes 3 and 4) replicons by PCR in newly developed leaves ofV. fabaagroinoculated with all eight FBNYV-EG DNA components, along with the R-6H-M repli-con. (D) Detection in secondary plants of DNA-R and R-6H-M rep-licons by PCR at 8 and 25 dpi as indicated. Numbers in the left and right panels correspond to the plant number. R, DNA-R-specific prim-ers; H, DNA-R-His-specific primprim-ers; M, molecular marker (in kilo-bases). Arrows indicate amplified DNA-R- or DNA-R-His-specific fragments (797 and 891 base pairs, respectively).
on November 8, 2019 by guest
http://jvi.asm.org/
4B, left panel). This indicates that wild-type FBNYV is readily transmitted by the aphids and that virus transmission is not affected by the multiplication of the R-6H-M or R-4H-A rep-licons. However, replicon R-6H-M was not detected in the secondary plants that had scored positive for DNA-R, i.e., had received virus from the aphids (Fig. 4B, right panel). The experiments were repeated three times with replicons R-6H-M and R-4H-A, respectively. Using these conditions, replicons expressing a tagged M-Rep were not transmitted by aphids along with the virus. These results show that, despite the sys-temic movement of the R-6H-M and R-4H-A replicons in a simultaneous infection with FBNYV, no transmission by aphids along with wild-type virus occurred, or that the repli-cons were rapidly lost after transmission.
In a different experimental setup, we used the established FBNYV infection by agroinoculatingV. fabaplants with eight cloned viral DNAs (T. Timchenko, et al., unpublished data). In addition to these eight DNAs, the R-GH-7 replicon was agroi-noculated into V. faba. Agroinoculated primary plants that developed symptoms at 21 dpa (4 of 34) were superinfected with viruliferousA. craccivoracarrying the isolate FBNYV-DZ for a 3-day inoculation access period. Viruliferous insects were removed and after a further 3 days nonviruliferous insects were allowed access to the primary plants for a 6-day acquisition access period. They were then transferred to healthyV. faba (secondary plants) for transmission, and the presence of rep-licon R-6H-M was assayed by PCR. DNA-R (no distinction is possible in this experimental set up between agroinoculated or aphid-delivered DNA-R) was found in primary plants 1 and 2 (Fig. 4C, lanes 1 and 2). By PCR, replicon R-6H-M could only be detected in primary plant 2 (Fig. 4C, lane 4), whereas it was detected in plant 1 by IC-PCR (data not shown). Transmission to secondary plants was analyzed at 8 and 25 dpi. DNA-R and replicon R-6H-M were detected in secondary plants 1 and 2 at 8 dpi (Fig. 4D). At 25 dpi, the amount of R-6H-M DNA had increased in both plants analyzed (Fig. 4D, right panel). These results show that aphids are able to transmit the modified replicon R-6H-M, provided it is agroinoculated, along with all eight FBNYV-EG DNAs and has spread systemically. Super-infection by viruliferous aphids containing FBNYV-DZ then allows for acquisition and transmission of the R-6H-M repli-con.
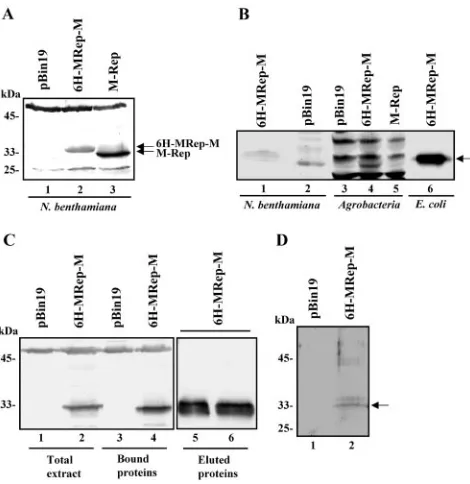
Affinity purification of oligohistidine-tagged FBNYV M-Rep proteins from plant tissue. The difficulty in purifying Rep proteins of nano- and geminiviruses under conditions of nat-ural infection has been a major problem in studying them in planta (24). Having demonstrated that oligohistidine-tagged FBNYV master Rep proteins were functional in planta, we assayed the expression of the tagged proteins by Western blot-ting with M-Rep-specific antisera. Figure 5A shows an example of 6H-MRep-M expressed inN. benthamianaleaf disks (lane 2). Similar levels of protein expression were observed for 6H-MRep-A and 4H-6H-MRep-A, and comparable results were ob-tained with agroinfiltrated N. benthamiana leaves (data not shown). Only one major protein species is produced from the R-6H-M replicon (Fig. 5A, lane 2), and the difference in size between the wild-type and tagged protein indicates the pres-ence of the tag (Fig. 5A, lanes 2 and 3, and Fig. 6). Therefore, we conclude that there is no translation initiation at the inter-nal methionine (methionine 1 of the wild-type M-Rep)
follow-ing the tag sequence. This was proven by replication of the 6H-MRep-A and 4H-MRep-A replicons (see above). To rule out that the Rep proteins detected were due to expression inA. tumefaciens, Western blots of protein extracts from agrobacte-ria carrying pBin19 alone (Fig. 5B, lane 3) or dimers of R-6H-M (Fig. 5B, lane 4) or DNA-R (Fig. 5B, lane5) were performed. Only nonspecific cross-reaction with other proteins was observed inA. tumefaciensirrespective of whether pBin19 carried a dimer of DNA-R or R-6H-M (Fig. 5B, lanes 3, 4, and 5, respectively). Similarly, nonspecific cross-reaction was ob-served in extracts ofN. benthamianainoculated with agrobac-teria carrying pBin19 (Fig. 5B, lane 2), whereas in extracts of N. benthamiana inoculated with agrobacteria carrying R-6H-M, 6H-MRep-M protein was detected (Fig. 5B, lane 1). Lane 6 shows 6H-MRep-M protein fromE. colithat served as control.
[image:6.585.303.538.68.310.2]We then progressed to purify oligohistidine-tagged M-Rep protein fromN. benthamianaplants by IMAC. The R-6H-M replicon was introduced by infiltration of agrobacteria into the FIG. 5.In planta expression and purification of oligohistidine-tagged M-Rep proteins. (A) Western blot analysis of crude extracts fromN. benthamianaleaf disks 4 days after inoculation with agrobac-teria containing the FBNYV DNA-R (lane 3) and the R-6H-M repli-con (lane 2). The positions of wild-type M-Rep of ca. 33 kDa (lane 3) and 6H-MRep-M (lane 2) proteins are indicated. The difference in migration between these two proteins is due to the additional His tag. An extract from inoculation with agrobacteria containing pBin19 was used as a control (lane 1). (B) Western blot analysis of extracts fromN. benthamianacompared to extracts fromA. tumefaciens. Extracts from N. benthamiana inoculated with agrobacteria carrying a dimer of R-6H-M in pBin19 (lane 1) or pBin19 alone (lane 2) and extracts from agrobacteria carrying pBin19 alone (lane 3), a dimer of R-6H-M (lane 4), or DNA-R (lane 5) were assayed. Lane 6 contains 6H-MRep-M protein fromE. coli. (C) Purification of 6H-MRep-M by affinity chro-matography from N. benthamiana leaves agroinfiltrated with the R-6H-M replicon. Western blot analysis of total extracts (lanes 1 and 2), bound proteins (lanes 3 and 4), and eluted proteins (lanes 5 and 6). (D) SDS-PAGE of the purified 6H-MRep-M protein stained with Coomassie brilliant blue (lane 2). The arrow indicates 6H-MRep-M.
on November 8, 2019 by guest
http://jvi.asm.org/
youngest leaves. The leaves were harvested 4 days postinfiltra-tion, and the tagged protein was extracted from plant tissue under nondenaturing conditions and bound to the Co2⫹-resin. The majority of 6H-MRep-M protein was eluted in the first four fractions containing 500 mM imidazole; fractions 1 and 2 are shown in Fig. 5C, lanes 5 and 6. The eluted fractions were pooled, analyzed by SDS-PAGE, and stained with Coomassie brilliant blue (Fig. 5D, lane 2). The results show that oligohis-tidine-tagged M-Rep proteins can readily be isolated fromN. benthamianaplants by a single purification step.
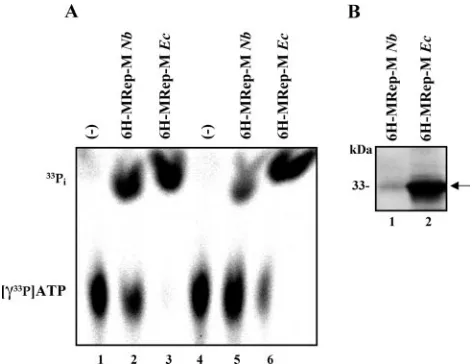
The 6H-MRep-M purified from plant tissue is enzymatically active.M-Rep possesses an ATPase activity that is essential for viral DNA replication in vivo (39). This ATPase activity and its requirement for DNA replication is also a characteristic fea-ture of geminivirus Rep proteins (11, 18). To prove that 6H-MRep-M isolated fromN. benthamianapossesses ATPase ac-tivity, the hydrolysis of [␥33P]ATP by the protein purified from
plant tissue was assayed. The same 6H-MRep-M expressed in and purified fromE. coliserved as a control. Figure 6A shows that the oligohistidine-tagged M-Rep has ATPase activity after purification fromE. coli(Fig. 6A, lanes 3 and 6) and fromN. benthamiana(Fig. 6A, lanes 2 and 5). Lanes 1 to 3 were from an assay at room temperature and 16 h of incubation; lanes 4 to 6 show the reaction products after 45 min at 37°C. A West-ern blot (Fig. 6B) illustrates the amount of protein used for the ATPase assay (175 ng fromN. benthamianaand 3.5g fromE. coli). These results show that 6H-MRep-M purified from plant tissue possesses comparable ATPase activity.
Similarly, origin DNA cleavage activity of 6H-MRep-M from N. benthamiana was compared to that of the protein purified fromE. coli(see Fig. S1 in the supplemental material). Both proteins were proficient in origin cleavage activity.
In planta interaction between FBNYV 6H-MRep-M and wild-type M-Rep.It had been reported that geminivirus Rep proteins form oligomers in solution (29, 33) and that the oli-gomerization state of Rep in vitro is pH dependent (15, 29). Further results suggested that an oligomeric complex may be essential for the initiation of DNA replication of geminiviruses (32). The nanovirus M-Rep protein also oligomerizes (T. Tim-chenko et al., unpublished results); however, and yet oligomer-ization was never shown in planta. Using the 6H-MRep-M protein, we demonstrated that interaction between tagged and wild-type proteins occurs in planta. Figure 7 (lanes 1 to 4) shows total protein extracts from N. benthamiana leaf disks agroinoculated with pBin19 (lane 1), DNA-R (lane 2), R-6H-M (lane 3), and coagroinoculated with R-6H-M and DNA-R (lane 4). The proteins eluted after incubation with the affinity resin are shown in lanes 5 to 8. M-Rep coelutes with 6H-MRep-M (Fig. 7, lane 8). M-Rep by itself was not bound by the resin (Fig. 7, lane 6) and was only detected in conjunction with 6H-MRep-M in extracts of plants expressing both wild-type M-Rep and 6H-MRep-M. This strongly suggests that both proteins interacted in planta, probably by forming mixed oli-gomers. Thus, the use of functional oligohistidine-tagged FBNYV master Rep variants allows isolation and purification of M-Rep protein complexes ex planta.
DISCUSSION
[image:7.585.299.542.67.225.2]Proteins involved in replication of ssDNA plant viruses have mostly been identified by virtue of their interaction with rep-lication initiator proteins by using the yeast two-hybrid system. In vitro pull down or immunoprecipitation of recombinant proteins confirmed complex formation by the respective inter-acting partners. For example, the interaction of the Wheat dwarf virus Rep protein with the large subunit of the wheat replication factor C has been observed (28). Moreover, it has been established that Tomato yellow leaf curl Sardinia virus (TYLCSV) Rep binds to the tomato proliferating cell nuclear FIG. 6. ATPase activity of plant- and E. coli-expressed
6H-MRep-M proteins. (A) 6H-6H-MRep-M purified fromN. benthamiana andE. coliwas incubated with␥-33P-labeled ATP. Liberation of33P
i
from␥-33P-labeled ATP by 6H-MRep-M was analyzed by thin-layer
[image:7.585.45.281.68.250.2]chromatography on PEI sheets. Lanes 1 and 4, no protein; lanes 2 and 5, 6H-MRep-M fromN. benthamiana; lanes 3 and 6, 6H-MRep-M fromE. coli. Lanes 1 to 3 indicate reactions performed at room tem-perature for 16 h; lanes 4 to 6 indicate reactions performed at 37°C for 45 min. (B) Amount of protein (175 ng of 6H-MRep-M from N. benthamiana and 3.5 g from E. coli) used for the ATPase assay analyzed by Western blotting.
FIG. 7. In planta interaction between FBNYV 6H-MRep-M and M-Rep. Total protein extracts (lanes 1 to 4) and eluted proteins after purification by IMAC (lanes 5 to 8) analyzed by Western blotting. Proteins fromN. benthamianaleaf disks agroinoculated with pBin19 (lanes 1 and 5), DNA-R (lanes 2 and 6), the R-6H-M replicon (lanes 3 and 7), and the R-6H-M replicon plus DNA-R (lanes 4 and 8) are shown. The positions of the tagged (6H-MRep-M) and wild-type (M-Rep) protein are indicated.
on November 8, 2019 by guest
http://jvi.asm.org/
antigen (9), and Tomato golden mosaic virus (TGMV) Rep binds to a retinoblastoma-related protein (1, 25). An N. benthamianasumoylation enzyme, NbSCE1, was also shown to interact with the Rep proteins from TYLCSV, TGMV, and African cassava mosaic virus-Kenia (10); curiously, the interac-tion site on Rep was mapped to a protein sequence that ex-tends over two opposite sides of the protein domain as re-vealed by its tertiary structure (8). This conundrum calls all the more for in planta confirmation of the complexes found by yeast two-hybrid assays and for a demonstration of the physi-ological relevance of such interactions. For example, in planta complex formation has been demonstrated for the nanovirus F-box protein Clink andMedicago sativaSKP1, a constituent of the ubiquitin-dependent protein turnover pathway (3).
The engineering of a functional Rep protein of an ssDNA plant virus, carrying a tag that allows its purification from the plant in the course of an infection, has been a challenge for the in planta study of gemini- and nanovirus replication. Hong et al. (20) showed that fusing green fluorescent protein (GFP) to the C terminus of ACMV Rep results in a protein with prop-erties similar to those of Rep with respect to viral DNA rep-lication and subcellular localization. The GFP tag permits vi-sual detection but no affinity purification. A functional replication initiator protein with such characteristics has not been reported for any ssDNA virus. Here we present the de-sign of an oligohistidine-tagged FBNYV M-Rep protein that is functional in planta and describe its use for the isolation and identification of proteins that interact in vivo with the tagged M-Rep. The tagged M-Rep was shown to trigger replication initiation inN. benthamianaof its cognate DNA component as well as that of other FBNYV DNAs.
The artificial replicon encoding the tagged M-Rep protein was able to spread throughout a plant when movement, en-capsidation, and other essential virus functions were provided by coinfection with a helper virus. Furthermore, when all es-sential virus FBNYV DNAs were introduced by agroinocula-tion along with the R-6H-M replicon, it was also transmitted by the aphid vector after superinfection with a helper virus. This proves that the R-6H-M replicon has all of the features of a genuine M-Rep-encoding artificial FBNYV genome component. In addition, the tagged protein was readily purified fromN. benthamianaleaves by native IMAC, and two of its key enzy-matic functions, the ATPase activity and the origin-specific ssDNA cleavage were demonstrated. To the best of our knowl-edge, this represents the first example of affinity purification of an enzymatically active ssDNA virus replication initiator pro-tein from the natural host of the virus. Moreover, we have shown the in planta interaction between an oligohistidine-tagged M-Rep and wild-type M-Rep, suggesting the formation of mixed protein oligomers. This will further allow the copu-rification of other viral and cellular partner proteins of M-Rep. Oligomerization is a common feature of replication initiator proteins, as has been shown for geminiviruses (32, 33) and replication-associated proteins of animal DNA viruses, such as the simian virus 40 large T antigen (36) and the adeno-associ-ated virus Rep78 protein (19).
The possibility that multifunctional Rep proteins are subject to posttranslational modifications to regulate their different functions is certainly possible. In the case of the SV40 large T antigen, some activities are regulated by phosphorylation (36).
Concerning Rep proteins of ssDNA plant viruses, there is very limited information about such modifications. Kong and Han-ley-Bowdoin (24) have shown that the TGMV Rep interacts with a protein kinase (GRIK), but no evidence of phosphory-lation of Rep in plant or insect cells was reported. Having at hand a tagged and functional M-Rep protein, the biological significance of phosphorylation or other protein modifications can now be readily studied in vivo.
Obtaining functionally intact modified replication initiator proteins of ssDNA viruses is difficult since, in the case of FBNYV, alteration of two amino acids at the N or C terminus of the M-Rep protein already abolishes its activity. The impor-tance of methionine 1 and arginine 3 for M-Rep is reflected by the fact that MRep-S is not functional, whereas 6H-MRep-M in which, apart from the tag, the wild-type amino acid sequence is conserved catalyzes replication initiation in planta. The lower replication level of the replicon R-6H-M compared to wild-type DNA-R (Fig. 2) indicates that the oli-gohistidine tag impairs to some extent the function of the protein. The replacement of methionine 1 by alanine in 6H-MRep-A or 4H-6H-MRep-A results in a further reduction of DNA replication. Whether the reduced activity of 6H-MRep-A and 4H-6H-MRep-A is solely due to the methionine 1 to alanine change or whether it is also influenced by the oligohis-tidine tag immediately preceding methionine 1 remains to be determined. Basic amino acids at the N terminus of the TYLCSV Rep have been suggested to be implicated in DNA recognition by the protein (8), an idea in line with the results presented here. An alternative or additional explanation for the observed reduction of DNA replication efficiency may be that important DNA-R elements required inciswere located in the sequence immediately preceding and/or encoding the amino terminus of M-Rep. The addition of the 27 nucleotides of the tag and base changes within the following sequence may interfere with the correct function of suchcis-acting elements. Experiments uncoupling M-Rep expression from cognate DNA (template) replication will provide distinctive information.
The ability of the DNA-R-His replicons to spread systemi-cally in the context of an infection with FBNYV indicates that the replicons move from cell to cell, either as DNA-protein complexes similar to geminivirus cell-to-cell movement (27) or as virions. The fact that we were able to amplify by IC-PCR the oligohistidine-tagged M-Rep encoding DNAs shows that they are at least tightly associated with virus capsid protein. Since the DNA-R-His replicons were transmitted by aphids, it is quite possible that they are also encapsidated into true virions. A tagged replication initiator protein of an ssDNA virus that is functional in planta represents a very useful tool for studying in planta protein-protein interactions and for identifying its viral and host partner proteins. It will also allow the study of host cell-dependent posttranslational modifications of Rep and its interacting partner proteins in the course of a virus infection.
ACKNOWLEDGMENTS
We thank H.-J. Vetten for providing monoclonal and polyclonal antibodies to FBNYV and polyclonal antibodies to M-Rep. We are indebted to L. Allala for providing the Algerian isolate of FBNYV and its vectorA. craccivoraand to the Cornell Research Foundation, Inc., forA. tumefaciensstrain COR308. We thank A.-L. Haenni, F. Ber-nardi, and two anonymous reviewers for valuable suggestions.
J.C.V.-A. was supported by a fellowship from CONACYT (Mexico).
on November 8, 2019 by guest
http://jvi.asm.org/
REFERENCES
1.Ach, R. A., T. Durfee, A. B. Miller, P. Taranto, L. Hanley-Bowdoin, P. C. Zambryski, and W. Gruissem.1997. RRB1 and RRB2 encode maize reti-noblastoma-related proteins that interact with a plant D-type cyclin and geminivirus replication protein. Mol. Cell. Biol.17:5077–5086.
2.Angulo, J. F., E. Rouer, A. Mazin, M. G. Mattei, A. Tissier, P. Horellou, R. Benarous, and R. Devoret.1991. Identification and expression of the cDNA of KIN17, a zinc-finger gene located on mouse chromosome 2, encoding a new DNA-binding protein. Nucleic Acids Res.19:5117–5123.
3.Aronson, M. N., A. Complainville, D. Clerot, H. Alcalde, L. Katul, H. J. Vetten, B. Gronenborn, and T. Timchenko.2002. In planta protein-protein interactions assessed using a nanovirus-based replication and expression system. Plant. J.31:767–775.
4.Aronson, M. N., A. D. Meyer, J. Gyorgyey, L. Katul, H. J. Vetten, B. Gronen-born, and T. Timchenko.2000. Clink, a nanovirus-encoded protein, binds both pRB and SKP1. J. Virol.74:2967–2972.
5.Boevink, P., P. W. Chu, and P. Keese.1995. Sequence of subterranean clover stunt virus DNA: affinities with the geminiviruses. Virology207:354–361. 6.Bradford, M. M.1976. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye bind-ing. Anal. Biochem.72:248–254.
7.Burns, T. M., R. M. Harding, and J. L. Dale.1995. The genome organization of banana bunchy top virus: analysis of six ssDNA components. J. Gen. Virol.
76:1471–1482.
8.Campos-Olivas, R., J. M. Louis, D. Clerot, B. Gronenborn, and A. M. Gronenborn.2002. The structure of a replication initiator unites diverse aspects of nucleic acid metabolism. Proc. Natl. Acad. Sci. USA99:10310– 10315.
9.Castillo, A. G., D. Collinet, S. Deret, A. Kashoggi, and E. R. Bejarano.2003. Dual interaction of plant PCNA with geminivirus replication accessory pro-tein (Ren) and viral replication propro-tein (Rep). Virology312:381–394. 10.Castillo, A. G., L. J. Kong, L. Hanley-Bowdoin, and E. R. Bejarano.2004.
Interaction between a geminivirus replication protein and the plant sumoy-lation system. J. Virol.78:2758–2769.
11.Desbiez, C., C. David, A. Mettouchi, J. Laufs, and B. Gronenborn.1995. Rep protein of tomato yellow leaf curl geminivirus has an ATPase activity re-quired for viral DNA replication. Proc. Natl. Acad. Sci. USA92:5640–5644. 12.Franz, A. W., F. van der Wilk, M. Verbeek, A. M. Dullemans, and J. F. van den Heuvel.1999.Faba bean necrotic yellows virus(genusNanovirus) requires a helper factor for its aphid transmission. Virology262:210–219. 13.Grimsley, N., B. Hohn, T. Hohn, and R. Walden.1986. “Agroninfection,” an
alternative route for viral infection of plants by using the Ti plasmid. Proc. Natl. Acad. Sci. USA83:3282–3286.
14.Grimsley, N., T. Hohn, J. W. Davies, and B. Hohn.1987.Agrobacterium -mediated delivery of infectious maize streak virus into maize plants. Nature
325:177–179.
15.Gutierrez, C., E. Ramirez-Parra, M. Mar Castellano, A. P. Sanz-Burgos, A. Luque, and R. Missich.2004. Geminivirus DNA replication and cell cycle interactions. Vet. Microbiol.98:111–119.
16.Hafner, G. J., R. M. Harding, and J. M. Dale.1997. A DNA primer asso-ciated with banana bunchy top virus. J. Gen. Virol.78:479–486.
17.Hamilton, C. M., A. Frary, C. Lewis, and S. D. Tanksley. 1996. Stable transfer of intact high molecular weight DNA into plant chromosomes. Proc. Natl. Acad. Sci. USA93:9975–9979.
18.Hanson, S. F., R. A. Hoogstraten, P. Ahlquist, R. L. Gilbertson, D. R. Russell, and D. P. Maxwell.1995. Mutational analysis of a putative NTP-binding domain in the replication-associated protein (AC1) of bean golden mosaic geminivirus. Virology211:1–9.
19.Hermonat, P. L., and R. B. Batchu.1997. The adeno-associated virus Rep78 major regulatory protein forms multimeric complexes and the domain for this activity is contained within the carboxy-half of the molecule. FEBS Lett.
401:180–184.
20.Hong, Y., J. Stanley, and R. van Wezel.2003. Novel system for the simulta-neous analysis of geminivirus DNA replication and plant interactions in
Nicotiana benthamiana. J. Virol.77:13315–13322.
21.Horsch, R., J. Fry, N. Hoffman, D. Eichholtz, S. Rogers, and R. Fraley.1985. A simple and general method for transferring genes into plants. Science
227:1229–1231.
22.Horser, C., R. Harding, and J. Dale.2001. Banana bunchy top nanovirus DNA-1 encodes the “master” replication initiation protein. J. Gen. Virol.
82:459–464.
23.Katul, L., T. Timchenko, B. Gronenborn, and H. J. Vetten.1998. Ten distinct
circular ssDNA components, four of which encode putative replication-associated proteins, are replication-associated with the Faba bean necrotic yellows virus genome. J. Gen. Virol.79:3101–3109.
24.Kong, L. J., and L. Hanley-Bowdoin.2002. A geminivirus replication protein interacts with a protein kinase and a motor protein that display different expression patterns during plant development and infection. Plant Cell14:
1817–1832.
25.Kong, L. J., B. M. Orozco, J. L. Roe, S. Nagar, S. Ou, H. S. Feiler, T. Durfee, A. B. Miller, W. Gruissem, D. Robertson, and L. Hanley-Bowdoin.2000. A geminivirus replication protein interacts with the retinoblastoma protein through a novel domain to determine symptoms and tissue specificity of infection in plants. EMBO J.19:3485–3495.
26.Laufs, J., I. Jupin, C. David, S. Schumacher, F. Heyraud-Nitschke, and B. Gronenborn.1995. Geminivirus replication: genetic and biochemical char-acterization of Rep protein function, a review. Biochimie77:765–773. 27.Lazarowitz, S. G., and R. N. Beachy.1999. Viral movement proteins as
probes for intracellular and intercellular trafficking in plants. Plant Cell
11:535–548.
28.Luque, A., A. P. Sanz-Burgos, E. Ramirez-Parra, M. M. Castellano, and C. Gutierrez.2002. Interaction of geminivirus Rep protein with replication factor C and its potential role during geminivirus DNA replication. Virology
302:83–94.
29.Missich, R., E. Ramirez-Parra, and C. Gutierrez.2000. Relationship of oligomerization to DNA binding of Wheat dwarf virus RepA and Rep proteins. Virology273:178–188.
30.Mozo, T., and P. J. Hooykaas.1991. Electroporation of megaplasmids into
Agrobacterium. Plant Mol. Biol.16:917–918.
31.Ooms, G., P. J. Hooykaas, R. J. Van Veen, P. Van Beelen, T. J. Regensburg-Tuink, and R. A. Schilperoort.1982. Octopine Ti-plasmid deletion mutants ofAgrobacterium tumefacienswith emphasis on the right side of the T-region. Plasmid7:15–29.
32.Orozco, B. M., L. J. Kong, L. A. Batts, S. Elledge, and L. Hanley-Bowdoin.
2000. The multifunctional character of a geminivirus replication protein is reflected by its complex oligomerization properties. J. Biol. Chem.275:6114– 6122.
33.Orozco, B. M., A. B. Miller, S. B. Settlage, and L. Hanley-Bowdoin.1997. Functional domains of a geminivirus replication protein. J. Biol. Chem.
272:9840–9846.
34.Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
35.Sano, Y., M. Wada, Y. Hashimoto, T. Matsumoto, and M. Kojima.1998. Sequences of ten circular ssDNA components associated with the milk vetch dwarf virus genome. J. Gen. Virol.79:3111–3118.
36.Simmons, D. T.2000. SV40 large T antigen functions in DNA replication and transformation. Adv. Virus. Res.55:75–134.
37.Stenger, D. C., G. N. Revington, M. C. Stevenson, and D. M. Bisaro.1991. Replicational release of geminivirus genomes from tandemly repeated cop-ies: evidence for rolling-circle replication of a plant viral DNA. Proc. Natl. Acad. Sci. USA88:8029–8033.
38.Studier, F. W., A. H. Rosenberg, J. J. Dunn, and J. W. Dubendorff.1990. Use of T7 RNA polymerase to direct expression of cloned genes. Methods En-zymol.185:60–89.
39.Timchenko, T., F. de Kouchkovsky, L. Katul, C. David, H. J. Vetten, and B. Gronenborn. 1999. A single rep protein initiates replication of multiple genome components of faba bean necrotic yellows virus, a single-stranded DNA virus of plants. J. Virol.73:10173–10182.
40.Timchenko, T., L. Katul, Y. Sano, F. de Kouchkovsky, H. J. Vetten, and B. Gronenborn.2000. The master rep concept in nanovirus replication: identi-fication of missing genome components and potential for natural genetic reassortment. Virology274:189–195.
41.Vetten, H. J., P. W. G. Chu, J. L. Dale, R. Harding, J. Hu, L. Katul, M. Kojima, J. W. Randles, Y. Sano, and J. E. Thomas.2004.Nanoviridae, p. 343–352.InC. M. Fauquet, M. A. Mayo, J. Maniloff, U. Desselberger, and L. A. Ball (ed.), Virus taxonomy: VIII Report of the ICTV. Elsevier/Aca-demic Press, London, England.
42.Wanitchakorn, R., G. J. Hafner, R. M. Harding, and J. L. Dale.2000. Functional analysis of proteins encoded by banana bunchy top virus DNA-4 to -6. J. Gen. Virol.81:299–306.
43.Wanitchakorn, R., R. M. Harding, and J. L. Dale.1997. Banana bunchy top virus DNA-3 encodes the viral coat protein. Arch. Virol.142:1673–1680.
on November 8, 2019 by guest
http://jvi.asm.org/
0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.16.10835.2005
ERRATUM
A Functional Histidine-Tagged Replication Initiator Protein:
Implications for the Study of Single-Stranded DNA
Virus Replication In Planta
Julio C. Vega-Arreguı´n, Tatiana Timchenko, Bruno Gronenborn,
and Bertha Cecilia Ramı´rez
Institut des Sciences du Ve´ge´tal, CNRS, 91198 Gif sur Yvette Cedex, France
Volume 79, no. 13, p. 8422–8430, 2005. Page 8427, column 1, first full paragraph, line 4: “R-GH-7” should read “R-6H-M.”