0022-538X/83/020489-07$02.00/0
Copyright ©1983,American Society forMicrobiology
Accumulation and
Breakdown of RNA-Deficient Intracellular
Virus
Particles in Interferon-Treated NIH 3T3 Cells
Chronically Producing
Moloney
Murine
Leukemia Virus
MORDECHAI ABOUD* AND YEHUDITH HASSAN
Departmentof Microbiology & Immunology, Faculty of Health Sciences, Ben Gurion University of the
Negev,BeerSheva,Israel
Received 27 August1982/Accepted 13 October 1982
Interferon treatment of NIH 3T3 cells chronically infected with Moloney
murine leukemia virus inhibited about95%of virus release. This inhibition was
accompanied byathree-totwofold accumulation of intracellularvirions. Howev-er,this accumulation could be demonstratedonly be exogenous reverse
transcrip-tasereactionassay orradioactivelabeling of the assembled viral proteins.Itcould
not be shown by the endogenous reverse transcriptase reaction assay, which
dependedonendogenousviralRNA, orby labeling the encapsidated viralRNA.
It was therefore evident that most ofthe intracellular virions accumulated in
interferon-treated cellswere RNAdeficient. Hybridization analysisrevealedthat
thesevirions weredeficient of genomic viral RNA, whereas size analysis by gel
electrophoresis suggested that the deficiency of 4S RNA normally packaged in
Moloneymurine leukemia viruswas evenstronger. Our data alsosuggestedthat
thisRNAdeficiencywasnotdueto adegradation ofthe encapsidated RNA, but morelikely to adefect in virus assembly. RNA-lacking intracellularvirionswere
unstable; theywerefoundto collapse before beingreleased.
TypeC retroviruses areusuallyassembledat
the inner side of the plasma membrane and
finally released from the cell surface by a
bud-dingprocess (13, 20, 30).Therefore, completed
virionscanbe detected in suchcasesonly
extra-cellularly. However, there are reportsofafew
exceptions. Forinstance, Friend leukemia virus
particleshave beenillustratedbyseveral
investi-gators to exist in cytoplasmic vacuoles of
di-methyl sulfoxide-activated erythroleukemic
cells (19, 23, 28, 35-37), in whichthey seem to
be stored until theyare expelled byopening of the vacuoles at the cell surface (28). Early
studies have shown vacuolartypeCparticlesof
Moloneymurine leukemia virus (MLV)inbone
marrowcells of leukemicrats(18)andparticles
of Moloney murine sarcoma virus in cells of
solid tumors of mice (17). Recently, we have
shown, by transmission electron microscopy,
the existenceinlarge numbers ofMLVparticles
incytoplasmic vacuoles ofthe NIH 3T3cellline
(2a, 6)and provided indicationsto suggest that
theyareformedintracellularly(1),apparently by budding intothespaceofthecytoplasmic
vacu-oles,wheretheyarekept untiltheirrelease from
the cells(M.Aboud etal., submittedfor
publica-tion).Onlyrarevirusparticleshave beenseen to
bud in the usual way through the outer
mem-brane of these cells. Furthermore, we have shown thatinterferon (IFN) treatment strongly
inhibitsvirus releasefromthesecells,leadingto
a remarkable accumulation of intracellular
viri-ons(2a,34).Thisfindingis instrikingcontrast to
the effect of IFN on Friend leukemia
virus-producing cells observed by Luftig et al. (28).
These authors have shown that, whereas control cells containedmanyvacuolarvirions,the
cyto-plasmicvacuolesof IFN-treated cellswere
emp-ty of virus particles, suggesting that in their
systemIFNinhibits the formationof the
vacuo-larvirions rather than their release.
However, in many repeated experiments we
havenoticed that IFN-inducedaccumulationof intracellular virions in our system was much
more apparent in thin-section electron
micro-scopic examinations or by measuring parame-ters representing assembled viral proteins than bymeasuring parameters representing
encapsi-dated viral RNA. Therefore, in this study we
carried out a systematic investigation of the
significanceof thisinconsistency and concluded
that mostoftheintracellular virions
accumulat-ed in IFN-treataccumulat-ed cells were RNA deficient.
Moreover, we presentdatasuggestingthatafter
alimitedstorageperiodtheRNA-lackingvirions
collapse within the cells, whereas the
RNA-489
on November 10, 2019 by guest
http://jvi.asm.org/
490 ABOUD AND HASSAN
containing virions seem to be rather stable and
apparently are released from the cells at a re-duced rate imposed by IFN.
MATERIALS AND METHODS
Cells and viruses. NIH 3T3mousefibroblasts chron-ically infected with MLV were used throughout this study. The cells were grown in Dulbecco modified Eagle mediumcontaining10% newborn calfserum.In experiments with IFN the serum concentration was reduced to2.5%.
IFN. Mouse IFN was prepared as described else-where (5). The IFN preparation used in this study contained 7x 106U/mgofproteinandwasappliedat a concentration of 80 U/ml.
Isolation of extracellular and intracellular virions. Extracellular virions were pelleted from the culture medium through a 20% sucrose cushion, after the medium was first cleared of cell debris. Thepelleted
virions were furtherpurifiedbybanding in a 15 to60%'o
sucrose gradient. To isolate intracellularvirions, the cells were thoroughly washed four times with cold
phosphate-bufferedsalinefor maximal removal of
re-sidualextracellular virions. The cellswerethen exten-sively treated withtrypsin-EDTAtofurtherdestroy any residue of extracellular virions. Although our previous electronmicroscopic studies (6; Aboud et al., in press) revealed that only rare virus particles were
externally associated with the outer membrane of
these cells,compared with thenumerousintracellular virions in cytoplasmic vacuoles, the trypsin-EDTA treatment was aimed also at removing these few external cell-associated virions, so that the isolated virusparticleswould be entirely of intracellular origin. Thecells werehomogenized,andapostmitochondrial
cytoplasmic fraction was prepared aspreviously
de-scribed(34). Thecytoplasmicfractionwastreated for 15 min with 5 mM EDTAtodissociatepolyribosomes
and further treated with 20 pLg of RNase A (Sigma
ChemicalCo.)perml for 30minat37°CtodigestRNA molecules which wereoutside of virusparticles. The
cytoplasmic virions werepelletedthrougha20%
su-crosecushion and then banded in 15 to60%sucrose
gradients. Unlabeled virionswerelocated in the
gradi-ent by reverse transcriptase assay ofeach fraction collected from the bottom of the gradient. Labeled virionswerelocatedbytrichloroaceticacid
precipita-tionofaliquots taken from each fraction of the
gradi-ent.
Reverse transcriptase assay. The exogenous reac-tion assay, using polyriboadenylate-oligothymidylate
[poly(rA) -oligo(dT)] as a template-primer, was
de-scribedby Aboudetal.(5). Theendogenous reaction assaywasperformedinthe same mannerexcept that poly(rA) -oligo(dT) wasomitted. In some
confirma-toryexperiments,75 ,ug ofactinomycinDper mlwas addedtotheendogenous reaction -mixture.
Radioactivelabeling. Cellswerelabeledwith 75,uCi
of[3H]uridine(50Ci/mmol; Nuclear Research Center, Negev, Israel)or 35 ,uCiof[3H]leucine 39Ci/mmol;
Nuclear ResearchCenter) perml.
Viral RNA extraction.Unlabeledorlabeledvirions, purified in a sucrose gradient, were lysed by 0.5% sodium dodecyl sulfate; RNA was extracted by a mixture of chloroform-isoamyl alcohol (24:1), and,
after50pLgof EscherichiacolitRNAper mlwasadded
as acarrier, RNA was concentrated by ethanol precip-itation.
Analysis ofgenomic RNA sequences. Concentrated viral RNA of unlabeled virions was centrifuged through a 15 to30% sucrose gradient; 3H-labeled 28S, 18S, and 4S RNA were used as size markers. Frac-tions were collected from the viral RNA-containing gradients, 50 ,ugof E. coli tRNA per ml was added to eachfraction, and RNA was precipitated from these fractions by ethanol and assayed for viral genomic RNA sequences by hybridization to MLV-specific
[3H]cDNAasdetailedby Salzberg et al. (34). By using purified 70S MLV RNA, this hybridization reaction was proved to belinear within a range of 15 to 65% hybridization.
Size analysis of viral RNA by gel electrophoresis. Equal amounts of[3H]uridine-labeled viral RNA (in termsof counts perminute) were boiled for 3 min with
0.5% sodium dodecyl sulfate and then analyzed by
1.5% agarose-1.8% polyacrylamide gel
electrophore-sis. The gel was sectioned into 2-mm slices, which wereextractedovernightby 0.5 mlof30%H202 and counted inInstagel scintillation liquid (Packard Instru-mentCo., Inc.), with correctionfor quenching of each sample.
RESULTS
Effect of IFN on reverse transcriptase activity
of intracellular virions. Virus-producing cells
were incubated for 48 h in the presence or
absence ofIFN. Extracellularvirions, released during this incubation period, were harvested from the culture medium and banded in 15 to
60% sucrosegradients. Each fraction collected
from thegradientswasassayed forreverse
tran-scriptase in exogenous reactions, using poly
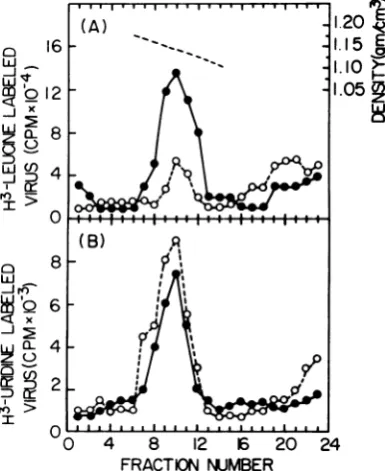
(rA) * oligo(dT)asanexogenous template-prim-er for virus quantitation. IFN inhibited virus releases by about95% (Fig.1A).
TodeterminetheeffectofIFN on the intracel-lular virions under these conditions, the cells
were disrupted and intracellular virions were
isolated in 15 to 60% sucrose gradients. An
exogenous reverse transcriptase reaction assay
of the gradient fractions (Fig. 1B) revealed a
three- to fourfold-higher activity in the viral
band of IFN-treated cells than in that of the
control. However, when the enzyme activity
wasassayed inendogenousreactions,no
signifi-cantdifference was observed(Fig. 1C). Similar results were obtained if actinomycin D was
added to the endogenous reaction mixture to
avoid DNA-dependent DNA synthesis, thus
eliminating a possible argument that cellular
DNApolymerasesweremeasured in this
experi-ment. Apossibleexplanation for this
discrepan-cyisthat theincreasedactivity ofthe
intracellu-lar virions ofIFN-treated cells detected in the
exogenousreactionrepresents ahigher
accumu-lation of virus particles, but most of them are
probably deficient of viral RNA, which is
re-quired for measuring the enzyme activity in
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
J(L)
-x
Q~o
a-Jx cr
Z5J
(I)
-JQ
3r x
J- 2.
a: L
4 31
i.20 f
1.15H 1.10
c-1.05ZH
0 4 8 12 16 20 24
FRACTION NUMABER
FIG. 1. Reversetranscriptase activity of extracel-lular and intracelextracel-lular virions of IFN-treated and
un-treated cells. Virus-producingcells weretreated with
80 U of IFNpermlfor 48 h(S).Untreated cells served
asacontrol(0).Extracellular virionswereharvested from the culture medium(A),and intracellular virions
were harvested from the postmitochondrial cytoplas-mic fractions of the cells (B and C). Virions were banded in 15 to 60% sucrose gradients. Fractions, collected from the gradients, were assayed for viral
reversetranscriptase activityinexogenous(AandB) andendogenous (C)reactions.
endogenous reactions. Another possible
expla-nation is based on the observation of
Bandyo-padhyay et al. (8) that IFN may interfere with
the formation ofapropercomplex between p30
andreversetranscriptase, which is important for
viral RNA-directed but not for poly(rA) *
oli-go(dT)-directed enzymeactivity. Such a
possi-ble defect in the intracellular virions of
IFN-treated cells would indeed make the virus
accumulation inthese cells much easierto
dem-onstrateby exogenous than by endogenous
en-zyme reactions. A third explanation could be
that thehigher activity of virions of IFN-treated
cellsmayreflectanincreased numberofenzyme
molecules per virion rather than virus
accumula-tion. This increased amountof enzyme is readily
detectable by exogenous reactions, inwhich a
vast excessoftemplate-primer is used.
Howev-er,since onlyone enzymemoleculepervirion is
supposedly involved in endogenous reactions,
this increased amount of enzyme per virion cannot be detected in such reaction conditions.
Effect of IFN on radioactivelabeling of
intracel-lular virions. To clarify which of the
above-mentioned possibilities indeed accounts for the discrepancy shown in Fig. 1B and C,
virus-producing cellswerelabeled for 48hwitheither
[3H]leucine or [3H]uridine in the presence or
absence of IFN. Labeled intracellular virions
were pelleted from the cytoplasmic fraction of
the cells and banded in 15 to 60% sucrose
gradients. Aliquots of each fraction of the
gradi-ent wereassayed for trichloroacetic
acid-precip-itableradioactivity. Figure 2A showsa remark-able accumulation of [3H]leucine-labeled intracellular virions in IFN-treated cells. This
finding eliminates the possibility that IFN treat-ment results in anincreased number ofreverse
transcriptase molecules per virion rather than virus accumulation, because such an increase wouldnotbe detected by[3H]leucine labeling of
o 12
J
z- 8
U) cr
'r
-r')
IJ
xQ
E
cj Cl >)
ESI
e
1.20 E 1.15 E 1.10
>--1.05
2
8
0 4 8 12 6 20 24
FRACTON NUMBER
FIG. 2. [3H]leucineand[3H]uridine labelingof
in-tracellular virions of IFN-treatedand untreated cells. Cells were labeled with [3H]leucine (A) and
[3H]uri-dine(B)for 48 h in thepresence(0)orabsence(0)of
IFN.Intracellular virions werepurifiedin 15 to60%o
sucrose gradients, and aliquots from each fraction,
collected from the gradients, were assayed for tri-chloroaceticacid-precipitableradioactivity.
(A xogenous Reaction
-I
I
II.,... ... ..,..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.496.53.244.89.418.2] [image:3.496.259.452.370.606.2]492 ABOUD AND HASSAN
virions. On the other hand, thisaccumulation of
intracellular virions could not be demonstrated in this particular experiment by [3H]uridine la-beling. It is therefore strongly suggested that the inabilitytodemonstrate intracellular virus
accu-mulationin IFN-treated cells by the endogenous
reverse transcriptase reactions wasdueto
defi-ciency of viral RNA rather thanto adefect in the
complex formation between p30 and reverse
transcriptase, although the latterpossibilitywas notdirectly examined and therefore couldnotbe totally excluded.
Effect ofIFN on viralgenomicRNA contentof intracellular virions. Sincereversetranscriptase
activity, measured inendogenousreactions,
de-pendsonboth thegenomicviral RNAservingas atemplate andon a primertRNA, the reduced endogenousenzymatic activityof the virions of IFN-treated cells could resultfromdeficiencyof
anyof thesevirus-packaged RNAspecies. Also,
the results of [3H]uridine labeling could not
provide any indication to suggest which of the RNA species was missing in the assembled
virions. To clarify this problem, the unlabeled
virions banded inthe sucrosegradients
present-ed in Fig. 1 were pelleted, and viral RNA was
extracted by chloroform-isoamyl alcohol and concentrated by ethanol precipitation. This
RNA was centrifuged through a 15 to 30%
sucrosegradient. Fractions werecollected and,
afteradditionof 50,gofE. coli tRNApermlas acarrier, theRNAof each fractionwas
concen-tratedby ethanol precipitation and analyzed for viralgenomic RNA sequences by hybridization
to MLV-specific
[3H]cDNA.
A major 70S peakofhybridizable RNA wasfound together witha
broad spectrum of smaller hybridizable RNA molecules, which are probably a degradation product of the 70S RNA (Fig. 3). More
impor-tant is that, although the amount of virions collected from thegradientof IFN-treated cells
was morethan threefold higher than that of the untreatedcontrol,nosignificantquantitative dif-ference could be detected between the
hybridi-zable RNA of the virions of IFN-treated and
untreated cells. It is therefore evident that the RNAdeficiency of intracellular virions of
IFN-treated cellsinvolves thegenomic viralRNA.
Effect of IFN on 4S RNAcontent of
intracellu-lar virions. To determine whether the RNA
deficiency of intracellular virions of IFN-treated
cells was restricted only to the viral genome,
RNA wasextractedfromthe
[3H]uridine-labeled
virionsof the experiment presented in Fig. 2B,
and the same amounts ofRNA (in counts per
minute) from virionsofIFN-treated and untreat-edcells wered analyzed by gelelectrophoresis.
Figure 4 shows that the ratio of 4S RNA/35S
RNA ofthe controlvirions(Fig. 4A)was consid-erably higher than that of virions from
IFN-treated cells (Fig. 4B). Since RNA of both
virions was similarly treated, it is unlikely that
this difference can be attributed tovariation in
degradation of 35S RNA during its preparation.
Since similar results were obtained in several
repeated experiments, it seems that IFN treat-ment results in deficiency of 4S RNA that is
evenstrongerthan thatof the viralgenome.
Fate of intracellular virions in IFN-treated and
untreated cells.Tofollow the fate of intracellular
virions underconditions ofanIFN-induced anti-viralstate,cellswerelabeledfor24hwitheither
[3H]leucine or [3H]uridine in the presence or
absence ofIFN. At the endof thislabeling, the cells were washed four times and further
incu-bated with nonradioactive medium. During this
additionalincubation,IFNcontinued to be
pres-ent in the medium of IFN-treated cultures. At
varioustimes afterremovaloflabels, extracellu-lar andintracellular labeled virionswereisolated
bysucrosegradient bandingandquantitated by
trichloroaceticacidprecipitation. Total incorpo-ration of labelsinto macromoleculeswas
deter-mined by trichloroacetic acid precipitation of cellaliquots. No increase in radioactive labeling
of cellular macromolecules occurred after label removal (Fig. 5A). Moreover, the cytoplasmic pool sizes of both labels were determined by
4°- 70S
28
S18
S40
z
30-~0
20
ol
0 4 8 12 16 20
FRACTION NUMBER
FIG. 3. Viral genome sequenceanalysis of intracel-lular virions of IFN-treated (0) and untreated (0)
cells. Unlabeled intracellular virions, banded in the
gradients presented in Fig. 1,werepelleted, and their RNAwasextractedbychloroform-isoamylalcohol and concentratedby ethanolprecipitation. TheRNA was
centrifugedthrough15 to30%osucrosegradients. Each
fraction, collected from the gradients, received50,ug of E. coli tRNA per ml and was precipitated with ethanol; the precipitates were hybridized with 2,500 cpmofMoloneyMLV-specific
[3H]cDNA.
Theextent of hybridization was determined by the fraction ofinput [3H]cDNA rendered resistant to Si nuclease.
This [3H]cDNA showed 2.7% self-annealing, which
wassubtractedfrom each value.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.496.263.434.387.545.2]RNA-LACKING CELLS
°x
200~4
I I I I> (B)35S
04 28S 18S 4S
3-2
0
0 10 20 30 40 50 60 70 80
SLICE NUMBER
FIG. 4. Sizeanalysisof RNAencapsidatedin intra-cellular virions ofIFN-treated and untreated cells. [3H]uridine-labeled intracellular virions, bandedinthe gradients shown in Fig. 2B, were sedimented, and RNAwasextracted asinthe legend toFig. 3. Equal
amounts of labeled RNA (in terms of counts per minute) from control virions (A)and from virions of IFN-treated cells (B)weresubjectedtogel electropho-resisasdescribedinthetext.The gelsweresectioned into 2-mmslices, whichwereextractedby 30% H202 andcounted inInstagel scintillation liquid,with
cor-rection forquenching.
directfiltering of cell aliquots without
trichloro-acetic acid treatment and subtracting from the
radioactivity of these aliquots the values of
trichloroaceticacid-precipitable radioactivity of
parallel aliqots. It was found that these pools
were promptly depleted after the cells were
washed(datanotshown). It is thusapparentthat
this wash was efficient for stopping further
sig-nificant incorporation of both labels into
pro-teins or RNA. Therefore, even if some new
labele&
virions ofprelabeledprecursorscontin-ued to be formed after removal of labels, they
would constitute only a small fraction of the
overall number of radioactive intracellular
viri-ons and therefore wouldnotraise serious
com-plications in interpreting the results. Both
[3H]uridine- and [3H]leucine-labeled
intracellu-lar virionsdisappeared from the control cellsata
rapid but approximately equal rate, which was
accompanied by an almost parallel appearance
of extracellular labeled virions (Fig. 5B and C).
(The appearance of only [3H]leucine-labeled
extracellular virionsisshowninFig. SD.) On the
other hand, in IFN-treated cells,
[3H]uridine-labeled intracellular virions disappeared at a
very slow rate (Fig. 5B), probably as a result of
the IFN-induced inhibition of their release.
However, the [3H]leucine-labeled virions dropped in these cells suddenly to about 35% of their initial level after a limited period of steady
state, and then they continued to decline at a
rate approximately equal to that of
[3H]uridine-labeled virions (Fig. 5C). It should be
empha-sized thatthe sudden fall in [3H]leucine-labeled
(A)
z15
9 15
M
6cr8
100~~~~~
~'(B)
15£
5Q 0
> 0~~~~~
3f~0(5
3
0
cc
cr7
a-0
(D)
20~~~
'~i
AA0
0 2 4 6 8 10
X TME AFTERLABEL REMMAL(h)
FIG. 5. Fateof labeled intracellular virions of IFN-treated and untreated cells. Cells were labeled with
[3H]uridine and [3H]leucinefor 24 h in the presence
(A)orabsence(0)of IFN. At the end of labeling the cellswerewashedfour times and received unlabeled fresh medium, with IFN addedtothemedium of the IFN-treated cultures. Atvarious times of incubation with the unlabeled medium, incorporation of labels into total trichloroacetic acid-precipitable materials wasdetermined in cell aliquots (A). Intracellular la-beled virionswere harvested from the
postmitochon-drial fraction of thecells(B andC) and extracellular virions wereharvested from the culture medium(D) andpurified by 15 to60%osucrose gradients. Values arethe total trichloroaceticacid-precipitable radioac-tivity in the 1.12- to 1.16-g/cm3band of these
gradi-ents.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.56.251.69.310.2] [image:5.496.262.449.210.525.2]494 ABOUD AND HASSAN
intracellularvirions ofIFN-treated cells wasnot
accompanied by acorresponding appearance of
labeled extracellular virions (Fig. SD). These
findings strongly suggest that most of the
RNA-deficient intracellular virions accumulated with-in IFN-treated cells collapse before being
re-leased from the cells. Furthermore, the slow
decline of[3H]uridine-labeled intracellular
viri-ons of IFN-treated cells, which was almost
parallel to the rate of virus release from these
cells,indicates thatthe RNAdeficiencyofthese
virionswas not due to itsdegradationafter being
encapsidated, butmorelikelyto adefectinvirus
assembly.
DISCUSSION
IFNis knowntointerfere with the replication of retroviruses(9,21).However,thetargetofits
actionseemsto varyfrom onesystem to anoth-er.Forinstance, inexogenousinfections, it has been found to inhibit someearly events before theintegration oftheproviralDNAintothehost
genome (2-4, 7, 31). In chronically infected
cells, IFN has been found to interfere with
various late steps, such as posttranslational
processing of viral precursor proteins (14, 29,
33,39),virusassembly (8, 15,32,38),oritsfinal release from the cell surface (10-12, 15). Since unlike mostothersystemsofretrovirus-infected cellsMoloneyMLVtypeCparticleswerefound
to be formed intracellularly in our chronically infected NIH 3T3 cells, apparently by budding into cytoplasmicvacuoles(2a), it was ofinterest
toinvestigate in detailthe effectof IFN on our
system. Inaccordancewith ourpreviousreports
(2a, 34), IFNwasfoundtostrongly inhibit virus release, and this inhibitionwasaccompanied by aremarkable accumulation of intracellularvirus
particles. However, our present data
demon-stratethatmostoftheintracellular virions
accu-mulating in IFN-treated cellsareRNAdeficient. This deficiency does not seem to result from a
degradation of the encapsidated RNA during
storage ofthe intracellular virions within
IFN-treated cells, but more likely from a defect in
virusassembly. It is interesting to mention that
formation ofRNA-deficient MLV particles has
been observed also in actinomycin D-treated,
virus-producing cells (22, 24-27). However,
whereas the RNAdeficiency observed in these
cells could be attributedtotheinhibition of viral
RNAsynthesis,in our cells IFN has beenfound
to have no effect on viral RNA synthesis (1).
RNAdeficiencyin oursystem involvedgenomic
viral RNA. Moreover, unlike the actinomycin
D-induceddeficiency, which is restricted to the
70S viral RNA(25, 26), thedeficiency imposed
in our systemby IFN was even stronger for the
small RNA species usually packaged in
retro-virus particles. Nevertheless, this uneven
defi-ciencyof the various RNAspeciesiscompatible withthe observationof Levin and Seidman(26)
thatpackaging oftRNA in MLV isindependent
of that of viral genomic RNA. Assembly of
reverse transcriptase invirus particleshas been
found also to be independent of packaging of
viral RNA (22, 27). This could be the reasonfor
our ability to demonstrate the accumulation of
intracellular virionsin IFN-treated cellsby
mea-suring their reverse transcriptase activity in an
exogenousreaction despite their RNA
deficien-cy. However,thisRNAdeficiencycould lead to
someadditionaldefects in the morphogenesisof
the virions,as reported by Jamjoomet al. (24).
Thesedefects could accountfor the breakdown
of the RNA-deficient intracellular virions
ob-served in ourIFN-treated cells. It isimportant
to notethat our data suggest that
RNA-contain-ing intracellular virions are rather stable, and it
is quite possible that the residual virus release
from IFN-treated cells involvesonlythiskindof
particle. Support for this proposal can be
provid-ed by our earlier observation (5) that IFN
inhib-its the relase of reversetranscriptase-containing
virus particles andinfectious virions to more or
less the same extent.
ACKNOWLEDGMENTS
Thisstudy wassupported by a grantfromtheChernow and StreinFoundation oftheIsraelCancerResearch Fund, New York,N.Y.
LITERATURE CITED
1. Aboud, M., R. Kimchi, M.Bakhanashvlli,andS. Salzberg. 1981.Intracellular production of virus particlesandviral components in NIH/3T3 cells chronically infected with Moloney murine leukemia virus: effect ofinterferon.J. Virol. 40:830-838.
2. Aboud, M., D. Moldovan-Levin, and S. Salzberg. 1981. CytoplasmicviralDNAsynthesis inexogenousinfection ofmurine leukemia virus: effect of interferonand cyclo-heximide. J. Virol. 37:836-839.
2a.Aboud, M.,R.Shoor,S.Bari,Y.Hassan,Z.Malik,andS. Salzberg.1982. Biochemicalanalysisand electron micro-scopic study onintracellular virions in NIH/3T3 mouse cellschronicallyinfected withMoloneymurine leukemia virus:effectof interferon. J. Gen. Virol.62:219-225. 3. Aboud, M., R. Shoor, and S. Salzberg. 1978. Effect of
interferononexogenousmurine leukemia virusinfection. Virology 84:134-141.
4. Aboud,M.,R.Shoor,andS.Salzberg.1980. Aneffectof interferonontheuncoatingof murine leukemiaVirusnot relatedtotheantiviralstate.J.Gen. Virol.51:425-429. 5. Aboud, M., 0. Weiss, and S. Salzberg. 1976. A rapid
quantitation of interferon withchronically oncornavirus-producingcells. Infect. Immun. 13:1626-1632.
6. Aboud, M., M. Wolfson, Y. Hassan, and M. Huleihel. 1982.Rapidpurification ofextracellular and intracellular Moloneymurineleukemiavirus. Arch.Virol.71:185-1%. 7. Avery, R.J., J.D.Norton, J.S.Jones,D.C.Burk,and A.G. Morris. 1980. Interferon inhibits transformationby murine sarcoma virus before integration of provirus. Nature(London)288:93-95.
8. Bandyopadhyay, A. K., E. H. Chang, C. C. Levi, and R. M. Friedman.1979.Structural abnormalitiesin murine leukemia viruses produced by interferon treated cells. Biochem. Biophys.Res. Commun. 87:983-988. 9. Billiau, A. 1977. Effect of interferon on RNA tumor
viruses. Tex.Rep.Biol. Med. 35:406-419.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
10. Billiau, A., V. G.Edy, H.Sobis,and P. DeSomer.1974.
Influence of interferon on virus particles synthesis in
oncornaviruscarrier lines. II. Evidence foradirect effect onparticles release. Int. J.Cancer 14:335-340. 11.Bilhlau, A., H. Heremans, P. T. Aflen, J. De
Maeyer-Guingard, and P. DeSomer. 1976. Trappingof oncorna-virus particles at the surface ofinterferon treated cells. Virology73:537-542.
12. Bilhlau,A., H. Heremans, P. T. Allen, and P. DeSomer. 1976. Influence of interferon on the synthesis of virus particles inoncornavirus carrier lines. IV. Relevance to potentialapplicationofinterferon in infectious diseases. J.Infect.Dis. 133:A51-A55.
13. Bishop,M.J. 1978. Retroviruses. Annu. Rev. Biochem. 47:35-88.
14.Chang, E. H., and R. M.Friedman. 1977. Alarge glyco-protein of Moloneymurine leukemia virusderivedfrom interferon treated cells. Biochem. Biophys. Res.
Com-mun.77:392-397.
15. Chang, E. H., M. W.Mayers, P. K. Y. Wong,andR.M. Friedman. 1977. Theinhibitoryeffect of interferonona temperature-sensitive mutant ofMoloney murine leuke-miavirus.Virology77:625-636.
16. Chang, E. H., S. H.Mis,T.J.Triche,andR. M. Fried-man. 1977.Interferon inhibits mouseleukemia virus re-lease: an electron microscopic study. J. Gen. Virol. 34:363-367.
17. Dalton, A. J. 1966. Anelectron microscopic study of a virus induced murine sarcoma (Moloney).Natl. Cancer Inst. Monogr.22:143-168.
18. Dalton, A. J., F. Haguenau, and J. B. Moloney. 1964. Further electronmicroscopicstudies on themorphology of the Moloney agent. J. Natl. Cancer Inst. 33:255-275. 19.De Harven, E., and C. Friend. 1966. Origin of viremia in
murine leukemia.Natl. Cancer Inst. Monogr. 22:79-105. 20. Fan, H. 1978. Expression of RNA tumor viruses at the
translationandtranscription levels.Curr.Top.Microbiol. Immunol.79:1-41.
21. Friedman,R. M. 1977. Antiviral activity of interferon.
Bacteriol.Rev.41:543-567.
22. Gerwin, B. I., and J. G. Levin. 1977. Interactions of murineleukemiavirus corecomponents: characterization ofreversetranscriptase packaged in the absence of 70S
genomicRNA. J. Virol.24:478-488.
23. Ikawa,Y., M.Furusawa, and H. Sugano. 1973.
Erythro-cytic membrane specific antigensin Friend virus induced
leukemiacells.Bibl. Haematol (Basel)39:955-967.
24.Jamjoom, G. A., R. B.Naso, and R. B.Arlinghaus.1976.
Selective decrease inthe rateofcleavageof an intracellu-lar precursor to Rauscher leukemia virusp30bytreatment ofinfectedcells withactinomycinD.J. Virol. 19:1054-1072.
25. Levin, J. G., P. M. Grimley, R. M.Ramseur, and I. K. Berezesky. 1974.Defficiencyof60 to 70S RNA in murine
leukemia virus particles assembled in cells treatedwith
actinomycinD.J. Virol. 14:152-161.
26. Levin, J.G.,andJ. G. Seidman. 1979. Selective packag-ingofhost tRNAbymurineleukemia virusparticlesdoes notrequire genomicRNA. J. Virol.29:328-335. 27. Levin,J. G.,andJ.G.Seldnan. 1981. Effect of
polymer-ase mutations on packaging ofprimer tRNAP° during
murineleukemia virusassembly.J.Virol.38:403-408. 28. Luftig,R.B., J.F.Conscience,A.Skoultchi,P.McMillan,
M.Revel,andF.H. Ruddle.1977. Effect ofinterferonon dimethyl sulfoxide-stimulated Friend erythroleukemic
cells: ultrastructural and biochemical study. J. Virol.
23:799-810.
29. Maheshwari, R.K., D. K.Banerjee, C.J. Waechter, K.
Olden, and R.M. Friedman. 1980. Interferon inhibits
glycosylationof viralprotein.Nature(London)
287:454-458.
30. Monteralo, R.C., and D. P. Bolognesl. 1978. Structure andmorphogenesisoftype-Cretroviruses. Adv. Cancer Res. 28:63-89.
31. Moris, A. G., and D. C. Burk. 1979. An interferon sensi-tiveearlystep in theestablishment ofinfection of murine
sarcoma/leukemia virus. J. Gen. Virol.43:173-181. 32. Pltha, P. M., B. Fernie, F. Maldarelli, T. Hattman, and
N.A. Wivel. 1979. The error inassembly of MuLV in interferon-treated cells. J. Supramol. Struct. 12:35-46. 33. Pltha, P. M., B.Fernie, F.Maldarell, T. Hattman, and
N.A.Wivel.1980. Effect of interferon on mouse leukemia virus (MuLV). V. Abnormal proteins in virions of Rauscher MuLV produced in the presence of interferon. J.Gen. Virol. 46:97-110.
34.Salzberg, S., M. ahshvili, S. Bari, I.Berman,and M. Aboud. 1980. Characterization ofintracellular viral RNA ininterferon-treated cells chronically infected with murine leukemia virus. J. Virol.35:694-703.
35. Sato,T., C.Friend,andE. DeHarven.1971. Ultrastructur-alchanges in Friend erythroleukemia cells treated with dimethylsulfoxide. Cancer Res.31:1402-1417.
36. Sato, T., C.Friend, C.Stackpole, and E. De Harven. 1972. Coating of Friend leukemia virus after treatment with specific antiserum. Cancer Res.32:2670-2678.
37. Sugano, H., M.Furusawa,T.Kawaguchi,and Y.Ikawa. 1973. Enhancement oferythrocyticmaturation of Friend virus induced leukemia cells in vitro. Bibl. Hematol. (Basel) 39:943-954.
38. Van-Griensvan, L. J., M. E. Baudelaire, J. Peries, R.
Emanoll-Ravicovitch,and M. Bolron. 1971. Studies on the biosynthesis of murine leukemia virus RNA. Some prop-erties Rauscher leukemia virus from interferon treated JLSV cells, p. 145-153. In Le Petit coloquia. North-Holland, Amsterdam.
39. Yagi, M. J., N. W.King,Jr., and G. Bekesd. 1980. Alter-ations of mousemammary tumor virusglycoproteinwith interferon treatment. J. Virol. 34:225-233.
VOL.45,1983
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG. 4.gradientsamountsandresiscellularRNAminute)intorectionIFN-treated[3H]uridine-labeled Size analysis ofRNA encapsidated in intra- virions of IFN-treated and untreated cells](https://thumb-us.123doks.com/thumbv2/123dok_us/1449198.97442/5.496.56.251.69.310/gradientsamountsandresiscellularrnaminute-intorectionifn-treated-uridine-labeled-analysis-encapsidated-untreated.webp)