0022-538X/83/010073-07$02.00/0
Copyright©1982, AmericanSocietyforMicrobiology
Genomic
Structure
of
Human
Polyoma Virus JC: Nucleotide
Sequence
of the Region
Containing Replication Origin and
Small-T-Antigen
Gene
TATSUOMIYAMURA,'HIROYUKIJIKUYA,2EIICHI SOEDA,2ANDKUNITOYOSHIIKEl*
DepartmentofEnteroviruses, National Institute of Health, Tokyo141,1and Departmentof Molecular
Genetics,National InstituteofGenetics, Misima
41J2
JapanReceived 5 August 1982/Accepted 23 September 1982
The nucleotide sequence of the region of human polyoma virus JC DNA
between 0.5 and0.7mapunitsfromaunique EcoRI cleavage sitewasdetermined and compared with thoseof the corresponding regions of another human polyoma virus, BK, andsimian virus 40DNAs. Within this region consisting of 945 base pairs, we located the origin ofDNA replication near 0.7 mapunits, the entire coding region for small T antigen, and the splice junctions for large-T-antigen mRNA. Thededuced amino acid sequences for smallTantigen and the partof largeTantigen markedly resembled those of polyoma virus BK and simian virus 40.The results stronglysuggestthatpolyomavirus JC has thesameorganization ofearly genome aspolyomavirus BKand simian virus40 on the physicalmap,
with theEcoRI site as areference point.
Human polyoma virus JC (JCV) has been isolated from brain tissues ofpatients with
pro-gressive multifocal leukoencephalopathy,arare
demyelinating disease, forwhich JCV is proba-bly responsible as an etiological agent (9, 19). Serological surveys have revealed thatJCV in-fection iscommonamonghumanpopulations(2, 18). Like other polyoma viruses, JCV has
tu-morigenic andtransforming capacities(7, 20, 33, 35).Therefore, studies of JCVareboth etiologi-callyandbiologically important.
Biological and biochemical studies on JCV, however, have been greatly hampered because JCVcangrowefficiently onlyinprimaryhuman fetalgliacell cultures rich in spongioblasts (17) andbecause viral DNA from the infected cells is usually heterogeneous in size (12, 16). Although JCV has been successfully adaptedtogrowth in
more readily available cells, such as human amnion cells (31) andhumanembryonic kidney (HEK) cells (14), the viral DNA thus obtained is heterogeneous. A JCV mutant growing well in HEKcellshasbeen foundtocontaintwoclasses
of defective DNAs, whichprobably complement
each otherforviralgrowth (38).
Molecularcloning of JCVDNAinabacterial
vector(10)from DNAobtainedby alow
multi-plicity of infection of primary human fetal glia
cells (12) hasnowmade detailed analyses ofthe
JCVgenome possible. Inthepresent study, we
determined the nucleotide sequence of the
re-gion near 0.7 map units fromEcoRI site in an
attempt to correlate the physical map of JCV
DNA withthe genomic organization. From the
similarity to simian virus 40(SV40) and human polyoma virus BK (BKV) DNAs, we could locate thereplication origin and the entire cod-ing region for the smallTantigen. Alsofound in
thisregionwerethesignal sequencesconserved
nearthe splicing sites of eucaryoticmRNA.
MATERIALSANDMETHODS
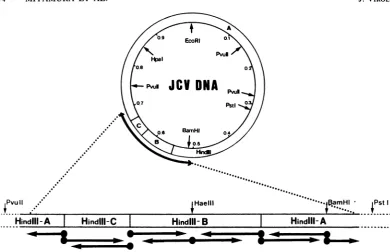
JCVDNA. Arestriction endonucleasecleavagemap of JCV(Mad-1)DNA(10, 12,22)is shown inFig.1. A recombinantplasmid,pJC(1-4), which hasaJCV DNA insertatthe BamHI siteofplasmid pBR322(10),was
used. DNA was extracted andpurified as described previously (38).
Enzymes. Restriction endonucleases BamHI,
HaeIII, HindIll, HpaI, PstI, andPvuII were
gener-ously provided by Takara Shuzo Co., Ltd., Kyoto,
Japan, andwereusedasrecommendedbythe
manu-facturer. Bacterial alkalinephosphatase and T4 poly-nucleotide kinase were purchased from Boehringei MannheimBiochemicals, Indianapolis,Ind.
DNAsequencing.Nucleotide sequenceswere deter-minedby the method ofMaxamand Gilbert(13) witha
strategyillustrated inFig. 1. pJC(1-4) DNAdigested
withHindIII,BamHI, HpaI,andPstIwassubjectedto
preparative electrophoresis in 5.5% polyacrylamide
gels. Afterelution, HindIII-B and the shorter
HpaI-cleavedHindIII-Awerefurther cleaved each intotwo
subfragments with HaeIII and PvuII, respectively.
Double-stranded fragments isolated from gels were
labeled at their 5' ends with [y-32PIATP (specific
activity, 2,000 Ci/mmol, Amersham International)
andT4polynucleotidekinase. Thelabeled DNA was
denatured by heatandseparated into eachstrand on
73
on November 10, 2019 by guest
http://jvi.asm.org/
74 MIYAMURA ET AL.
PvujU
PstI
PvuII .-... .I...
HindIll-A
I
HindIll-C Hindi^_-
-~---*w *- -0 I
lPstI
FIG. 1. Restriction endonuclease cleavage map of JCV(Mad-1) DNA and schematic representation of the strategy forsequencing. Thecleavagesitesarefrompublisheddata(10,12, 22).Thestraightarrowsindicate the direction andapproximateextentof sequence determination from the indicated restriction enzyme sites.
5% polyacrylamide-0.1% bisacrylamide gels. Frag-mentslabeled atoneendweresubmitted tothe four setsof partialdegradation reaction of A > C, G, C, andC+T.Thedegradation productswererunon
0.5-mm-thick 12%polyacrylamide-0.62% bisacrylamide-7
Mureagels.Autoradiographywasdoneat-70°Cwith
anintensifyingscreen (KodakX-Omatic).
RESULTS
LikeSV40 and BKVDNAs, JCV DNA hasa
unique EcoRI site that can be used as a
refer-ence point (Fig. 1). Cross-hybridization under
lessstringent conditions between restriction
en-donuclease cleavage fragments of these three
polyoma viruses has shown that theirphysical
maps could be colinearly aligned by using the
EcoRIsiteasthe0mapposition (11). To locate
the replication origin precisely on the physical
mapofJCVDNA,wedetermined the nucleotide
sequenceof theareapresumably containing the
replication origin and coding for 5' end of the
early mRNAs and compared it with the
corre-spondingareasof BKVandSV40 DNAs, whose
entire sequences and relation to the genetic
maps have been established (5, 21, 24, 37).
Figure 2 shows the nucleotide sequence of 945
basepairscovering theareafrom0.5to0.7map
units.
Theorigin of JCV DNA replicationwas
locat-edneartheHindIII-A/C junction (Fig. 1). Since
the origin of SV40 DNA replication has been
confinedwithinanarrowregion around the BglI
cleavage site(8, 25), the similar nucleotide
se-quence to that region was searched for in the
JCV DNA sequence. The replication origin of
JCVDNA wasassignedtotheregionof
nucleo-tides 36 to 121 (Fig. 2), which shows extensive
homology with the replication origin of both
SV40 (30) and BKV DNA(3). Thestructuresof replication origins are compared among the
threepolyoma virusesin Fig. 3.
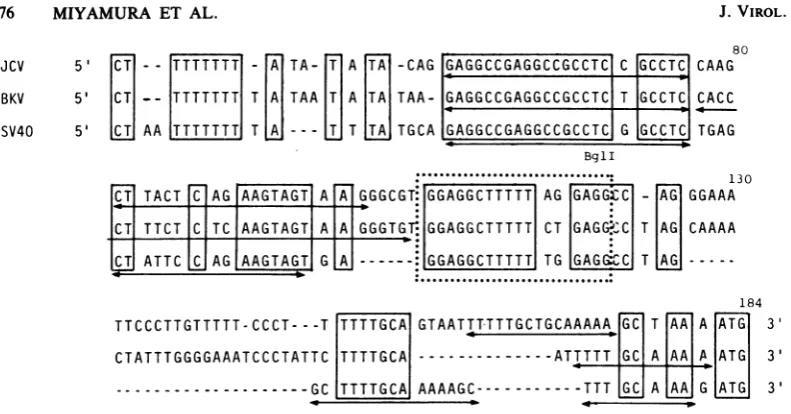
The nucleotide sequence of the replication
origincanbe divided into three parts with differ-ent structuralfeatures, ATstretch (nucleotides from 36to50; boxedinFig. 2),twopairsof the
inverted repeat sequences (nucleotides 54 to
102), and a palindromic sequence with mirror image form (nucleotides 105 to 121; boxed by
dotted line in Fig. 2 and 3). Goldberg-Hogness
box structure (24)was detectedattherightend
of the AT stretch (nucleotides 43to50in Fig. 2
and 3). The regionbetween ATstretchand the
palindrome was rich in GC, with intrastrand
symmetrywhich could formtwopairsof double
hairpin loops, asindicatedbythearrowsinFig.
3.
The coding region for small T antigen was
foundtoinitiate from ATGatnucleotides182to
184andterminateatTAAatnucleotides 698to
700 (Fig. 2). This ATG was the first initiation codonin thelongest openreadingframe
down-streamfromthereplicationorigin. Thetopology
oftheinitiation codonissimilartothatof BKV and SV40 (4, 34). Consequently, JC small-T-J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.489.53.444.68.320.2]5' ATGAGCTCAT TACTCGAGTA
CTTACTCAGA GAATGAGTCT
80 GCTTGGCTGG CAGCCATCCC TTCC TTTTT TTTATATATA CAGGAGGCCG AGGCCGCCTC CGCCTCCAAG CGAACCGACC GTCGGTAGGG AAGGGAAAAA AAATATATAT GTCCTCCGGC TCCGGCGGAG GCGGAGGTTC
HindIII
AGTAGTAAGG
GCG$g.GAGGC
TTTTTAGGAG G3CAGGGAAA TTCCCTTGTT TTTCCCTTTT TTGCAGTAATTCATCATTCC CGCACCTCCG AAAAATCCTC CGGTCCCTTT AAGGGAACAA AAAGGGAAAA AACGTCATTA
* ...** @
-240
TTTTTGCTGC AAAAAGCTAA
PEACAAA
GTGCTGAATA GGGAGGAATC CATGGAGCTT ATGGATTTAT TAGGCCTTGA AAAAACGACG TTTTTCGATT TTACCTGTTT CACGACTTAT CCCTCCTTAG GTACCTCGAA TACCTAAATA ATCCGGAACT 320 TAGGTCTGCA TGGGGGAACA TTCCTGTCAT GAGAAAAGCT TATCTGAAAA AATGCAAAGA ACTCCACCCT GATAAAGGTG ATCCAGACGT ACCCCCTTGT AAGGACAGTA CTCTTTTCGA ATAGACTTTT TTACGTTTCT TGAGGTGGGA CTATTTCCACHindIII
400 GGGACGAAGA CAAGATGAAG AGAATGAATT TTTTATATAA AAAAATGGAA CAAGGTGTAA AAGTTGCTCA TCAGCCTGAT CCCTGCTTCT GTTCTACTTC TCTTACTTAA AAAATATATT TTTTTACCTT GTTCCACATT TTCAACGAGT AGTCGGACTA
1 480
TTTGGTACAT GGAATAGTTC AGAGGTTGGT TGTGATTTTC CTCCTAATTC TGATACCCTT TATTGCAAGG AATGGCCTAA
AAACCATGTA CCTTATCAAG TCTCCAACCA ACACTAAAAG GAGGATTAAG ACTATGGGAA ATAACGTTCC TTACCGGATT
560 CTGTGCCACT AATCCTTCAG TGCATTGCCC CTGTTTAATG TGCATGCTAA AATTAAGGCA TAGAAACAGA AAATTTTTAA GACACGGTGA TTAGGAAGTC ACGTAACGGG GACAAATTAC ACGTACGATT TTAATTCCGT ATCTTTGTCT TTTAAAAATT 640 GAAGCAGCCC ACTTGTGTGG ATAGATTGCT ATTGCTTTGA TTGCTTCAGA CAATGGTTTG GGTGTGACTT AACCCAAGAA
CTTCGTCGGG TGAACACACC TATCTAACGA TAACGAAACT AACGAAGTCT GTTACCAAAC CCACACTGAA TTGGGTTCTT 720 GCTCTTCATT GCTGGGAGAA AGTTCTTGGA GACACCCCCT ACAGGGATCT
AAAGCTTE2
GGTAAACCAC TATTTTTCTTCGAGAAGTAA CGACCCTCTT TCAAGAACCT CTGTGGGGGA TGTCCCTAGA TTTCGAAATT CCATTTGGTG ATAAAAAGAA HindIII
1 800
TTGCAGGGCA TTTTGTTTTT TACAATAAAC TTAATTGTTT TTTTTTAGGT GCCAACCTAT GGAACAGATG AATGGGAATC
AACGTCCCGT AAAACAAAAA ATGTTATTTG AATTAACAAA AAAAAATCCA CGGTTGGATA CCTTGTCTAC TTACCCTTAG 880
CTGGTGGAAT ACATTTAATG AGAAGTGGGA TGAAGACCTG TTTTGCCATG AAGAAATGTT TGCCAGTGAT GATGAAAACA GACCACCTTA TGTAAATTAC TCTTCACCCT ACTTCTGGAC AAAACGGTAC TTCTTTACAA ACGGTCACTA CTACTTTTGT
CAGGATCCCA ACACTCTACC CCACCTAAAA AGAAAAAAAA GGTAGAAGAC CCTAAAGACT TTCCT
GTCCTAGGGT TGTGAGATGG GGTGGATTTT TCTTTTTTTT CCATCTTCTG GGATTTCTGA AAGGA 5' BamHI
FIG. 2. Nucleotidesequenceof JCVDNA:theregioncontainingreplicationoriginandsmall-T-antigengene.
Numbering isgiven fromneartheHindIII-A/Cjunction(nucleotide 1)to neartheBamHI site(nucleotide 945).We
identified the orientation ofthestrands fromthesimilaritytoBKV andSV40DNAs. The upperstrand,readfrom lefttoright, is believedtohavethesamepolarityasearlymRNA. The AT stretch within thereplication originis
indicatedin thesolid line box(nucleotides36to50). The dotted line box indicates thepalindrome (nucleotides
105to121). ATGatnucleotides182to 184 isconsidered tobe theinitiation codon forTantigen mRNA, and TAA at nucleotides 698 to 700 is considered to be the termination codon. The arrows refer to the splice
junctions forlarge-T-antigenmRNA. Thearrowbetweennucleotides424and 425isfor the donorsite,and the
other, between nucleotides768and769,is forthe acceptorsite(see Fig.6).
on November 10, 2019 by guest
http://jvi.asm.org/
76 MIYAMURA ET AL.
CT
-- TTTTTTT - A TA-T
A TA-CAG
GAGGCCGAGGCCGCCTCCT -- TTTTTTT T A TAA
T
A TA TAA- GAGGCCGAGGCCGCCTCCT
AATTTTTTTT
T A ---- TA TGCAGAGGCCGAGGCCGCCTC
80
C4GCCTC CAAG T GCCTC CACC
G
GCCTC
TGAG
BglI
CTI TACT m
AGIAAGTAGT|
A:...
S@*:
_1307AGGGCGT:
GGAGGCTTTTT AG GAGGfC - AG GGAAAA|GGGTGT'
GGAGGCTTTTT
CTGAGG;ZC
T AG CAAAAA-!GGAGGCTTTTT
TGGAGGCC
TAG-* d .... . ...s@*@ *@ @
184 TTCCCTTGTTTTT-CCCT - T TTTTGCA GTAATTTTTTGCTGCAAAAA GC T AA A ATG 3'
CTATTTGGGGAAATCCCTATTC TTTTGCA --- --ATTTTT GC A AA A ATG 3'
---GC TTTTGCA AAAAGC-TTT GC A AA G ATGL 3'
FIG. 3. Comparison of the sequences around thereplicationoriginsofJCV, BKV,andSV40DNAs. Boxes indicate the sequence homology among the three DNAs in this region. Nucleotides have been aligned to
maximize homology. Arrows indicate the areas with intrastrand symmetry which could formhairpin loops.The 17-nucleotide palindromic structures areboxedbydotted lines. TheBglIrestriction site(GCCN5GGC) isfor SV40 DNA. Numbering for JCV DNA is fromFig. 2.InBKVDNA, the first C and the lastGcorrespondto
nucleotides 140 and 5151, respectively, from Seifetal. (24). InSV40 DNA, the first C and the last G are at
nucleotides 5193 and 5079,respectively, from Reddyetal.(21).
antigengene cancode for 172 amino acids, like those of BKV and SV40encoding 172 and 174 aminoacids,respectively. Furthermore, the de-ducedaminoacidsequencesof smallTantigens showed extensive homology among the three polyoma viruses (Fig. 4).
Fromthehomologywith thesignalsequences
proposed forsplicing oflarge-T-antigen mRNA in SV40, BKV, and mousepolyoma virus (23),
we identified the sequence 5'TCAGAG/ GTTGGT3' atnucleotides 419to430as adonor
site and the sequence 5'TTTTTTTAG/
GTGCCA3' at nucleotides 760 to 774 as an
acceptor site oflarge-T-antigen mRNA of JCV
(Fig.5). In the nucleotidesequencedownstream from theacceptorsiteofJCV,onereading frame
was left open and the other two were closed.
The amino acid sequence predicted from the
openreading framedownstreamfromthe accep-torsitewassimilartothosefromBKVandSV40
T antigen genes in the corresponding regions (Fig. 6).
DISCUSSION
The complete nucleotide sequences and the
genomic organization for BKV, SV40, and
mouse polyoma virus have been established,
andseveralrecognitionsequencesforregulatory
functions have been mapped on the nucleotide
sequences(5,21, 24,28, 37). From the
homolo-gydeterminedbyDNA-DNAhybridization, the
cleavage map of JCV DNA has been aligned, with reference to EcoRI site, to those ofBKV
andSV40DNAs (11), but little is known about the genetic map and genomic organization of JCV. In the present study, we attempted to
locate the origin of DNA replication and the proximalpartof theearlyregionof JCVgenome
by comparingJCV sequences tothose of BKV and SV40 to correlate the physical map to the genomicorganization. Apparently,thesimilarity of thesequencesforreplication originandof the aminoacid sequencesforputative small T
anti-gen between JCV and other primate polyoma virusesproves thatJCV hasearlygenome
orga-nization similartothatof BKV andSV40onthe physicalmap of DNA with the conservedsingle EcoRIsite in each genome as areferencepoint. Since the structural features of the region around the HindIII-A/Cjunction of JCV DNA (at nucleotides 36 to121)were strikinglysimilar
to those ofthe replication origins ofBKV and
SV40 (3, 30), this area was regarded as the
replicationorigin of JCVDNA(Fig. 3). Thearea
iscomposed ofalongATstretch, twoinverted
repeatsequences,andapalindrome. At theright
end ofthe AT stretch, the three primate
poly-omaviruses had sequencessimilartothe Gold-berg-Hogness boxsequenceof 5'TATAAATA3' (24). By analogy to SV40 (1), sequence
5'TATATATA3'of JCV(nucleotides43to50) is probably related to the early promoter region.
TheAT-rich sequencewasfollowedbythe
GC-rich region which could form double hairpin loops (Fig. 3). Although it is unclear whether
this structure is actually formed in vivo, it is
JCv 51
BKV 5'
SV40 5'
CTITTCT
CITC
AAGTAGT Al CT ATTC C AG AAGTAGT GJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.489.50.446.54.261.2]JCV Met Asp Lys Val Leu Asn Arg Glu Glu Ser Met Glu Leu Met Asp Leu Leu Gly Leu Asp Arg Ser Ala Trp Gly
BKV Met Asp Lys Val Leu Asn Arg Glu Glu Ser Met Glu Leu Met Asp Leu Leu Gly Leu Glu Arg Ala Ala Trp Gly
SV40 Met Asp Lys Val Leu Asn Arg Glu Glu Ser Leu Gln Leu Met Asp Leu Leu Gly Leu Glu Arg Ser Ala Trp Gly
Asn Ile Pro Val Met Arg Lys Ala
Asn Leu Pro Leu Met Arg Lys Ala
Asn Ile Pro Leu Met Arg Lys Ala
Tyr Leu Lys Lys Cys Lys Glu Leu His Pro Asp Lys Gly Gly Asp Glu Asp Tyr Leu Arg Lys Cys Lys Glu Phe His Pro Asp Lys Gly Gly Asp Glu Asp
Tyr Leu Lys[ Cys Lys Glu Phe His Pro Asp Lys Gly Gly Asp Glu Glu Lys Met Lys Arg Met Asn Phe Leu Tyr Lys Lys Met Glu Gln Gly Val Lys Val Ala His Gln Pro Asp Phe Gly Lys Met Lys Arg Met Asn Thr Leu Tyr Lys Lys Met Glu Gln Asp Val Lys Val Ala His Gln Pro Asp Phe Gly Lys Met Lys Lys Met Asn Thr Leu Tyr Lys Lys Met Glu Asp Gly Val Lys TyrAla His Gln Pro Asp Phe Gly Thr - Trp Asn Ser Ser Glu Val Gly Cys - Asp - Phe Pro Pro Asn Ser Asp Thr Leu Tyr Cys Lys|Glu
Thr - Trp Ser Ser Ser Glu Val - Cys Ala Asp - Phe Pro Leu Cys Pro Asp Thr Leu Tyr Cys Lys Glu
Gly Phe Trp Asp Ala Thr Glu Val - Phe Ala Ser Ser Leu Asn Pro Gly Val AsAla MetjTyr Cys LysjGln
Trp Pro Asn Cys Ala Thr Asn ProS Val His sPro Cys Leu Met Cys Met Leu Lys Leu Arg His Arg Asn
Trp Pro Ile Cys Ser Lys Lys Pro Ser Val His Cys Pro Cys Met Leu Cys Gln Leu Arg Leu Arg His Leu Asn
Trp Pro Glu C Ala Lys Lys Met S Ala Asn C Ile&s Leu Leu C Leu L Arg Met Lys His Glu Asn
Arg Lys Phe Leu Arg Ser Ser Pro Leu Val Trp Ile Asp Cys Tyr Cys Phe Asp Cys Phe Arg Gln Trp Phe Gly Arg Lys Phe Leu Arg Lys Glu Pro Leu Val Trp Ile Asp Cys Tyr Cys Ile Asp Cys Phe Thr Gln Trp Phe Gly
Arg Lys Leu Tyr A Lys Asp Pro Leu Val Trp Val Asp Cys Tyr Cys Phe Asp Cys Phe Arg Met|Trp Phe Gly
Cys Asp Leu Thr Gln Glu Ala Leu His Cys Trp Glu Lys Val Leu GlAsp Thr Pro Tyr Arg Asp Leu Lys Leu Leu Asp Leu Thr Glu Glu Thr ILeul Gin Trp ITrpI Val Gin Ile Ile Gly Glu Thr Pro Phe Arg Asp Leu Lys Leu Leu Asp Leu Cys Glu Gly ThrL Leu Leu Trp Cys Asp Ile Ile Gln Thr Thr Tyr Arg Asp Leu Lys Leu
FIG. 4. Amino acidsequencesofJCV, BKV,andSV40small Tantigens.Thesequencewasdeducedfrom
the nucleotide sequence.Thesequencesfor BKV andSV40arefrompublisheddata(4, 34, 36). Gapswereleft
wherenecessarytomaximizehomology.The boxes indicate thehomologousamino acidsamongJCV, BKV,and
SV40.Thearrowbetweenglutamineand valine residues divides the amino terminalregion (commontolargeT
antigens)from thecarboxylterminalregionof theseproteins.
possiblethat thehigherstructuresof DNAnear
thereplicationorigin affecttheexpressionof the viralgenome. In the firsthairpin loop indicated
by an arrow in Fig. 3, 22 out of23 base pairs
wereidentical toeach otheramongJCV,BKV, andSV40. Thecomposition ofthe second
hair-pin structure, on the otherhand, was less
ho-mologous, but exactly the same sequence of
5'AAGTAGTAAGGG3' was found. Another
highly homologous sequence among the three polyomavirus DNAswasfound in 17-base pair palindromes. Of17nucleotides, 15 were identi-cal. Althoughthegeneralfeatures ofreplication originweresharedby mouse polyomavirus (6, 29), the strong homology of nucleotide
se-quences was detected only among the three
primate polyomaviruses.
There was a stretchof60nucleotidesbetween
the palindrome and the initiation codon for T
antigens (Fig. 3). Although thehomology in this noncoding regionamongthe threeprimate
poly-omaviruseswasweaker than thatinthe replica-tion origin, they had repeats of5'TnGC3' and
5'GCAn3',which could formhairpin loops (indi-cated by arrows in Fig. 3), in this region. The
structure ofthe corresponding region inmouse
polyomavirusDNA(27) is distinct from those of the primate polyoma viruses.
It seems reasonable to consider the ATG
codon at nucleotides 182to 184 tobean initia-tion signal for translation of T antigen messen-gers for the following reasons: (i) this was the firstinitiationcodon in the only longopen read-ing frame in this region; (ii) its topology was
almost the same as thatof its BKV and SV40
counterparts; (iii) the deduced amino acid
se-quencefor JCV smallTantigen resembled those
of BKV and SV40 (Fig. 4).
Thus, the small-T-antigen gene of JCV can
code for 172 amino acids (Fig. 4). In the
N-terminal regions, where the small T antigen sharesits amino acids with the largeTantigen,
JCV 5' TCAGA TTGGT...TTTTTTTA TGCCA 3' BKV 5' TCAGA TTTGT...TTTTTATAGGTGCCA 3'
SV40 5' ACTGAG TATTT...GTATTTTAG TTCCA 3'
Py 5' TTCCA TAAGA...TTCTTACAG GCTCT 3'
FIG. 5. Splicing signal sequences for large-T-anti-gen mRNA ofpolyoma viruses. The intervening se-quences of JCV, BKV, SV40, and mouse polyoma (Py) virus are 344, 344, 346, and 386 nucleotides long, respectively. The sequences for BKV, SV40, and
mousepolyoma virus are frompublished data (5, 21, 24, 28).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.489.47.443.61.335.2]78 MIYAMURA ET AL.
JCV Val Pro BKV Val Pro
SV40 Il1e Pro
Thr PheAsnGliu Lys
Ser Phe AsnGlu Lys'
A1lA Php Ac,n rlij-l_
I--I I'-u I J'U"AJ-IU U
Asp Asp Len Cy: His Gilu 'la er spAsp Asn rGly AspGiu Asp Leu Cys His Glu Asp Met
'Ala
Ser AspGiuuG1u Ala ThrAla AspAsn Leu Cys Ser Glu Glu Met Pro Ser AspAspGlu Ala Ala Asp
Ser Gln His SerThr Pro Pro Lys Lys Lys Lys LysVal Glu Asp Pro Lys Asp Pro...
Ser Gln His SerThr Pro Pro Lys Lys Lys Arg Lys Val Glu Asp Pro Lys Asp Pro ... Ser GlnHis Ser Thr Pro Pro Lys Lys Lys Arg Lys Val GluAsp Pro Lys Asp Pro ...
FIG. 6. Amino acid sequences encoded by the areas just downstream from the splice junction for large-T-antigen mRNA in JCV, BKV, and SV40. Homologous amino acids are shown by boxes. Aromatic amino acids (e.g., tyrosine, tryptophan, and phenylalanine) are shown by shading. Sequences forBKVandSV40 are from published data (5, 21, 24).
JCV and BKV shared 72 of 81 amino acids
(89%). The homology between JCV and SV40
was83%(67of81), and that between BKV and
SV40wasalso83%. Amino acids shared by the threeviruses in theproximal half of the smallT
antigen were as many as 63 (78%). The restof thesmall-T-antigen coding region seemedto be diverged somewhat more from BKV and SV40 than the former region shared by the large T
antigen. In this region, 63 of 91 amino acids (69%)werecommon toJCV and BKV, 49(54%)
to JCV and SV40, and 54 (59%) to BKV and SV40. The three viruses shared only 43 of 91 amino acids(47%) but hada commonstretchof five amino acids, Arg-Asp-Leu-Lys-Leu, fol-lowed by stop codon TAA. These data are
consistent with the earlier reports showing the relatedness ofT antigens of the three primate polyoma viruses by immunological
cross-reac-tivity (32, 35) or 35S methionine-labeled tryptic peptide patterns (26).
The signal sequences for splicing of mRNA are conserved among eucaryotic species, and
the consensus sequences have been proposed
(15). The latterhalf of the small-T-antigengene
is splicedout inlarge-T-antigen mRNAs of
pa-povaviruses. Thesplicing signals for JCV large-T-antigenmRNA agreewellwith those of
poly-oma viruses (23) (Fig. 5) and the consensus
sequences (15). The amino acid sequence
en-coded downstreamfromthesplicing site
resem-bled those of BKV and SV40 (Fig. 6). Like
BKV, SV40, and mouse polyoma virus (24),
JCV had a stretch of aromatic amino acids
sequence in theregion justdownstreamfromthe
splicing signal.
It has not been determined yet whetherJCV
has a virus-coded middle Tantigen like that of
mouse polyoma virus. Downstream from the
splice junction (acceptor site), mouse polyoma virus DNA has two long open reading frames,
which are used for encoding large and middle T
antigens, respectively (28).Inthecorresponding region of JCV DNA,thetwoframesnotusedfor encoding largeTantigenwereshort(Fig. 2),and
the amino acid sequences deduced from these
twoframes (data not shown) hadno homology with that of mouse polyoma virus middle T
antigen (28). The topology ofcodingframes of JCV in this arearesembled those of SV40 and BKV. Weconclude,therefore,thatJCVcannot
code for the protein like mouse polyoma virus middleTantigen.
The present study showed that, in the region
containing the replication origin and small-T-antigengene, the structureof JCVDNA
resem-bles those of the other polyoma viruses. The homology in the nucleotide or amino acid
se-quence washigherbetween theprimate polyoma
viruses than between JCV andmouse polyoma virus (28) andwasthehighest between JCV and BKV. Despite the similarity between JCV and BKVinthisregion, thetwoviruses have
differ-ent pathogenicity in humans and different host
rangesfor cultured cells. Thesequences
respon-sible for the biological difference between the
twoviruses remainto be studied.
A host range mutant ofJCV, JC-HEK, can
grow well in HEK cell cultures (14), whereas wild-type JCV (Mad-1)cangrowefficiently only in primary human fetal glia cell cultures (17). Interestingly,mutantJC-HEK hasnowthesame
hostrange asBKV. Itispossible that
compari-son of the nucleotide sequence between JCV
andJC-HEK, and between JCV andBKV, will
reveal the structure that determines the host
rangeofthese viruses. Astudy ofthe structure
of JC-HEKDNA isnow in progress.
ACKNOWLEDGMENTS
This workwassupportedin partbygrants-in-aidforcancer researchfrom theMinistry ofEducation, Science,and Cul-ture,Japan.
We thank Peter M. Howley, National Cancer Institute, Bethesda, Md., for his generousgiftofcloned JCV DNA.
LITERATURECITED
1. Benoist, C., and P. Chambon. 1981. In vivo sequence requirementsof theSV40earlypromoterregion.Nature (London) 290:304-310.
J.VIROL.
I'
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.489.94.391.72.163.2]2. Brown, P., T. Tsai, and D. C.Gajdusek.1975. Seroepide-miology of human papovaviruses. Am. J. Epidemiol. 102:331-340.
3. Dhar, R., C.-J. Lai, and G. Khoury. 1978. Nucleotide sequence of the DNAreplication origin for human papo-vavirus BKV: sequence and structural homology with SV40. Cell 13:345-358.
4. Dhar, R., I.Self,and G. Khoury. 1979. Nucleotide se-quenceof the BK virus DNA segment encoding smallt antigen. Proc. Natl. Acad. Sci. U.S.A. 76:565-569. 5. Fiers, W., R. Contreras, G. Haegeman, R. Rogiers, A. Van
deVoorde, H. Van Heuverswyn, J. VanHerreweghe, G. Volckaert,and M. Ysebaert. 1978. Complete nucleotide sequenceofSV40 DNA. Nature (London) 273:113-120. 6. Friedmann, T., P. LaPorta, and A.Esty.1978. Nucleotide
sequencestudies of polyoma DNA. TheHpaII3/5 junc-tion to theHpaII4/HaeIII18junction,encodingtheorigin of DNAreplicationand the 5' end of theearly region. J. Biol.Chem. 253:6561-6567.
7. Frisque, R. J., D. B. Riflin, and D. L. Walker. 1980. Transformation of primary hamster brain cells with JC virus andits DNA. J. Virol. 35:265-269.
8. Hay, R. T., and M. L. DePamphilis. 1982. Initiation of SV40 DNAreplicationin vivo:location andstructureof 5' ends of DNAsynthesizedin the oriregion. Cell 28:767-779.
9. Howley, P. M. 1980.Molecularbiology of SV40and the humanpolyomaviruses BKand JC, p. 489-550. In G. Klein(ed.), Viral oncology. Raven Press, New York. 10. Howley, P. M., F.Rentier-Delrue, C. A.Heilman,M.-F.
Law, K. Chowdhury, M. A. Israel, and K. K. Takemoto. 1980. Cloned human polyomavirus JC DNA can trans-formhumanamnioncells. J. Virol. 36:878-882. 11. Law, M.-F., J. D. Martin, K. K. Takemoto, and P. M.
Howley. 1979. The colinear alignment of genomes of papovaviruses, JC, BK, and SV40. Virology 96:576-587. 12. Martin, J. D., R. J. Frisque, B. L. Padgett, and D.L. Walker.1979. Restriction endonucleasecleavage map of the DNA of JC virus. J. Virol.29:846-855.
13. Maxam, A. M., and W. Gilbert. 1980. Sequencing end-labeled DNA with base-specific chemical cleavages. MethodsEnzymol.65:499-560.
14. Miyamura,T., K.Yoshilke,and K. K. Takemoto. 1980. Characterization of JCpapovavirus adaptedtogrowthin humanembryonickidneycells. J. Virol. 35:498-504. 15. Mount, S. M. 1982. A catalogue ofsplice junction
se-quences.Nucleic Acids Res. 10:459-472.
16. Osborn, J. E., S. M. Robertson, B. L. Padgett, G. M. ZuRhein, D. L.Walker,andB.Weisblum.1974. Compari-sonof JC and BK humanpapovaviruseswithsimian virus 40:restriction endonucleasedigestionandgel electropho-resis of resultantfragments.J. Virol. 13:614-622. 17. Padgett, B. L., C. M. Rogers, and D.L.Walker.1977.JC
virus,ahumanpolyomavirus associated with progressive multifocal leukoencephalopathy: additional biological characteristics and antigenic relationships. Infect. Im-mun. 15:656-662.
18. Padgett, B.L., and D. L. Walker. 1973. Prevalence of antibodiesin human seraagainst JC virus, an isolate from acaseofprogressivemultifocalleukoencephalopathy.J. Infect.Dis. 127:467-470.
19. Padgett, B. L., and D. L. Walker. 1976. New human papovaviruses. Prog. Med. Virol. 22:1-35.
20. Padgett,B.L.,D. L.Walker,G.M.ZuRhein,andJ.N. Varakis.1977.Differentialneurooncogenicityof strainsof JC virus, a human polyoma virus, in newborn Syrian
hamsters. Cancer Res. 37:718-720.
21. Reddy, V. B., B.Thlimmappaya, R. Dhar, K. N. Subra-mnanan, B. S. Zain, J. Pan, P. K. Ghosh, M. L. Celma, and S. M. Weissman. 1978.The genome of simian virus 40. Science 200:494-502.
22.Rentier-Delrue, F., A. Lubnlecki, and P. M. Howley. 1981. Analysis of JC virus DNA purifieddirectly from human progressive multifocal leukoencephalopathy brains. J. Virol. 38:761-769.
23. Seif, I., G. Khoury, and R. Dhar. 1979. BKV splice sequences based on analysis of preferred donor and acceptorsites. Nucleic Acids Res. 6:3387-3398. 24. Self,I., G. Khoury, and R. Dhar. 1979. The genomeof
humanpapovavirus BKV. Cell 18:963-977.
25. Shortle, D. R., R. F.Margolskee,and D. Nathans.1979. Mutational analysis of the simian virus 40 replicon: pseu-dorevertants of mutants with a defective replication ori-gin. Proc. Nati. Acad. Sci. U.S.A. 76:6128-6131. 26. Shnmons, D. T., and M. A. Martin. 1978. Common
methionine-tryptic peptides near the amino-terminal end ofprimate papovavirustumorantigens.Proc. Natl.Acad. Sci.U.S.A. 75:1131-1135.
27. Soeda, E., J. R. Arrand, N. Smolar, and B. E. Griffin. 1979. Sequence from early region of polyoma virus DNA containing viral replication origin and encoding small, middle and (part of) large T antigens.Cell17:357-370. 28. Soeda, E., J. R. Arrand, N. Smolar, J. E. Walsh, and B. E.
Griffin. 1980.Coding potential andregulatory signalsof thepolyoma virus genome. Nature(London)283:445-453. 29. Soeda, E., G.Kimura,andK.Miura.1978.Similarityof
nucleotide sequences around theoriginof DNA replica-tion in mousepolyoma virus and simian virus 40. Proc. Natl. Acad. Sci. U.S.A. 75:162-166.
30. Subrama ,K.N., R. Dhar, and S.M.Weissman.1977. Nucleotide sequence of afragment of SV40 DNA that contains theoriginof DNAreplicationandspecifiesthe5' ends of "early" and "late" viral RNA. III. Construction ofthe totalsequenceofEcoRII-GfragmentofSV40 DNA. J. Biol.Chem. 252:355-367.
31. Takemoto, K. K., P. M. Howley, and T. Miyamura. 1979. JC humanpapovavirusreplicationin human amnion cells. J. Virol. 30:384-389.
32. Takemoto,K.K., and M.A.Martin.1976. Transformation of hamster kidney cells by BK papovavirus DNA. J. Virol.17:247-253.
33. Varakis, J., G. M. ZuRhein, B. L. Padgett, and D.L. Walker.1978. Induction ofperipheralneuroblastomas in Syrianhamstersafterinjectionas neonateswithJCvirus, ahumanpolyomavirus.Cancer Res. 38:1718-1722. 34. Volckaert, G., A. VandeVoorde, andW. Fkers. 1978.
Nucleotide sequenceof the simian virus 40 small-t gene. Proc. Natl. Acad. Sci. U.S.A.75:2160-2164.
35. Walker, D. L., B. L. Padgett, G.M. ZuRhein, A. E. Albert,andR.F.Marsh.1973. Humanpapovavirus (JC): inductionofbrain tumors in hamsters. Science 181:674-676.
36. Yang, R. C. A., and R. Wu. 1979. BK virus DNA sequencecodingfor theamino-terminus of theT-antigen. Virology92:340-352.
37. Yang, R. C. A., and R. Wu. 1979. BK virus DNA: complete nucleotide sequenceofa humanpapovavirus. Science206:456-462.
38. Yoshlke, K., T. Mlyamura, H. W. Chan, and K. K. Takemoto.1982. Twodefective DNAs of human polyoma-virusJC adapted to growth in humanembryonic kidney cells. J. Virol.42:395-401.
on November 10, 2019 by guest
http://jvi.asm.org/