0022-538X/81/060805-10$02.00/0
Polyoma Virus Minichromosomes:
aSoluble In Vitro
Replication Systemt
BRIAN B. GOURLIE, MARC R. KRAUSS, ALICIA J. BUCKLER-WHITE, ROBERT M. BENBOW,
ANDVINCENT PIGIET*
McCollum-PrattInstitute and theDepartment of Biology, Johns Hopkins University, Baltimore, Maryland 21218
Received21October 1980/Accepted17February 1981
Polyoma virus minichromosomes were isolated from infected 3T6 cells by
hypotonic extraction of isolated nuclei. The kinetics of in vitroDNA synthesis in
thenuclearextractwassimilartothat observed with intact nuclei.The majority
of theproducts ofin vitro DNAsynthesissedimented with replicative interme-diate (RI) minichromosomes and migratedastwobands (RI-a andRI-b)on 1.4%
agarosegels.Thekineticsofdeoxynucleotide monophosphate incorporationinto
thesespecieswasconsistentwith the existence of several rate-limitingstepsin in
vitro replication by polyoma minichromosomes. Electron microscope analysis
showedthat the RI-a band consisted almost entirely of RI structures ranging
from 46to87% replicated, with one-half ofall structures 67 + 4% replicated.
TheRI-b materialwas morecomplex, consisting ofaandastructureswith tails
ranging from7 to 114%ofpolyomagenomelength and, less frequently, of linked
andmultiplylinked dimeric structures.
As a foundation for the eventual
reconstitu-tion of chromatin replication in vitro, we have
studied thereplicationof isolatedpolyoma
min-ichromosomes. These small, well-defined viral
genomeshave a chromatinstructure similar to
thatof the host (3, 7, 20) and provide anexcellent
model foreucaryotic replication (5, 22, 28, 29).
With theexceptionof the virus-coded Tantigen,
which is responsible for initiating each new
roundofDNAreplication(26), the hostprovides
allof themachinerynecessaryfor viral
replica-tion.
In this paper, wehave characterized
endoge-nous DNA synthesis by isolated minichromo-somesand have shown the accumulationof
rep-licative intermediates (RI-a and RI-b). In the
second paper (13), the electronmicroscope was
usedtoquantitateand characterize DNAspecies associatedwithmature (form I) and
replicating
(RI) minichromosomes. In the third paper (8),
we show that several enzyme activities
impli-cated in DNAreplicationcosediment with
pol-yomaminichromosomes.
MATERIALS AND METHODS
Virus and cells.Asingle-plaqueisolate of a large-plaquestrain ofpolyomaviruswaskindlyprovided by
M. Martin andpropagated atlowmultiplicity (10-3)
onprimarybabymousekidneycells.3T6mouse fibro-blasts were seeded at 2.5 x 106 cells per 150-mm
t Publication 1087 from the McCollum-Pratt Institute.
culture plate (Falcon Plastics) inDulbecco modified Eagle medium (GIBCO Laboratories) supplemented with 5% calfserum (GIBCO)and grown at 38'Cina 5% CO2 atmosphere. Cells were infectedon the 2nd
day after seeding (approximately80% confluency) at amultiplicity of infection of approximately15.
Reagents. HEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid), dithiothreitol, phenylmeth-ylsulfonyl fluoride, ethylene glycol-bis(B-aminoethyl
ether)-N,N-tetraacetic acid, and unlabeled deoxyri-bonucleoside 5'-triphosphates were obtained from Sigma Chemical Co. Pyruvate kinase was obtained
from Calbiochem. [3H]thymidine (55 Ci/mmol) and
[a-32P]dTTP(317Ci/mmol)wereobtained from ICN.
[a-32P]dGTPwasobtained fromAmershamCorp. The
restrictionenzymeBglIwasobtainedfrom Bethesda Research Laboratories.
Preparation of polyoma nuclearextracts(23). Twenty-sevenhours afterinfection, cell cultureswere incubatedin vivo either with 20,uCiof[3H]thymidine
(55 Ci/mmol) in 10 ml of Dulbecco modified Eagle
medium for 90 minorwith 100 LCiof[3H]thymidine
in 2ml for5min.Theplates (nowcontaining4x 107 cells andapproximately20 LgofpolyomaDNA)were
rinsed three times with 5 ml of ice-cold hypotonic bufferA(10mMHEPES, pH7.8,5mMKCI,0.5mM
MgCl2,0.5mMdithiothreitol,and1mM phenylmeth-ylsulfonyl fluoride) and drained. The cells were
re-moved with arubber scraper, transferred to an ice-cold Dounce homogenizer, and disrupted with 10
strokes oftight-fittingpestle.Nucleiwerepelletedat
3,000 xg inaSorvall typeSS-34rotorfor 5 min. The supernatantwasremoved,and the nuclearpelletwas suspended with hypotonic extraction buffer (0.5 ml perplate) and extracted at0°Cfor2 h. Nucleiwere
805
on November 10, 2019 by guest
http://jvi.asm.org/
removed bycentrifugation (6,800xg) for15min, and the supernatant solution of the nuclear extract was
used for further study.
In vitro DNAsynthesis (30).ViralDNAsynthesis wasassayed in a total volume of100,llin the presence of: 40 mMNaCl,5mMMgCl2,5mMethylene glycol-bis(B-aminoethyl ether-N,N'-tetra-acetic acid, 4 mM phosphoenol pyruvate,2 mMATP,3.0IUof pyruvate kinase per ml, and 100,uM each dATP, dCTP, and dGTP.Experiments using [3H]- and [32P]dTTPwere
performedat25ytCi/ml(18 and300Ci/mmol, respec-tively). Nucleiornuclearextract wasadded ina vol-umeof90,dl,and incubationswereperformedat30°C. Duplicateportions of10 or 20 ulwereremoved and precipitated with2ml of cold 10% trichloroacetic acid containing10mMsodiumpyrophosphate. Acid-insol-uble materials were collected on glass fiber filters (WhatmanGF/C) and washed four times with2mlof cold 5%trichloroacetic acid-5 mM sodium pyrophos-phateand oncewith cold95% ethanol. Radioactivity wasdetermined on the dried filters byliquid scintil-lationcounting in4mlof Beckman EP fluor.
Analysis of viral chromatin by sucrose gra-dient sedimentation. Nuclearextracts(0.5ml)were
sedimentedthrough linearsucrosegradients(10ml,5 to20%,wt/vol) prepared in buffer B (10mMHEPES, pH 7.8,5 mMKCI, and0.5mMMgCl2) for90minat
255,000xg inaBeckmanSW41rotor.Fractionswere
collected from thebottom, andportions of each frac-tionwereanalyzed for acid-insoluble radioactivityas
described above. Relative sedimentation velocity of minichromosomes was determined relative to the sed-imentationof form IpolyomaDNA(20S)on aparallel but separatesucrosegradient inhigh-salt buffer C (0.5
MNaCl, 10mMEDTA, and 10mMsodium acetate, pH 6.0).
Analysis of viral DNA by sedimentation on
neutral and alkaline sucrose gradients.After in vitroincubation, viralDNA wasprepared from viral chromatin by the method of Hirt (11).Samples (100 y1)weresedimentedthrough neutral sucrose gradients (3.7 ml, 5 to 20%, wt/vol) in buffer C above a 70%
sucrosecushion (100,l)inbuffer Coralkalinesucrose
gradients which also contained 0.05% sodium
sarcosi-nateand0.3MNaOH. Gradientswerecentrifuged in
a Beckman SW60 rotor at350,000 x g for 110 min (neutral) or 200 min (alkaline). Fractions were col-lected from thebottom, and acid-precipitable radio-activitywasdeterminedas above.
Agarose gel electrophoresis of viral DNA (27). Electrophoresis was performed by using 1.4% (wt/vol) agaroseslabgels (18cmby 19 cm by 3 mm) in 40 mM Tris-hydrochloride,5 mMsodiumacetate, and 1 mM EDTA, pH 7.6. Samples of nuclear extract were
treated with0.6% sodium dodecyl sulfate, 10 mM Tris-hydrochloride, and 10 mM EDTA (pH 7.6) for 20 min
at30°C andelectrophoresed for12to 18 h at 2.7 V/
cmalongwithpolyoma form I and II DNA markers. The gelwasstained with ethidiumbromide (0.5,ug/ ml, 30min), and the fluorescentbands were photo-graphed under UV illumination (254 nm) through a Wratten no. 25 (red) filteronPolaroid type 55 P/N film. Autoradiography wasperformed at 4°C on wet gels, using Kodak X-Omat R film. BglI restriction enzyme digestionofdeproteinized DNA synthesized
bythenuclearextract invitrowasperformedas de-scribed by thesupplier.
Electron microscope analysis of viral DNA formsseparated byagarosegelelectrophoresis.
Sections of 1.4% agarose gels corresponding to the form I, RI-a and RI-b bandswereexcised and dissolved in 5 volumes of saturated potassium iodide, 10 mM
sodiumphosphate (pH 7.0), and 1 mMsodium thio-sulfate.Sampleswerespreadfor electronmicroscopy without furtherprocessing, usingthe aqueousmethod of Davis et al. (la). Every DNA molecule seen was photographedat x20,000magnification withaJEOL JEM 100Belectronmicroscope,projectedwitha
Ni-kon Shadowgraph comparator (total magnification, approximately x200,000), and traced with a Keufel and Esser map measurer. Photographswere printed
onIlfobrom 5.1M resin-coated paper without further
contrastenhancement.
RESULTS
Kinetics of in vitroDNA
synthesis by
the nuclear extract.Polyoma
minichromosomeswere extracted fromvirus-infected 3T6 cells at a time corresponding to maximal viral DNA
synthesis (27 h
postinfection
in our system),using a hypotonic extraction procedure
origi-nally described forisolation of simian virus40
(SV40)minichromosomes (22).Thenuclear
ex-tract, preparedfrom cells labeled foreither5 or
90minwith
[3H]thymidine
beforeharvest,con-tained 10 to20%of thetotallabeledviral DNA
as determinedby theHirt procedure (11). This protocol allowed
preferential
labeling of eitherthe RI (5-min pulse) or mature (90-min pulse) formIspecies.The efficiencyofextraction was
comparable
to results obtained with SV40-in-fected cells (5, 22), but was less efficient than methods usingTritonandhigh salt (9).Theability ofnuclearextracts toincorporate
[32P]dTMP was determined by using reaction
conditionsoptimizedforpolyomaDNA
synthe-sis in isolated nuclei (30).
[32P]dTMP
incorpo-rationinto DNA was rapid during the first 5 min and reached a plateau after 10 min (Fig. 1).
Maximalinvitro DNAsynthesisby the nuclear extract (both initial rate andplateau) was
ob-tainedafter extraction ofnuclei for2 h at0°C.
EndogenousDNA synthesis in the nuclear
ex-tract wasstable for at least 8 h and decreased
only 20% after storage overnight at 0°C. The
rateof DNAsynthesis in the nuclear extract was
compared with the rate of synthesis by nuclei
isolated by an isotonic procedure (20). Both
reactions reached a plateau after 10 min. The
initialrateof dTMP incorporation was 0.10 and
1.14pmol/min per plate for the nuclear extract
and the intact nuclear preparations,
respec-tively. Since the nuclear extract contained 10 to 20% of the total polyoma DNA in the cell (based on the efficiency ofextracting DNA labeled in
on November 10, 2019 by guest
http://jvi.asm.org/
E
O 10
z 0
0.
Os
0 5 10 20 30 60
INCUBATION TIME (min)
FIG. 1. Kinetics ofDNA synthesis bythenuclear extract. Nuclearextract(correspondingto3.5x JOr
cells) in 500 ,Il was assayed for endogenous DNA synthesis in thepresenceof [a-32P]dTTP (158x 103 cpm/pmol) asdescribed in Materials andMethods.
At the indicatedtimes, 10-,ld portionswereremoved,
and the amount ofacid-precipitable [a-32P]dTMP wasdeternined.
vivo), the initialrateof DNA synthesis in both
preparationswasroughly comparable, assuming
equal extraction ofarepresentative population
of functionalreplicating complexes (see
Discus-sion). The extent ofdenovo synthesis in vitro
after 10 min canbe calculated fromthe initial
rate (see above) and the amount ofreplicative
intermediates(0.6pmol/plate)asapproximately
120 pmol of nucleotide per mol ofreplicative
intermediate. This calculation isanaverageand
does not discriminate between forks that may
varyintheir relativeefficiency. After hypotonic
extraction ofminichromosomes,nuclei still
con-tained 60 to 80% of the viral DNA labeled in
vivo, but less than 5% of the DNA synthetic
capacity of either the nuclear extract or the
intactnuclei. Thecytoplasmicsupernatant
con-tained the remainderof the totalpolyomaDNA
in thecell (10to20%).
Characterization of the reaction
prod-uctsby sedimentation analysis. The
32P-la-beled productsof in vitro DNA synthesis were
characterized either as viral chromatin or as
deproteinized DNA. The 32P-labeled products
synthesized bythe nuclear extractduringa
30-min incubation sedimented between 50S and
90S, withamajor peakat84Sandashoulderat
56S relative to polyoma form I DNA (Fig. 2).
For comparison, mature polyoma
minichromo-somes (labeledwith[3H]thymidinefor 90minin
vivo) sedimentedatapproximately56S,whereas
minichromosomes labeled in vivo foronly5min
(predominantly RI minichromosomes)
sedi-mented atapproximately 84S (Fig. 3). This
sed-imentation behavior was comparable to results obtained by others using different extraction
conditions for polyoma virus (29) or for SV40
minichromosomes (5, 23).
The 32P-labeled products, deproteinized by
treatmentwith0.6%sodiumdodecyl sulfateand
3
E
4
in
22
z
0
to
0 5 10 15 20
[image:3.495.157.446.76.307.2]FRACTION
FIG. 2. Characterization of the chromatin product of DNA synthesis in the nuclear extract. After incu-bation ofthe nuclear extract for 30 min with
[a-32P]dTTP (0)asdescribed in Fig. 1, the sample was
sedimented withoutfurther treatment through a neu-tral sucrosegradient as described in Materials and Methods. Radioactivity was determined after acid precipitation of 10 ul of each fraction. The viral DNA
was labeled in vivo with 11H]thymidine (0) for 30 minimmediately before harvest. The arrow indicates thepeakofform Ipolyoma DNA (20S) sedimented on
a separate 5 to 20% sucrose gradient in high-salt buffer C.
E
0
4-0
[image:3.495.259.453.156.304.2]FRACTION
FIG. 3. Sedimentation of mature (form I) and RI viral chromosomes. Infected cells (3.5 x 107 cells/ plate) were labeled either with 20,uCi of 19H]thymi-dine(55 Ci/mmol) in 10 ml for90min (0)or with 100
,iCi of/5H]thymidine in 2 mlfor 5mmn (0) before
harvest. Portions (500,ul)of thenuclear extracts were sedimented asdescribed in Materials and Methods. Radioactivity was determined after acid precipita-tion of10O,ufor thesample labeled for 90minand the entire 0.5-ml fraction for the sample labeled for5mmi.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.495.257.453.439.565.2]808 GOURLIE ET AL.
1 MNaCl (11), were characterizedby
sedimen-tationonbothneutral and alkaline sucrose
gra-dients. The in vitro DNA productssedimented
in aneutralsucrosegradient slightlyfasterthan polyoma form I DNA (20S) (Fig. 4), which is
consistent with the sedimentation behavior of RI DNA (21). In contrast to the SV40 system
(23), the majority of the invitro-labeled DNA
remained associatedwith the parental genome
whenanalyzed eitheras intact chromatin (Fig.
2) or as deproteinized DNA (Fig. 4).
Sedimen-tation under alkaline conditions of the
depro-teinized, 32P-labeled product of a 30-min
incu-bationinvitrodemonstrated thatthedaughter strandsrangedinsizefrom 16S (full-length lin-earmolecules) to4S (Okazaki fragments) (Fig.
5). Significantamountsof mature form I DNA
(53S) were notobserved.
Characterization
of the reactionprod-ucts by agarose gel electrophoresis. The
results of agarose gel electrophoresis of
32P-la-beledmaterialsynthesizedin vitro are shown in
theautoradiograminFig.6.Themajorproducts
ofvitrosynthesis appearedas twobands
migrat-ingslower than relaxed circular polyoma DNA
(form II). We refer to these bands as RI-a and RI-b, based on their cosedimentation with RI minichromosomes labeled in vivo (Fig. 3). Su-percoiled Cairns-type replicative intermediates would be expected to migrate as a spectrum
from form I (earlyreplicativeintermediates) to
6
I
0
-4
z 03
0.
0 10 20 30 40 50 60
FRACTION
FIG. 4. Characterization of the deproteinized product ofDNAsynthesisbysedimentation inneutral sucrose.Afterincubation of the nuclearextractfor30
min with[a-32P]dTTP(0)asdescribed in Fig.1,the
sample was treated according to the procedure of
Hirt(11). The extracted DNA wasthen sedimented
on aneutralsucrosegradientasdescribed in
Mate-rials andMethods. Fractionswerecollectedfrom the
bottom, and acid-precipitableradioactivity was de-terminedasdescribedabove. Viral DNAwaslabeled in vivofor30min immediately before harvest with [3H]thymidine (0).
E
Iy
,ss
60
60
z
0.
Ni
40-0
20-0 (0 20 30 40
FRACTION
FIG. 5. Characterization of the deproteinized
productof DNAsynthesis bysedimentationin alka-linesucrose.Aportionofthesamesamplelabeled in vitro and in vivo as described in Fig. 4 was sedi-mentedonanalkalinesucrosegradientasdescribed inMaterials and Methods. Fractionswerecollected from the tube bottom andanalyzedfor32P in vitro
(0)
or3Hinvivo(0)
label.a position slowerthan form II
(late
replicative
intermediates)
(16,25).
Thesamespecies
in thesame relative amountswere present in vivo as
determined
bypulse-labeling
with [3H]thymi-dine. Therelativelabelings
ofbothRI-aandRI-b in vitro were also
comparable
when intactnucleiwereused
(Gourlie
andPigiet, manuscript
in
preparation).
Since variations in agarose
concentration,
electricfield, and temperature
differentially
af-fectthemigration
oflinear,
circular,
andsuper-coiled DNA ofthe same molecular
weight (4),
we
investigated
the effect of agaroseconcentra-tions from 0.8 to
1.4%
(wt/vol).
The RI-a and RI-bbands maintainedthesamerelative mobil-itiestothepolyoma
formIImarker,
but became broaderandmorediffuseatlowergelconcentra-tions, indicating thatthere may be further
het-erogeneity withinthebands.
Labeling of the RI-a band wasrapid and
es-sentially
complete after 5 min (Fig. 6). Incon-trast,labeling of the RI-b band increased from
about 20% of the totalincorporationat 5minto
about 40%of the total at60min.Thus,
substan-tiallabelingofRI-b continuedaftertotal
incor-poration had reached a plateau (10 min).
La-beled
form
I DNA accumulated toapproxi-mately3% of the total
incorporation by
60min,consistent withresultsof sucrosegradient
sedi-mentation (Fig. 5).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.495.256.447.65.277.2] [image:4.495.51.246.399.655.2]Incubation Time
(min)
5 10 30 60
-RI-b
-_
[image:5.495.58.240.69.304.2]-I
FIG. 6. Characterizationof the products of in vitro DNA synthesis by agarosegel electrophoresis. Nu-clear extract was incubated asdescribed in Fig. 1
with[a-32P]dTTP.Attheindicatedtimes,
50-,ullpor-tionswereremovedfromthe incubation,treated with 0.6% sodium dodecyl sulfate, 10 mM EDTA, and 10 mM Tris-hydrochloride (pH 7.6) for 1 h at 20°C, mixedwith 1/10volumeof 0.05%bromophenol blue, 25 mMEDTA, and 50% glycerol, andapplied to a 1.4%agarosegel. Electrophoresiswascarriedoutas
describedinMaterials andMethods. Autoradiogra-phywasperformedat4°C for14 h.Migration ofform
I and form II DNA is indicated.
Tocomparethedistributionof RI-aand RI-b
DNAs as a function of sedimentation of the
chromatinproducts,agarosegelelectrophoresis
was performed on the DNA from individual
fractions of thesucrosegradient showninFig.2.
Theautoradiogram of this gel (Fig. 7A) showed
the distribution of label incorporated into the
RI-a andRI-bbands.Directquantitation ofthe
radioactivity in the excised bands (Fig. 7B)
showed that RI-a sedimented as a single peak
with anaveragerate of 84S (fraction 9),
corre-sponding to the fraction which contained the
maximum in vitro label. The RI-b DNA, in
contrast, wasdistributed into twopeaks of
ap-proximately equal magnitudeatfractions 9(84S)
and 11 (68S). The ratio of label incorporated
into the RI-aandRI-b bands(Fig. 7C)increased
as afunctionofgreatersedimentationrate, with
RI-abeingthe dominantcomponentin the
sed-imentationregion shown tocontain replicative
intermediates (50to 90S) (see Fig. 3and
refer-ence13).
Characterization of the reaction
prod-ucts by electron microscopy. To determine
thecomposition of the RI-a and RI-b bands,we
excised theseregions directly from a 1.4%
aga-rosegelrun asdescribedin Fig. 7and prepared
theDNA for electron microscopy (see Materials
-RI-i and
Methods).
Of the 22 DNA moleculesob-served from the RI-aband, 78%were
unambig-I 5 7 9 10 11 12 1316 19
f~
-RI-a
-RI-b
-m
-M
I
B
601
40(
c
0
E
cil
0.
-0
204
E
z
0
FRACTION
FIG. 7. Gel electrophoretic analysis ofDNA in
fractions from aminichromosome sucrosegradient.
Portions (20ul) takenfrom thepreparative sucrose
gradientdescribed inFig.3wereanalyzed byagarose
gel electrophoresisasdescribed inFig.6. Electropho-resiswascarriedoutfor 16 h at 2.5 V/cm.
Autora-diography ofthegel (A)wascarried outfor 4°C for
12days. Bandscorrespondingto RI-a(0) andRI-b
(0)werecut outofthegeland melted with 100,ulof
water for 10 min at 100°C, andradioactivity was
quantified by usingBeckman HPfluor (B). The
dot-ted line represents [3H]thymidine incorporated in
vivofrom Fig. 3. The ratiosof isotope incorporated
into RI-a andRI-bareplottedin C.
0
0
-4
0
3~~~~~~;1
I 2~~~~~~~~~~~I
- - -.---
I~~~~~%-~o ClI\
0 5 10 15
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.495.267.442.163.523.2]810 GOURLIE ET AL.
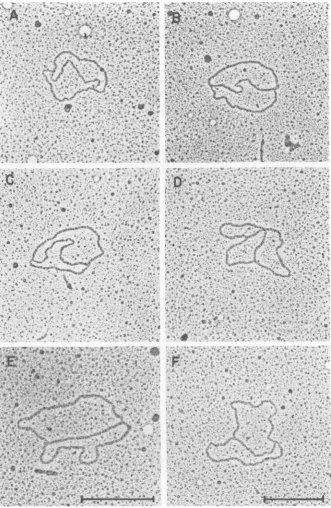
uous9structures. Otherstructuresobserved
in-cluded "connected" monomers (two monomers
joined by a double-stranded linker of variable
length) (6%) and a structures (3%). Figure 8
showsrepresentative micrographs ofthe DNA
moleculesfound in theRI-aband. The putative
replicated regions of the 9 structures ranged
from 46 to87%ofthe total genomelength.Ten
of the 18 unambiguous 9 structures were
clus-tered at 67 +4%replicated. Of the 28 molecules
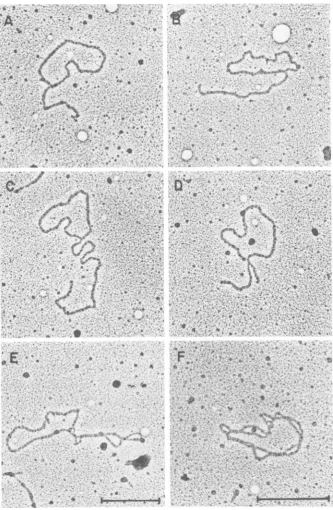
observed from the RI-bband, 48%were
unam-biguousa structures and 18% wereastructures.
Other structures observedinthe RI-b band
in-cludedlinkedandmultiplylinkeddimers and9
structures.Figure9showsrepresentative
micro-graphs ofmoleculesfound in the RI-b band. As
a control, DNA molecules extracted from the
form I band were monomeric, with fewif any
complexstructures.
Characterization of the reaction prod-ucts by
BglI
restriction enzyme digestion.We further characterized the 32P-labeled
reac-tionproducts afterdigestionwiththerestriction
endonuclease BglI. Cleavageofpolyoma DNA
occurs once near the origin ofreplication (10)
andwould convertreplicativeintermediatesinto
H-likestructures.Restrictiondigestionwas
per-formed on the DNA from a sample ofnuclear
extractincubated in vitro with[a-32P]dGTPfor
30 min. Theautoradiogram of this material after
electrophoresis on a 1.2% agarose gel (Fig. 10) shows a gradation of radioactivity extending
fromthe migration position of late Cairns species
totheearliest Cairnsstructuresmigratingclose
to form III. A second spectrum ofreplicative
intermediates wasapparentmigratingin a range
intermediate between the late Cairns species
andformIII. Thisspectrum represents
replica-tive intermediates that hadreplicatedina
uni-directed manner and assumed aY-type
config-uration after digestion with BglI
(Buckler-White, Krauss, Benbow, and Pigiet, submitted for publication). Molecules comigrating with
form IIIincluded all double-stranded linear
mol-ecules produced from cleavage of form I, form
II,oranyconcatenated dimers. Thispatternof
isotope incorporation revealed a spectrum of
replicativeintermediates and was comparable to
that obtained for DNA labeled in vitro with
[ax32P]dGTP
bynuclei prepared by an isotonicmethod(15) or DNAlabeled in vivo with
32Pi.
DISCUSSION
We have demonstrated that
minichromo-somespresent insoluble nuclear extracts
incor-porate deoxynucleotide monophosphates into
polyoma DNA in vitro. The distinctive advan-tage of the soluble system lies in the ability to
isolate mature and replicating
minichromo-somes (50 to90S) as intactnucleoprotein
com-plexes and toidentifyindividual DNA molecules
and enzymeactivitiesassociated with these
com-plexes.
Interpretation of studies usingisolated
mini-chromosomesdependsonunderstandingthe
na-tureof thematerialsextracted and whetherthey
arerepresentativeof the minichromosomes
pres-ent in the intact cell. The lower rate ofDNA
synthesis by the nuclearextractrelativeto
iso-latednuclei(about10%)raises several
questions
about the
synthetic capacity
of theminichro-mosomes. Thesimplest interpretationis that the
low
incorporation
ofdeoxynucleotide
mono-phosphates by the nuclear extract reflects the
low yieldof extraction ofreplicating
minichro-mosomecomplexes. This argument issupported
bytheinitialrateofdeoxynucleotide
monophos-phateincorporation,which wasroughly
propor-tionaltotheefficiency (10to20%)of extraction. Thesyntheticrateofnuclearextracts was
com-parabletothesyntheticrateobserved innuclei afteradjusting for this extraction
efficiency.
Thevalidityofthisargument rests on theassumption
thattheextracted minichromosomesare
repre-sentative of the total
population
of functional complexesinnuclei.The retention of labeled viral DNA in the
nucleus, even after extensive extraction with
hypotonicorhypertonic detergent buffers (1), is
consistent with the finding that substantial
amountsofviral DNA aretenaciouslybound to
the nuclear matrix. If the matrix is the site of
some stages of viral replication,
minichromo-somes may represent a stage with no
require-mentfor matrixattachment.
Alternatively,
ifallstages of replication take place on the matrix
(19), minichromosomes may represent species released from cellsbyourmanipulations. Itwas
ofinterest that
approximately
10%ofthemini-chromosomes were associated with amorphous material (possibly matrix components) even
afterhigh-saltextraction (13). In any event, the
isolated minichromosomes are likely to prove
especiallyvaluable for identification of the
pro-teins involved in replication even if the DNA
synthesiscarried out invitro is not
fully
repre-sentative of events inside the nucleus.Sedimentation analysisof the products of in
vitro DNA synthesis by the nuclear extract
showed that themajority of labeled DNA
sedi-mented with the same distribution
(approxi-mately 84S) as the viral minichromosomes
la-beledduringashort (5-min) pulsein vivo (Fig.
2 and 3). The smaller amount of in vitro label
which sedimented with mature
minichromo-somes (Fig. 2) may correspond either to early
on November 10, 2019 by guest
http://jvi.asm.org/
FIG. 8. Electron microscopeanalysis of DNA molecules comigrating with the RI-a band on agarose gels. TheRI-aband was excised from an agarose gel, dissolved in potassium iodide, and prepared for electron microscopy asdescribed in Materials and Methods. (A) B Structure where the putative unreplicated region measures 0.90 um andthe replicated arms measure 0.76 and 0.76
tum.
(B) BStructure where the putative unreplicated region measures 0.80pm and the replicated armsmeasure 0.90 and 0.85tum.
(C) B Structurewheretheputative unreplicatedregion measures 0.60pmand thereplicatedarmsmeasure 1.03and 0.99 ,um. (D) BStructurewhere theputativeunreplicated region measures 0.59pmand thereplicated arms measure 1.10 and 1.06
tum.
(E) B Structure where theputative unreplicated region measures 0.57pmand thereplicatedarmsmeasure 1.17 and 1.16pm.(F) BStructure where the putative unreplicated region measures 0.49pumand the replicated arms measure 1.20 and
1.19ptm.
Molecules A, B, C, D, and F were printed at the samemagnification;molecule E wasprintedat25% higher magnification. Themarkerscorrespondto 0.5pm.
811
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.495.80.412.55.564.2]812 GOURLIE ET AL.
FIG. 9. Electronmicroscopeanalysisof DNA comigrating with theRI-bbandonagarosegels. The RI-b
band was excisedfromanagarosegel andpreparedforelectron microscopy as described inFig. 8. (A) a
Structure where the circlemeasures 1.70
Aim
and thetailmeasures 0.66,um. (B)aStructure wherethecirclemeasures 1.56,Im and the tail measures 0.99jim.(C)aStructurewhere the circle measures 1.64jimand thetail
measures 1.87jim. (D)aStructure where the circle measures 1.70jimandthetails measure 0.51and0.22jim. (E) Linked dimerstructurewherethe two circles measure 1.74 and 1.62jim.(F)Multiplylinked dimer structure where the circles measure 1.61 and 1.59 jim. Molecules A, B, C, D, and E were printed at the same
magnification;molecule Fwasprintedat25%highermagnification. The markerscorrespondto0.5jum.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.495.93.427.72.583.2]FIG. 10. BglI digestion oftheproducts ofin vitro DNA synthesis. Nuclear extract, incubated in vitro
for60mmnwith[a_3'P]dGTP,wasdeproteinizedand
digested with BglI as described in Materials and Methods. Electrophoresis in 1.2% agarose was
per-formnedat5 V/cmfor18hat4i C.Migration of form
IIIand late Cairns(LC)structuresisindicated.
replicativeintermediatesasdetectedbyelectron
microscopy (13) or to termination products
re-semblingmatureform I DNA(14, 17, 24).
Com-parisonofFig. 3and4showsthatsedimnentation
of viral minichromosomes resolved the
replica-tive intermediates from the mature forms far betterthan sedimentation ofdeproteinizedDNA
(Fig.4).
Agarosegelelectrophoresis of theproductsof DNA synthesis in vitro demonstrated that the
majority oflabel appeared astwo bands (RI-a
and RI-b), each migrating slower than form II
polyoma DNA (25), and that these formswere
also present in RI DNA pulse-labeled in vivo. Thecomplexcontributions of molecularweight, superhelicity, and topology to the behavior of DNAsonagarosegels(4)complicateanycertain identification of these bands at this timne. Evi-dence that they are members of different
con-formnational families (24) is supported by the increased band width observedatloweragarose
concentrations and is consistent with the spec-trum of intermediates seen in the BgllI digest
pattern (Fig. 10) and the electron microscope
evidence discussed below. The kinetics of
label-ingof the RI-a and RI-b bands is consistent with RI-a being an initial product, rather than the
result of conversion from RI-b. In contrast to
the rapid labeling of RI-a, the slower rate of
labeling of RI-b is consistentwithconversionof some RI-a into RI-b and/or conversion from
early replicative intermediates migrating as a
continuum from formI tothe late Cairns posi-tion. The appearanceof RI-b in tworegions of
the sucrose minichromosome gradient (Fig. 7)
mayindicate thatbothof thesepossibilitiesare
true.
The results obtained byelectronmicroscopy
(Fig. 8 and 9) suggest that the RI-a band
con-sisted almost entirely ofunambiguous 6
struc-turesrangingfrom46 to 87% replicated with a
modal value of 67 + 4%
replicated.
The RI-bband was more complex, consistingof a and a
structures which either could have arisen by
breakage of 9 structures or could have been
generated by a
rolling
circle mechanism and branchmigration. In addition, linked and mul-tiply linkeddimeric
species reminiscent of theconformational families described by Sundin
andVarshavsky (24) werealsoseen,althoughat
lowerfrequencies. Althoughwecannot
unequiv-ocally
ruleoutthepossibility that RI-aandRI-b correspond to the 80 and 90% late Cairns species observedinSV40-infectedcells(25), the
kinetics oflabeling and sedimentation properties ofRI-aandRI-b,aswellasthe electron
micros-copy, render this possibility extremely unlikely
for polyoma-infected cells. The results of this
paper areconsistentwith, butdo not prove,the
hypothesis that incorporation into RI-b is due to either breakage ofRI-a or maturation into topologically linked speciesasdescribedby
Sun-din and
Varshavsky
(24), orboth.Mature form I polyoma DNA was not
pro-ducedinvitroto anappreciableextent,
amount-ingtoless than 5%of the total incorporation by
60 min (Fig. 7A). This lack of conversion into
form
Imolecules,
as observedby
others forintactnuclei (2,6,25) andsoluble
minichromo-somesystems (5, 22) maybe due tothe release
of factors essential for termination intothe
cy-toplasm during nuclearisolation.
ACKNOWLEDGMENTS
We acknowledge the contributions of Glen Humphrey, CarolBreaux, and MarvinBaynefortheircritical evaluation of ourworkandmanuscript.We alsoacknowledgethe contri-butionsofMitchellK.Hobish,who initiated thisstudy.
This research was supported by Public Health Service grantsGM-23813 and GM-23970 andpostdoctoral fellowship
CA-06304toB.B.GfromNationalInstitutesofHealth.
LIrERATURE CITED
1. Buckler-White,A.J.,G.N.Humphrey,and V.Pigiet.
1980.AssociationofpolyomaTantigenand DNA with the nuclearmatrixfromlyticallyinfected3T6cells. Cell 22:37-46.
la.Davis,R.W.,M.Simon,and N.Davidson.1971.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.495.123.173.72.301.2]tron microscope heteroduplex methods for mapping regions of base sequence homology in nucleic acids. Methods Enzymol. 21D:413-428.
2. DePamphilis, M. L., P. Beard, and P. Berg. 1975. Synthesis of superhelical simian virus 40 DNA incell
lysates. J.Biol. Chem. 250:4340-4347.
3. DePamphilis, M. L., and P. M. Wasserman. 1980. Replication of eukaryotic chromosomes: a close-up of thereplication fork. Annu. Rev. Biochem. 49:627-666. 4. Dingman, C. W., M. P.Fisher, and T. Kakefuda. 1972.
Role of molecular conformation in determining the electrophoreticproperties ofpolynucleotidesin
agarose-acrylamidegels. Biochemistry11:1242-1250. 5. Edenberg, H. J., M. J. Wagar, and J. Huberman.
1976.Subnuclear systems for synthesis ofSV40 in vitro. Proc.Natl. Acad.Sci. U.S.A. 73:43924396.
6. Franke, B., and T. Hunter.1974.In vitropolyoma DNA synthesis: discontinuous chaingrowth. J. Mol. Biol. 83: 99-121.
7. Germond,J.-E., B.Hirt,P.Oudet, M. Gross-Bellard, and P. Chambon. 1975.Foldingofthe DNAdouble helix inchromatin-like structures from simian virus 40. Proc.Natl. Acad. Sci.U.S.A. 72:1843-1850.
8. Gourlie,B.B., V.Pigiet, C. B. Breaux, M. R. Krauss, C. R.King, and R. M. Benbow.1981.Polyoma virus minichromosomes:associated enzyme activities. J. Vi-rol. 38:826-832.
9. Green,M.H.1972.Biosyntheticpropertiesof apolyoma nucleoproteincomplex: evidence for replication sites. J. Virol. 10:32-41.
10. Griffith,J.D. 1975.Chromatinstructurededuced from aminichromosome.Science 187:1202-1204.
11. Hirt, B. 1967. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 26:365-369. 12.Jaenisch, R.,A.Mayer,and A. Levine.1971.
Replicat-ing SV40 molecules containReplicat-ing closed circular template DNA strands. Nature(London)New Biol. 233:72-75. 13. Krauss,M.R., and R. M. Benbow.1981.Polyoma virus
minichromosomes:associated DNA molecules. J. Virol. 38:815-825.
14. Laipis,P.J., A.Sen,A. J. Levine,and C. Mulder. 1975.DNAreplication in simian virus 40-infectedcells:
the structure of the 16S gap circle intermediate in simian virus40DNAsynthesis. J. Virol. 68:115-123. 15. Magnusson, G., V. Pigiet, E. L. Winnacker, R.
Abrams, and P. Reichard. 1973. RNA-linked short DNAfragments duringpolyoma replication. Proc. Natl. Acad. Sci.U.S.A. 70:412-415.
16. Martin, R. F. 1977. Analysis of polyoma virus DNA replicative intermediates by agarose gelelectrophoresis.
J.Virol. 23:827-832.
17. Meinke, W., and D. A. Goldstein.1972.Studiesonthe structure and formation ofpolyomaDNAreplicative
intermediates.J.Mol. Biol. 61:543-563.
18. Meinke, W.,M. R. Hall,and D. A. Goldstein.1975. Proteins in intracellular simian virus nucleoprotein complexes: comparison withsimianvirus40 core pro-teins. J. Virol. 15:439-442.
19. Pardoll,D.M.,B.Vogelstein,andD.S.Coffey.1980. Afixed site of DNAreplication in eukaryoticcells.Cell 19:527-536.
20.Pigiet, V., R.Eliasson,andP.Reichard.1974.
Repli-cation of polyoma DNA in isolated nuclei. III. The nucleotide sequenceatthe RNA-DNAjunction of nas-centstrands. J. Mol. Biol. 84:197-216.
21. Seidman,M.M.,C. F.Garon,and N. P. Salzman.1978. TheRelationship of SV40 replicating chromosomes to twoformsofthenon-replicating SV40 chromosomes. Nucleic Acids Res. 5:2877-2893.
22. Su,R.T.,and M. L.DePamphilis.1976.In vitro repli-cation of SV40 DNA inanucleoprotein complex. Proc. Natl. Acad. Sci.U.S.A. 73:3466-3470.
23. Su,R.T.,and M. L.DePamphilis.1978.Simian virus40 DNAreplicationinisolatedreplicating viral chromo-somes. J.Virol.28:53-65.
24. Sundin,O., and A.Varshavsky.1980.Terminal stages ofSV40 DNAreplication proceed via multiply inter-twinedcatenated dimers.Cell21:103-114.
25. Tapper,D.P.,and M. L.DePamphilis.1978. Discontin-uousDNAreplication: accumulation of SV40 DNA at specific stages in itsreplication. J. Mol. Biol. 120:401-422.
26. Tegtmeyer, P.1972.Simian virus40DNAsynthesis: the
viralreplicon. J. Virol.10:591-598.
27. Tegtmeyer, P., and F. Macasaet. 1972. SV40 DNA synthesis: analysis bygelelectrophoresis. J. Virol. 10: 599-604.
28. Tsubota, Y.,M.A.Waqar,J.F.Burke,B. I.Milavetz,
M. J.Evans, D.Kowalski,and J. Huberman. 1979. Association of enzymes withreplicating and non-repli-cating SV40 chromosomes. ColdSpring Harbor Symp. Quant. Biol.43:693-704.
29. Waldeck, W.,U.Sparen,G.Mastromei,R.Eliasson,
and P. Reichard.1979.Replicationofpolyoma DNA innuclear extracts andnucleoprotein complexes. J. Mol. Biol. 135:675-689.
30. Winnacker,E.-L., G.Magnusson,and P. Reichard. 1972.Replication of polyomaDNA inisolated nuclei. I. Characterization of the system from mouse fibroblast 3T6cells.J.Mol. Biol. 72:523-537.
![FIG.1.Atsynthesiscpm/pmol)wasextract.andcells) the Kinetics of DNA synthesis by the nuclear Nuclear extract (corresponding to 3.5 x JOr in 500 ,Il was assayed for endogenous DNA in the presence of [a-32P]dTTP (158 x 103 as described in Materials and Method](https://thumb-us.123doks.com/thumbv2/123dok_us/1477944.100428/3.495.157.446.76.307/atsynthesiscpm-wasextract-kinetics-synthesis-corresponding-endogenous-described-materials.webp)