0022-538X/09/$08.00⫹0 doi:10.1128/JVI.01307-09
Copyright © 2009, American Society for Microbiology. All Rights Reserved.
A B-Box 2 Surface Patch Important for TRIM5
␣
Self-Association,
Capsid Binding Avidity, and Retrovirus Restriction
䌤
†
Felipe Diaz-Griffero,
1Xu-rong Qin,
3Fumiaki Hayashi,
3Takanori Kigawa,
3,4Andres Finzi,
1Zoe Sarnak,
1Maritza Lienlaf,
1Shigeyuki Yokoyama,
3,5and Joseph Sodroski
1,2*
Department of Cancer Immunology and AIDS, Dana-Farber Cancer Institute, Department of Pathology, Division of AIDS, Harvard Medical School, Boston, Massachusetts 021151; Department of Immunology and Infectious Diseases, Harvard School of
Public Health, Boston, Massachusetts 021152; Systems and Structural Biology Center, Yokohama Institute, Riken,
1-7-22 Suehiro-cho, Tsurumi, Yokohama 230-0445, Japan3; Department of Computational Intelligence and Systems Science,
Interdisciplinary Graduate School of Science and Engineering, Tokyo Institute of Technology, 2-12-1 Ookayama, Meguro-ku, Tokyo 152-8550, Japan4; and Department of Biophysics and Biochemistry, Graduate School of
Science, University of Tokyo, 2-11-16 yayoi, Bunko-Ku, Tokyo 113-0032, Japan5
Received 25 June 2009/Accepted 29 July 2009
TRIM5␣is a tripartite motif (TRIM) protein that consists of RING, B-box 2, coiled-coil, and B30.2(SPRY) domains. The TRIM5␣rhprotein from rhesus monkeys recognizes the human immunodeficiency virus type 1
(HIV-1) capsid as it enters the host cell and blocks virus infection prior to reverse transcription. HIV-1-restricting ability can be eliminated by disruption of the B-box 2 domain. Changes in the TRIM5␣rhB-box 2
domain have been associated with alterations in TRIM5␣rhturnover, the formation of cytoplasmic bodies and
higher-order oligomerization. We present here the nuclear magnetic resonance structure of the TRIM5 B-box 2 domain and identify an unusual hydrophobic patch (cluster 1) on the domain surface. Alteration of cluster 1 or the flanking arginine 121 resulted in various degrees of inactivation of HIV-1 restriction, in some cases depending on compensatory changes in other nearby charged residues. For this panel of TRIM5␣rhB-box 2
mutants, inhibition of HIV-1 infection was strongly correlated with higher-order self-association and binding affinity for capsid complexes but not with TRIM5␣rhhalf-life or the formation of cytoplasmic bodies. Thus,
promoting cooperative TRIM5␣rhinteractions with the HIV-1 capsid represents a major mechanism whereby
the B-box 2 domain potentiates HIV-1 restriction.
Mammalian proteins that can dominantly block retroviral in-fection by interfering with the early phase of viral replication have been identified (1a, 47, 50). One member of the tripartite motif (TRIM) family of proteins, TRIM5␣, mediates species-specific restriction of particular retroviruses from diverse retrovirus subfamilies. For example, TRIM5␣ from rhesus macaques (TRIM5␣rh) blocks infection of human immunodeficiency virus type 1 (HIV-1) soon after viral entry but prior to reverse tran-scription (22, 50). Although human TRIM5␣(TRIM5␣hu) only weakly inhibits HIV-1 infection, this protein potently inhibits in-fection by N-tropic murine leukemia virus (N-MLV) (16, 39, 42, 47, 57). Some species of Old World and New World monkeys encode TRIMCyp instead of TRIM5␣; TRIMCyp is a fusion of TRIM5 and cyclophilin A and restricts some retroviruses, includ-ing HIV-1 (4, 16, 38, 39, 42, 47, 55–57).
TRIM5␣ is composed of RING, B-box 2, and coiled-coil domains typical of TRIM proteins and a carboxy-terminal B30.2(SPRY) domain. The RING domain is not absolutely required for, but contributes to the potency of, retroviral re-striction; the RING domain plays an important role in the rapid degradation of TRIM5␣(11). Some changes in the B-box
2 domain exert dramatic effects on retrovirus-restricting ability (10, 19, 40). The coiled-coil domain allows oligomerization of TRIM5␣(20, 36), which recently has been shown to be a dimer (16, 39, 42, 47, 57). Dimerization contributes to the avidity of TRIM5␣ for the HIV-1 capsid (51). The B30.2(SPRY) do-main, which mediates capsid recognition, dictates the viral specificity of restriction (46, 52, 58). Sequence divergence within variable regions of the B30.2(SPRY) domain accounts for species-specific differences in the potency of restriction (46, 52, 58).
Structural and functional similarities exist between the RING and B-box domains of TRIM proteins. Both domains bind two zinc atoms tetrahedrally in a cross-brace conforma-tion (3, 17, 34, 35). Furthermore, both domains have been shown to be involved in protein-protein interactions. In non-TRIM proteins, RING domains play the role of molecular scaffolds, allowing formation of supramolecular complexes by self-association, which in some cases improves enzymatic ac-tivity (2, 23, 24, 43). Interestingly, RING-RING interactions have been reported to be functionally important for genomic stability, as in the heterodimer formed by the RING domain of BRCA1 with the RING domain of BARD-1 (5).
The B-box domain is found in extremely diverse proteins; in TRIM proteins, the B-box domain is present in one or two copies (44, 54). B-box domains have been implicated in pro-tein-protein interactions (32, 53) and have been shown to play a role in protein localization, growth control, differentiation, and transcriptional regulation (54). The B-box domain usually adopts a␣conformation reminiscent of the RING domain.
* Corresponding author. Mailing address: Dana-Farber Cancer In-stitute, 44 Binney Street-CLSB 1010, Boston, MA 02115. Phone: (617) 632-3371. Fax: (671) 632-4338. E-mail: [email protected] .edu.
† Supplemental material for this article may be found at http://jvi .asm.org/.
䌤Published ahead of print on 5 August 2009.
10737
on November 8, 2019 by guest
http://jvi.asm.org/
Some TRIM proteins, such as MID1 (TRIM18), contain two B-box domains in tandem. Structural studies demonstrated that the tandem B-boxes of MID1 pack against each other (53). The contact interface between the two B-boxes is made by the short -strands of each B-box domain. Mutations af-fecting either one of these B-box domains occur in patients with Opitz G/BBB syndrome, a severe developmental disorder (48). Recent structural studies of the B-box 2 domain of MuRF1 (TRIM63) identified a dimer interface that consists of a hydrophobic surface and polar residues (46, 52, 58). These findings suggest that B-box–B-box interactions may have func-tional significance.
Using models based on the B-box 1 or B-box 2 structures of other proteins, mutagenesis of putative surface-exposed resi-dues of the TRIM5␣rh B-box 2 domain has been conducted (10, 19, 40). These studies have revealed several functions dependent upon the intact TRIM5␣rhB-box 2 domain. First, the wild-type TRIM5␣ proteins of humans and rhesus mon-keys are turned over rapidly, with a half-life of less than 1 h (11). Alteration of specific basic residues in the TRIM5␣rh
B-box 2 domain results in much more stable proteins; because some of these mutants retain antiretroviral activity, rapid turn-over of TRIM5␣ appears not to be essential to retrovirus-restricting ability (10).
Second, a change in the B-box 2 domain of TRIM5␣ can affect the higher-order self-association of TRIM5␣ dimers (32). This self-association contributes to the ability of TRIM5␣ to bind the retroviral capsid, perhaps by increasing avidity (32). The contribution of the B-box 2 domain to antiretroviral ac-tivity is particularly important when the interactions of the B30.2(SPRY) domain with the viral capsid are weak (31, 32). Like many TRIM proteins (44), TRIM5␣rhforms aggregates called cytoplasmic bodies when overexpressed (11). A third phenotype associated with TRIM5␣B-box 2 changes is a de-crease in the propensity of the protein to coalesce into cyto-plasmic bodies; this phenotype is particularly striking, because many TRIM5␣ B-box 2 mutants are expressed at higher steady-state levels than the wild-type protein (10). Although cytoplasmic bodies per se are not necessary for TRIM5␣ -me-diated retroviral restriction (46, 52, 58), the relationship be-tween higher-order TRIM5␣ self-association, which contrib-utes to capsid binding (32), and the tendency to form cytoplasmic aggregates is not clear.
Lastly, some TRIM5␣B-box 2 mutants apparently lose the ability to block retrovirus infection out of proportion to any measured reductions in capsid binding (10, 32). This observa-tion hints that the B-box 2 domain may mediate funcobserva-tions beyond higher-order self-association and capsid binding. Pre-sumably, such functions could contribute mechanistically to the process whereby TRIM5␣disrupts the early events in retrovi-rus infection following the recognition of the capsid after it enters the cytosol. Studies following the fate of the HIV-1 capsid in the cytosol of infected cells have correlated restric-tion of infecrestric-tion with a decrease in the amount of cytosolic particulate capsids (51). Similar changes have been observed in the amount of cytosolic particulate capsids during infection of HIV-1 and N-MLV in the presence of the restriction factors TRIMCyp and TRIM5␣hu, respectively (10, 41). In contrast to the wild-type TRIM5␣huprotein, a TRIM5␣huB-box 2 mutant defective for N-MLV restriction did not cause a reduction in
the amount of particulate capsid in the cytosol (46, 52, 58). Thus, B-box 2 functions may contribute to the accelerated uncoating of the retroviral capsid associated with TRIM5␣ restriction.
We present here a nuclear magnetic resonance (NMR) structure of the TRIM5 B-box 2 domain and demonstrate a weak tendency of the purified B-box 2 domains to self-associ-ate. Alteration of the unusually hydrophobic surface or flank-ing charged amino acid residues of the TRIM5␣rh B-box 2 domain revealed structures important for rapid turnover, higher-order self-association, cytoplasmic body formation, cap-sid binding, and retrovirus restriction. To understand which B-box 2 functions contribute to retroviral restriction, the rela-tionships among these TRIM5␣properties were investigated. Both higher-order self-association and binding to the viral capsid strongly correlated with inhibition of HIV-1 infection, implying that the B-box 2 domain potentiates retroviral restriction by pro-moting cooperative TRIM5␣interactions with the capsid.
MATERIALS AND METHODS
Sample preparation of the TRIM5 B-box 2 domain. The DNA fragment
encoding the B-box 2 of human TRIM5␣(amino acid residues S86 to T131
[Swiss-Prot Q9C035]) was amplified via PCR from the Research Association for Biotechnology Clone FLJ14687 and was cloned into the plasmid vector pCR2.1 (Invitrogen, Carlsbad, CA) as a fusion with an N-terminal His tag and a tobacco
etch virus protease cleavage site. The13
C- and15
N-labeled protein was synthe-sized by a cell-free protein expression system (25). Purification was performed by a standard procedure (29). For structure determination, a single 1.13 mM
uni-formly13C- and15N-labeled sample was prepared in 20 mM Tris-HCl buffer (pH
7.0), 100 mM NaCl, 1 mM dithiothreitol (DTT), 0.02% NaN3, 0.05 mM ZnCl2,
and 1 mM iminodiacetic acid, with the addition of D2O to 10% (vol/vol). The
engineered protein sample used for the NMR measurements was comprised of 46 amino acid residues from human TRIM5 spanning the B-box 2 domain
(residues 86 to 131 of TRIM5␣hu); the first seven and last six residues of the
protein consist of tags. The residues of this engineered protein are referred to as 79 to 137, so that the numbers of the B-box 2 residues correspond to those of human TRIM5.
NMR spectroscopy, structure determination, and analysis.All of the NMR spectra were recorded at 20°C on Varian INOVA 600, 800, and 900 spectrometers equipped with pulsed-field gradient triple-resonance probes. Sequence-specific res-onance assignments were made using the standard triple-resres-onance techniques. The backbone assignment was achieved by the combined analysis of HNCO, (HCA)CO (CA)NH, HN(CO)CA, HNCACB, and CBCA(CO)NH spectra. The side chain resonances were identified by the combinational use of HBHANH, HBHA (CO)NH, (H)CC(CO)NH, (H)CCH-TOCSY, HCCH-TOCSY, and
two-dimen-sional1
H-15
N HSQC and1
H-13
C HSQC spectra. Nuclear Overhauser effect (NOE)
data for structure determination were extracted from three-dimensional15N- and
13
C-edited NOESY spectra, recorded with mixing times of 75 and 65 ms, respec-tively.
The NMRpipe software package (7) and the program KUJIRA (26), created on the basis of NMRView (21), was used for optimal visualization and spectral analysis. Automated NOE cross-peak assignments (18) and structure calcula-tions with the torsion angle dynamics were performed using the software package CYANA2.0.17 (15).
Dihedral angle restraints were derived using the program TALOS (6). A total of 100 conformers were calculated independently. The 20 conformers with the lowest final CYANA target function values were finally selected. The structures were validated by using PROCHECK-NMR (28). The program MOLMOL (27) was used to analyze the resulting 20 conformers and to prepare drawings of the structures, unless noted otherwise in the legends.
The selected 20 conformers have been deposited in the Protein Data Bank (PDB entry 2YRG).
Dynamic light-scattering experiment.The data were measured at 4°C by using a DynaPro light scattering system (Protein Solutions, Lakewood, NJ) and a microsampler, with a protein concentration of 2.2 mg/ml in 20 mM Tris-HCl buffer (pH 7.0) containing 100 mM NaCl and 1 mM iminodiacetic acid. Dynam-ics Version 5.0 software (Protein Solutions) was used in the data collection and analysis.
10738 DIAZ-GRIFFERO ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
Analytical ultracentrifugation.Sedimentation equilibrium experiments were performed on a Beckman Optima XL-I centrifuge, in an eight-position An-50Ti rotor with equilibrium six-channel centerpieces and quartz windows. Experi-ments were carried out at 4°C with the B-box 2 domain protein in solution at concentrations of 1.2, 0.8, 0.4, and 0.2 mg/ml and at rotor speeds of 20,000, 22,000, and 24,000 rpm for the dimeric molecular weight range and 30,000, 31,000, and 32,000 rpm for the monomeric molecular weight range. The data were corrected after 12, 14, and 16 h at each speed. Protein concentration profiles were monitored by the absorbance at 280 nm. To generate the baseline, the samples were then completely sedimented at 40,000 rpm for 6 h at 4°C. For the calculation of the molecular weight, corrected datasets were analyzed in the self-association model with a partial specific volume of 0.705 ml/g and a solvent density of 1.003 g/ml, using the Beckman XL-A/XL-I data analysis software (version 6.03).
Creation of cells stably expressing TRIM5␣variants.Retroviral vectors
en-coding wild-type or mutant rhesus monkey TRIM5␣rhproteins were created
using the pLPCX vector. The TRIM5␣rhproteins contained an influenza
hem-agglutinin (HA) epitope tag at the C terminus or FLAG epitope tags at the N terminus. Recombinant viruses were produced in 293T cells by cotransfecting the pLPCX plasmids with the pVPack-GP and pVPack-VSV-G packaging plasmids (Stratagene). The pVPack-VSV-G plasmid encodes the vesicular stomatitis virus G envelope glycoprotein, which allows efficient entry into a wide range of
ver-tebrate cells (59). Cf2Th canine thymocytes were transduced and selected in 5g
of puromycin (Sigma)/ml.
Infection with viruses expressing GFP. Recombinant HIV-1 and N-MLV expressing green fluorescent protein (GFP) were prepared as described previ-ously (12, 50). All recombinant viruses were pseudotyped with the vesicular
stomatitis virus G glycoprotein. For infections, 3⫻104
Cf2Th cells seeded in 24-well plates were incubated at 37°C with virus for 24 h. Cells were washed and returned to culture for 48 h and then subjected to fluorescence-activated cell sorting (FACS) analysis with a FACScan (Becton Dickinson). HIV-1 and N-MLV viral stocks were titrated by serial dilution on Cf2Th cells to determine the concentration of infectious viruses.
Protein analysis.Cellular proteins were extracted with radioimmunoprecipi-tation assay (RIPA) buffer (10 mM Tris [pH 7.4], 100 mM NaCl, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate [SDS], 1% NP-40, 2 mg of aprotinin/ ml, 2 mg of leupeptin/ml, 1 mg of pepstatin A/ml, 100 mg of phenylmethylsul-fonyl fluoride/ml). The cell lysates were analyzed by SDS-polyacrylamide gel electrophoresis (PAGE; 10% acrylamide), followed by blotting onto nitrocellu-lose membranes (Amersham Pharmacia Biotech). Detection of protein by West-ern blotting utilized monoclonal antibodies directed against the hemagglutinin (HA) epitope tags (Roche) and FLAG epitope tags (Sigma) and monoclonal
antibodies to-actin (Sigma). All monoclonal antibodies were directly
conju-gated to horseradish peroxidase. Proteins were detected by enhanced chemilu-minescence (NEN Life Sciences Products) using the FluorChem FC2 detection system (Alpha Innotech). Signals were acquired as an image file (TIFF) and quantified by using QuantityOne software (Bio-Rad Laboratories).
TRIM5␣dimerization.Approximately 107293T cells transfected with 5g of
plasmids expressing TRIM5␣rhvariants were lysed in NP-40 lysis buffer (0.5%
NP-40 in phosphate-buffered saline [PBS]) and cross-linked with different con-centrations (0to 10 mM) of ethylene glycolbis-succinimidylsuccinate (EGS) for 30 min as previously described (10). The cross-linking reaction was stopped by
adding 10l of 1 M Tris-HCl. After cross-linking, the samples were resuspended
in 2⫻sample buffer, followed by incubation at 37°C for 30 min. Samples were
analyzed by SDS-PAGE and Western blotting with an anti-HA antibody (Roche).
Higher-order self-association of TRIM5␣.Human 293T cells were indepen-dently transfected with plasmids encoding FLAG-tagged and HA-tagged mutant
and wild-type TRIM5␣rh proteins. After 48 h, the cells expressing each
TRIM5␣rhvariant were lysed in 1 ml of whole-cell extract buffer (50 mM Tris
[pH 8.0], 280 mM NaCl, 0.5% IGEPAL–10% glycerol, 1 mM DTT, protease inhibitor cocktail [Roche]). Lysates were centrifuged at 14,000 rpm for 1 h at 4°C. Post-spin lysates were then precleared using protein A-agarose (Sigma) for 1 h at 4°C; a small aliquot of each of these lysates was stored as an INPUT sample. Precleared lysates containing the differently tagged proteins were mixed in a 1/1 ratio and incubated with anti-FLAG-agarose beads (Sigma) for 2 h at 4°C to precipitate the FLAG-tagged proteins. Beads containing the immunoprecipitate were washed four times in whole-cell extract buffer. Subsequently, immune
complexes were eluted using 200g of FLAG tripeptide/ml in whole-cell extract
buffer. The eluted samples were separated by SDS-PAGE and analyzed by Western blotting using anti-HA or anti-FLAG antibodies conjugated to horse-radish peroxidase.
TRIM5␣ turnover.The turnover rates of wild-type and mutant TRIM5␣ proteins were estimated by pulse-chase experiments. Cf2Th cell lines stably expressing TRIM5 proteins were metabolically labeled for 30 min with 0.1 mCi
of [35S]methionine-cysteine (35S protein-labeling mix; Perkin-Elmer)/ml in
Dul-becco modified Eagle medium lacking methionine and cysteine and supple-mented with 5% dialyzed fetal bovine serum. The cells were then chased for different time intervals in Dulbecco modified Eagle medium containing an excess of unlabeled methionine and cysteine. Cells were subsequently lysed in a
mod-ified RIPA buffer (140 mM NaCl, 8 mM Na2HPO4, 2 mM NaH2PO4, 1% NP-40,
0.25% SDS). Samples were subsequently immunoprecipitated with anti-HA an-tibody-Sepharose beads for 16 h at 4°C. After two washes with modified RIPA buffer, samples were separated by SDS-PAGE (Invitrogen) and analyzed by autoradiography and PhosphorImager (Molecular Dynamics).
Localization of wild-type and mutant TRIM5␣proteins.The localization of
TRIM5␣rhand variants was investigated as previously described (8). Briefly, cells
were grown overnight on 12-mm-diameter coverslips and fixed in 3.9% parafor-maldehyde (Sigma) in PBS (Cellgro) for 30 min. Cells were washed in PBS, incubated in 0.1 M glycine (Sigma) for 10 min, washed in PBS, and permeabilized with 0.05% saponin (Sigma) for 30 min. Samples were blocked with 10% donkey serum (Dako, Carpinteria, CA) for 30 min and incubated for 1 h with antibodies. The anti-HA fluorescein isothiocyanate-conjugated 3F10 antibody (Roche) was used to stain HA-tagged proteins. Subsequently, samples were mounted for fluorescence microscopy by using the ProLong Antifade kit (Molecular Probes, Eugene, OR). Images were obtained with a Bio-Rad Radiance 2000 laser
scan-ning confocal microscope with Nikon 60⫻N.A.1.4 optics.
HIV-1 CA-NC expression and purification.The HIV-1 CA-NC protein was expressed, purified, and assembled as previously described (13, 14). The pET11a expression vector (Novagen) expressing the CA-NC protein of HIV-1 was used
to transform BL-21(DE3)E. coli. CA-NC expression was induced with 1 mM
IPTG (isopropyl--D-thiogalactopyranoside) when the culture reached an optical
density of 0.6 at 600 nm. After 4 h of induction, the cells were harvested and
resuspended in 20 mM Tris-HCl (pH 7.5), 1M ZnCl2, 10 mM
2-mercaptoeth-anol, and protease inhibitors (Roche). Lysis was performed by sonication, and
debris was pelleted for 30 min at 35,000⫻g. Nucleic acids were stripped from the
solution by using 0.11 equivalents of 2 M (NH4)2SO4and the same volume of
10% polyethylenimine. Nucleic acids were removed by stirring and
centrifuga-tion at 29,500⫻gfor 15 min. The protein was recovered by addition of 0.35
equivalents of saturated (NH4)2SO4. The protein was centrifuged at 9,820⫻gfor
15 min and resuspended in 100 mM NaCl, 20 mM Tris-HCl (pH 7.5), 1M
ZnCl2, and 10 mM 2-mercaptoethanol. The CA-NC protein was dialyzed against
50 mM NaCl, 20 mM Tris-HCl (pH 7.5), 1M ZnCl2, and 10 mM
2-mercapto-ethanol, and stored at⫺80°C.
In vitro assembly of CA-NC complexes.HIV-1 CA-NC particles were assem-bled in vitro by diluting the CA-NC protein to a concentration of 0.3 mM in 50
mM Tris-HCl (pH 8.0), 0.5 M NaCl, and 2 mg of DNA oligo-(TG)50/ml. The
mixture was incubated at 4°C overnight and centrifuged at 8,600⫻gfor 5 min.
The pellet was resuspended in assembly buffer (50 mM Tris-HCl [pH 8.0], 0.5 M NaCl) at a final protein concentration of 0.15 mM (13, 14) and stored at 4°C until needed.
Binding of TRIM5␣rhvariants to HIV-1 capsid complexes.293T cells were
transfected with plasmids expressing wild-type or mutant TRIM5␣rhproteins. At
48 h after transfection, cell lysates were prepared as follows: previously washed cells were resuspended in hypotonic lysis buffer (10 mM Tris [pH 7.4], 1.5 mM
MgCl2, 10 mM KCl, 0.5 mM DTT). The cell suspension was frozen, thawed, and
incubated on ice for 10 min. Afterward, the lysate was centrifuged at full speed
in a refrigerated Eppendorf microcentrifuge (⬃14,000⫻g) for 5 min. The
supernatant was supplemented with 1/10 volume of 10⫻PBS and then used in
the binding assay. In some cases, samples containing the TRIM5␣rhvariants were
diluted with extracts prepared in parallel from untransfected cells. To test
bind-ing, 5l of CA-NC particles assembled in vitro were incubated with 200l of cell
lysate at room temperature for 1 h. A fraction of this mixture was stored (input).
The mixture was spun through a 70% sucrose cushion (70% sucrose, 1⫻PBS,
and 0.5 mM DTT) at 100,000⫻gin an SW55 rotor (Beckman) for 1 h at 4°C.
After centrifugation, the supernatant was carefully removed and the pellet was
resuspended in 1⫻SDS-PAGE loading buffer (pellet). The level of TRIM5␣rh
proteins was determined by Western blotting with an anti-HA antibody, as described above. The level of HIV-1 CA-NC protein in the pellet was assessed by Western blotting with an anti-p24 CA antibody.
Quantitative real-time PCR.Cf2Th cells expressing wild-type and mutant
TRIM5␣rhproteins were challenged with HIV-1 at a multiplicity of infection of
0.2. Viruses were pretreated with DNase to prevent contamination from car-ryover plasmid DNA. An infection using a virus that was heat-inactivated (60°C for 30 min) was performed as a control for carryover plasmid DNA in the PCR.
on November 8, 2019 by guest
http://jvi.asm.org/
After 6 h, the cells were lysed and DNA was extracted by using a Qiagen blood tissue DNA extraction kit. PCRs were prepared by using the QuantiTect probe PCR kit. Each sample contained 100 ng of total cellular DNA. PCR was carried
out using two primers that amplify a 263-bp fragment of GFP: GFP-fwd, 5⬘-GAC
GTA AAC GGC CAC AAG-3⬘; GFP-rev, 5⬘-GGT CTT GTA GTT GCC GTC
GT-3⬘; and GFP-Probe, 5⬘-56-FAM-CCT ACG GCA AGC TGA CCC
TGA-36-TAMRA-3⬘. A calibration curve was prepared using an HIV-1-GFP plasmid.
RESULTS
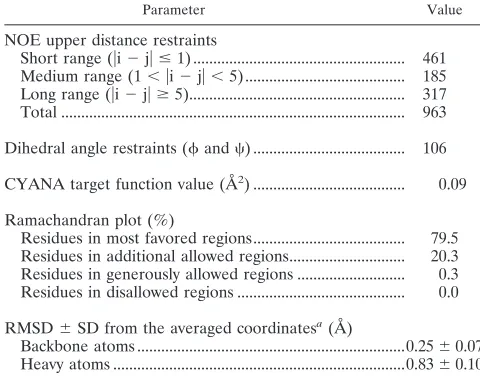
Solution structure of the TRIM5 B-box 2 domain.A frag-ment (residues 86 to 131) of human TRIM5 that encompasses the B-box 2 domain was expressed and labeled in a cell-free system. The solution structure of the monomeric protein was determined by using multidimensional NMR spectroscopy (PDB entry 2YRG). The1H-15N and 1H-13C HSQC spectra
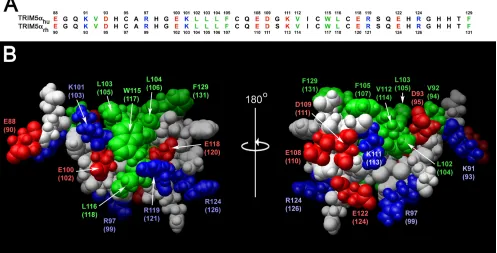
were broad for the size of the protein, but all of the resonances were assigned except for the amide proton of Q121. The so-lution structures were well defined from residues 93 to 131 (Fig. 1). The root mean square deviations (RMSDs) from the mean structure were 0.25⫾ 0.07 Å for backbone atoms and 0.83⫾ 0.10 Å for heavy atoms in the defined region of the domain. The structural statistics are summarized in Table 1. Like most B-box domains (34, 35), the B-box 2 domain from TRIM5 adopts a␣RING-like fold, which is maintained by two zinc ions and CHCDC2H2-type zinc coordination. There is also an additional very short-strand just before the helix. A noteworthy feature of the TRIM5␣B-box 2 domain is that the majority of hydrophobic residues are solvent exposed and clustered, forming two hydrophobic patches on the domain surface. These hydrophobic surface patches are conserved in the B-box 2 domains of TRIM5␣proteins from different
[image:4.585.86.502.67.320.2]spe-cies; for example, the human and rhesus monkey B-box 2 domains differ in only one amino acid residue, which does not reside within either hydrophobic patch (Fig. 2). One of the hydrophobic patches (cluster 1) is located on the outer surface of the helix and the first -strand and is formed in human TRIM5␣by residues L103, L104, W115, and L116. The cluster 1 patch is surrounded by the negatively charged residues E118 and E100 and positively charged residues R119 and K101. The
FIG. 1. Solution structure of the human TRIM5 B-box 2 domain. (A) Superposition of 20 NMR structures showing the␣-helix in red, the -strands in blue, and the Zn2⫹in green. (B) Ribbon diagram of the NMR structure shown in panel A from the same perspective. The Zn2⫹
coordinating residues (CHCDC2H2) are also shown in green. (C) Electrostatic mapping of the B-box 2 surface, highlighting the positions of the positive (blue), negative (red), or neutral (white) charges.
TABLE 1. Summary of conformational constraints and statistics of the final 20 structures of the B-box 2 domain from TRIM5␣
Parameter Value
NOE upper distance restraints
Short range (兩i⫺j兩ⱕ1) ... 461
Medium range (1⬍兩i⫺j兩⬍5) ... 185
Long range (兩i⫺j兩ⱖ5)... 317
Total ... 963
Dihedral angle restraints (and) ... 106 CYANA target function value (Å2) ... 0.09
Ramachandran plot (%)
Residues in most favored regions... 79.5 Residues in additional allowed regions... 20.3 Residues in generously allowed regions ... 0.3 Residues in disallowed regions ... 0.0
RMSD⫾SD from the averaged coordinatesa(Å)
Backbone atoms ...0.25⫾0.07 Heavy atoms ...0.83⫾0.10
a
That is, for residues 93 to 131.
10740 DIAZ-GRIFFERO ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.299.539.525.713.2]second hydrophobic patch (cluster 2) is formed in human TRIM5␣by L102, V112, F105, and F129.
Concentration dependence of NMR signals.The residues on the human TRIM5 B-box 2 protein exhibited broadened sig-nals in the1H-15N HSQC spectrum; this contrasts with
previ-ous observations with other B-box 2 domains, which usually give sharp signals because of their compact size and stable fold (34). To investigate the basis of the observed broadening, the concentration dependence of the 1H-1D and 1H-15N HSQC
spectra was measured (Fig. 3A). As the protein concentration decreased, the peaks of the amide protons from K101, L116, and W115 and side chain CH protons from L116 were shifted; the broadened peaks of the amide protons from L104 and E118, and the imino proton from the W115 indole ring, were sharpened or appeared. These results indicate that the B-box 2 domains associated via the cluster 1 region in the millimolar range of concentration and dissociated at lower concentrations (Fig. 3B). We also observed additional chemical shift pertur-bations (CSPs) in the very low concentration range⬍100M) involving the very narrow region near E108 and H127; this phenomenon may result from the zinc-mediated B-box 2 asso-ciation at H127 (see below).
Size and affinity of the associated state of the B-box 2 do-main.To estimate the oligomeric state of the associated B-box 2 domain in solution, we performed a high-speed sedimenta-tion equilibrium study by analytical ultracentrifugasedimenta-tion (Fig. 3C). The radial distributions at 20,000, 22,000, and 24,000 rpm with a B-box 2 domain concentration of 1.2 mg/ml (⬃180M)
fitted to a molecular mass of 1.05⫻104Da, which corresponds
to 1.57 molecules of the B-box 2 domain. This implies that the B-box 2 domain exists between monomeric and dimeric forms in solution at this concentration. In addition, dynamic light scattering experiments revealed that the B-box 2 domain in solution had a monomodal profile with a narrow peak corre-sponding to particles with an average hydrodynamic diameter of 3.3 nm and an estimated molecular mass of 11 kDa (data not shown). Although it is difficult to calculate an exact molecular mass by dynamic light scattering, this result is consistent with the TRIM5 B-box 2 domain existing in solution at an average size between those of monomers and dimers. The partial dimer content of TRIM5 B-box 2 solutions may reflect a zinc-medi-ated self-association of the B-box 2 domain, because we de-tected a chemical shift change specifically in H127 in the low protein concentration range (⬍100M); in contrast, we did not observe any additional chemical shift changes in other residues in the concentration range from 10 to 660M (data not shown). We also found that reducing the amount of the weak chelator iminodiacetic acid in the buffer resulted in a much higher average molecular mass in the sedimentation equilibrium measurement and reduced solubility of the B-box 2 protein (data not shown). These results support the inter-pretation that most of the dimer estimated from the sedimen-tation equilibrium data resulted from zinc-mediated associa-tion. Collectively, the results suggest two types of B-box association, a zinc-mediated association and a direct associa-tion involving the region from W115 to E118. The
zinc-medi-FIG. 2. B-box 2 domains of human and rhesus monkey TRIM5. (A) The human and rhesus monkey TRIM5 B-box 2 domain sequences are aligned and numbered. Residues with surface-accessible side chains are colored as follows: green, hydrophobic; red, acidic; and blue, basic. (B) A Corey-Pauling-Koltun model of the human and rhesus monkey TRIM5 B-box 2 domain is shown. The residue numbers correspond to those of human TRIM5, with the residue numbers of rhesus monkey TRIM5 in parentheses. The orientations of the left and right figures are the same as those of the upper and lower figures, respectively, in Fig. 1; the right image represents a rotation of the left image 180° around the vertical axis. Acidic and basic residues are shown in red and blue, respectively. Hydrophobic residues are shown in green. Cluster 1 is evident in the left figure, and cluster 2 is evident in the right figure.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.585.45.542.74.327.2]10742
on November 8, 2019 by guest
http://jvi.asm.org/
ated association appears to exhibit a higher affinity but affects the HSQC spectra less. The direct association has a lower affinity but causes major concentration-dependent shifts of NMR peaks that reflected the dynamic equilibrium between zinc-mediated associates.
Since the zinc-mediated association did not cause CSPs ex-cept for H127, the CSP data in the other areas of the domain could be fitted independently. The equilibrium dissociation constant (KD) was calculated to be 9.33 ⫾ 5.40 mM using gnuplot in the dimer model by the analysis of chemical shift changes of the W115 amide proton in the concentration-de-pendent two-dimensional 1H-15N HSQC spectra (Fig. 3D).
The fitting error was very large because of the low affinity and limited solubility of the protein. We estimated that only ca. 10% of the protein contributed to dimerization even at the highest concentration (0.66 mM). This explains the small ob-served chemical shift changes, even though the indole ring of W115 is apparently included in the binding interface. The small chemical shift changes of the backbone amide protons also suggest that the association does not involve a change of the protein fold and is stabilized by inter-side chain interac-tions.
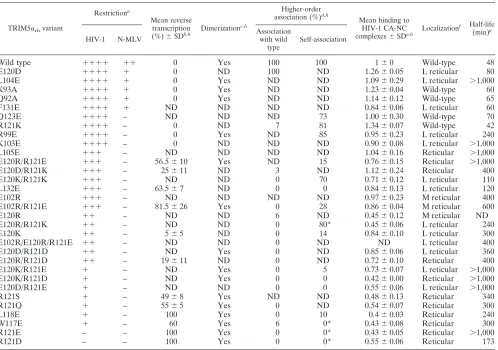
Mutagenic analysis of the TRIM5␣rh B-box 2 surface. To
investigate the functional implications of the TRIM5 B-box struc-ture, we modified specific amino acid residues of TRIM5␣rh. We chose the TRIM5␣rhprotein because it potently restricts HIV-1
infection (50), allowing quantitation of phenotypes over a wide dynamic range. Moreover, most of the assays to evaluate TRIM5␣phenotypes have been developed using the rhesus mon-key protein as a prototype (11, 32, 36, 50, 51). Because the TRIM5␣rh and TRIM5␣hu B-box 2 primary amino acid se-quences differ by only one residue (see the alignment in Fig. 2A), structure-function relationships are expected to be similar be-tween these two domains. Amino acid residues predicted to be surface accessible on the TRIM5␣rhB-box 2 domain, including those predicted to reside in clusters 1 and 2, were altered (Fig. 2B). Particular attention was paid to the charged residues sur-rounding cluster 1, with various combinations of charge substitu-tions being introduced into this region. The wild-type and mutant TRIM5␣rhproteins were expressed stably in Cf2Th canine
thy-mocytes, which do not express a TRIM5 ortholog (45). All of the TRIM5␣rhmutants studied were expressed at least as well as the
wild-type TRIM5␣rhprotein (see Fig. S1 in the supplemental material). The steady-state levels of expression of several of the B-box 2 mutants were greater than that of wild-type TRIM5␣rh,
which has a half-life of⬃48 min (see Fig. S2 in the supplemental material). Indeed, prolonged half-lives, relative to that of the wild-type TRIM5␣rh, were associated with many of the TRIM5␣
mutants (Table 2). This result is consistent with previous studies that implicated the TRIM5␣B-box 2 domain in the regulation of TRIM5␣turnover (10, 11). All of the TRIM5␣mutants tested formed dimers detected by EGS cross-linking (see Fig. S3 in the supplemental material and Table 2). This result is consistent with previous studies indicating that the TRIM5␣B-box 2 domain can be deleted without disrupting dimerization (10).
Retroviral restriction by TRIM5␣rh B-box 2 mutants. To
examine the ability of the TRIM5␣rhB-box 2 mutants to
in-hibit retroviral infection, Cf2Th cells expressing these mutants were challenged with recombinant HIV-1 expressing GFP. The TRIM5␣rhmutants exhibited a range of HIV-1-restricting abil-ities (Fig. 4), and are rank-ordered in Table 2 according to restriction potency. Substitution of an acidic residue for argi-nine 121 (R121E and R121D) completely eliminated the abil-ity of TRIM5␣rhto inhibit HIV-1 infection, as has been pre-viously seen (10, 32). Of note, some changes in the glutamic acid residues at 102 and 120, which flank the hydrophobic patch in the cluster 1 region, restored various degrees of HIV-1-restricting activity to the R121D or R121E mutants (Table 2). For TRIM5␣rhmutants with an acidic residue at position
121, an arginine substitution for glutamic acid 120 resulted in the best improvement in HIV-1 restriction; lysine and aspartic acid substitutions at residue 120 were less effective. An argi-nine substitution for E102 was as effective as an argiargi-nine sub-stitution for E120 in restoring HIV-1-restricting activity to TRIM5␣rhvariants with the R121E change. Thus, while the
presence of three negatively charged residues at positions 102, 120, and 121 was associated with only a low level of TRIM5␣rh
function, arginine substitutions at position 102 or 120 could partially compensate for the presence of an acidic residue at position 121.
The TRIM5␣rhmutants that retained the wild-type arginine or a lysine residue at position 121 exhibited HIV-1-restricting ability, regardless of the particular amino acid at residue 120. However, the wild-type glutamic acid and aspartic acid resi-dues at position 120 generally allowed the most potent HIV-1 restriction in this case. The alteration of glutamic acid 102 to arginine resulted in only a slight decrease in the potency of HIV-1 restriction when the wild-type arginine 121 was retained on TRIM5␣rh. These results indicate that the nature of the
FIG. 3. HSQC perturbation analysis in concentration-dependent experiments. (A) Eleven1H-15N HSQC spectra were measured at protein
concentrations of 660, 560, 490, 400, 300, 250, 170, 110, 40, 20, and 10M and superimposed, with colors ranging from purple (highest concentration) to green (lowest concentration). Signal appearances were observed for the backbone amide proton of E118 and imino proton of the W115 indole ring at the lower concentrations. Chemical shift changes and sharpening were observed for other signals. (B) Mapping of perturbed residues on the B-box 2 structure. In the figure on the left, the size of the CSP is indicated by color. The red color means more than 0.03 ppm of CSP were observed, and yellow indicates more than 0.01 ppm of CSP. The magenta color indicates the appearance of the signal at a lower concentration. Most of the implicated residues are proximal to cluster 1 on one face of the B-box 2 domain (dotted circle on the electrostatic surface on the right). (C) Equilibrium analytical ultracentrifugation of the TRIM5 B-box 2 domain. The radial distribution of the absorbance in the centrifuge cell at equilibrium at 22,000 rpm, with a protein concentration of 1.2 mg/ml (⬃180M), is shown in the lower panel. The red line through the data represents the fit to a single species with a molecular weight corresponding to 10,502; the theoretical molecular weight for the single polypeptide chain is 6,682. The upper panel shows the residuals for the fit. (D) The concentration dependence of chemical shift perturbation for the W115 backbone amide proton in1H-15N HSQC spectra. Total chemical shift change was calculated by using the relation: ⌬␦tot⫽[(␦HN)
2⫹(␦ N/6.5)
2]1/2(1). Curve fitting was carried out by using the program gnuplot. The calculated equilibrium dissociation constant
KDwas 9.33⫾5.40 mM (see the text).
on November 8, 2019 by guest
http://jvi.asm.org/
charged residues that rim the hydrophobic center of cluster 1 influence the efficiency of restriction by TRIM5␣rh.
Of the hydrophobic B-box 2 residues that were altered, changes in W117 and L118, in the cluster 1 hydrophobic patch, exerted the greatest impact on HIV-1-restricting activity. Thus, the TRIM5␣rhB-box 2 residues important for HIV-1
restric-tion (arginine 121, leucine 118, and tryptophan 117) occupy a localized area on the domain surface.
The TRIM5␣rhB-box 2 mutants exhibited a range of HIV-1-restricting activities. The ability of the mutants to restrict N-MLV infection was examined. The wild-type TRIM5␣rh protein exhibited a modest level of restriction against N-MLV (Table 2). Whereas a few of the B-box 2 mutants that exhibited wild-type anti-HIV-1 activity retained the ability to inhibit N-MLV weakly, most of the mutants were devoid of detectable N-MLV-restricting ability. Thus, several of the mutants,
in-TABLE 2. Phenotypes of TRIM5␣B-box 2 mutants
TRIM5␣rhvariant
Restrictiona
Mean reverse transcription
(%)⫾SDb,h Dimerization
c,h
Higher-order
association (%)d,h
Mean binding to HIV-1 CA-NC
complexes⫾SDe,h Localization
f Half-life
(min)g
HIV-1 N-MLV
Association with wild
type
Self-association
Wild type ⫹⫹⫹⫹ ⫹⫹ 0 Yes 100 100 1⫾0 Wild-type 48
E120D ⫹⫹⫹⫹ ⫹ 0 ND 100 ND 1.26⫾0.05 L reticular 80
L104E ⫹⫹⫹⫹ ⫹ 0 Yes ND ND 1.09⫾0.29 L reticular ⬎1,000
K93A ⫹⫹⫹⫹ ⫹ 0 Yes ND ND 1.23⫾0.04 Wild-type 60
Q92A ⫹⫹⫹⫹ ⫹ 0 Yes ND ND 1.14⫾0.12 Wild-type 65
F131E ⫹⫹⫹⫹ ⫹ ND ND ND ND 0.84⫾0.06 L reticular 60
Q123E ⫹⫹⫹⫹ – ND ND ND 73 1.00⫾0.30 Wild-type 70
R121K ⫹⫹⫹⫹ – 0 ND 7 81 1.34⫾0.07 Wild-type 42
R99E ⫹⫹⫹⫹ – 0 Yes ND 85 0.95⫾0.23 L reticular 240
K103E ⫹⫹⫹⫹ – 0 ND ND ND 0.90⫾0.08 L reticular ⬎1,000
L105E ⫹⫹⫹ – ND ND ND ND 1.04⫾0.16 Reticular ⬎1,000
E120R/R121E ⫹⫹⫹ – 56.5⫾10 Yes ND 15 0.76⫾0.15 Reticular ⬎1,000
E120D/R121K ⫹⫹⫹ – 25⫾11 ND 3 ND 1.12⫾0.24 Reticular 400
E120K/R121K ⫹⫹⫹ – ND ND 0 70 0.71⫾0.12 L reticular 110
L132E ⫹⫹⫹ – 63.5⫾7 ND 0 0 0.84⫾0.13 L reticular 120
E102R ⫹⫹⫹ – ND ND ND ND 0.97⫾0.23 M reticular 400
E102R/R121E ⫹⫹⫹ – 81.5⫾26 Yes 0 28 0.86⫾0.04 M reticular 600
E120R ⫹⫹ – ND ND 6 ND 0.45⫾0.12 M reticular ND
E120R/R121K ⫹⫹ – ND ND 0 80* 0.45⫾0.06 L reticular 240
E120K ⫹⫹ – 5⫾5 ND 0 14 0.84⫾0.10 L reticular 300
E102R/E120R/R121E ⫹⫹ – ND ND 0 ND ND L reticular 400
E120D/R121D ⫹⫹ – ND Yes 0 ND 0.85⫾0.06 L reticular 360
E120R/R121D ⫹⫹ – 19⫾11 ND 0 ND 0.72⫾0.10 Reticular 400
E120K/R121E ⫹ – ND Yes 0 5 0.73⫾0.07 L reticular ⬎1,000
E120K/R121D ⫹ – ND Yes 0 0 0.42⫾0.00 Reticular ⬎1,000
E120D/R121E ⫹ – ND ND 0 0 0.55⫾0.06 L reticular ⬎1,000
R121S ⫹ – 49⫾8 Yes ND ND 0.48⫾0.13 Reticular 340
R121Q ⫹ – 55⫾5 Yes 0 ND 0.54⫾0.07 Reticular 300
L118E ⫹ – 100 Yes 0 10 0.4⫾0.03 Reticular 240
W117E ⫹ – 60 Yes 6 0* 0.43⫾0.08 Reticular 300
R121E – – 100 Yes 0 0* 0.43⫾0.05 Reticular ⬎1,000
R121D – – 100 Yes 0 0* 0.55⫾0.06 Reticular 173
a
Restriction was measured by infecting cells expressing the indicated TRIM5␣rhvariants with HIV-1 and N-MLV expressing the GFP protein. After 48 h, the
percentage of GFP-positive cells (infected cells) was determined by flow cytometry. For HIV-1:⫹⫹⫹⫹, 100% restriction;⫹⫹⫹,⬃75% restriction;⫹⫹,⬃50%
restriction;⫹,⬃25% restriction; and⫺,⬍25% or no restriction. For N-MLV:⫹⫹,⬃50% restriction;⫹,⬃25% restriction; and⫺,⬍25% restriction. Experiments were
performed at least three times, and typical results are shown.
b
HIV-1 reverse transcription was measured by real-time PCR 6 h after infection as described in Materials and Methods. The number represents the percentage of
late reverse transcripts observed in cells expressing the indicated TRIM5␣rhvariant relative to the level of late reverse transcripts in control HIV-1-infected cells
transduced with the empty LPCX vector. Experiments were performed at least two times.
c
Dimerization of TRIM5␣rhvariants was measured by the use of the cross-linker EGS as described in Materials and Methods.
d
Each TRIM5␣rhvariant was assayed for higher-order association with wild-type TRIM5␣rh(association with wild type) or with itself (self-association), as described
in Materials and Methods. In the case of association with wild-type TRIM5␣rh, the percentage represents the fraction of the TRIM5␣rhvariant coprecipitated with
wild-type TRIM5␣rhrelative to the amount of wild-type TRIM5␣rhcoprecipitated with itself. For self-association, the percentage represents the fraction of the
TRIM5␣rhvariant coprecipitated with itself relative to the coprecipitation of wild-type TRIM5␣rhwith itself.ⴱ, A high background was observed for the TRIM5␣rh
variant in the control sample without a TRIM5␣target protein. The reported values may be less reliable as a result of this background.
e
Binding to the HIV-1 capsid complexes was determined for each TRIM5␣rhvariant as described in Materials and Methods. Binding is expressed as the amount
of the TRIM5␣rhvariant bound to HIV-1 capsid complexes divided by the amount of bound wild-type TRIM5␣rhat a similar input level. Experiments were repeated
at least three times. Note that, because the binding ratios are calculated at input levels at which some binding of the mutant TRIM5␣rhprotein to the HIV-1 capsid
complexes can be detected, these ratios overestimate the relative capsid-binding affinities of the mutant proteins.
f
The localization of each TRIM5␣rhvariant was determined as described in Materials and Methods. For each TRIM5␣rhvariant, cytoplasmic bodies were counted
for five independent cells and compared to the number in cells expressing wild-type TRIM5␣rh. “L” means fewer cytoplasmic bodies than wild-type TRIM5␣rh; “M”
means more cytoplasmic bodies than the wild type, and “reticular” indicates a diffuse, reticular pattern of staining throughout the cytoplasm.
g
The half-life of each TRIM5␣rhvariant was estimated by pulse-chase experiments. Every experiment was repeated at least two times.
h
ND, not determined.
10744 DIAZ-GRIFFERO ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:8.585.46.544.79.429.2]cluding R121K, exhibited a conditional phenotype, restricting HIV-1 potently but exhibiting no inhibitory activity against the more weakly restricted N-MLV.
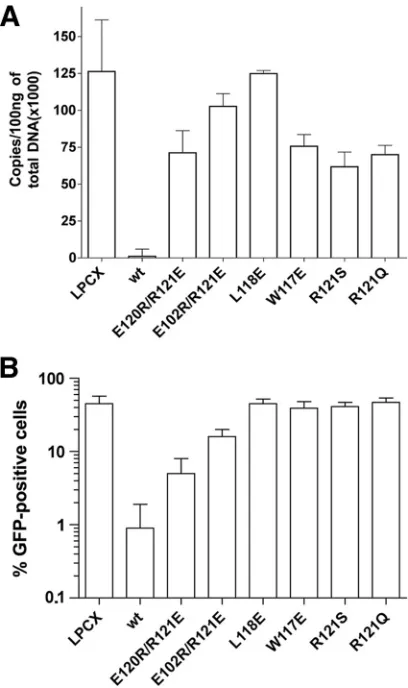
Inhibition of HIV-1 reverse transcription by TRIM5␣rh
mu-tants.Potent HIV-1 restriction by TRIM5␣rhoccurs prior to the initiation of reverse transcription (22, 50). To examine the ability of the TRIM5␣rh B-box 2 mutants to block HIV-1 reverse transcription, we assayed the level of late reverse tran-scripts in mutant-expressing cells challenged with HIV-1-GFP. The percentage of GFP-positive cells was assessed in parallel to provide an indicator of successful infection (Fig. 5). HIV-1 reverse transcripts were not detected in any of the cells ex-pressing potently restricting TRIM5␣rhB-box 2 mutants (Ta-ble 2). In contrast, in cells expressing B-box 2 mutants that did not restrict HIV-1 infection, the levels of HIV-1 reverse tran-scripts were similar to those in control cells not expressing
TRIM5␣rh(Table 2). Intermediate levels of reverse transcripts were found in most cells expressing TRIM5␣rhmutants with modest HIV-1-restricting ability (Table 2). Thus, for this panel of TRIM5␣rhB-box 2 mutants, the levels of HIV-1 late reverse transcripts were lowest when the degree of restriction was most potent.
Effect of TRIM5␣B-box 2 changes on HIV-1 capsid binding.
[image:9.585.45.538.66.481.2]Based on early, qualitative assays measuring the binding of TRIM5␣mutants to HIV-1 capsid complexes, an effector func-tion was proposed for the TRIM5␣ B-box 2 domain (10). Subsequently, when more quantitative capsid-binding assays were utilized, the effects of B-box 2 changes on capsid binding were recognized (32, 33). Because the expression of some TRIM5␣B-box 2 mutants is elevated compared to that of the wild-type protein (9), the input levels of TRIM5␣variants must be adjusted to compare capsid-binding abilities accurately. We
FIG. 4. Restriction of HIV-1 infection by TRIM5␣rh mutants. Cf2Th cells were transduced with the LPCX vector expressing HA-tagged
wild-type and mutant TRIM5␣rhproteins. Control cells were transduced with the empty LPCX vector. Stably expressing cell lines were selected
with 5g of puromycin/ml. The cells were incubated with HIV-1-GFP, and the percentage of GFP-positive cells was measured 48 h later by FACS. The results of two independent experiments were similar; the results of a single experiment are shown.
on November 8, 2019 by guest
http://jvi.asm.org/
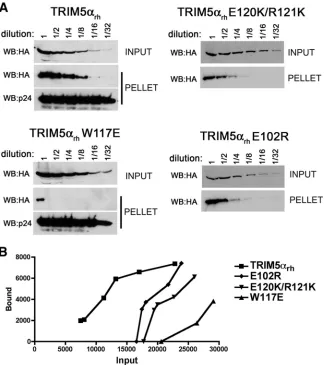
tested the hypothesis that the major function of the TRIM5␣ B-box 2 domain in retrovirus restriction is to increase the avidity of capsid binding. To this end, the ability of the panel of TRIM5␣rhB-box 2 mutants to bind HIV-1 CA-NC complexes assembled in vitro was assessed. The level of each TRIM5␣rh variant in the cell lysate was first measured, and each lysate was diluted with control cell lysates so that comparable levels of TRIM5␣ proteins were present in each input sample. The adjusted cell lysates containing the TRIM5␣rhvariants were then serially diluted and incubated with assembled HIV-1 CA-NC complexes. The mixtures were layered on a 70% su-crose cushion and centrifuged. The amount of TRIM5␣rh pro-tein that cosedimented with the HIV-1 CA-NC complexes was measured. Comparison of the input and bound TRIM5␣rh levels for each mutant with those of the wild-type TRIM5␣rh
protein allowed an estimation of the relative capsid-binding affinity of the mutants (Fig. 6). The relative capsid-binding ability of each mutant was calculated at the lowest input con-centration of the mutant protein that yielded a detectable amount of capsid binding, thus allowing a comparison among the mutants (Fig. 7A and Table 2). A strong correlation (rs⫽
0.8421,P⬍0.001) between binding to HIV-1 capsid complexes and HIV-1 restriction was observed for the panel of TRIM5␣rh
variants (Fig. 7B). These results support the hypothesis that the enhancement of capsid-binding avidity represents a major contribution of the TRIM5␣ B-box 2 domain to retroviral restriction.
Some of the B-box 2 mutants were expressed at higher steady-state levels than the wild-type TRIM5␣rhprotein (see
Fig. S1 in the supplemental material). Although this variable is compensated for by adjusting the input levels of TRIM5␣rhin
the capsid-binding assay, varying expression levels could influ-ence the assessment of HIV-1 restriction potency. Thus, we reanalyzed the data in Fig. 7B, separating the TRIM5␣rh mu-tants into those that were highly expressed and those expressed at wild-type TRIM5␣rhlevels. The strength of the correlation was not improved by taking expression level into consideration (data not shown). Thus, TRIM5␣rhoverexpression only mini-mally affects HIV-1 restriction potency in this context.
Higher-order association of TRIM5␣rh B-box 2 mutants.
The B-box 2 domain has been shown to be important for the ability of TRIM5␣ dimers to associate in higher-order com-plexes (32). To test the ability of the B-box 2 mutants to form higher-order complexes with the wild-type TRIM5␣rhproteins, cell lysates containing FLAG-tagged wild-type TRIM5␣rhand
HA-tagged mutant TRIM5␣rhproteins were mixed. After pre-cipitation with an anti-FLAG antibody, the precipitates were Western blotted with antibodies directed against the FLAG and HA epitope tags. The wild-type TRIM5␣rh-HA protein
was efficiently coprecipitated by the anti-FLAG antibody (Fig. 8), which is consistent with the ability of wild-type TRIM5␣rh
to self-associate in higher-order complexes. Of the TRIM5␣rh mutants tested, only E120D was able to associate efficiently with wild-type TRIM5␣rh(Fig. 8 and Table 2). Apparently, the ability of TRIM5␣rh to form higher-order hetero-oligomers
with the wild-type TRIM5␣rh protein is very sensitive to changes in different surface residues of the B-box 2 domain.
We examined the ability of a subset of TRIM5␣rhmutants to self-associate into higher-order homo- oligomers (Fig. 8D and see Fig. S4 in the supplemental material). TRIM5␣rhmutants that exhibited efficient ability to form higher-order homo-oligomers restricted retrovirus infection with modest or high efficiency (Table 2). Conversely, most of the weakly restricting TRIM5␣ mutants either did not self-associate into higher-order homo-oligomers or did so weakly. Thus, there is a sig-nificant correlation between homologous higher-order self-as-sociation of the TRIM5␣rh variants and HIV-1-restricting ability (rs⫽ 0.862,P⬍ 0.001) (Fig. 8E). This observation is consistent with a model in which homologous higher-order self-association contributes to the HIV-1-restricting ability of TRIM5␣.
Cellular localization of TRIM5␣rhB-box 2 mutants.Changes
[image:10.585.61.265.68.412.2]in the B-box 2 domain can change the cytoplasmic staining of the overexpressed TRIM5␣rhprotein from cytoplasmic bodies to a diffuse, reticular pattern (10). Although a few of the B-box
FIG. 5. Blockade of HIV-1 reverse transcription by the TRIM5␣rh
B-box 2 mutants. Cf2Th cells expressing the indicated wild-type and mutant TRIM5␣rhproteins or containing the empty LPCX vector were
challenged at a multiplicity of infection of 0.4 with DNase-pretreated HIV-1–GFP. After 6 h, the cells were lysed, and the total DNA was extracted. (A) The levels of viral DNA were measured by quantitative real-time PCR, using a probe against GFP, as described in Materials and Methods. The means and standard deviations of the results de-rived from three independent experiments performed in triplicate are shown. (B) In some cases, infections were left to proceed for 48 h, and infectivity was measured as the percentage of GFP-positive cells by FACS. The values shown represent the means and standard deviations of the results obtained in three independent experiments.
10746 DIAZ-GRIFFERO ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
2 mutants localized in well-defined cytoplasmic bodies, most of the mutants exhibited reticular staining patterns (see Fig. S5 in the supplemental material and Table 2). These results support previous studies that suggested that cytoplasmic body forma-tion is not necessary for HIV-1 restricforma-tion by TRIM5␣rh(10,
40, 49).
DISCUSSION
We report here the structure of the TRIM5␣B-box 2 do-main, which reveals its unusual surface properties. We utilize this information to investigate which of the many reported functions of this domain relate to retroviral restriction. The TRIM5␣B-box 2␣fold is similar to that of other B-box 2 and RING-like domains. One notable feature is the surface exposure of hydrophobic residues, which are organized into two clusters, cluster 1 and cluster 2. We provide evidence that the B-box 2 domain contributes to the self-association of
TRIM5␣proteins. A stronger, zinc-mediated interaction con-tributed to the formation of dimers between isolated TRIM5␣ B-box 2 domains. Since the TRIM5␣B-box 2 domain is not required for dimerization (20) and free zinc ion concentrations in the cytoplasm are low, this zinc-mediated interaction is unlikely to contribute to the association of the subunits in the TRIM5␣dimer. All of the TRIM5␣rhB-box 2 mutants studied
herein efficiently dimerized. We identified a second, weaker interaction between the B-box 2 domains involving residues near or within cluster 1. Cluster 1 is likely exposed on the surface of the TRIM5␣dimer, since it apparently contributes to the higher-order self-association of TRIM5␣ proteins. Al-teration of W117, L118 and R121 within or near Cluster 1 resulted in TRIM5␣rhproteins that dimerized, but could not further associate to form higher-order oligomers. Higher-order self-association of the panel of B-box 2 mutants correlated with the ability of TRIM5␣rhto restrict HIV-1 infection. The
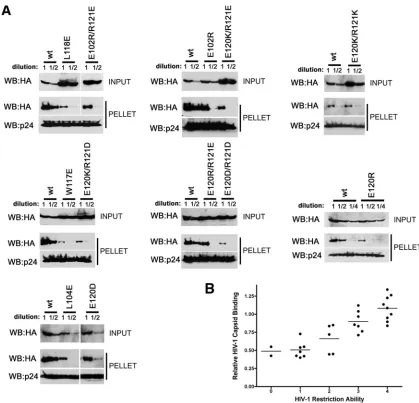
for-FIG. 6. Binding of TRIM5␣rhB-box 2 mutants to assembled HIV-1 capsids. (A) 293T cells were transfected with plasmids expressing the
indicated wild-type and mutant TRIM5␣rhproteins tagged with HA epitopes. At 36 h after transfection, the cells were lysed. Serial dilutions of
the cleared lysates were made by mixing with cleared lysates from untransfected 293T cells. The lysates were incubated at room temperature for 1 h with HIV-1 CA-NC complexes that had been assembled in vitro. The mixtures were applied to a 70% sucrose cushion and centrifuged. Input represents the mixtures analyzed by Western blotting before being applied to the 70% cushion. The input mixtures were Western blotted for the HA tag. The pellet from the 70% cushion (pellet) was analyzed by Western blotting with antibodies against the HA tag and HIV-1 CA-NC protein. (B) The Western blots shown in panel A were quantitated as described in Materials and Methods. The amounts of TRIM5␣rhprotein in the input
and pellet (bound) fractions are shown in arbitrary units.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:11.585.131.457.68.433.2]mation of short-range, higher-order TRIM5␣ arrays might contribute to capsid-binding avidity (Fig. 9). If the long-range geometry of these TRIM5␣arrays is incompatible with that of the retroviral capsid, higher-order self-association could pro-mote the dissolution of the viral capsid during the restriction process (9, 41, 51).
Specific changes in or near cluster 1 were shown to decrease the ability of TRIM5␣rhto bind HIV-1 capsid-like complexes. The TRIM5␣B-box 2 mutants exhibited a wide range of HIV-1-restricting abilities, affording us the opportunity to test the hypothesis that the major contribution of the B-box 2 domain to TRIM5␣ restriction function is mediated through capsid binding. The strong correlation that we observed between cap-sid-binding ability and HIV-1 restriction potency supports this hypothesis. Notably, we did not identify any B-box 2 mutants that bound the HIV-1 capsid complexes efficiently but failed to
restrict HIV-1 infection; in other words, no candidate for a B-box 2 mutant defective in an “effector” function was appar-ent in this sizable mutant panel. Collectively, more than 80% of the TRIM5␣B-box 2 domain surface has been altered (10, 33), increasing the likelihood that the major functions of this domain have already been uncovered.
Coprecipitation assays were used to assess the ability of TRIM5␣rhmutant dimers to associate with wild-type TRIM5␣rh
dimers (higher-order heterologous association) or with them-selves (higher-order homologous association). The higher-order homologous association of TRIM5␣rhmutants correlated with HIV-1-restricting activity better than heterologous association. This is expected, because in the cells tested for restriction activity, only mutant TRIM5␣rhproteins were present. Some of the
[image:12.585.80.499.65.468.2]clus-ter 1 mutants formed higher-order homologous associations bet-ter than hebet-terologous associations, indicating that the introduced
FIG. 7. Binding of additional TRIM5␣rhmutants to assembled HIV-1 capsid complexes. (A) The binding of the indicated TRIM5␣rhvariants
to HIV-1 CA-NC complexes assembled in vitro were assessed as described in the Fig. 6 legend and Materials and Methods. (B) The relative capsid-binding ability and HIV-1-restricting activity of each TRIM5␣rhB-box 2 variant were assessed as described in the Table 2 footnotes and
in Materials and Methods. The means for the relative capsid-binding abilities associated with each HIV-1 restriction group are indicated by horizontal bars. The Spearman rank correlation coefficient,rs, is 0.8421, with a 95% confidence interval of 0.779 to 0.905 (two-sidedPvalue of ⬍0.001).
10748 DIAZ-GRIFFERO ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
changes were self-compatible but disruptive of interactions with the wild-type TRIM5␣rhprotein. This observation supports the importance of the B-box 2 surface proximal to cluster 1 in medi-ating the intermolecular contacts that stabilize the higher-order self-association of TRIM5␣dimers.
The hydrophobic cluster 1 surface of the TRIM5␣rhB-box 2 domain is dominated by the indole ring of W117 (W115 in human TRIM5). W117, along with L118 (L116 in human TRIM5), appears to contribute significantly to the higher-or-der self-association, capsid-binding avidity and HIV-1-restrict-ing ability of TRIM5␣rh. In addition, a basic residue, prefera-bly an arginine residue, flanking this hydrophobic patch contributes to these functions; an arginine at the equivalent position of human TRIM5␣ has previously been shown to contribute to N-MLV restriction (10). Some flexibility with respect to the position of this arginine residue appears to be tolerated in TRIM5␣rh, with arginine at positions 102 or 120
able to compensate for the alteration of R121. The W117, L118, and R121 residues are all conserved in TRIM6 and TRIM34, whose B-box 2 domains can functionally substitute for those of TRIM5 (31). These residues diverge in more distant TRIM proteins such as TRIM21, for example; the
FIG. 8. Higher-order self-association of TRIM5␣rh B-box 2 mutants. 293T cells were transfected with plasmids expressing the indicated
wild-type or mutant TRIM5␣rhproteins with a FLAG or an HA epitope tag. Cells expressing wild-type and mutant TRIM5␣rhproteins were lysed
48 h after transfection. (A to D) The cell lysates were mixed, and the indicated mixtures were used for immunoprecipitation by an antibody directed against the FLAG epitope, as described in Materials and Methods. Elution of the immunocomplexes was performed with a FLAG tripeptide and analyzed by Western blotting with anti-HA and anti-FLAG antibodies. WB, Western blot; IP, immunoprecipitation. (E) The higher-order self-association and HIV-1-restricting ability, determined as described in the Table 2 footnotes and in Materials and Methods, of each TRIM5␣rh
[image:13.585.44.544.66.376.2]variant were compared. The means of the values for higher-order self-association within each restriction group are indicated by horizontal bars. The Spearman rank correlation coefficient,rs, is 0.862, with a 95% confidence interval of 0.665 to 1.0 (two-sidedPvalue of⬍0.001).
FIG. 9. Model for the role of the TRIM5␣ B-box 2 domain in HIV-1 restriction. The TRIM5␣rhdimer is depicted, with the B-box 2
domains (magenta) and B30.2(SPRY) domains (blue) highlighted. In-teractions involving the cluster 1 region of the B-box 2 domain increase the avidity of TRIM5␣rhfor the retroviral capsid, promoting the
ag-gregation of the TRIM5␣rhprotein on the capsid surface.
on November 8, 2019 by guest
http://jvi.asm.org/
RING domain, but not the RING and B-box 2 domains, of TRIM5␣rhcan be functionally replaced by those of TRIM21
(31). Although TRIM6 and TRIM34 exhibit at best marginal ability to restrict retrovirus infection (30, 60), higher-order self-association might contribute to other functions of these proteins. In this respect, it is notable that the dimeric contacts in the crystallized MuRF1 (TRIM63) B-box domain corre-spond to the cluster 1 surface of TRIM5 (37). Thus, many TRIM proteins may utilize the corresponding surface of the B-box domain to modulate proteprotein interactions, in-cluding self-association.
The levels of HIV-1 reverse transcripts detected in the cells expressing the TRIM5␣rhmutants after HIV-1 challenge were lowest when infection was potently restricted. Conversely, in cells expressing the most restriction-defective TRIM5␣rhproteins, the levels of reverse transcripts were comparable to those in control cells lacking TRIM5␣rh. In cells expressing TRIM5␣rhmutants with intermediate levels of restriction potency, a wide range of reverse transcript levels were observed following HIV-1 chal-lenge. These results suggest that TRIM5␣rhrestriction of HIV-1
occurs early in the infection process, precluding reverse transcrip-tion when potent and allowing various degrees of reverse tran-scription when less potent.
Several TRIM5␣rhB-box 2 did not form cytoplasmic bodies
but still exhibited fairly potent restriction of HIV-1 infection. This observation is consistent with previous studies that sug-gested that cytoplasmic body formation by TRIM5␣ is not required for retroviral restriction (40, 49). Our data do not rule out the possibility that the B-box 2-mediated higher-order self-association of TRIM5␣is required for cytoplasmic body for-mation; however, higher-order self-association appears not to be sufficient for cytoplasmic body formation.
As previously observed (10), some changes in the B-box 2 domain can increase the longevity of the relatively short-lived TRIM5␣protein. Our data confirm that rapid turnover is not required for the HIV-1-restricting activity of TRIM5␣; no cor-relation between the half-lives and retrovirus-inhibiting abili-ties of the TRIM5␣rhB-box 2 mutants was apparent. Our data also indicate that cytoplasmic body formation is not necessary for the rapid turnover of TRIM5␣.
The importance of B-box 2-mediated higher-order self-as-sociation and capsid-binding avidity to retroviral restriction should motivate efforts to understand the TRIM5␣and capsid structures that contribute to these processes. Such understand-ing may suggest new interventional approaches to HIV-1 in-fection.
ACKNOWLEDGMENTS
We thank Takashi Umehara for analytical ultracentrifugation and dynamic light scattering measurements and useful discussions and Toshio Nagashima for molecular dynamics calculation of the associa-tion model and useful discussions. We also thank Satoru Watanabe, Takushi Harada, Takeshi Nagir, Yasuko Tomo, Masaomi Ikari, Kazu-haru Hanada, Yukiko Fujikura, and Akiko Tanaka for sample prepa-ration and showing the screening data of B-box 2 domains. We thank Yvette McLaughlin and Elizabeth Carpelan for manuscript prepara-tion.
We acknowledge the National Institutes of Health (grants AI063987 and AI076094 and a Center for AIDS Research Award AI60354), the International AIDS Vaccine Initiative, the Bristol-Myers Squibb Foun-dation, and the late William F. McCarty-Cooper for research funding. F.D.-G. is a recipient of a K99/R00 Pathway to Independence Award
from the National Institutes of Health (1K99MH086162-01), an Amer-ican Foundation for AIDS Research Mathilde Krim fellowship in basic biomedical research (106987-43-RFHF), and a Claudia Adams Barr award from the Dana-Farber Cancer Institute. The B-box 2 structure determination was supported by the Riken Structural Genomics/Pro-teomics Initiative of the National Project on Protein Structural and Functional Analyses, Ministry of Education, Culture, Sports, Science, and Technology of Japan.
REFERENCES
1.Ayed, A., F. A. Mulder, G. S. Yi, Y. Lu, L. E. Kay, and C. H. Arrowsmith.
2001. Latent and active p53 are identical in conformation. Nat. Struct. Biol.
8:756–760.
1a.Best, S., P. Le Tissier, G. Towers, and J. P. Stoye.1996. Positional cloning of
the mouse retrovirus restriction gene Fv1. Nature382:826–829.
2.Borden, K. L.2000. RING domains: master builders of molecular scaffolds?
J. Mol. Biol.295:1103–1112.
3.Borden, K. L.1998. RING fingers and B-boxes: zinc-binding protein-protein
interaction domains. Biochem. Cell Biol.76:351–358.
4.Brennan, G., Y. Kozyrev, and S. L. Hu.2008. TRIMCyp expression in Old
World primates Macaca nemestrinaandMacaca fascicularis. Proc. Natl.
Acad. Sci. USA105:3569–3574.
5.Brzovic, P. S., P. Rajagopal, D. W. Hoyt, M. C. King, and R. E. Klevit.2001. Structure of a BRCA1-BARD1 heterodimeric RING-RING complex. Nat.
Struct. Biol.8:833–837.
6.Cornilescu, G., F. Delaglio, and A. Bax.1999. Protein backbone angle re-straints from searching a database for chemical shift and sequence
homol-ogy. J. Biomol. NMR13:289–302.
7.Delaglio, F., S. Grzesiek, G. W. Vuister, G. Zhu, J. Pfeifer, and A. Bax.1995. NMRPipe: a multidimensional spectral processing system based on UNIX
pipes. J. Biomol. NMR6:277–293.
8.Diaz-Griffero, F., S. A. Hoschander, and J. Brojatsch.2002. Endocytosis is a
critical step in entry of subgroup B avian leukosis viruses. J. Virol.76:12866–
12876.
9.Diaz-Griffero, F., A. Kar, M. Lee, M. Stremlau, E. Poeschla, and J. Sodroski.
2007. Comparative requirements for the restriction of retrovirus infection by
TRIM5␣and TRIMCyp. Virology369:400–410.
10.Diaz-Griffero, F., A. Kar, M. Perron, S. H. Xiang, H. Javanbakht, X. Li, and J. Sodroski.2007. Modulation of retroviral restriction and proteasome
in-hibitor-resistant turnover by changes in the TRIM5␣B-box 2 domain. J.
Vi-rol.81:10362–10378.
11.Diaz-Griffero, F., X. Li, H. Javanbakht, B. Song, S. Welikala, M. Stremlau, and J. Sodroski.2006. Rapid turnover and polyubiquitylation of the
retro-viral restriction factor TRIM5. Virology349:300–315.
12.Diaz-Griffero, F., N. Vandegraaff, Y. Li, K. McGee-Estrada, M. Stremlau, S. Welikala, Z. Si, A. Engelman, and J. Sodroski.2006. Requirements for capsid-binding and an effector function in TRIMCyp-mediated restriction of
HIV-1. Virology351:404–419.
13.Ganser, B. K., S. Li, V. Y. Klishko, J. T. Finch, and W. I. Sundquist.1999.
Assembly and analysis of conical models for the HIV-1 core. Science283:
80–83.
14.Ganser-Pornillos, B. K., U. K. von Schwedler, K. M. Stray, C. Aiken, and W. I. Sundquist.2004. Assembly properties of the human immunodeficiency
virus type 1 CA protein. J. Virol.78:2545–2552.
15.Guntert, P.2009. Automated structure determination from NMR spectra.
Eur. Biophys. J.38:129–143.
16.Hatziioannou, T., D. Perez-Caballero, A. Yang, S. Cowan, and P. D. Bien-iasz.2004. Retrovirus resistance factors Ref1 and Lv1 are species-specific
variants of TRIM5␣. Proc. Natl. Acad. Sci. USA101:10774–10779.
17.Hennig, J., A. Bresell, M. Sandberg, K. D. Hennig, M. Wahren-Herlenius, B. Persson, and M. Sunnerhagen. 2008. The fellowship of the RING: the RING-B-box linker region interacts with the RING in TRIM21/Ro52, con-tains a native autoantigenic epitope in Sjogren syndrome, and is an integral
and conserved region in TRIM proteins. J. Mol. Biol.377:431–449.
18.Herrmann, T., P. Guntert, and K. Wuthrich.2002. Protein NMR structure determination with automated NOE assignment using the new software CANDID and the torsion angle dynamics algorithm DYANA. J. Mol. Biol.
319:209–227.
19.Javanbakht, H., F. Diaz-Griffero, M. Stremlau, Z. Si, and J. Sodroski.2005. The contribution of RING and B-box 2 domains to retroviral restriction
mediated by monkey TRIM5␣. J. Biol. Chem.280:26933–26940.
20.Javanbakht, H., W. Yuan, D. F. Yeung, B. Song, F. Diaz-Griffero, Y. Li, X. Li, M. Stremlau, and J. Sodroski.2006. Characterization of TRIM5␣ trimer-ization and its contribution to human immunodeficiency virus capsid binding.
Virology353:234–246.
21.Johnson, B. A.2004. Using NMRView to visualize and analyze the NMR
spectra of macromolecules. Methods Mol. Biol.278:313–352.
22.Keckesova, Z., L. M. Ylinen, and G. J. Towers.2004. The human and African
green monkey TRIM5␣genes encode Ref1 and Lv1 retroviral restriction
factor activities. Proc. Natl. Acad. Sci. USA101:10780–10785.
23.Kentsis, A., and K. L. Borden.2004. Physical mechanisms and biological
10750 DIAZ-GRIFFERO ET AL. J. VIROL.