0022-538X/96/$04.0010
Copyrightq1996, American Society for Microbiology
Plasmid-Like Replicative Intermediates of the Epstein-Barr
Virus Lytic Origin of DNA Replication
ROGER PFU¨ LLERANDWOLFGANG HAMMERSCHMIDT* Institut fu¨r Klinische Molekularbiologie und Tumorgenetik, GSF-Forschungszentrum
fu¨r Umwelt und Gesundheit GmbH, D-81377 Munich, Germany
Received 4 August 1995/Accepted 10 March 1996
During the lytic phase of herpesviruses, intermediates of viral DNA replication are found as large concate-meric molecules in the infected cells. It is not known, however, what the early events in viral DNA replication that yield these concatemers are. In an attempt to identify these early steps of DNA replication, replicative intermediates derived from the lytic origin of Epstein-Barr virus,oriLyt, were analyzed. As shown by density shift experiments with bromodeoxyuridine,oriLytreplicated semiconservatively soon after induction of the lytic cycle andoriLyt-containing DNA is amplified to yield monomeric plasmid progeny DNA (besides multimeric forms and high-molecular-weight DNA). A new class of plasmid progeny DNA which have far fewer negative supercoils than do plasmids extracted from uninduced cells is present only in cells undergoing the lytic cycle of Epstein-Barr virus. This finding is consistent with plasmid DNAs having fewer nucleosomes before extrac-tion. The newly replicated plasmid DNAs are dependent on a functionaloriLytincisand support an efficient marker transfer into Escherichia coli as monomeric plasmids. Multimeric forms of presumably circular progeny DNA oforiLyt, as well as detected recombination events, indicate thatoriLyt-mediated DNA replication is biphasic: an early theta-like mode is followed by a complex pattern which could result from rolling-circle DNA replication.
Epstein-Barr virus (EBV), a human herpesvirus, is able to infect human B lymphocytes in vitro and in vivo. These target cells for EBV become latently infected with the virus, and its genetic information is maintained in the dividing cell popula-tion as extrachromosomal multiple copies (see reference 31 for a review). Thecis-acting element which mediates DNA repli-cation during the latent phase of the viral life cycle has been identified as the plasmid origin of DNA replication,oriP, to-gether with its viral transactivator, EBNA1 (35, 48). In the latently infected cells, only a fraction of the more than 80 viral genes are found expressed and no virus is produced. In con-trast, during the lytic phase of the viral life cycle the viral DNA is amplified several hundredfold via a different origin of DNA replication,oriLyt(20). The lytic phase is characterized by the sequential activation of viral gene expression, which is initiated by the transcriptional activation of a single viral gene, the BZLF1 gene (9, 45).
During lytic-cycle replication, herpesviruses have to over-come the cellular barrier of constraints on DNA replication. Since lytic herpesvirus replication results in a burst-like ampli-fication of DNA, all herpesviruses provide replication proteins that specifically interact with the lytic origins of DNA replica-tion and sustain the key funcreplica-tions of DNA replicareplica-tion, includ-ing polymerases, helicases, primases, DNA-bindinclud-ing proteins, and associated factors, as well as enzymatic activities involved in the biochemical pathways of nucleotide synthesis and phos-phorylation (6, 14, 46). Although thecis-acting origins of DNA replication of different members of the herpesvirus family have been identified and the trans-acting viral factors have been genetically and biochemically dissected, the mechanism which
underlies lytic herpesvirus replication has not been investi-gated thoroughly to our knowledge. To address this question from a mechanistic point of view, we analyzed the mode with which the EBV lytic origin of DNA replication replicates.
The structure of the EBV lytic origin of DNA replication is complex. oriLyt consists of several essential and auxiliary re-gions which are required for its full activity (19, 20). One essential component colocalizes with a promoter which is di-rectly transactivated by the viral gene product BZLF1. This protein is related to the AP1 family of transcription factors (12), is a sequence-specific DNA-binding protein (12, 30), and is directly involved in oriLyt-mediated DNA replication (38, 39). The second essential component oforiLyt(40) could con-tain the origin recognition element of EBV (10) to which several cellular proteins, including Sp1, bind specifically (18). In contrast tooriLyt of EBV, the members of the alphaher-pesvirus family have less complicated origins which neverthe-less need a viral DNA-binding protein for activation (see ref-erence 7 for a review).
The intermediate product of lytic DNA replication of all herpesviruses is a large concatemeric molecule in which the single genome units are arranged head to tail (23), an arrange-ment which has also been demonstrated for EBV (20, 21). This finding is consistent with the idea that a circularized mono-meric DNA molecule which is either formed upon infection of a cell through circularization of linear virion DNA or provided by the plasmid genomes present in latently infected cells serves as a template for DNA replication (36). For such a model, it is conceivable that the concatemers are formed by a rolling-circle mechanism analogous to that used by several bacteriophages. Such a model would account poorly, however, for the rapid accumulation of concatemeric molecules. In herpes simplex virus type 1-infected cells, 20 to 40 min is needed to complete the replication of one viral molecule but the herpesviral DNA is replicated several hundredfold in several hours to yield up to 1,000 genomic copies per cell (24). A pure rolling-circle
mech-* Corresponding author. Mailing address: Institut fu¨r Klinische Molekularbiologie und Tumorgenetik, GSF-Forschungszentrum fu¨r Umwelt und Gesundheit GmbH, Marchioninistr. 25, D-81377 Mu¨nchen, Germany. Phone: 49/89/7099-506. Fax: 49/89/7099-500. Electronic mail address: [email protected].
3423
on November 9, 2019 by guest
http://jvi.asm.org/
anism of DNA replication produces progeny DNA linearly with time, not exponentially, and appears inadequate to ex-plain the observed amplification of viral DNA. Alternatively, an initial phase of semiconservative plasmid replication of the primary circular DNA template via theta-like or Cairns struc-tures would provide many circular progeny molecules which could all serve as templates for DNA synthesis via a subse-quent rolling-circle mode of DNA replication. We have tested the validity of such a biphasic model for the EBV lytic origin of DNA replication and found evidence for the amplification of early replicative DNA intermediates which are plasmid-like.
MATERIALS AND METHODS
Cell lines.D98HR1 cells were derived from a somatic cell hybrid composed of the EBV genome-positive Burkitt’s lymphoma cell line P3HR1 and the human epithelial cell line D98 (16). This cell line was maintained in Dulbecco’s modified Eagle’s medium containing 5% fetal and 5% newborn calf sera. The Akata Burkitt’s lymphoma cell line (44) was grown in RPMI 1640 medium supple-mented with 15% fetal calf serum. The lytic cycle of EBV was induced in Akata
cells at a density of 43106/ml with an anti-human immunoglobulin G (IgG)
antibody at 0.1 mg of immunoglobulin derived from goats (Cappel) per ml as described elsewhere (44).
Bromodeoxyuridine labeling and isolation of replicative intermediates from stably transfected Akata cells.Akata cells were electroporated with the 12.6-kb plasmid p562 (20), which carries both the plasmid and lytic origins of replication,
oriPandoriLyt, and the selectable marker gene encoding hygromycin phospho-transferase. The transfected cells were selected for maintenance of p562 in RPMI 1640 medium supplemented with 15% fetal calf serum in the presence of
200mg of hygromycin per ml. The selected cell line was kept at a density of about
106
cells per ml, and the cells were induced to undergo the lytic cycle by an anti-human IgG antibody as described above. For labeling experiments with the base analog bromodeoxyuridine, bromodeoxyuridine (final concentration, 60 mM) and deoxycytidine (final concentration, 0.2 mM) (33) were added to the medium 2 h after induction of the lytic cycle and total cell DNA was prepared after various time periods, as described in Results. For control experiments with uninduced cells, the cells were grown under the same conditions except that the
incubation with the antibody was omitted. Total DNA was prepared from 108
cells, and 100mg of the isolated DNA was separated on neutral CsCl
buoyant-density gradients with an initial buoyant-density of 1.75 g/ml at 38,000 rpm for 72 h in a Beckman 70.1 TI rotor. The gradients were fractionated into 30 samples. Defined portions of the fractions were slot blotted onto nylon membranes which were
hybridized to [32
P]dCTP-random-primed pUC DNA to detect p562 DNA only. The slot blot autoradiograms were quantified by liquid scintillation counting. DNAs from certain fractions were subsequently analyzed on agarose gels. The p562 DNA was detected after Southern blot transfer of the agarose gels onto
nylon membranes (Zetaprobe; Bio-Rad) by hybridization to [32P]dCTP-labeled
pUC plasmid DNA, which detected the prokaryotic plasmid backbone of p562 only.
Two-dimensional agarose gel electrophoresis.Total cellular DNA was pre-pared from cells which had been washed once in TEN (50 mM Tris-HCl [pH 7.6],
20 mM EDTA, 100 mM NaCl) and resuspended in TEN at 23105cells per ml.
Cells were immediately lysed by adding sodium dodecyl sulfate (SDS) (final
concentration, 0.1 g/ml) and incubation with proteinase K (0.5 mg/ml) at 508C for
2 h. After addition of RNase I (1mg/ml) for 5 min, the DNA was phenol
ex-tracted, precipitated, and resuspended in Tris-EDTA (TE) buffer according to standard procedures (37). Total cellular or gradient-fractionated DNA was
pre-pared, and samples were loaded on 0.5% agarose gels in 13TAE buffer (40 mM
Tris-acetate [pH 8.0], 1 mM EDTA). After electrophoresis at 1.2 V/cm for at least 20 h in the first dimension (from left to right in the figures), the gel slabs
were incubated in 13TAE containing 1.5 or 50mg of chloroquine per ml, turned
at an angle of 908, and embedded in 0.5% agarose in the same buffer and then
second-dimension electrophoresis (from top to bottom in the figures) was per-formed at 1.6 V/cm for at least 24 h. Electrophoretic runs in the presence of chloroquine were performed in the dark to prevent photodamaging of DNA. The
oriLyt-containing DNAs were detected as described above.
Transient replication assays and plasmid rescue experiments.The plasmid p526, which carries the lytic origin of DNA replication of EBV only and lacks
oriP, contains aBamHI-SalI fragment with the completeoriLytelement
(nucle-otide coordinates 48,848 to 56,084) (2) from the EBV strain B95-8 and is
described in detail elsewhere (20, 40). TheoriLytmutant plasmid p1026 carries
a nonfunctionaloriLytwith a 10-bp substitution which was introduced by
oligo-nucleotide-directed mutagenesis as described previously (40). Transient
replica-tion assays were performed with D98HR1 cells by cotransfecting 10mg each of
theoriLytwild-type plasmid p526, the mutantoriLytplasmid p1026, and the plasmid pCMV-BZLF1. The latter plasmid is an expression vector which effi-ciently induces the lytic cycle of EBV (20). The BZLF1-encoding gene is driven in this retroviral vector construct by the promoter of the immediate-early genes of the human cytomegalovirus. The transfection experiments were performed by
electroporation as described elsewhere (40). At various time points after
trans-fection, DNA was prepared (22) and digested withDpnI andXbaI to cleave the
input plasmid DNAs and cellular DNAs, respectively. Five-microgram amounts
of the DNA samples were transfected subsequently intoEscherichia coli
Epicu-rian SCS1 [recA1 endA1 gyrA96 thi-1 hsdR17[rk2mk1)supE44 relA1], and the
numbers of colonies which grew on ampicillin plates at 378C were counted. As a
control, theoriPplasmid p152.22 (pHEBo) (43) was used. To meet the same
conditions as used with the three plasmids p526, p1026, and pCMV-BZLF1,
20mg of pBluescript SKII(2) DNA was cotransfected with 10mg of p152.22 into
D98HR1 cells and 5mg of Hirt-extracted DNA was transfected intoE. coli.
Propidium diiodide CsCl gradients.Low-molecular-weight DNA from Akata
cells stably transfected with theoriP-oriLytplasmid p562 was prepared by the
Hirt protocol (22), phenol extracted, precipitated, and resuspended in a CsCl solution with a density of 1.54 g/ml containing 0.5 mg of propidium diiodide (Sigma) per ml. The different forms of topoisomeric DNAs were separated in a Beckman 70.1 TI rotor at 35,000 rpm at 168C for 72 h. The gradients were fractionated, the dye was removed by several rounds of extraction with CsCl-saturated isopropanol, and the DNA was precipitated with ethanol and resolved in standard TE buffer (37).
Topoisomerase assays.Purified topoisomerase I from calf thymus (Gibco/ BRL) or wheat germ topoisomerase (Promega) was used in buffers as recom-mended by the manufacturers.
RESULTS
oriLytreplicates semiconservatively soon after induction of the lytic cycle.The lytic mode of DNA replication of herpes-viruses ensures massive DNA replication. It is known that during the lytic phase of herpesviruses, intermediates of DNA replication are found as large concatemeric molecules, but what the early events are that yield these concatemers is un-known. The size of herpesvirus DNA genomes (.100 kbp) makes it difficult to analyze initiation, elongation, and replica-tive intermediates directly during lytic DNA replication, even with pulsed-field gel electrophoresis techniques (4, 49). To overcome this technical problem and to identify the mode of DNA replication of EBV early after onset of lytic DNA rep-lication, we used the vector plasmid p562, which combines both the lytic and latent origins of EBV DNA replication,oriPand
oriLyt. This 12.6-kbp plasmid replicates extrachromosomally via oriP in latently EBV-infected cells but can also replicate throughoriLytafter induction of the lytic phase of the endog-enous virus (20). Therefore, theoriP-oriLytvector serves as a mini replicator model to reflect viral DNA replication in both phases of EBV’s life cycle. Since the oriP-oriLytvector p562 also carries the gene for hygromycin phosphotransferase, the plasmid was stably introduced into the EBV-positive Burkitt’s lymphoma cell line Akata, where the plasmid replicates extra-chromosomally viaoriPduring the latent phase of EBV. Akata cells are also known to undergo the lytic phase of EBV’s life cycle upon cell surface IgG cross-linking with IgG anti-bodies (44). Induction of the lytic cycle is fast and DNA am-plification viaoriLytstarts as early as 4 h after induction (data not shown), although not all Akata cells support the lytic phase of EBV. The viral replication factors needed fororiLyt -medi-ated DNA replication are supplied intransby the endogenous EBV (13, 14, 38). DNAs isolated before and after induction of the lytic cycle of EBV in these cells carrying the 12.6-kbp
oriP-oriLytvector p562 have been analyzed by different tech-niques.
First, in a classical Meselson-Stahl experiment (32) we ana-lyzed the incorporation of the base analog bromodeoxyuridine into p562 plasmid DNA in Akata cells in which the vector replicates extrachromosomally in synchrony with the cellular DNA in a theta-like mode viaoriPduring the latent phase (1, 15, 47). Uninduced, asynchronously proliferating Akata cells were cultivated in the presence of the base analog bromode-oxyuridine (final concentration, 60mM) and deoxycytidine (fi-nal concentration, 0.2 mM) (33) for 8 h; total cellular DNA was prepared and separated in neutral CsCl density gradients; and
3424 PFU¨ LLER AND HAMMERSCHMIDT J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
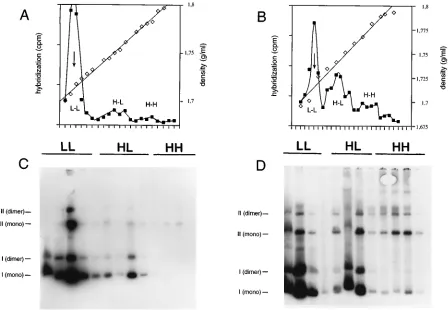
the p562 plasmid DNA was detected by slot blot hybridization after fractionation of the gradient as described in detail in Materials and Methods. After a labeling period of 8 h, approx-imately 10% of theoriP-oriLytvector had replicated semicon-servatively in uninduced cells, as indicated by the detection of heavy-light (H-L) DNA (Fig. 1A), whereas most of the p562 DNA had not incorporated bromodeoxyuridine and localized at the position of light-light (L-L) DNA (Fig. 1A). Fractions of the gradient were also analyzed on agarose gels by Southern blot hybridization (Fig. 1C). Individual bands which were vi-sualized in the autoradiogram indicate different species of mo-nomeric form I and form II [I (mono) and II (mono)] plasmid DNA and probably a dimeric version of theoriP-oriLytplasmid [I (dimer) and II (dimer)] replicating in the Akata cells.
The experiment was repeated, but the lytic phase of EBV’s life cycle was now induced and 2 h later bromodeoxyuridine was added to the medium. After induction of the lytic cycle in Akata cells, a marked proportion (about 40%) of p562 DNA banded at the position of H-L DNA after an 8-h bromode-oxyuridine labeling period and a smaller proportion (about 10%) in which both DNA strands were substituted by bro-modeoxyuridine could be found (Fig. 1B). Selected fractions of the gradients were analyzed on one-dimensional agarose gels,
and Southern blot hybridizations detected predominantly su-percoiled plasmid DNAs of monomeric and presumably dimeric natures (Fig. 1D) which were identical to the signals detected in uninduced cells (Fig. 1C). This was unexpected for
oriLyt-mediated DNA replication since the replicative inter-mediates of the proposed rolling-circle model would not nec-essarily be plasmid-like but are expected to consist of concate-mers.
Both concatemers and Cairns circles, however, would give rise to a small fraction of DNA substituted on one strand in the first round and mixed H-H and H-L DNA species in subse-quent rounds of DNA replication. Noncircular, high-molecu-lar-weight DNA could mostly be found as a smear in the upper part of the gel in the H-L and H-H fractions in the induced cells (Fig. 1D), whereas such signals were lacking in cells in which p562 replicates viaoriPonly (Fig. 1C). The amount of this high-molecular-weight DNA relative to that of plasmid form DNA is small (see also Fig. 2 and 3 and reference 20), and H-H DNA fractions contained very little form I DNA of plasmid nature, unlike the situation with the L-L and H-L DNA fractions, which contained an excess of form I DNA (Fig. 1D). These observations are most likely due to inhibitory ef-fects of bromodeoxyuridine on oriLyt-mediated DNA
repli-FIG. 1.oriLytreplicates semiconservatively soon after induction of the lytic cycle. The EBV-positive Burkitt’s lymphoma cell line Akata was stably transfected with
theoriP-oriLytplasmid p562. Total cellular DNA from noninduced (A and C) or lytically induced (B and D) Akata cells was separated on neutral cesium chloride density gradients, and the plasmid DNA was detected by slot blot hybridization after fractionation. (A) After an 8-h period of labeling with bromodeoxyuridine,
approximately 10% of thisoriP-oriLytvector has replicated semiconservatively in uninduced cells, as indicated by the detection of H-L DNA. Most of the plasmid DNA
has not yet replicated and is detected at the position of L-L DNA. (B) After induction of the lytic cycle for 10 h, approximately 30% of p526 is found in the H-L fraction and a minor proportion of fully bromodeoxyuridine-substituted p562 DNA is found at the expected position of H-H DNA. (C and D) Most of the p562 DNA in noninduced (C) and induced (D) cells is present as different mono- and multimeric forms of circular plasmid DNA, as revealed by Southern blot hybridization of agarose
gels on which selected fractions from the cesium chloride buoyant-density gradients were analyzed. (A and B)h, cesium chloride density of individual fractions;■,
liquid scintillation counting of individual fractions after slot blot hybridization. The arrows point to the calculated density of native p562 DNA (1.717 g/ml), based on a GC content of 57.6%. (C and D) Bars labeled I (mono), II (mono), I (dimer), and II (dimer) indicate form I and II DNAs of monomeric and presumably dimeric p562 plasmid form DNAs.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.82.530.70.380.2]cation. Bromodeoxyuridine pulses at later time points after induction revealed a shift to high-molecular-weight DNA products labeled on both strands, consistent with a shift to a rolling-circle mechanism (data not shown).
Despite such limitations of the bromodeoxyuridine labeling experiments, our results indicated that circular plasmid-like progeny DNA can easily be detected. Its detection is depen-dent on a functionaloriLyt element on theoriP-oriLytvector (data not shown) and does not arise from enhancedoriP rep-lication during the lytic phase of EBV DNA reprep-lication (see below). It thus appeared thatoriLyt-mediated DNA replication can give rise to plasmid-like DNA as a replicative intermedi-ate. The finding of this kind of replication, which is nearly indistinguishable from semiconservativeoriP-mediated DNA replication (47), could also be the result of an artifact due to bromodeoxyuridine toxicity and needed independent verifica-tion.
oriLytgives rise to progeny plasmid DNA in transient rep-lication experiments. To verify the findings of the bromode-oxyuridine labeling experiments, we used a transient replica-tion assay which allowed the analysis of oriLyt-derived replication intermediates in the absence of inhibitory base an-alog and with anoriLytplasmid lacking the plasmid origin of replication, oriP. DNA replication of oriLyt-containing plas-mids isolated fromE. coliin EBV-positive cells can be easily monitored in a transient replication assay (20). The basis for this assay is the presence or absence of E. coli-borne dam -specific replication in one or both strands of DNA isolated from transfected cells. Plasmid DNA from E. coli becomes resistant to cleavage by the restriction enzyme DpnI after 1 round of replication in mammalian cells. We have previously used this transient replication assay to define theoriLyt ele-ments (18, 20, 39, 40). Were plasmids true replicative interme-diates duringoriLyt-mediated replication, rescue experiments withE. colishould be a biological means to detect such plas-mid-like intermediates, sinceE. colican be transformed easily only with circular plasmid DNA.
The latently EBV-infected D98HR1 cell line was used in the transient replication assays (16). This cell line was cotrans-fected with the 8.4-kbporiLytplasmid p526 together with the expression vector pCMV-BZLF1 and a null mutant oforiLyt
(p1026) (40). pCMV-BZLF1 efficiently induces the lytic cycle in D98HR1 cells (20), and the nonreplicatingoriLytplasmid p1026 served as an internal control for completeDpnI diges-tion of input DNA which had not undergone replicadiges-tion in D98HR1 cells. For comparison, the 7.1-kbp p155.22 plasmid (43), which encompasses the plasmid origin of DNA of repli-cation,oriP, only, was cotransfected with pBluescript plasmid DNA (in order to transfect the same amount of DNA) or the pCMV-BZLF1 plasmid into D98HR1 cells. Immediately after DNA transfection into D98HR1 cells and after defined time intervals, low-molecular-weight DNA was prepared and di-gested withDpnI to cleave multiply input DNA from E. coli
and XbaI to cleave cellular DNA in order to reduce the vis-cosity of the DNA preparation (XbaI does not cleave in p526 or p1026 DNA). An E. colistrain was transformed with the DNA samples, and ampicillin-resistant colonies were counted at various time intervals after transfection. The data are sum-marized in Table 1. Although the kinetics of induction of the lytic cycle in D98HR1 cells are much slower than in Akata cells, ampicillin-resistant colonies could be detected 8 h after transfection and an nearly exponential increase in colony num-bers could be observed. These colony numnum-bers correlated di-rectly with the increase in the amount ofDpnI-resistant p526 DNA seen in Southern blot hybridization experiments (Fig. 2 and data not shown). No colonies could be obtained
immedi-ately after DNA transfection (Table 1), which indicates that
DpnI cleavesE. coliinput DNA completely in this very sensi-tive biological assay. Analysis of about 50 colonies for the type of the rescued DNAs indicated that all colonies contained monomeric p526 DNA. In no case could theoriLytnull-mutant plasmid p1026 be detected (it can be distinguished from p526 by its restriction pattern). TheoriP-carrying plasmid, p155.22, replicated much more slowly than theoriLytplasmid did and could only be consistently detected 48 h after transfection (Table 1). Cotransfection of theoriPplasmid, p155.22, with the expression plasmid pCMV-BZLF1 did not alter the number of ampicillin-resistantE. colicolonies (data not shown).
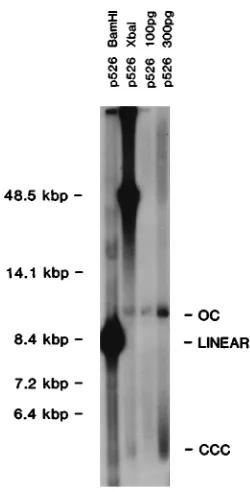
[image:4.612.314.554.92.226.2]In an attempt to directly show the fraction of molecules participating inoriLyt-mediated DNA replication, we analyzed newly replicated p526 DNA by Southern blot hybridization. After 24 h, total cellular DNA was extracted from D98HR1 cells cotransfected with pCMV-BZLF1 and the 8.4-kbp plas-mid p526, which carriesoriLytonly. The DNA preparation was digested first with DpnI and subsequently with BamHI, to linearize p526 DNA, orXbaI, which cleaves only cellular DNA. As a control for the positions of form I and form II DNAs, uncut p526 DNA (100 and 300 pg) isolated fromE. coliwas loaded on the same agarose gel together with the DNA sam-ples. After Southern blot hybridization with a radioactively labeled prokaryotic probe, most of the DpnI-resistant p526 DNA was of high molecular weight and migrated above the position of linear lphage DNA (Fig. 2). In addition, form I (covalently closed circular) (CCC) and form II (open circular) (OC) DNAs which migrated at the same positions as p526 plasmid DNA isolated directly from E. coliwere clearly de-tectable (Fig. 2). The supercoiled form I DNA (CCC) (Fig. 2) is displayed as a smear on the gel resulting from an artifact that is seen when very small amounts of supercoiled plasmids are analyzed on ethidium bromide-containing gels (data not shown). The amount of plasmid-like p526 DNA was in the low picogram range, which is in the same order as in the plasmid rescue experiments (Table 1). In dramatic contrast to results shown in Fig. 1, however, high-molecular-weight DNA was prevalent in the experiment shown in Fig. 2, indicating that bromodeoxyuridine artifactually impaired the generation of such DNA duringoriLyt-mediated DNA replication of EBV.
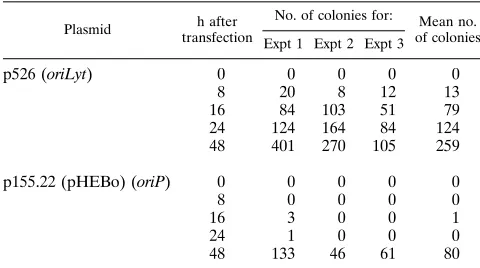
TABLE 1. Plasmid rescue in transient replication assays withoriLytandoriPplasmids in D98HR1 cellsa
Plasmid h after
transfection
No. of colonies for: Mean no.
of colonies Expt 1 Expt 2 Expt 3
p526 (oriLyt) 0 0 0 0 0
8 20 8 12 13
16 84 103 51 79
24 124 164 84 124
48 401 270 105 259
p155.22 (pHEBo) (oriP) 0 0 0 0 0
8 0 0 0 0
16 3 0 0 1
24 1 0 0 0
48 133 46 61 80
aCellular DNA was isolated and digested with a combination ofXbaI and
DpnI to restrict cellular DNA and unreplicated input plasmid DNA, respectively.
The test plasmids do not containXbaI sites, and newly replicated DNA is not cut
byDpnI since the DNA lacks theE. coli dam-specific adenosine methylation
required forDpnI cleavage. The DNA samples were transfected intoE. coli
Epicurian SCS1, and ampicillin-resistant colonies were counted. In reconstruc-tion experiments, cellular DNA samples were doped with defined amounts of pUC18 DNA and the transfection efficiency was calculated to be about 10
colonies per 10212
g of plasmid DNA inE. coliEpicurian SCS1.
3426 PFU¨ LLER AND HAMMERSCHMIDT J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
TheoriLytproducts obtained in transient replication assays are in complete agreement with results of our bromodeoxyuri-dine labeling experiments and indicate that DNA replication via oriLytgives rise to plasmid-like progeny DNA intermedi-ates which resemble Cairns circles.
oriLyt-mediated DNA replication yields plasmid-like iso-mers.InoriLytreplication assays we regularly observed signals, namely smears and bands, which could not easily be explained. Some of the signals are faintly visible in Fig. 2, and we also observed them in our previous work (20). We wondered if these additional signals might arise from replicative interme-diates which stem from transitional DNA forms to yield either plasmid-like progeny DNAs or higher-order DNA complexes. Since the resolution of one-dimensional gels is not sufficient to analyze complicated structures, we performed two-dimen-sional agarose gel electrophoresis in which the first dimension was run in the absence of an intercalating dye to separate DNAs mainly according to molecular weight. The second di-mension (run at a 908 angle) was done in the presence of different concentrations of chloroquine, which intercalates to various extents into different topological forms of double-stranded DNAs and alters their migration. This effect occurs because intercalation changes the hydrodynamic shape of closed circular DNA. Intercalation by chloroquine reduces or removes helical twists in negatively supercoiled DNA and
in-creases the number of positive supercoils in already positively supercoiled DNA. Therefore, negatively supercoiled DNA be-comes less supercoiled at low concentrations of chloroquine, and as the chloroquine concentration is raised DNA becomes more relaxed until increasing intercalation introduces positive supercoils into form I CCC molecules. Relaxed or positively coiled CCC DNA can only be positively supercoiled by inter-calation of chloroquine, such that the intercalator always in-creases the mobility of these DNAs relative to that of nicked, OC DNA.
In an attempt to analyze replicative intermediates from the
oriP-oriLytvector p562, Akata cells which carried this plasmid in a stable extrachromosomal form were used. Total cell DNA was isolated from uninduced cells in which p562 replicates via
oriP and from cells in which the lytic phase of the EBV life cycle had been induced for 10 h. The DNAs were digested with
XbaI, which does not cut within p562 (and does not have a detectable nicking activity) but cleaves cell DNA frequently to reduce the viscosity of the DNA solution, thereby preventing overloading effects on the gels. Twelve-microgram amounts of each DNA preparation were run on two-dimensional agarose gels which contained 50mg of chloroquine per ml only in the second dimension, and p562 DNA was detected after Southern blotting with a probe directed against the prokaryotic plasmid backbone of theoriP-oriLytplasmid.
Highly negatively supercoiled form I DNA (Fig. 3A, black box) which stems from the removal of histones from the depro-teinized DNAs runs as an entity in the first dimension but is separated in the second dimension in the form of a vertical streak due to intercalation of chloroquine (Fig. 3A shows a schematic drawing of the expected migration behavior of form I and II plasmid DNAs). This finding is expected and is the case for p562 DNA in uninduced cells (Fig. 3B) as well as in induced cells (Fig. 3C, supercoiled form I DNA). The signal intensities shown in Fig. 3B and C differ remarkably; much more material can be detected in cells in which lytic DNA replication is induced. Besides high-molecular-weight DNA of p562, which is dominant in induced Akata cells (Fig. 3C, upper left corner), different oligomers of p562 can occasionally be detected, as well as a new class of p562 DNA which is clearly visible as a horizontal bar at the bottom of Fig. 3C (labeled ‘‘T’’). This DNA class, together with the negatively supercoiled DNA in the vertical streak, forms a rectangular pattern, indi-cating that both sides of the rectangle are composed of mono-meric CCC DNA. The new class of p562 DNA is separated into an apparent streak in the first dimension, but the very high concentration of chloroquine introduces positive supercoils and makes this DNA move uniformly and rapidly in the second dimension. The horizontal streak ‘‘T’’ actually consists of a stripe-like arrangement of narrowly spaced vertical bands, as can be seen in the enlarged reproduction shown in Fig. 3D. Similar horizontal streaks are also seen with oligomers of the p562 plasmid in induced cells (Fig. 3C); they were not detect-able in uninduced cells, even in very long exposures of such autoradiograms (data not shown). In contrast to the clearly interpretable second-dimension pattern, smearing between the supercoiled form I DNA and the nicked form II DNA in the autoradiograms in Fig. 3B and C indicates that nicking oc-curred between performance of the first and second dimen-sions and during electrophoresis in the second dimension. These streaks are artifactual.
[image:5.612.113.238.67.313.2]oriLyttopoisomers with fewer negative supercoils.The find-ings reported in the previous section are indicative of different isomers of p562 progeny DNA which are plasmid-like but differ in their respective linking numbers. These DNAs are detectable only after induction of the lytic phase of EBV’s life
FIG. 2. Replication of theoriLytplasmid p526 in transient replication assays
in D98HR1 cells. The autoradiogram shows different forms of p526oriLyt
plas-mid DNA which had been isolated from D98HR1 cells in the transient
replica-tion assay. Total cell DNA was digested withDpnI to multiply cleave input
plasmid DNA and withBamHI to linearize p526 orXbaI to cleave cellular DNA
only. Controls for supercoiled form I DNA (CCC) or OC form II DNA of p526
derived from CsCl-purifiedE. colipreparations were included (100 and 300 pg)
in the electrophoretic analysis.DpnI-XbaI-digested cellular DNA from D98HR1
cells transfected with p526 shows a strong signal above the 14.1-kbp marker fragment, and faint but clearly visible signals were detected at the positions of
OC and supercoiled (CCC) plasmid DNA.DpnI-BamHI-digested DNA from the
same DNA preparation shows a very strong signal at the expected position for linearized p526 plasmid DNA which is 8.5 kbp in size. In addition, faint bands which we and others have consistently observed in longer-exposed autoradio-grams from transient replication assays are visible above and below this signal
(20, 35). We interpret these bands as partiallyDpnI-digested DNA fragments
which could arise from passing replication forks in newly replicated DNAs.
on November 9, 2019 by guest
http://jvi.asm.org/
cycle. The topological state of this DNA class is unusual in that the DNA is separated on a native agarose gel in the absence of intercalating dye but in the presence of high concentrations of chloroquine moves in a very condensed form. Since the cellular DNA was treated with RNase, proteinase K, and SDS and subsequently extracted with phenol, an alternative factor which contributes to the unusual migration behavior of the DNAs is unlikely. It appears that this class of plasmid DNA has an unusual range of linking numbers. Two different types of ex-periments were performed to evaluate this assumption.
Since a change in linking number can be gauged on the basis of the degrees of supercoiling under different conditions (25, 34), we repeated the experiment described above.Total cellular DNA extracted from induced Akata cells carrying the p562 plasmid was electrophoresed in the first dimension in the ab-sence of intercalating dye, but in the second dimension 1.5mg of chloroquine per ml was added (1/33 of the concentration used in the previous experiment). p562 DNA was again de-tected by Southern blot hybridization.
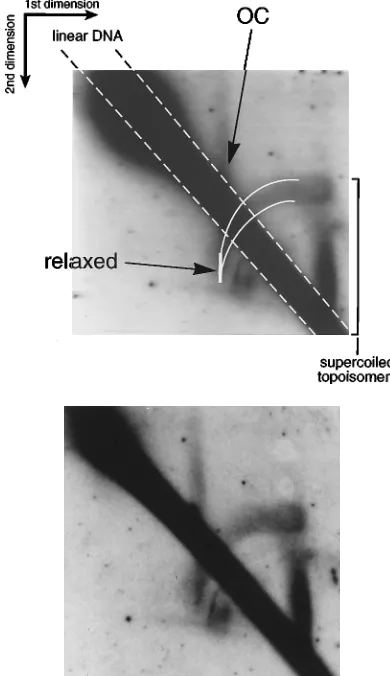
In contrast to the horizontal ‘‘T’’ streak visible in Fig. 3C and D, under these conditions an arc is formed between the posi-tions of the relaxed DNA and the highly negatively supercoiled DNA molecules which migrated fastest in the first dimension. Relaxed form I DNA (Fig. 4) is the most slowly migrating monomeric CCC form in the first dimension and marks the leftmost band of the arc. In the second dimension, these re-laxed molecules migrated faster than nicked molecules (OC) (Fig. 4) because chloroquine introduced positive supercoils during the second electrophoresis. The arc which presumably originates at this position of relaxed topoisomers extends up-ward and rightup-ward (Fig. 4). Although smearing makes it im-possible to see individual topoisomers in the arc, the presence of topoisomers with progressively lower degrees of superhelic-ity than that of the relaxed form of DNA (under native con-ditions) explains the positioning of the arc. The migration pattern resulted from the removal of helical twists in negatively supercoiled DNA at a low concentration of chloroquin. As a consequence, negatively supercoiled DNA becomes less super-coiled and migrates more slowly than relaxed form I DNA.
[image:6.612.85.267.71.724.2]In contrast, relaxed or positively supercoiled DNA can only be positively supercoiled by intercalation of chloroquine. As a consequence, the intercalator increases the mobility of these DNAs relative to that of form I DNA with reduced (i.e., neg-ative) superhelicity. Such positively supercoiled DNA was not
FIG. 3.oriLytgives rise to plasmid progeny DNA. Total cellular DNAs from
noninduced and induced Akata cells carrying theoriP-oriLytvector p562 were
analyzed on two-dimensional agarose gels (top) and detected after Southern blot hybridization. (A) The schematic drawing indicates the expected migration behavior of negatively supercoiled (form I) and nicked (OC or form II) plasmid DNA during
the first (no intercalating dye) and second (50mg of chloroquine per ml) dimensions
of electrophoresis. In the second dimension, chloroquine introduces positive super-helical turns, i.e., reduces the number of negative supercoils and increases the number of positive supercoils. Individual topoisomers overlap and cannot be clearly
separated, which results in a vertical line. (B and B9) In noninduced cells p562
replicates viaoriP, and different plasmid forms are detected as indicated. Form I
DNA is separated into a streak of topoisomers. Form II DNA of monomeric p562
(OC) is indicated (II), as well as linear DNA (diagonal line). (C and C9) Ten hours
after induction of the lytic cycle, p562 plasmid DNA was amplified viaoriLytand
additional signals were visible on the autoradiogram. A new class of plasmid DNA present only in induced cells gives rise to a horizontal streak. This streak (indicated by ‘‘T’’) is composed of 30 to 40 vertical bands, as shown in panels D and D9. This new class consists of plasmid DNA (mono-, di-, and trimers are visible) which is separated in the first dimension (from left to right) in the absence of an intercalating dye but migrates uniformly and faster than ‘‘normal’’ form I DNA in the second dimension. Linear forms (form III) and OC (form II) of monomeric p562 DNA are indicated. High-molecular-weight DNA, probably consisting of concatemeric forms of p562, is not resolved in this autoradiogram and shows up as a blob in the upper left corner.
3428 PFU¨ LLER AND HAMMERSCHMIDT J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
detectable because no signals which migrated faster than re-laxed DNA were detected. Since the arc formed by p562 DNA is located above the position of the relaxed closed-circle DNA and extends clockwise to the upper right, this DNA consisted of topoisomers which had a net negative supercoiling in the first dimension. This class of topologically different DNA mol-ecules is dependent onoriLytincisas well as on the induction of the lytic phase of EBV’s life cycle (data not shown). More-over, this finding is not unique to theoriP-oriLytvector p562, as indicated by transient experiments with a plasmid containing
oriLyt only (data not shown). The data suggested that the molecules present in the arc might consist of topoisomers with a low degree of negative supercoiling.
Enrichment and analyses of p562 topoisomers on buoyant-density gradients.Smearing of individual topoisomers shown
in Fig. 3 and 4 makes a clean interpretation of the data difficult. In order to obtain independent information onoriLyt topoiso-mers, p562 plasmid DNAs isolated from noninduced and from induced Akata cells were analyzed on CsCl-propidium diiodide buoyant-density gradients. These gradients allow the isolation of different isomers, since the buoyant densities of individual topoisomers are a function of their superhelical state. In the presence of an intercalating dye, the buoyant density of relaxed form I plasmid DNA is higher than that of negatively super-coiled form I DNA (17). This is because under saturating conditions negatively supercoiled DNA takes up more dye than does relaxed or even positively supercoiled CCC DNA. DNA molecules with slightly increased linking numbers (i.e., with intermediate negative superhelicity under native condi-tions) take up intermediate levels of intercalator and therefore have intermediate densities (for an excellent overview, see reference 28).
Low-molecular-weight DNA from uninduced and induced Akata cells harboring p562 was extracted and separated on CsCl buoyant-density gradients in the presence of saturating concentrations of propidium diiodide. DNA was prepared from individual gradient fractions and run on agarose gels, and p562 DNA was detected by Southern blot hybridization. Dur-ing electrophoresis (in the absence of intercalatDur-ing dye), dif-ferent forms of monomeric and oligomeric plasmid DNAs were detected; most noticeably, fractions 4 and 5 from the induced Akata cells contained a ladder of bands extending from the position of OC to the fastest-migrating supercoiled topoisomers of p562 DNA (Fig. 5A, bottom). The ladder of bands, which cannot be detected in uninduced cells (Fig. 5A, top panel), corresponds most likely to the stripe-like signal visible in Fig. 3C and D and the arc in Fig. 4. These DNAs were monomeric covalently closed plasmids as determined by elec-tron microscopy (data not shown).
DNA in fraction 4 shown in the bottom panel of Fig. 5A was further analyzed on one-dimensional agarose gels in the pres-ence of chloroquine and for its sensitivity to topoisomerase. Wheat germ topoisomerase I completely relaxed supercoiled DNA present in fraction 4 from induced Akata cells, which resulted in a single band (I* in Fig. 5B). The comparison of fraction 4 from induced cells in gels lacking chloroquine (Fig. 5A, bottom panel) with fraction 4 in a gel containing 1.5mg of chloroquine per ml (Fig. 5B) was also informative because the way in which chloroquine changes the electrophoretic mobility aids interpretation. The low concentration of chloroquine shifted the pattern of topoisomers in fraction 4 of induced cells to slower mobility (compare fraction 4 bands, induced, in Fig. 5A and B). This shift indicated that the topoisomers in fraction 4 of induced cells are significantly less negatively supercoiled than is the case for plasmids in cells before lytic induction. Chloroquine removes helical twists and introduces positive supercoils into DNA with negative superhelicity. As a conse-quence, these molecules migrate more slowly than does re-laxed form I DNA in the presence of low concentrations of chloroquine, shown in Fig. 5B (compare the position of relaxed [I*] molecules). No positively supercoiled p562 DNA is de-tected, since already positive supercoiled DNA in the presence of chloroquine becomes even more tightly packed than relaxed DNA and would migrate at least as fast as or faster than re-laxed (I*) DNA (Fig. 5B).
[image:7.612.77.272.73.411.2]Our finding, which is based on various independent analyses, is consistent with the presence of newly replicatedoriLyt plas-mid DNAs which are formed during lytic-cycle DNA replica-tion of EBV. In addireplica-tion, a certain proporreplica-tion of the plasmid-like intermediates consists of topoisomers which are only slightly negatively supercoiled.
FIG. 4. Topoisomers oforiLytplasmids with reduced superhelicity. Total
cellular DNA from induced Akata cells harboring theoriP-oriLytplasmid p562
was analyzed on a two-dimensional agarose gel in the absence of intercalating
dye in the first dimension and with 1.5mg of chloroquine per ml in the second
dimension. p562 DNA was visualized after probing of the Southern blot and autoradiography similar to what was done for the analysis shown in Fig. 3. In contrast to the conditions in the experiment shown in Fig. 3, the chloroquine
concentration used was very low (1.5mg/ml) and the class of plasmid topoisomers
which migrated in a horizontal line in the blot shown in Fig. 3B and D (indicated by ‘‘T’’) is now resolved into an arc extending from the relaxed-circle position to the upper right. Mostly negatively supercoiled topoisomers which were not re-solved in the first dimension are separated into a vertical streak, as indicated by a bracket on the right of the scanned and labeled image. OC DNA and the diagonal smear of linear DNA are indicated. The top panel shows the digital image of the autoradiograph shown below, with relevant features and annota-tions.
on November 9, 2019 by guest
http://jvi.asm.org/
DISCUSSION
Our knowledge about DNA replication in the lytic phase of the herpesvirus life cycle is very limited. Although the viralcis -andtrans-acting functions in many different herpesviruses have been identified, insight into the mechanism of herpesvirus DNA replication is restricted to the accumulation of concate-meric high-molecular-weight DNAs in the infected cells and their topographical composition (4, 41, 49). The aspects of initiation and elongation of DNA replication, as well as the structure of early intermediates, have not been investigated previously. This ignorance results partly from the size of
geno-mic DNA (;100 to.250 kbp), which limits manipulation of its replicative intermediates. The construction of a plasmid with two EBV origins of replication and of relatively small size al-lowed the analysis of the DNA’s replicative intermediates. Unexpectedly, early after the onset of lytic replication we de-tected DNAs which are similar to unit-length plasmids and contain one or two newly replicated DNA strands. Rescue experiments with plasmids carryingoriLytonly indicated that monomeric progeny plasmids are true replicative intermedi-ates which do not depend on the design of a dual-origin vector. In addition, part of the newly replicated progeny DNA has an unexpectedly low degree of negative supercoiling.
We do not know how plasmid progeny DNAs fromoriLyt
arise, nor do we know the origin of topoisomers with a reduced negative superhelicity. It appears that induction of lytic repli-cation leads to a loss of negative supercoiling in a part of the plasmid-like progeny DNA, as this could mean that replicated DNA contains only a reduced number of nucleosomes. A re-duction in the number of nucleosomes appears to be the most probable explanation for the low degree of negative supercoil-ing for the products of lytic replication, for two reasons. (i) Herpesvirus DNA is packaged without nucleosomes, and avoiding nucleosome assembly after lytic DNA replication, at least at late times, could be an economical way of doing things. (ii) Induction of transcription of late genes requires DNA replication. Removal of nucleosomes from late promoters by replication could be an obvious factor in, and perhaps a nec-essary component of, switching on EBV late genes.
Another possibility is that histones were lost during isolation of DNA while topoisomerases had the chance to relax the DNA, especially since cellular topoisomerases localize to sites of active viral DNA synthesis (11). This artifact is not very like-ly, since cellular DNA was prepared with the aid of a very rapid lysis protocol in the presence of ionic detergent and chelator. Alternatively, part of these plasmids could be embedded in nucleosomes and part could be positively supercoiled to give a net supercoiling that is only slightly negative. How could par-tially positively supercoiled DNA then be generated during the process of lytic replication? Positively supercoiled DNA orig-inates either through the activity of a reverse gyrase gene pro-duct in archaebacteria (8, 26), through a proposed DNA helix-tracking protein (27) found in Saccharomyces cerevisiae, or upstream of the moving transcription complex of highly active genes under conditions in which the endogenous topoisomer-ases are inactive (29). One intrinsic property of such DNA is its sensitivity toward DNase I (29).
[image:8.612.65.298.75.385.2]This observation is especially interesting since it might provide a clue to the proposed two-stage model of herpesvirus DNA replication: an initial phase of semiconservative plasmid replica-tion via theta-like or Cairns structures provides many cir-cular progeny molecules. A certain proportion of these DNAs contains positively supercoiled regions which might be prefer-entially nicked by DNase I to provide the ideal template DNA for rolling-circle replication. Although herpes simplex virus DNA replication in vitro has been successfully demonstrated with six of seven viral proteins required in vivo to yield con-catemeric molecules, this in vitro synthesis requires neither a specific origin nor the origin-binding protein (42). This obser-vation could support the hypothesis that herpesviruses have a biphasic mode of DNA replication: an early mode of semicon-servative DNA replication which does depend on a functional origin in cis and one viral (and additional cellular) origin-binding protein(s) intransto yield plasmid-like progeny DNA with a range of different linking numbers and a late phase which initiates at nicks introduced preferentially into topoiso-mers with reduced linking numbers to give rise to rolling-circle
FIG. 5. Preparation of p562 topoisomers on CsCl buoyant-density gradients in the presence of propidium diiodide. (A) DNA isolated from noninduced (top)
and lytically induced (bottom) Akata cells carrying theoriP-oriLytvector p562
was prepared on propidium diiodide-containing CsCl buoyant-density gradients. Different fractions (lanes 1 to 5) of the gradients were analyzed on one-dimen-sional agarose gels, and p562 was detected after Southern blot hybridization. As
an internal control,3
H-labeled simian virus 40 form I DNA was included in the gradients, and it localized to fractions 2 and 3 (data not shown). In lytically induced Akata cells, p562 molecules with an increased buoyant density can be detected in fractions 4 and 5, which show a striking pattern on native one-dimensional agarose gels. These topoisomers were separated into 30 to 40 bands (bottom). No such molecules of p562 DNA were detected in uninduced cells in
which this plasmid replicated viaoriP(top). The gels were run in 0.53TAE
buffer without intercalating dye. (B) The topoisomers are completely relaxed by wheat germ topoisomerase, resulting in one major band migrating fast in the
presence of 1.5mg of chloroquine per ml (labeled ‘‘Ip’’). The comparison of
fraction 4 DNA in gels lacking chloroquine (A) with DNA in the gel with chloroquine (B) reveals that the pattern shifts to slower mobility in the presence of chloroquine. This change in migration indicates that in induced cells there are isomers that are significantly less negatively supercoiled than in cells before lytic induction. On CsCl buoyant-density gradients in the presence of propidium diiodide, these isomers take up less intercalating dye than does highly negatively supercoiled DNA; therefore, they are denser than DNA in fraction 2 or 3. The
agarose gel was electrophoresed in 0.53TAE buffer in the presence of 1.5mg of
chloroquine per ml.
3430 PFU¨ LLER AND HAMMERSCHMIDT J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
DNA replication. Such a model would be akin to bacterio-phagelDNA replication (3, 5), including complicated recom-bination intermediates which result from homologous and il-legitimate recombination events (unpublished data).
ACKNOWLEDGMENTS
We thank Bill Sugden for intriguing ideas, continuous support, help-ful discussions, and comments on the manuscript. We also thank Ross Inman for electron microscopy of DNA intermediates, Noreen Warren for her valuable help, and Joachim Mankertz for suggesting the tran-sientoriLytrescue experiment.
Part of this work was initiated at McArdle Laboratory for Cancer Research, University of Wisconsin, Madison, while W.H. was a recip-ient of a personal grant from the Deutsche Forschungsgesellschaft. Also acknowledged are NIH grants to Bill Sugden (CA-22443 and CA-07175). This work was also supported by grant Fa138/6-1 from the Deutsche Forschungsgemeinschaft to W.H.
REFERENCES
1.Adams, A.1987. Replication of latent Epstein-Barr virus genomes in Raji
cells. J. Virol.61:1743–1746.
2.Baer, R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P. J. Farrell, T. J. Gibson, G. Hatfull, G. S. Hudson, S. C. Satchwell, C. Seguin, P. S. Tufnell, and B. G. Barell.1984. DNA sequence and expression of the B95-8
Epstein-Barr virus genome. Nature (London)310:207–211.
3.Bastia, D., and N. Sueoka.1975. Studies on the late replication of phage lambda: rolling-circle replication of the wild type and a partially suppressed
strain, Oam29 Pam80. J. Mol. Biol.98:305–320.
4.Bataille, D., and A. Epstein.1994. Herpes simplex virus replicative
concate-mers contain L components in inverted orientation. Virology203:384–388.
5.Carter, B. J., B. D. Shaw, and M. G. Smith.1969. Two stages in the
repli-cation of bacteriophage lambda DNA. Biochim. Biophys. Acta195:494–505.
6.Challberg, M. D.1986. A method for identifying the viral genes required for
herpesvirus DNA replication. Proc. Natl. Acad. Sci. USA83:9094–9098.
7.Challberg, M. D.1991. Herpes simplex virus DNA replication. Semin. Virol.
2:247–256.
8.Confalonieri, F., C. Elie, M. Nadal, C. de La Tour, P. Forterre, and M. Duguet. 1993. Reverse gyrase: a helicase-like domain and a type I
topo-isomerase in the same polypeptide. Proc. Natl. Acad. Sci. USA90:4753–
4757.
9.Countryman, J., and G. Miller. 1985. Activation of expression of latent Epstein-Barr herpesvirus after gene transfer with a small cloned subfragment
of heterogeneous viral DNA. Proc. Natl. Acad. Sci. USA82:4085–4089.
10.DePamphilis, M. L.1993. Eukaryotic DNA replication: anatomy of an
ori-gin. Annu. Rev. Biochem.62:29–63.
11.Ebert, S. N., D. Subramanian, S. S. Shtrom, I. K. Chung, D. S. Parris, and M. T. Muller.1994. Association between the p170 form of human topo-isomerase II and progeny viral DNA in cells infected with herpes simplex
virus type 1. J. Virol.68:1010–1020.
12.Farrell, P. J., D. T. Rowe, C. M. Rooney, and T. Kouzarides.1989. Epstein-Barr virus BZLF1 trans-activator specifically binds to a consensus AP-1 site
and is related to c-fos. EMBO J.8:127–132.
13.Fixman, E. D., G. S. Hayward, and S. D. Hayward.1995. Replication of Epstein-Barr virus oriLyt: lack of a dedicated virally encoded origin-binding
protein and dependence on Zta in cotransfection assays. J. Virol.69:2998–
3006.
14.Fixman, E. D., G. S. Hayward, and S. D. Hayward.1992.trans-acting
re-quirements for replication of Epstein-Barr virus ori-Lyt. J. Virol.66:5030–
5039.
15.Gahn, T. A., and C. L. Schildkraut.1989. The Epstein-Barr virus origin of
plasmid replication,oriP, contains both the initiation and termination sites of
DNA replication. Cell58:527–535.
16.Glaser, R., and M. Nonoyama.1974. Host cell regulation of induction of
Epstein-Barr virus. J. Virol.14:174–176.
17.Gray, H. B. J., W. B. Upholt, and J. Vinograd.1971. A buoyant method for the determination of the superhelix density of closed circular DNA. J. Mol.
Biol.62:1–19.
18.Gruffat, H., O. Renner, D. Pich, and W. Hammerschmidt.1995. Cellular proteins bind to the downstream component of the lytic origin of DNA
replication of Epstein-Barr Virus. J. Virol.69:1878–1886.
19.Hammerschmidt, W., and J. Mankertz.1991. Herpesviral DNA replication:
between the known and the unknown. Semin. Virol.2:257–269.
20.Hammerschmidt, W., and B. Sugden.1988. Identification and
characteriza-tion oforiLyt, a lytic origin of DNA replication of Epstein-Barr virus. Cell55:
427–433.
21.Hammerschmidt, W., and B. Sugden.1989. Genetic analysis of immortaliz-ing functions of Epstein-Barr virus in human B lymphocytes. Nature
(Lon-don)340:393–397.
22. Hirt, B.1967. Selective extraction of polyoma DNA from infected mouse cell
cultures. J. Mol. Biol.26:365–369.
23. Jacob, R. J., L. S. Morse, and B. Roizman.1979. Anatomy of herpes simplex virus DNA. XII. Accumulation of head-to-tail concatemers in nuclei of infected cells and their role in the generation of the four isomeric
arrange-ments of viral DNA. J. Virol.29:448–457.
24. Jacob, R. J., and B. Roizman.1977. Anatomy of herpes simplex virus DNA.
VIII. Properties of the replicating DNA. J. Virol.23:394–411.
25. Keller, W.1975. Determination of the number of superhelical turns in simian
virus 40 DNA by gel electrophoresis. Proc. Natl. Acad. Sci. USA72:4876–
4880.
26. Kikuchi, A., and K. Asai.1984. Reverse gyrase—a topoisomerase which
introduces positive superhelical turns into DNA. Nature (London)309:677–
681.
27. Koo, H. S., K. Lau, H. Y. Wu, and L. F. Liu.1992. Identification of a DNA
supercoiling activity in Saccharomyces cerevisiae. Nucleic Acids Res.20:
5067–5072.
28. Kornberg, A.1980. DNA replication. W. H. Freeman and Company, New York.
29. Lee, M. S., and W. T. Garrard.1991. Positive DNA supercoiling generates a chromatin conformation characteristic of highly active genes. Proc. Natl.
Acad. Sci. USA88:9675–9679.
30. Lieberman, P. M., J. M. Hardwick, J. Sample, G. S. Hayward, and S. D. Hayward.1990. The Zta transactivator involved in induction of lytic cycle gene expression in Epstein-Barr virus-infected lymphocytes binds to both
AP-1 and ZRE sites in target promoter and enhancer regions. J. Virol.64:
1143–1155.
31. Liebowitz, D., and E. Kieff.1993. The Epstein-Barr virus, p. 107–172.InB. Roizman, R. J. Whitley, and C. Lopez (ed.), The human herpesviruses. Raven Press, New York.
32. Meselson, M., and F. W. Stahl.1958. The replication of DNA in Escherichia
coli. Proc. Natl. Acad. Sci. USA44:671–682.
33. Meuth, M., and H. Green.1974. Induction of a deoxycytidineless state in
cultured mammalian cells by bromodeoxyuridine. Cell2:109–112.
34. Pruss, G. J., and K. Drlica.1986. Topoisomerase I mutants: the gene on pBR322 that encodes resistance to tetracycline affects plasmid DNA
super-coiling. Proc. Natl. Acad. Sci. USA83:8952–8956.
35. Reisman, D., J. Yates, and B. Sugden.1985. A putative origin of replication
of plasmids derived from Epstein-Barr virus is composed of twocis-acting
components. Mol. Cell. Biol.5:1822–1832.
36. Roizman, B.1979. The structure and isomerization of herpes simplex virus
genomes. Cell16:481–484.
37. Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
38. Schepers, A., D. Pich, and W. Hammerschmidt.1993. A transcription factor with homology to the AP-1 family links RNA transcription and DNA
repli-cation in the lytic cycle of Epstein-Barr virus. EMBO J.12:3921–3929.
39. Schepers, A., D. Pich, and W. Hammerschmidt.Activation oforiLyt, the lytic origin of DNA replication of Epstein-Barr virus by BZLF1. Virology, in press.
40. Schepers, A., D. Pich, J. Mankertz, and W. Hammerschmidt.1993.cis-acting elements in the lytic origin of DNA replication of Epstein-Barr virus. J.
Virol.67:4237–4245.
41. Severini, A., A. R. Morgan, D. R. Tovell, and D. L. Tyrrell.1994. Study of the structure of replicative intermediates of HSV-1 DNA by pulsed-field gel
electrophoresis. Virology200:428–435.
42. Skaliter, R., and I. R. Lehman.1994. Rolling circle DNA replication in vitro by a complex of herpes simplex virus type 1-encoded enzymes. Proc. Natl.
Acad. Sci. USA91:10665–10669.
43. Sugden, B., K. Marsh, and J. Yates.1985. A vector that replicates as a plasmid and can be efficiently selected in B-lymphoblasts transformed by
Epstein-Barr virus. Mol. Cell. Biol.5:410–413.
44. Takada, K., K. Horinouchi, Y. Ono, T. Aya, T. Osato, M. Takahashi, and S. Hayasaka.1991. An Epstein-Barr virus-producer line Akata: establishment
of the cell line and analysis of viral DNA. Virus Genes5:147–156.
45. Takada, K., N. Shimizu, S. Sakuma, and Y. Ono.1986.transactivation of the latent Epstein-Barr virus (EBV) genome after transfection of the EBV DNA
fragment. J. Virol.57:1016–1022.
46. Wu, C. A., N. J. Nelson, D. J. McGeoch, and M. D. Challberg.1988. Iden-tification of herpes simplex virus type 1 genes required for origin-dependent
DNA synthesis. J. Virol.62:435–443.
47. Yates, J. L., and N. Guan.1991. Epstein-Barr virus-derived plasmids repli-cate only once per cell cycle and are not amplified after entry into cells. J.
Virol.65:483–488.
48. Yates, J. L., N. Warren, and B. Sugden.1985. Stable replication of plasmids derived from Epstein-Barr virus in various mammalian cells. Nature
(Lon-don)313:812–815.
49. Zhang, X., S. Efstathiou, and A. Simmons.1994. Identification of novel herpes simplex virus replicative intermediates by field inversion gel
electro-phoresis: implications for viral DNA amplification strategies. Virology202:
530–539.