Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Measles Virus-Induced Disruption of the Glial-Fibrillary-Acidic
Protein Cytoskeleton in an Astrocytoma Cell Line (U-251)
W. PAUL DUPREX,1* STEPHEN MCQUAID,2ANDBERT K. RIMA1
School of Biology and Biochemistry, The Queen’s University of Belfast, Belfast BT9 7BL,1and Neuropathology
Laboratory, Royal Group of Hospitals Trust, Belfast BT12 6Bl,2Northern Ireland, United Kingdom
Received 20 October 1999/Accepted 9 December 1999
A recombinant measles virus which expresses enhanced green fluorescent protein (MVeGFP) has been used to infect two astrocytoma cell lines (GCCM and U-251) to study the effect of virus infection on the cytoskeleton. Indirect immunocytochemistry was used to demonstrate the cellular localization of the cytoskeletal compo-nents. Enhanced green fluorescent protein autofluorescence was used to identify measles virus-infected cells. No alteration of the actin, tubulin, or vimentin components of the cytoskeleton was observed in either cell type, whereas a disruption of the glial-fibrillary-acidic protein filament (GFAP) network was noted in MVeGFP-infected U-251 cells. The relative amounts of GFAP present in MVeGFP-infected and unMVeGFP-infected U-251 cells were quantified by image analysis of data sets obtained by confocal microscopy by using vimentin, another inter-mediate filament on which MVeGFP has no effect, as a control.
The advent of reverse genetics for negative-stranded RNA viruses provides new opportunities for the examination and reassessment of various aspects of the virus infection process.
Measles virus (MV) is a Morbillivirus which belongs to the
Paramyxoviridae. Like the other members of this family, MV
has a single-stranded negative-sense RNA genome which is encapsidated by nucleoprotein (N). Six structural genes are encoded by the genome. The polymerase (L) and phosphopro-tein (P) associate with the N prophosphopro-tein to generate the helical ribonucleocapsid structure. Two glycoproteins, fusion (F) and hemagglutinin (H), are embedded in the pleomorphic virion envelope, and these mediate cell entry and fusion (9, 13, 39, 55). The matrix protein (M) associates with both the glycop-roteins and the ribonucleocapsid structure and plays a key role in virion assembly (8, 41).
Many viruses have been shown to cause alterations to the cytoskeleton during in vitro infection (6, 10, 43, 47). For a review, see the work of Cudmore et al. (12). TheMorbillivirus
Canine distemper virus(CDV), has been reported to cause a
total reorganization of the cytoskeleton, with the most notable alterations being in the microtubule and intermediate-filament networks (26).Vesicular stomatitis virus(VSV) infection, first, causes disassembly of the actin filaments and, second, alters the distribution of the microtubules and intermediate filaments (44, 47).Respiratory syncytial virus(RSV) also causes a disrup-tion of the cytoskeleton (7, 21, 52). The effect of MV on the actin cytoskeleton is less clear. One group has reported a striking decrease in the overall number of actin bundles in human fibroblasts infected with MV. They also show a similar disruption upon infection with otherParamyxoviridae(16, 17). Contrary to this, a second group has not been able to demon-strate alterations to the actin cytoskeleton in MV-infected Vero cells (2).
Treatment of MV-infected cells with the actin-depolymeriz-ing agent cytochalasin B (CB) results in the inhibition of virus maturation. This suggests that microfilaments play a role in the
release of budding virions (2, 48, 51). Actin filaments have been shown to have a role in the movement of MV glycopro-teins on the surfaces of infected cells (14). The involvement of actin filaments in the budding of MV has been examined by electron microscopy (4, 5). Again, a close association exists between actin filaments from the outer part of the cytoskeletal network and budding virus, with the filaments protruding into the particles. It has been suggested that budding is possibly the result of a vectorial growth of actin filaments (4). CB inhibits the production of infectious virus particles of other paramyxo-viruses (7, 11, 24). Interestingly, CB has no effect on the mat-uration of VSV (23), which has been unequivocally shown to disrupt the actin cytoskeleton (44, 47). Recently the essential role of cellular actin in the gene expression and morphogenesis of RSV has been described. In this instance RSV infection causes a gross disruption of the actin cytoskeleton (7). Thus, there appears to be confusion in the literature. Additionally, it is not clear whether these alterations are active, i.e., induced to facilitate virus growth, or passive, i.e., simply caused as a result of infection but playing no formal role in virus replication.
A number of virus genomes, such asSimian virus 5,Mouse
hepatitis virus,Human herpesvirus, and Simian varicella-zoster
virus, have been engineered to express green fluorescent
pro-tein (GFP) (18, 19, 25b, 32). Recently the gene encoding en-hanced GFP (EGFP) has been introduced into the MV ge-nome, and a recombinant virus (MVeGFP) has been rescued (25a). We have demonstrated that EGFP is detectable in cells in the early stages of infection (13a). In all cases diffuse EGFP autofluorescence was detectable before viral antigen was de-tected by immunocytochemistry. Therefore, EGFP appears to be an ideal indicator of early MV cell infection and the recom-binant virus appears to be very useful for in vitro studies and may also be beneficial for in vivo investigations. The diffuse nature of EGFP autofluorescence makes MVeGFP an ideal candidate for assessing the effects of virus replication on the cytoskeletons of MVeGFP-infected cells. As tubulin has been shown to stimulate MV RNA synthesis in vitro (36) and the fate of actin within MV-infected cells remains unclear (2, 16, 17), we decided to use MVeGFP to investigate the effects of MV infection on the cytoskeleton. Confocal scanning laser microscopy (CSLM) was used to gain maximal resolution in dually labeled specimens.
* Corresponding author. Mailing address: School of Biology and Biochemistry, The Queen’s University of Belfast, Medical Biology Centre, 97 Lisburn Rd., Belfast BT9 7BL, Northern Ireland, United Kingdom. Phone: 01232 272060. Fax: 01232 236505. E-mail: p.duprex @qub.ac.uk.
3874
on November 9, 2019 by guest
http://jvi.asm.org/
MVeGFP infects astrocytoma cells. MV infection of the central nervous system (CNS) is a rare event (27, 30). Oligo-dendrocytes and neurons are the predominantly infected cell types, although infected astrocytes have also been described (1, 30, 31, 34). In this study we have used two astrocytoma cell lines, the first being GCCM cells, which have been used to study MV cell-to-cell spread (13a). This cell line was derived from an anaplastic astrocytoma (grade IV). The second cell line used was U-251 MG cells, which were established from a human glioma (3). These cells have been used to examine the induction of inflammatory cytokines upon MV infection (22, 45). Both cell lines were maintained in RPMI 1640 medium supplemented with 5% fetal calf serum. MVeGFP virus was rescued from a full-length antigenomic clone (25a) by using a cell line which expresses T7 RNA polymerase and the N and P proteins of MV (42). MVeGFP was propagated in African green monkey kidney cells (Vero). The gene encoding EGFP is present in an additional transcription unit (ATU) which is inserted before the N gene in the MV genome in the most promoter-proximal position. Due to this location, large amounts of EGFP are produced in infected cells. No major effects on the replication of the virus and the type of cell-pathogenic effect generated was observed (25a). Autofluores-cence was readily observed during virus rescue and propaga-tion by UV microscopy. Cells which showed none of the well-characterized signs of MV-induced cell-pathogenic effects were frequently observed. This demonstrates the strength of using MVeGFP for these experiments in that observations of the cytoskeletons of cells in the very early stages of virus in-fection can be made.
MVeGFP has no effect on the actin-, tubulin-, or
vimentin-based cytoskeletons of astrocytoma cells.GCCM and U-251
cells were grown to a confluence of 80% on glass coverslips. Cells were infected with MVeGFP at multiplicity of infection of 0.01 for 1 h at 37°C, after which time the inoculum was removed and maintenance medium, RPMI 1640 containing 2% fetal calf serum, was added. Infections were carried out for 50 h at 37°C. During this time the cells attained 95 to 100% confluence. Cells were permeabilized and fixed by using freshly prepared 4% paraformaldehyde. The cytoskeletons of the cells were visualized with a monoclonal antibody specific for either tubulin (Sigma) or vimentin (Dako). Antivimentin and antitu-bulin antibodies were diluted in phosphate-buffered saline (PBS) containing 0.5% Triton X-100 (1:100 and 1/1,500, re-spectively). The detergent was included to increase the perme-ability of the paraformaldehyde-fixed cells. Antibodies were incubated on the coverslips for 20 h at 4°C. Unbound antibod-ies were removed by three washes in PBS, each lasting 5 min. CY3-conjugated sheep anti-mouse immunoglobulin G (Sigma) was used as a secondary antibody. Dilutions (1:40) were made in PBS containing 0.5% Triton X-100, and the antibody was incubated on the coverslips for 3 h at 37°C. Unbound antibody was removed as described above. Tetramethyl rhodamine iso-thiocyanate (TRITC)-conjugated phalloidin (Sigma), a fluo-rescently conjugated phallotoxin from Amanita phalloides
which specifically binds to F-actin, was used to directly stain the microfilaments. TRITC-conjugated phalloidin (200 ng/ml) in PBS was incubated on the coverslips for 2 h at 37°C. Excess phalloidin was removed by a single PBS wash. Coverslips were mounted with Citifluor (Amersham). A Leica TCS/NT confo-cal microscope equipped with a krypton-argon laser as the source for the ion beam was used to examine the samples for fluorescence. CY3-stained samples were imaged by excitation at 568 nm with a 564- to 596-band-pass emission filter. EGFP was visualized by virtue of its autofluorescence by excitation at 488 nm with a 506- to 538-band-pass emission filter. Data sets
were collected by dual excitation, and image stacks were accu-mulated every 0.5m through an optical plane of 5m. Com-posite images were generated for the separate EGFP (green) and TRITC (red) channels in single-excitation mode to pre-vent spillover artifacts. Images were accumulated from regions of the monolayer which contained uninfected and infected cells and thereby permitted direct comparison of their cy-toskeletal networks.
MVeGFP infection of GCCM and U-251 cells led to exten-sive fusion. Syncytia which are typical of MV-infected cells were observed. Nuclei clustered in the centers of the syncytia, and possibly due to a nonspecific accumulation of EGFP, these were brightly autofluorescent, as is shown for both cell types in Fig. 1. EGFP was present diffusely throughout the cytoplasm, and no overlap was observed between the green and red chan-nels. This is particularly important as the most readily detect-able MV antigens, N and P, produce a pronounced punctate staining pattern in extensively fused syncytia. Under these con-ditions, any overlap between channels by standard, dual-label-ing indirect immunofluorescence may give the impression that alterations have occurred in the cytoskeleton. This effect is absent when MVeGFP infection is used in conjunction with CSLM, and thus this combined technology provides an excel-lent approach for examination of the effects of MV on the cytoskeleton. Using this approach, we investigated the effects of infection on microtubule, intermediate-filament, and micro-filament components of the cytoskeletons of U-251 and GCCM cells.
Actin. No disruption of the actin-based cytoskeleton was
observed upon MVeGFP infection of either cell type (Fig. 1A). Actin is present in two forms within cells, a globular, mono-meric form (G-actin) and a polymerized, filamentous form (F-actin). It is the latter form which contributes to the cytoskel-eton and is detected by phalloidin (29). The distributions of F-actin in GCCM and U-251 cells were similar to that observed in a previous study (20). In uninfected cells F-actin was present in long parallel stress fibers. These ran along the long axes of the cells. Cortical filaments outlined the peripheries of both cell types. In MVeGFP-infected cells which formed syncytia, the microfilaments were integrated into a larger, but organi-zationally similar, network. Extended fibers, which were greater in length than those of the single cells, spanned the syncytia (Fig. 1A, GCCM), indicating that actin polymerization does not seem to be inhibited by virus infection. Clumping of actin was noted in the U-251 cells (Fig. 1A). Equivalent amounts of F-actin appear to be present in both uninfected and infected cells. Actin bundles were more prevalent in MVeGFP-infected GCCM cells (Fig. 1A) than in U-251 cells. These observations confirm what was previously shown for nonrecombinant MV-infected Vero cells (2) and contrast with the results of two studies (16, 17) which observed severe actin disruption during MV infection of a human lung cell line. We have confirmed that MVeGFP infection of Vero cells causes no disruption of the actin cytoskeleton (data not shown). In a recent report, (24) colocalization of human parainfluenza virus type 3 (HPIV3) ribonucleoprotein (RNP) and actin microfila-ments was observed in infected CV-1 cells by confocal micros-copy with an HPIV3 polyclonal antiserum. The extent of co-localization was striking and demonstrates the usefulness of CSLM in this type of investigation. In MV-infected cells we have never observed a close relationship between MV RNP and the actin cytoskeleton using either polyclonal or monoclo-nal antibodies (anti-P or anti-N antibodies) for staining. This result is in spite of the fact that actin is known to associate with MV nucleocapsid (36). Rather, a punctate perinuclear staining pattern in which antigen is detected in MV-induced
cytoplas-VOL. 74, 2000 NOTES 3875
on November 9, 2019 by guest
http://jvi.asm.org/
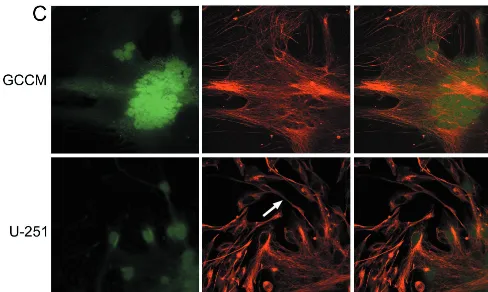
FIG. 1. Effect of MVeGFP infection on the cytoskeletons of GCCM and U-251 astrocytoma cells. Astrocytoma cells were infected with MVeGFP at a multiplicity of infection of 0.01 for 50 h. Cells were fixed and examined by CSLM for autofluorescence and immunoreactivity. Micrographs represent an 8- to 10-m-deep composite optical section, and all images were obtained in double-excitation mode. EGFP was detected by virtue of its autofluorescence (green). (A) Actin microfilaments in GCCM and U-251 cells were visualized using TRITC-labeled phalloidin (red). The arrow indicates a single actin stress fiber in a GCCM cell. The arrowhead indicates a nonfibrillary aggregation of actin in a U-251 cell. (B) Tubulin was visualized using a monoclonal antibody and a CY3-conjugated secondary antibody (red). The arrow indicates a tubulin-rich astrocytic process from a GCCM cell. The arrowhead in the U-251 panel indicates tubulin accumulation around the nucleus of an unaffected cell. (C) Vimentin was visualized using a monoclonal antibody and a CY3-conjugated secondary antibody (red). The arrowhead indicates an astrocytic process from a U-251 cell. Magnification,⫻400.
on November 9, 2019 by guest
http://jvi.asm.org/
mic inclusion bodies is observed. The HPIV3 system appears to be unique in that the actin microfilaments seem to be di-rectly involved in viral-RNA synthesis in vivo (24). Interest-ingly, no disruption of the actin cytoskeleton was observed upon HPIV3 infection.
Tubulin.This component of the cytoskeleton has been
im-plicated as having a role in the MV life cycle, possibly as a subunit of the viral RNA polymerase (35). The distribution of the microtubules in MV-infected cells has not been examined previously by immunocytochemistry. The tubulin-based cy-toskeletons are quite similar in organization in both the GCCM and U-251 cells. The microtubule bundles are thinner and more filamentous than F-actin stress fibers which cross syncytia. Generally, tubulin was present throughout the cell at similar levels, although the processes seemed to be particularly rich in microtubules (Fig. 1B, GCCM). There may be a slight accumulation of tubulin around the nuclei of the U-251 cells (Fig. 1B). Tubulin distribution was examined in MVeGFP-infected GCCM cells. No disruption of the cytoskeleton was observed in infected cells. Filaments were longer in the syncy-tia, indicating that the dynamic process of microtubule assem-bly is not perturbed in infected cells engulfed in syncytia. One investigation has reported a thickening of the microtubules in Hep-2 cells infected with the closely relatedMorbillivirusCDV (26). Bright foci and thick, long bundles crossing near or among the multiple nuclei were also observed in these syncytia. We have not been able to detect any such accumulation in MV-infected astrocytoma cells. Disruption of the microtubules has been suggested to have a role in the bipolar budding of
Sendai virus(50). A mutant virus which buds in a bipolarized
manner has alterations in the M protein, and it has been suggested that this protein may cause the alteration of micro-tubules. Involvement of the VSV M protein in microtubule
disruption has also been suggested (47). In that study major changes in tubulin distribution were detected soon after VSV infection. As is the case for MV, a role has also been suggested for tubulin in VSV viral transcription (35).
Vimentin. The intermediate filament, vimentin, is a major
[image:4.612.62.550.73.365.2]component of the cytoskeleton. The fate of the vimentin net-work in MV-infected cells has not been examined previously. In uninfected GCCM and U-251 cells the overall structure of the vimentin component of the cytoskeleton was similar in organization to the fine structure of filamentous tubulin. It appears, however, that the filaments are less well organized in parallel arrays than either the microtubules or the microfila-ments. Astrocytic processes were particularly detectable in the U-251 cells by vimentin staining (Fig. 1C). We have previously shown that these processes mediate cell-to-cell spread of MV in vitro (13a). Once again no disassembly of this intermediate filament was observed in MVeGFP-infected cells (Fig. 1C). Extended filaments which were longer than those present in single cells were visible in syncytia, again indicating that as-sembly is not noticeably impaired within infected cells, as was also the case for actin and tubulin. A number of viruses have been shown to cause alterations in the intermediate-filament-based cytoskeletons (37, 46). RSV infection of Hep-2 cells leads to morphological changes of vimentin and an overall reduction in abundance, possibly due to proteolytic degrada-tion (21). CDV infecdegrada-tion of epithelial cells has been shown to lead to a disruption of the intermediate-filament network (26). Disruption of the cytoskeleton therefore seems to depend on the virus studied. Here we clearly demonstrate that MV infection does not perturb the microfilament-, intermediate-filament-, or microtubule-based cytoskeletons of the two astro-cytoma cell lines even when cell-pathogenic effect has pro-gressed to form large, but intact, syncytia.
FIG. 1—Continued.
VOL. 74, 2000 NOTES 3877
on November 9, 2019 by guest
http://jvi.asm.org/
MVeGFP disrupts the GFAP-based cytoskeleton of
astrocy-toma cells.Glial-fibrillary-acidic protein (GFAP) is an
inter-mediate filament of the astrocytic cytoskeleton. This protein is found almost exclusively in astrocytes and is therefore com-monly used as a marker for these cells (33). Its initial expres-sion marks the differentiation of precursor cells into astrocytes, and its up-regulation accompanies the reactive response to
[image:5.612.78.525.78.539.2]CNS injury (15). Due to the contribution of GFAP to the astrocyte cytoskeleton, we examined the effects of MVeGFP infection on the organization of this protein. Immunocyto-chemistry was carried out as described above. GFAP was de-tected using a rabbit polyclonal antiserum (Dako) at a dilution of 1:100 in PBS containing 0.5% Triton X-100. CY3-conju-gated sheep anti-rabbit immunoglobulin G (Sigma) was used
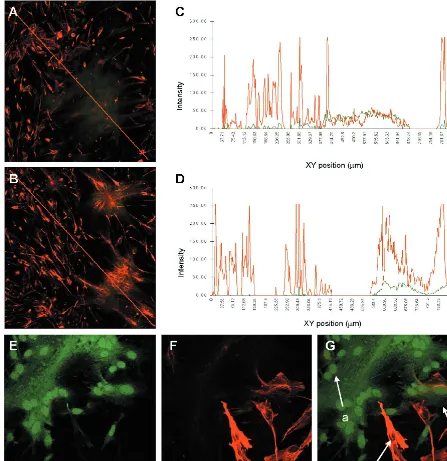
FIG. 2. Effect of MVeGFP infection on the GFAP and vimentin cytoskeleton. U-251 cells were infected, fixed, and examined by CSLM for autofluorescence and immunoreactivity, as described for Fig. 1. GFAP was visualized using a polyclonal antiserum and CY3-conjugated secondary antibody (red). Vimentin was visualized using a monoclonal antibody and a CY3-conjugated secondary antibody (red). EGFP autofluorescence (green) indicates infected cells. (A) Disruption of GFAP within MVeGFP-infected syncytia. Magnification,⫻160. The red line crossing the image indicates the region selected for quantification of green and red fluorescence using the TCS/NT software. (B) Lack of disruption of the vimentin cytoskeleton. The line used for subsequent quantification is shown in red. Magnification,⫻160. (C) Intensity profile obtained from panel A showing a correlation between the decrease in GFAP (red) and the increase in EGFP (green) autofluorescence. (D) Intensity profile obtained from panel B showing no alteration in vimentin (red) staining within areas of infection (green). (E to G) Severe disruption of GFAP cytoskeleton in U-251 cells. (E) Infection of cells shown by EGFP autofluorescence. (F) Vimentin staining. (G) Overlaid image. Magnification,⫻400. Strongly positive GFAP cells were readily observed (arrow c); these were not present within the syncytia. In more recently infected cells at the peripheries of the syncytia (arrow b), the intermediate-filament network was partially disrupted and the overall amount of GFAP staining was diminished. A residual amount of GFAP occasionally remained associated with the nuclei of cells in the central areas of the syncytia (arrow a).
on November 9, 2019 by guest
http://jvi.asm.org/
as the secondary antibody. Immunocytochemical detection of GFAP in paraformaldehyde-fixed GCCM cells proved prob-lematic (data not shown). This was disappointing, as we wished to be consistent and continue to examine the cytoskeleton in MVeGFP-infected cells using EGFP autofluorescence as an indicator of infection. For this, paraformaldehyde fixation was a prerequisite, as EGFP autofluorescence is rapidly lost when all organic fixatives are used. Detection of GFAP expression in the U-251 cells was more satisfactory. Nevertheless, it was important to use the cells at a low passage number due to the overall decrease in the levels of expression of GFAP as cells were cultured. Not all U-251 cells stained equally for GFAP, and it proved essential to carry out incubations in the presence of detergent to improve the detection of the fibrillary network. However, greater than 99% of cells were GFAP positive, per-mitting a satisfactory examination of the effects of MVeGFP infection on this intermediate filament in these cells. Unin-fected U-251 cells stain brightly for GFAP, and MVeGFP was observed to efficiently infect GFAP-positive cells and form syncytia (Fig. 2A). At this low magnification large numbers of infected and uninfected cells are shown. In the uninfected cells GFAP was present as a fibrillary network which was similar in organization to that of vimentin. Astrocytic processes, which connect the cells, also stained positive for GFAP. A severe disruption of the GFAP cytoskeletal network was seen in MVeGFP-infected cells, and the overall amount of GFAP staining was reduced compared to that of the uninfected cells. Intensity profiles were plotted, using quantification software installed on the Leica TCS/NT confocal microscope, to assess the overall levels of fluorescence derived from the presence of GFAP and EGFP. A line was drawn across the composite data sets through a region of uninfected and infected cells. The analysis software was used to determine the total intensity of red and green in each pixel present along the length of this line, and the results were obtained as graphs (Fig. 2C and D). A direct correlation between infection (EGFP positive) and decrease in GFAP levels was observed (Fig. 2C). The line from which this profile was obtained is shown in red in Fig. 2A. This type of analysis was repeated for complete data sets collected from 10 distinct syncytia present in different areas of the in-fected monolayer. Similar profiles were obtained in all cases. Duplicate coverslips, infected at the same time with the same virus pool, were stained, as described above, for the interme-diate filament vimentin and are shown at the same magnifica-tion for comparison (Fig. 2B). No disrupmagnifica-tion of the vimentin cytoskeleton had been observed previously (Fig. 1C). There-fore, this intermediate-filament protein served as the best con-trol for this type of analysis. In this case no decrease in the intensity of the vimentin staining was observed in the infected area (Fig. 2D). Again, the line chosen from which this profile was produced is shown in red in Fig. 2B. Ten control data sets showed no diminution of the amount of vimentin in infected cells. An extreme example of GFAP disruption is shown at a higher magnification (Fig. 2E to G). In this case GFAP was barely detectable within the main body of the syncytium. Pos-itive staining was observed within the syncytium, albeit at a very low level. This residual GFAP seemed to aggregate around the nucleus. Cells on the periphery of a syncytium can be assumed to be more recently infected than those in the center. GFAP staining in these infected cells was diminished compared to that in the uninfected cells, and the cytoskeletal network showed a certain degree of reorganization. This seems, therefore, to represent an intermediate stage in the disintegration of the GFAP cytoskeletal network. Association of residual GFAP with the nucleus was also observed. Exam-ination of the GFAP fibrils demonstrated that they were much
shorter than those in uninfected cells. Therefore, a reorgani-zation of the GFAP-based cytoskeleton occurs in many of the infected astrocytoma cells upon MVeGFP infection. This has not been previously reported for MV.
The closely relatedMorbillivirusCDV causes a demyelinat-ing disease (reviewed by Summers and Appel [49]). CDV pri-marily infects astrocytes producing intracytoplasmic and in-tranuclear inclusion bodies (40). A small percentage (5%) of macrophages are also thought to be infected in the acute de-myelinating lesions (38). Double labeling-immunohistochem-istry has been used to detect viral antigen and GFAP in astro-cytes present in, or derived from, brain tissues of CDV-infected animals. In vitro-CDV-infected primary cultures have also been examined (25, 54, 56). An overall decrease has been observed, both in vivo and in vitro, in the numbers of GFAP-positive cells, the prevalence of astrocytic processes, and GFAP staining in some cells (53, 57). Neither of these studies, however, links this diminution in GFAP staining with a reor-ganization of the cytoskeleton. Infection of primary fetal as-trocytes with a lytic varicella-zoster virus causes a down-regu-lation or modification of GFAP expression (28). Morphological changes in the GFAP cytoskeleton which were very similar to those observed for MV infection were observed (Fig. 2), dem-onstrating that an alteration of the GFAP organization is not without precedent, albeit, in the case of varicella-zoster virus, in a lytic virus.
In this study we set out to examine the effect of MV infection on the cytoskeleton. MVeGFP was used to facilitate the ex-amination of the cytoskeleton by CSLM because of its ability to express EGFP, which produces a diffuse cytoplasmic autofluo-rescence. We observed no disruption of the actin-, vimentin-, or tubulin-based cytoskeletal networks. A disruption of the GFAP cytoskeleton was observed. A direct correlation be-tween MVeGFP infection and a decrease in GFAP amount was confirmed using quantitative confocal fluorescence micros-copy. This is the first time that this effect has been noted for a fusogenic virus. Whether this disruption is a passive or active phenomenon remains unclear. It also remains to be seen if this decrease in GFAP mirrors the in vivo situation. This is an in vitro study which has examined effects in transformed cell lines because they expressed appreciable levels of GFAP. Neverthe-less, this finding is important as the effect of MV on GFAP may give rise to an underestimation of the numbers of infected astrocytes in MV infection of the CNS.
We are very grateful to Martin Billeter for advice and constructive criticism throughout the course of this study. We acknowledge the help of Uta Gassen in the critical reading of the manuscript. We thank Roy Creighton for photographic work, Paula Haddock for excellent tech-nical assistance, and Aaron Maule for advice on phalloidin staining.
This work was supported by the Wellcome Trust (grant 047245).
REFERENCES
1.Allen, I. V., S. McQuaid, J. McMahon, J. Kirk, and R. McConnell.1996. The significance of measles virus antigen and genome distribution in the CNS in SSPE for mechanisms of viral spread and demyelination. J. Neuropathol. Exp. Neurol.55:471–480.
2.Bedows, E., K. M. Rao, and M. J. Welsh.1983. Fate of microfilaments in Vero cells infected with measles virus and herpes simplex virus type 1. Mol. Cell. Biol.3:712–719.
3.Bigner, D. D., S. H. Bigner, J. Ponten, B. Westermark, M. S. Mahaley, E. Ruoslahti, H. Herschman, L. F. Eng, and C. J. Wikstrand.1981. Heteroge-neity of genotypic and phenotypic characteristics of fifteen permanent cell lines derived from human gliomas. J. Neuropathol. Exp. Neurol.40:201–229. 4.Bohn, W., G. Rutter, H. Hohenberg, K. Mannweiler, and P. Nobis.1986. Involvement of actin filaments in budding of measles virus: studies on cy-toskeletons of infected cells. Virology149:91–106.
5.Bohn, W., K. Mannweiler, H. Hohenberg, and G. Rutter.1987. Replica-immunogold technique applied to studies on measles virus morphogenesis. Scanning Microsc.1:319–330.
VOL. 74, 2000 NOTES 3879
on November 9, 2019 by guest
http://jvi.asm.org/
6.Bowden, D. S., J. S. Pedersen, B. H. Toh, and E. G. Westaway. 1987. Distribution by immunofluorescence of viral products and actin-containing cytoskeletal filaments in rubella virus-infected cells. Arch. Virol.92:211–219. 7.Burke, E., L. Dupuy, C. Wall, and S. Barik.1998. Role of cellular actin in the gene expression and morphogenesis of human respiratory syncytial virus. Virology252:137–148.
8.Cathomen, T., H. Y. Naim, and R. Cattaneo.1998. Measles virus with altered envelope protein cytoplasmic tails gain cell fusion competence. J. Virol.
72:1224–1234.
9.Cattaneo, R., and J. K. Rose.1993. Cell fusion by the envelope glycoproteins of persistent measles viruses which cause lethal human brain disease. J. Vi-rol.67:1493–1502.
10.Ceccaldi, P. E., F. Valtorta, S. Braud, R. Hellio, and H. Tsiang.1997. Alteration of the actin-based cytoskeleton by rabies virus. J. Gen. Virol.
78:2831–2835.
11.Chen, W. F., M. M. Soong, S. C. Liang, and D. M. Wang.1986. Cytochalasin B changes the cytoskeletal organisation in Newcastle disease virus-infected cells. Proc. Natl. Sci. Counc. Repub. China10:137–144.
12. Cudmore, S., I. Reckmann, and M. Way.1997. Viral manipulations of the actin cytoskeleton. Trends Microbiol.5:142–148.
13. Do¨rig, R. E., A. Marcil, A. Chopra, and C. D. Richardson.1993. The human CD46 is a receptor for measles virus (Edmonston strain). Cell75:295–305. 13a.Duprex, W. P., S. McQuaid, L. Hangartner, M. A. Billeter, and B. K. Rima.
1999. Observation of measles virus cell-to-cell spread in astrocytoma cells by using a green fluorescent protein-expressing recombinant virus. J. Virol.
73:9568–9575.
14. Ehrnst, A., and K. G. Sundqvist.1976. The mechanisms of appearance of viral glycoproteins at the cell surface membrane. Exp. Cell Biol.44:198–225. 15. Eng, L. F., and R. S. Ghirnikar.1994. GFAP and astrogliosis. Brain Pathol.
4:229–237.
16. Fagraeus, A., D. L. Tyrrell, R. Norberg, and E. Norrby.1978. Actin filaments in paramyxovirus-infected human fibroblasts studied by indirect immunoflu-orescence. Arch. Virol.57:291–296.
17. Fagraeus, A., M. Bottiger, L. Heller, and E. Norrby.1981. Replication of poliovirus and measles virus in cultures of human lymphoblastoid and of Burkitt lymphoma cell lines. Arch. Virol.69:229–237.
18. Fischer, F., C. F. Stegen, C. A. Koetzner, and P. S. Masters.1998. Construc-tion of a mouse hepatitis virus recombinant expressing a foreign gene. Adv. Exp. Med. Biol.440:291–295.
19. Foster, T. P., G. V. Rybachuk, and K. G. Kousoulas.1998. Expression of the enhanced green fluorescent protein by herpes simplex virus type 1 (HSV-1) as an in vitro or in vivo marker for virus entry and replication. J. Virol. Methods75:151–160.
20. Furukawa, R., and M. Fechheimer.1997. The structure, function, and as-sembly of actin filament bundles. Int. Rev. Cytol.175:29–90.
21. Garcia-Barreno, B., J. L. Jorcano, T. Aukenbauer, C. Lopez-Galindez, and J. A. Melero.1998. Participation of cytoskeletal intermediate filaments in the infectious cycle of human respiratory syncytial virus (RSV). Virus Res.
9:307–321.
22. Ghali, M., and J. Schneider-Schaulies.1998. Receptor (CD46)- and repli-cation-mediated interleukin-6 induction by measles virus in human astrocy-toma cells. J. Neurovirol.4:521–530.
23. Griffin, J. A., and R. W. Compans.1979. Effect of Cytochalasin B on the maturation of enveloped viruses. J. Exp. Med.150:379–391.
24. Gupta, S., B. P. De, J. A. Drazba, and A. K. Banerjee.1998. Involvement of actin microfilaments in the replication of human parainfluenza virus type 3. J. Virol.72:2655–2662.
25. Hamburger, D., C. Griot, A. Zurbriggen, C. Orvell, and M. Vandevelde.
1991. Loss of virulence of canine distemper virus is associated with a struc-tural change recognised by a monoclonal antibody. Experientia47:842–845. 25a.Hangartner, L.1997. M.S. thesis. University of Zurich, Zurich, Switzerland. 25b.He, B., R. G. Paterson, C. D. Ward, and R. A. Lamb.1997. Recovery of infectious SV5 from cloned DNA and expression at a foreign gene. Virology
237:249–260.
26. Howard, J. M., B. S. Eckert, and L. Y. Bourguignon.1983. Comparison of cytoskeletal organisation in canine distemper virus-infected and uninfected cells. J. Gen. Virol.64:2379–2385.
27. Jabbour, J. T., D. A. Duenas, J. L. Sever, H. M. Krebs, and L. Horta-Barbosa.1972. Epidemiology of subacute sclerosing panencephalitis (SSPE). A report of the SSPE registry. JAMA220:959–962.
28. Kennedy, P. G., E. O. Major, R. K. Williams, and S. E. Straus.1994. Down-regulation of glial fibrillary acidic protein expression during acute lytic varicella-zoster virus infection of cultured human astrocytes. Virology205:
558–562.
29. Korn, E. D.1982. Acanthamoeba castellanii: methods and perspectives for study of cytoskeleton proteins. Methods Cell Biol.25:313–332.
30. Liebert, U. G.1997. Measles virus infections of the central nervous system. Intervirology40:176–184.
31. Liebert, U. G., K. Baczko, H. Budka, and V. ter Meulen.1986. Restricted expression of measles virus proteins in brains from cases of subacute scle-rosing panencephalitis. J. Gen. Virol.67:2435–2544.
32. Mahalingam, R., M. Wellish, T. White, K. Soike, R. Cohrs, B. K. Klein-schmidt-DeMasters, and D. H. Gilden.1998. Infectious simian varicella virus expressing the green fluorescent protein. J. Neurovirol.4:438–440. 33. McLendon, R. E., and D. D. Bigner.1994. Immunohistochemistry of the glial
fibrillary acidic protein: basic and applied considerations. Brain Pathol.
4:221–228.
34. Mesquita, R., E. Castanos-Velez, P. Biberfeld, R. M. Troian, and M. M. de Siqueira.1998. Measles virus antigen in macrophage/microglial cells and astrocytes of subacute sclerosing panencephalitis. APMIS106:553–561. 35. Moyer, S. A., S. C. Baker, and J. L. Lessard.1986. Tubulin: a factor
neces-sary for the synthesis of both Sendai virus and vesicular stomatitis virus RNAs. Proc. Natl. Acad. Sci. USA83:5405–5409.
36. Moyer, S. A., S. C. Baker, and S. M. Horikami.1990. Host cell proteins required for measles virus reproduction. J. Gen. Virol.71:775–783. 37. Murti, K., and R. Goorha.1989. Synthesis of frog virus 3 proteins occurs on
intermediate filament-bound polyribosomes. Biol. Cell65:205–214. 38. Mutinelli, F., M. Vandevelde, C. Griot, and A. Richard.1989. Astrocytic
infection in canine distemper virus-induced demyelination. Acta Neuro-pathol.77:333–335.
39. Naniche, D., G. Varior-Krishnan, F. Cervoni, T. F. Wild, B. Rossi, C. Ra-bourdin-Combe, and D. Gerlier.1993. Human membrane cofactor protein (CD46) acts as a cellular receptor for measles virus. J. Virol.67:6025–6032. 40. Pearce-Kelling, S., W. J. Mitchell, B. A. Summers, and M. J. Appel.1990. Growth of canine distemper virus in cultured astrocytes: relationship to in vivo persistence and disease. Microb. Pathog.8:71–82.
41. Peebles, M. E.1991. Paramyxovirus M proteins. Pulling it all together and taking it on the road, p. 247–256.InD. W. Kingsbury (ed.), The paramyxo-viruses. Plenum Press, New York, N.Y.
42. Radecke, F., P. Spielhofer, H. Schneider, K. Kaelin, M. Huber, C. Dotsch, G. Christiansen, and M. A. Billeter.1995. Rescue of measles virus from cloned DNA. EMBO J.14:5773–5784.
43. Rubino, S., A. Monaco, A. Degener, and P. Cappuccinelli.1991. Cell micro-filaments disorganisation is an early cytopathic effect in mengo virus repli-cation. Microbiologica14:95–102.
44. Rutter, G., and K. Mannweiler.1977. Alterations of actin-containing struc-tures in BHK21 cells infected with Newcastle disease virus and vesicular stomatitis virus. J. Gen. Virol.37:233–242.
45. Schneider-Schaulies, J., S. Schneider-Schaulies, and V. ter Meulen.1993. Differential induction of cytokines by primary and persistent measles virus infections in human glial cells. Virology195:219–228.
46. Sharpe, A. H., L. B. Chen, and B. N. Fields. 1982. The interaction of mammalian reoviruses with the cytoskeleton of monkey kidney CV-1 cells. Virology120:399–411.
47. Simon, K. O., P. A. Whitaker-Dowling, J. S. Youngner, and C. C. Widnell.
1990. Sequential disassembly of the cytoskeleton in BHK21 cells infected with vesicular stomatitis virus. Virology177:289–297.
48. Stallcup, K. C., C. S. Raine, and B. N. Fields.1983. Cytochalasin B inhibits the maturation of measles virus. Virology124:59–74.
49. Summers, B. A., and M. J. Appel.1994. Aspects of canine distemper virus and measles virus encephalomyelitis. Neuropathol. Appl. Neurobiol.20:525– 534.
50. Tashiro, M., J. T. Seto, H. D. Klenk, and R. Rott.1993. Possible involvement of microtubule disruption in bipolar budding of a Sendai virus mutant, F1-R, in epithelial MDCK cells. J. Virol.67:5902–5910.
51. Tyrrell, D. L., and A. Ehrnst.1979. Transmembrane communication in cells chronically infected with measles virus. J. Cell Biol.81:396–402.
52. Ulloa, L., R. Serra, A. Asenjo, and N. Villanueva.1998. Interactions between cellular actin and human respiratory syncytial virus (HRSV). Virus Res.
53:13–25.
53. Vandevelde, M., P. Bichsel, S. Cerruti-Sola, A. Steck, F. Kristensen, and R. J. Higgins.1983. Glial proteins in canine distemper virus-induced demy-elination. A sequential immunocytochemical study. Acta Neuropathol.59:
269–226.
54. Vandevelde, M., A. Zurbriggen, R. J. Higgins, and D. Palmer.1985. Spread and distribution of viral antigen in nervous canine distemper. Acta Neuro-pathol.67:211–218.
55. Wild, T. F., and R. Buckland.1995. Functional aspects of envelope-associ-ated measles virus proteins, p. 51–64.InV. ter Meulen and M. A. Billeter (ed.), Measles virus. Springer-Verlag KG, Berlin, Germany.
56. Zurbriggen, A., M. Vandevelde, and M. Dumas.1986. Secondary degenera-tion of oligodendrocytes in canine distemper virus infecdegenera-tion in vitro. Lab. Investig.54:424–431.
57. Zurbriggen, A., and M. Vandevelde.1983. Canine distemper virus-induced glial cell changes in vitro. Acta Neuropathol.62:51–58.