JOURNAI OFVIROLOGY., Oct. 1973,p.919-930
CopyrightO 1973 American Society forMicrobiology
Vol. 12. No. 4
Printedin U.S.A.
Herpes Simplex
Virus
and Human
Cytomegalovirus
Replication
in
WI-38
Cells
I.
Sequence
of
Viral
Replication
JANET DUYCKINCK SMITH' AND ETIENNE DE HARVEN
Sloan-Kettering Institute for Cancer Research and Cornell University Graduate School of Medical Sciences,
New
York,New York 10021Received forpublication 26 April 1973
A comparison, under standardized conditions, of herpes simplex virus
(HSV) and human cytomegalovirus (CMV) revealeddifferences in viral
morphol-ogy, inthetimingof their infectiouscycles, and in several morphological events
during those cycles. Structuraldistinctionsbetween thetwoviruses included the
coatingofunenveloped cytoplasmic CMV capsids, but not those ofHSV, and a
variation in thestructureoftheircores. Since thetwo cycleswerecarried outin
thesame host cell strain under conditions ofone-stepgrowth (input multiplicity
= 10 PFU/cell), itwaspossibleto constructtime scaleslocating the majorevents
of each cycle. Comparison of thetwoshowed that HSVreplicated and released
progeny within 8 hpostinfection, whereas CMV required 4days. These results
correlated well with those ofconcurrent plaque assays. Within the longer CMV
cycle, most of the major events appeared retarded toa similar degree, and no
obvious limiting step in particle production could be identified. Distinctions
between thetwo cyclesincluded thefollowing: condensation of the chromatin in
HSV- but not CMV-infected cells; the greater tendency of HSV to produce
membranealterations; and theappearanceofcytoplasmicdense bodies in
CMV-but not HSV-infected cells. Identification of these differences even under
identical conditions of culture and infection strongly implies that they result from intrinsic differences in the nature of the viruses, and are not caused by variations in experimental conditions.
The members of theherpesvirus groupshare
common features including a similar
morphol-ogy, a nuclear site of capsid assembly, and similar modes of maturation and release, yet differ significantly in otherparameterssuch as
the length of the replication cycle, the host range, and theyield ofinfectious cell-freevirus (12, 17, 32). To date many of the studies of individual herpesviruses have been carried out
under different conditions in different host cell systems, making it difficult to determine
whether observed dissimilaritieswereduetothe intrinsic nature ofthe viruses or to the
varia-tions in the systems employed. We report here
on acomparison ofthereplicative cycles of two
herpesviruses carried out under standardized conditions in the same host cell system, using equivalent multiplicities ofinfection. To estab-lish a time scale for each cycle, a multiplicity
lPresent address: Department of Anatomy, The Medical
CollegeofPennsylvania. Philadelphia. Pa. 19144.
919
sufficient to produce a synchronous one-step growth cycle wasused.
MATERIALS AND METHODS
Cells. WI-38cellswereobtainedfromthe American
Type Culture Collection in the 16th passage. They
were grown as stationary monolayers in minimal
essentialmedium (MEM) with Earle's saltsplus10%
fetal calfserum (FCS), 7.7 mM NaHCO3, 100jgof
streptomycinperml, and 100Uofpotassium
penicil-lin G per ml. Tests for mycoplasma contamination
were negative.
Virus stocks. Herpes simplex virus(HSV), strain
HF (8)wasreceivedfromthe AmericanType Culture
Collection, as was human cytomegalovirus (CMV),
strain AD-169 (28). Both were free of mycoplasma.
Virus-infected cells were maintained on MEM with 2%agammacalfserum(MEM2), and were harvested
when85 to 90%showedcytopathic effects.
Mechani-calhomogenizationwasfollowedbycentrifugation at
500 x g for 10 minand filtration through prewashed
Milliporefilters with pore size of 0.8 and then 0.45jm
(6).Clarifiedsuspensions ofHSVwerefrozen
on November 10, 2019 by guest
http://jvi.asm.org/
SMITH AND DE HARVEN
ately, whereas those ofCMV were concentrated
10-to20-fold by negativepressureultrafiltration through
dialysis tubing(25). Thefrozen stocksweretitered by
plaque assay.
Plaque assay. Cell-free virus wasassayed bythe
procedures of Wentworth and French for both HSV
(29) and CMV(30), except that CMVwas dilutedin
medium lacking bicarbonate, and fixed monolayers
werestained with 1%aqueousmethyleneblue.
Infection of cell monolayers. Preconfluent
mono-layerswereinfected with0.1 ml/plateofcellfree virus
ata multiplicityof 10PFU/cell.After adsorptionfor
90minat37C and5%CO2, thecultureswerewashed
onceand incubated in 2 ml offresh MEM 2.
HSV-infected samplesweretakenat0, 2, 3,4, 5,6, 8, 10, 12,
and24 hpostinfection, with uninfected controlsat0
and 24 h. Asample of medium from each plate was
frozen and subsequently titered for released virus.
CMV-infectedcellswerefixedat12-h intervalsover a
periodof 8days,withuninfected controlsat0 and 7.5
days. Duplicate plates were takenat eachsampling
time, onefor titration of released and cell-associated
virusand the other for electron microscopy.
Electron microscopy. The monolayers werefixed
and embeddedin situbyamodification of the method
of Brinkley et al. (3). Sections were stained with
uranylacetate and lead citrate, coated withcarbon,
and examined in a modified Siemens 1A electron
microscope. The main modification was a shorter
objectivelens focal distance(H. Armbruster,Siemens
Inc., Iselin, N.J.).
Fornegative staining,unfixed viralspecimenswere
adsorbedto grids covered bythin carbon films, and
then stained with 2% aqueous phosphotungstic acid
(PTA), pH4.8.
RESULTS
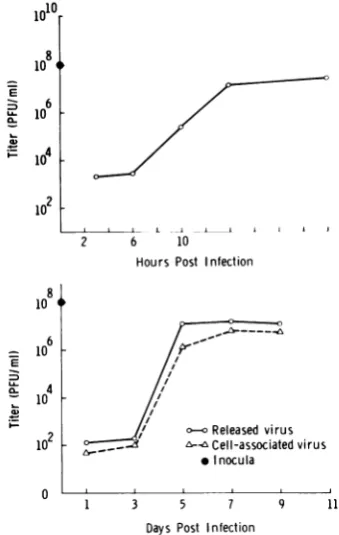
Virusgrowthcurves. The growth curvesfor
thecycles studied by electron microscopy (Fig. 1) indicate that, as expected with an input
multiplicity of 10 PFU/cell, conditions of
one-step infectionwere achieved with bothviruses.
This fact was independently confirmed by
in-fectious center assays.
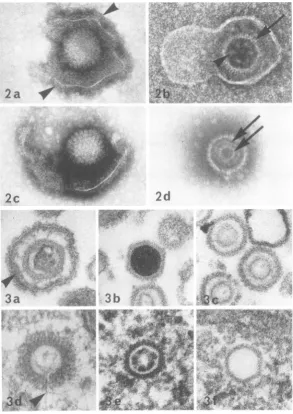
Viralmorphology. The major structural dif-ferences between the two viruses were as
fol-lows: (i) In negatively stained samples, the
cores ofHSV appeared to becontinuous
struc-tureswithawall of uniformthickness (Fig. 2d),
whereas those ofCMV resembledaggregates of globular subunits (Fig. 2b). This distinction in
corestructure wasalso evidentinthinsections,
where CMV but not HSV cores had a beaded appearanceincross section(Fig. 3c, e). Despite a recent report on herpes simplex morphology
(9), we were unable tofind any evidence indi-cating a toroidal structure for the core. (ii)
Unenveloped CMV capsids located free in the
cytoplasm were coated by a layer of fibrillar
material applied directly to the exterior of the capsid (Fig. 3d), whereas free cytoplasmic
cap-sids of HSV werenever coated (Fig. 3f). Coats
were never present on those capsids ofHSVor
CMV located within intact nuclei (Fig. 3e). Other features of viral morphology, including the variety of structural types usually observed with herpesviruses (25), areillustratedinFig. 2
and 3.
Comparison of paired HSV and CMV means, using Student's t test, showed no significant differences between any dimensions of the two (a = 0.1). In thin section, enveloped particles averaged 174.0 nm in diameter, capsids
aver-aged 106.4 nm, andcoresaveraged 64.3nm.The average thickness of the coat on unenveloped cytoplasmic CMV capsids was24.4 nm.
CMV replication cycle. By electron micros-copy the first visible sign of infection was the
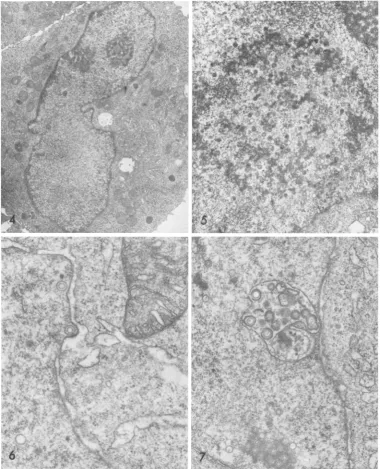
rounding up of individual uninucleate cells
between 12and24 hpostinfection. The reniform nuclei were displaced to the periphery of the cytoplasm, while a prominent Golgi apparatus
(Fig.4) occupied the centerofthe cell.
Less frequently cells fused to produce small
polykaryocytes usuallycontaining 3 to 4nuclei.
Thesewereoftenarranged in aringsurrounding
theGolgielements.
By day 3 capsids were assembling in the nucleus. At this stage they were all of one
morphological type, namely those containing
"empty" cores. Occasionally a capsid and core
appeared to be developing in synchrony. Free cores without a surrounding capsid were not observed. Capsids with dense cores began to appear between 3 and 3.5 days.
At 3.5 days mostofthe capsidswere closely
associated with a distinctiveskein-like nuclear
inclusion (Fig. 5), although smaller numbers
were found in the clear zone of nucleoplasm which characteristically separated these
inclu-sionsfromthe nuclearmembrane.
Envelopmentatthenuclear membranebegan between3and3.5days.
Capsids
ofall morpho-logicaltypes buddedthrough the innernuclearmembrane, either directly into the perinuclear cisterna (Fig. 6), or into sac-like membranous invaginations commonly found in the nucleo-plasm (Fig. 7). Rupture of the nuclear
mem-brane was not observed attheseearly stages. In theperiodbetween2and 3days postinfec-tion, there was a transient increase in the number of 55- to 60-nm intranuclear
granules
(Fig. 8, 9). Although previously reported
by
others (23) and interpreted as
possible
viralnucleoids, these seem morecloselyto resemble theperichromatingranulesofnormal
cells,
both920 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HSV AND CMV REPLICATION IN WI-38 CELLS
in size and in the presence of a clear area or
"halo" separating many ofthem from hetero-chromatin.
From theperinuclearcisterna, enveloped par-ticles usually moved into the cytoplasm within smooth vacuoles, formed by the evagination of the outer nuclear membrane. Nonlytic release of viruses within vacuoles occurred when the vacuolar membrane fused with the
plasma-lemma, thereby extruding the enveloped parti-cle.Altematively, particles could leave the
peri-nuclear cisterna via the channels of the rough
endoplasmic reticulum (Fig. 10), and could
travel through these channels for variable dis-tances. Since no continuities were ever found
between the rough endoplasmic reticulum and theplasmalemma, theviruses within the
reticu-lum must exit from the cell by a discontinuous route involving intermediate vacuoles.
The effect of CMV on cellular membranes was minimal. The nuclear membrane redu-plication which is characteristic of herpes-viruses occurred only to a limited extent (Fig. 11). Reduplication seldom involved more than
one extra membrane pair or more than a short section of the nuclearmembrane. Similarly, cell fusion was not common, and those
polykaryo-cytes observed rarely had more than 3 or 4
nuclei. In the cytoplasm there was a
prolifera-tion of
Golgi
vesiclesandstackedsaccules(Fig.
12), but these retained their normal
morpholo-gies.
Within the nuclei there was no noticeable
condensation or margination of the chromatin either at this or later stages of the infection.
Despite the apparently intact nuclear
mem-branes, unenveloped capsids were routinely
ob-served in the cytoplasm beginning at 4 days.
These capsids, all of which were coated, were mostnumerous in theimmediate vicinity of the nucleus and in theGolgiregion (Fig. 12e). In the
Golgi
zone many budded into a variety ofcytoplasmic vacuoles including multivesicular bodies,
phagolysome-like
vacuoles, and a char-acteristic small, clear vacuole with acrescent-shapedcrosssection(Figs. 12a-c).As aresult of this process the capsids simultaneously
ac-quired an envelope and became enclosed in a
surrounding vacuole, making them virtually identical to viruses enveloped at the nuclear membrane.
Concurrent with capsid assembly and
en-velopment, a homogenous electron-dense mate-rial began to accumulate in the cytoplasm, especially in the Golgi region (Fig. 12e). It
appeared first in small, scattered foci which lacked a limiting membrane. These retained
iolO
10f1
10 4
1o6~ L-
0-
1-10
102
k10oi
2 6 10
HoursPost Infection
16
~
U-a- 104
102
- Released virusn-n,Cell-associated virus
* nocula
1 3 5 7 9 11
0
Days Post Infection
FIG. 1. One-step growth curves showing the titers
of released virus for HSV (upper curve), and both
released and cell-associated virus for CMV (lower
curve).
regular spherical outlines while gradually in-creasing in size. By day 5 many foci were acquiring anenvelopebybuddingintothe same types ofvacuolesas CMV capsids (Fig. 12d, e).
Individual vacuoles often contained a mixed populationofboththeseenvelopeddense bodies and enveloped CMV. The dense bodies were
released from the cell in the same manner as virus.
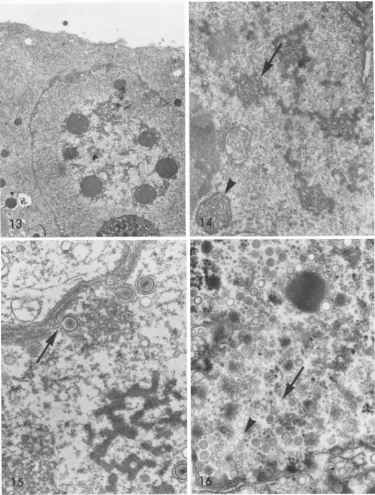
In later stages, a variety of distinctive
nu-clear inclusions were observed
(Fig.
13, 14).These may represent sequential stages in a
gen-eral compaction or condensation ofthe
skein-like inclusions described above.
Cell lysis commonly occurred at 7 to8 days postinfection.
HSV replication cycle. The most obvious dissimilarity between the cycles of HSV and
CMV was in the length of time necessary for
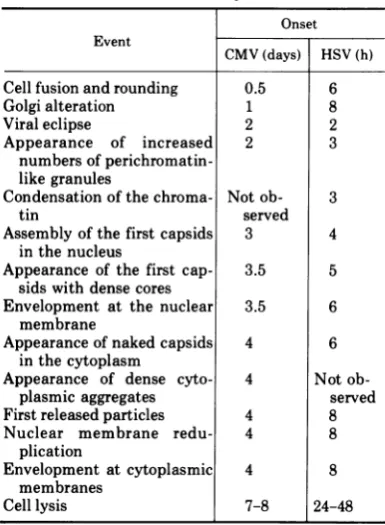
replication (Fig. 1). A comparison of the timing ofthemajor events in the two cycles is shown in Table 1.
Although the replication times differ signifi-cantly, most of the major events occur in the same sequence in both cycles. The exceptions 921
VOL.12,1973
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.495.262.432.74.342.2]SMITH AND DE HARVEN
2a
1
'IO. 1~ .
2--b
t.:
2d
2c
3a
3b
(I
--W~~~~~~~~~~.f
. .;F.
. , r I
*i.s
;*
c
FIG. 2. Negatively stained HSV and CMV particles. (a) Short 12- to 15-nm projections (arrowheads)
sometimes seen on the outer surface of enveloped particles. HSV. (b) Enveloped CMV with individual
capsomeresvisibleattheperiphery of the capsid (arrow). The core appears as an aggregate of globular subunits
(arrowhead),sixormoreperipheral and one central. (c) HSVparticle showing the hollow polygonal shape of the
capsomeres seen end-on. (d)AnHSV capsid containing a central core made up of two continuous concentric
structures (arrows). (A magnification ofx127,000 was used for a-d.)
FIG. 3. HSV and CMV particlesinthinsection. (a) Enveloped HSV. Note the dense material marginated
against the inner surface of the envelope (arrowhead), the capsid, and an apparently ruptured core of intermediate electron density. (b) An unsual HSV capsid completely filled by the electron dense material
thought to represent viral nucleoprotein. (c)HSV capsids containing "empty" cores. Note that as in Fig. 2d, the
wallof thecore seemscomposed oftwolayers, theinner here appearing slightly more electron opaque. (d) An
unenveloped, cytoplasmic CMV capsid, with fibrillar coat. Traces ofa coreremain intheinterior,along with an
unknown materialforminganunusual tail-like structure (arrowhead). (e) ACMV capsid within the nucleus.
Note the absenceof any coat and the beaded appearance of the core. (f)A cytoplasmic HSV capsid lacking a
core entirely and which, like all HSV capsids, nuclear or cytoplasmic, is not coated. (A magnification of
approximately x 127,000 wasusedfora-f.)
922 J. VIROL.
;.s
:4.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.495.112.406.78.493.2]HSV AND CMV REPLICATION IN WI-38 CELLS
r
*-.
:,.4;,;,>;:>s'
'II. *1
f '0
:
*'^; - ^
wf ...
.
b ,# F '. s
>,,@ .1 :, ,\
tr/) fE.
( "-e
.s , Z.l, 's. f
es
FY
t.ts t ,
'--^ - -X}t* \ ' I_.j%i,.
+lf -- s Kbi\t r
., :;
.. ..
-v
<. .. s
87
-.1 N I
.: "..I
.4'.
4
6
*''
'v{'4
*'*
'
'.;
a
(_
.:S
s le'% ' j -v~~~%
FIG. 4. A rounded cell 12h after CMVinfection. To the right of the reniform nucleus is aregion ofthe
cytoplasm where the accumulationand proliferationofGolgi elements have begun. Magnification x6,700.
FIG. 5. A typical nuclearinclusion3.5daysafter CMV infection.Itis composedofirregularanastomosing
strandsof electron dense materialand is in close proximity to mostof thedeveloping capsids.Magnification
x13,000.
FIG. 6. Asingle CMVcapsid acquiringanenvelope by buddingthroughthe inner nuclear membrane intothe
perinuclearcisterna. Three and one-halfdays postinfection. Magnification x37,000.
FIG. 7. CMVcapsidenvelopmentby buddingintoamembrane-boundedsacwhich ispresumablycontinuous
withtheperinuclearcisterna. Fourdays postinfection. Magnificationx25,500.
VOL.12,1973 923
-;.r,
.11
fz
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.495.54.435.80.550.2]SMITH AND DE HARVEN
pi.
., . . ,
.49
144
. X
x
r>* ;
_ t.;
Ab.. ;
'q-,.*
S
!
~ rf
rAr
AA'tf.
4
:V
k .r*
_,p
1||
f0,,Y.
9FIG. 8. A large accumulation of "perichromatin-like" granules inanHSV-infected nucleus. The granules occur in association with heterochromatin and occasionally near characteristic granular aggregates. 2 h postinfection. Magnification x36,000.
FIG. 9. "Perichromatin-like"granules inanucleus 2days after infection with CMV. Note the clear regionor
"halo"separating them from the surrounding heterochromatin. Magnification x38,000.
FIG. 10. Atangential section ofanenveloped particle in theperinuclear cisternanearthepoint of continuity
with therough endoplasmic reticulum. HSV, 8hpostinfection. Magnification x75,000.
FIG. 11. At severalpoints (arrowheads) short segments of reduplicated membranes separate the nucleus
(lower right) from the cytoplasm ofacell 4days after infectionwith CMV.Magnification x48,000.
924 J.VIROL.
I
I _
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.495.67.447.74.552.2].12.a.
t.
ze rr--n x
12b
12c.-,¢;
r,,^
A..
... ,
..I
¢2r -?
fr..'.:."
*; 1
4.
,t ir t*C., g 2,
*Xso;
sr,
f ib.sOX
Xs sS..
ffi X
#,,.. r e
,.;
Xt_
.. i'.I ~~~~.
-. i ..
.-, x
A-z
'-
1>sr,
A*. ;iXZ [.4: A
PYtS'
FIG. 12. Cytoplasmic envelopmentinCMVinfection. (a-c)NakedCMVcapsids budding throughonesideof
acytoplasmicvacuole. Note that inbudding particlesthe thicknessofthecapsidcoatissignificantlyreduced.
Magnification approximately x100,000. (d)A typical dense bodyundergoing cytoplasmic envelopment in a mannersimilartoviralcapsids. Six dayspostinfection. Magnification x32,000. (e)Anareaof cytoplasmrichin
Golgi elements and containing several unenveloped, coated capsids (arrows, upper left), one of which
(arrowhead)isbuddingintoasmallcytoplasmicvacuole.Manydensebodies,bothenvelopedandunenveloped
(arrow,lowerright)arealso present. Four andone-halfdays postinfection. Magnification x24,000.
925
~~rv
12 d
--*4
.4
.*W
so ,@.
, v ;t
> e
.- i: vi_
s 7 .t
,
*!^
.
._-
c :o
O
' .. .:1:
i 'W
4
..
.4 It,
i.
12
e
s.,
.".1.4
-V.
%. i!
ll.,.-. I
i .
-. , .1.
I'l.l-*
on November 10, 2019 by guest
http://jvi.asm.org/
SMITH AND DE HARVEN
TABLE 1. Comparison of timing of major events in
HSV and CMV cycles.
Onset Event
CMV(days) HSV(h)
Cell fusion androunding 0.5 6
Golgialteration 1 8
Viral eclipse 2 2
Appearance of increased 2 3
numbersof
perichromatin-like granules
Condensationofthe chroma- Not ob- 3
tin served
Assembly of the first capsids 3 4
inthenucleus
Appearance of the first cap- 3.5 5
sids with dense cores
Envelopment at the nuclear 3.5 6
membrane
Appearance of naked capsids 4 6
inthecytoplasm
Appearance of dense cyto- 4 Not
ob-plasmic aggregates served
Firstreleased particles 4 8
Nuclear membrane redu- 4 8
plication
Envelopment at cytoplasmic 4 8
membranes
Celllysis 7-8 24-48
arethose reflecting thereorganization of
cellu-lar architecture. These, including cell fusion, cellrounding, and the proliferation and consoli-dation of theGolgi elements,occurwithinhours
after infection by either virus, and therefore
relativelyearlier in the longer cycle of CMV. Inadditiontothesetemporal variations there
werequalitative differences whichmaybe
sum-marized asfollows:
(i) The naked cytoplasmic capsids of HSV,
unlike thoseofCMV, were nevercoated.
(ii) The cytoplasmic dense bodies (Fig. 12e) characteristic of CMV infectionwerenotfound
inHSV-infected cells.
(iii) The skein-like nuclear inclusions of
CMV-infected cells were not observed after
infection with HSV. Nuclear inclusions were
not commonuntil late inthereplicationcycle,
when two characteristic types appeared (Fig. 15, 16).
(iv)By3h postinfection with HSV therewas a marked condensation and marginationof the
chromatin, which persistedthroughout replica-tion (Fig. 17, 18). Such condensation was
en-tirely absentwith CMV.
(iv) By3 h postinfectionwithHSV therewas
a marked condensation and margination of the
chromatin, which persistedthrough replication (Fig. 17, 18). Such condensation was entirely
absent with CMV.
(v) Extensive membrane changes followed
infection bythis strain ofHSV. These included
the following: (i) cell fusion with formation of
large polykaryocytes often containing 12 or
more nuclei; (ii) reduplication of the nuclear membrane, with two or more extra membrane pairs over much of the periphery of thehighly lobulated nucleus; and (iii) proliferation and
extensive morphological alterationof the
mem-branous elements in the Golgi region. With respect to this last point, by 8 h postinfection
the typical vesicles and saccules of the Golgi
had been almost entirely replaced by two new
elements, namely, concentric cisternae
bounded by smooth membranes (Fig. 18) and
large smooth-surfaced vacuoles arranged in
characteristic clusters (Fig. 17, 18).
Cytoplas-mic envelopment ofnaked HSV capsids often
occurred attheselarge vacuoles.
DISCUSSION
Since both viral cycles described above were carried out under standardized conditions, the observed differences between the two presum-ably reflectdifferencesinthe geneticmakeupof the viruses rather than variation inthe experi-mental system.
To the extent that other strains of HSV or CMV are related to those used here, they should showsimilar distinctive patterns of replication when compared inthe same cell system. Indeed, earlier investigations with other strains have shown similar patternswith regard to chroma-tinmargination andcondensation byHSV(15); lack of condensation with CMV (5, 10, 13, 18); extensive membrane changes with HSV, i.e., eitherpolykaryocytosisorextensive cell round-ing (12, 16); and onlyminimaleffects with CMV (13, 18). Nor are the patterns ofreplicationwe observed limited to laboratory strains. Thus, extensive polykaryocytosis has been observed notonly with adaptedstrains ofHSV, but also with recently isolated virus (1, 7, 22), andeven inbiopsy material(2, 20,21).Asimilar concord-ance between many of the major effects of an adapted strain and afreshisolatewasreported
recentlyforCMV (11).
Withregardtoviralmorphology,weobserved
many of the same featuresreportedby othersfor
HSV (15, 31) and CMV (13, 26). The
double-walledstructure oftheHSVcoreresembles that describedfortheLucke virus (27), whereas the beaded orglobularappearance of theCMVcore has been illustrated by a number ofpublished micrographs (5, 10, 11, 13). We did not find evidence witheither HSVorCMV foratoroidal structure ofthe core, suchasreportedrecently
for HSV(9).Thiswas true eveninlatestages of
926 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.495.58.251.89.352.2]0
' X
-;1I-e
4'-*
I
4.
13
.'
2.
.\.
.
4:
#
w
el
ti/eA~-t;t
.t--<-7
FIG. 13. A late CMV inclusioncomposed of multiple roundmasses,associatedattheirperiphery with viral
capsids andwith theremnantsof theearlierskein-like inclusions,andoccupyingan areaof reduced electron
opacity inthecenterof thenucleus. A rim of nucleoplasmseparatestheinclusionfromthe nuclear membrane andfromthesegregated nucleolusatthe lowerright. Six dayspostinfection. Magnification x6,300.
FIG. 14. Several inclusionsresembling granulesorshortfibrils (arrow) andone morecompactfibrillar form
(arrowhead) are closely associated with the remnants of the skein-like CMV nuclear inclusion. Six days
postinfection.Magnification x18,000.
FIG. 15. A late HSV inclusion ispresent in the lowerright. Note also the extensive reduplication ofthe
nuclear membrane as well as the viruses acquiring an envelope at their inner face (arrow). Twelve hours
postinfection.Magnification x34,000.
FIG. 16. Around inclusionbody (upperright)commoninlateHSVinfection.Thesewerescatteredsinglyin
thenucleoplasmrather thaningroups,andshowednotraceofafibrillarsubstructure. Note also thenumerous
small, ring-likestructuresfreeinthenucleoplasm (arrow). Althoughsmaller thannormal viralcores,several
appearto besurrounded by typical capsids (arrowhead), suggesting they may be theresultofaberrantcore
assembly in late infection. Twenty-fourhourspostinfection. Magnification x36,000. 927
F.,)
ion November 10, 2019 by guest
http://jvi.asm.org/
[image:9.495.55.431.38.534.2]p
'a
I
.1
S
,
17
J.
-t
A.
,,Nki. - ic~ AO
FIG. 17. An HSV-infected cell showing long stretches of nuclear membrane reduplication as well as
condensation and margination of the chromatin intoseveral large masses. Within the cytoplasm the typical
Golgielements havebeenreplaced by large, apparentlyemptyvacuoles; andnumerousunenvelopedcapsids,
alllackingcoats,arepresent. Tenhpostinfection. Magnification x75,500.
FIG. 18. AnHSV-infected cell in which the typical Golgi membranes have been replaced by concentric
arraysof membrane-boundedcisternae(arrow), andlarge vacuoles.Oftennakedcytoplasmic capsids became
enveloped by buddinginto the latter (inset). Ten h postinfection. Magnification x14,000; inset, x21,000.
928
'$.
.-0 h;0
,:.v., .
N
i;
s
s*S-t Se
v
.. *
t._....-r
b'v. i''w'
% ..
*q!\t;*;...<
.<. ..... "
w..
$,_
.s @ w
;r2+ R iF
*S .;-
*-r;-.wY . ;
'A
N
..-, I
1,, ..i. .I;illw.
:
x .I ,,
,. e
I ..
.1.
wt
.. k '.
O..:
;#K.
I
V
t
'.-i
"I
I
4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.495.66.458.43.620.2]HSV AND CMV REPLICATION IN WI-38 CELLS
infection when bizarre core forms often appear
(15).
In a system identical toour own, i.e., CMV
strain AD169 growing in WI-38 cells, other
investigators were not able to document capsid
envelopment at the nuclear membrane (11). However, weregularly observed thefullprocess of CMV envelopment at the nucleus, as have
other workers (13).
Despite the observation of envelopment at the nuclear membrane, other capsids of both
HSV and CMV are consistently found unen-veloped in the cytoplasm. Unenunen-veloped
cyto-plasmic CMV capsids were coated, whereas those ofHSVwere not.These results differ from
those in an early report (5) which described
CMV capsids in monkey kidney cells as
un-coated. We found coating tobe a strictly
cyto-plasmicphenomenon, in agreement with the
ob-servation ofStackpole (27) forLuckevirus.
Our studies revealed only one method of nonlytic viral release, namely,fusionbetween a
vacuole carrying an enveloped virus and the
plasmalemma, a process which has been aptly
described as reverse phagocytosis (14). Other investigators (19) have described a tubular
system continuous at one end with the
perinu-clear cisterna, and with theextracellular space at the other, which might serve as a direct
avenueof viral release inHSV-infected HEp-2 cells. Despite numerous attempts to observe such tubules by through sectioning of in situ
embedded monolayers in a variety of selected orientations, we saw no evidence for them in
eitherHSV-orCMV-infected cells.
Wedid observe theformation of
cytoplasmic
densebodiesinCMV-infectedcells inamanner
reported by others (10, 11, 13, 18).
However,
on the basis of
cytochemical
datareported
else-where(manuscriptin preparation), we feelthat the common assumption that these representlysosomesisprobablyerroneous.
With regard to the timing ofthe two cycles, our results agree well with those of previous authors for HSV (15, 24) and with some of the more limited data available for CMV (13, 18).
However, we were not able to identify any
limiting steps in CMV reproduction which
might explain the much longer cycle time. As wehaveshown(manuscript in preparation), the limitation does not appear to involve viral
adsorptionorpenetration, since capsids of HSV and CMV entering the cell from the inoculum both reach the periphery of the nucleus in approximately the same time.
Finally, it should be pointed out that
al-though we have defined several criteria for
discriminating between HSV and CMV
infec-tions in this system, we do not as yet
under-stand the biochemical basis for any of these
distinctions. Especially intriguing isthefailure
of CMV to cause condensation of host cell
chromatin, since this may reflect an effect on host cell macromolecular synthesis which is
different from that of most herpesviruses (12, 17).High-titerCMVinocula, which can now be obtained in a variety of ways (4, 10, 25) should makethis and othersimilarquestionsaccessible
to investigation bybiochemical methods.
ACKNOWLEDGMENTS
We aregratefultoChristopherStackpoleforreading and commentingonthismanuscript.Inaddition,we areindebted toMark Van Boeckelforexcellentphotographicalassistance
andtoDorothy Saltzer for the experttypingofthe
manu-script.
Thesedata were partof aPh.D. thesispresentedby J. D. S. to the Cornell University Graduate School of Medical Sciences.
This investigationwassupported byPublic Health Service Grant CA-08748 from the NationalCancer Institute andby
the Health Research Council of the City of New York,
Contract1-325.
LITERATURE CITED
1. Beale,A.J., and H. C. Hair.1956.Rapiddiagnosisofthe herpetic variety ofKaposi's varicelliform eruptionby
tissueculturemethods. Can. Med. Ass. J.74:443-448. 2. Blank, H., C. F. Burgoon, G. D. Baldridge, P. L.
McCarthy, and F.Urbach.1951.Cytologicalsmearsin diagnosis ofherpessimples, herpeszosterand varicella.
J. Amer.Med. Ass. 146:1410-1412.
3. Brinkley, B.R.,P.Murphy,and L. C. Richardson. 1967.
Procedure forembedding in situ selected cells cultured invitro. J. Cell Biol. 35:279-283.
4. Chambers,R.W.,J. A.Rose,A.S.Rabson,H. E.Bond,
and W.T. Hall. 1971.Propagationandpurificationof
high-titer human cytomegalovirus. Appl. Microbiol. 22:914-918.
5. Dalton,A.I., and R.A.Manaker.1967.Thecomparison of virus particles associated with Burkitt lymphoma
with other herpes-like viruses. In Carcinogenesis: a broad critique, p. 59-90. The Williams& Wilkins Co., Baltimore.
6. deHarven, E., and C. Friend.1965.Originofthe viremia
in murineleukemia. Nat.Cancer Inst.Monogr.no.22, p. 125-134.
7. Doane,F.,A.J.Rhodes, andH.L.Ormsby. 1955.Tissue culturetechniquesinthestudyofherpeticinfections of the eye.Amer.J.Ophthalmol.40:189-193.
8. Flexner, S., and H. L. Amoss. 1925. Contributiontothe
pathologyofexperimentalvirusencephalitis. II. Her-petic strainsofencephalitogenic virus. J. Exp. Med. 41:233-244.
9. Furlong,D., H. Swift, and B. Roizman. 1972.
Arrange-ment ofherpesvirusdeoxyribonucleic acidinthecore. J. Virol. 10:1071-1074.
10. Iwasaki, Y.,T.Furukawa,S.Plotkin, andH.Koprowski.
1973.Ultrastructuralstudyonthe sequenceof human
cytomegalovirusinfection inhumandiploid cells. Arch. GesamteVirusforsch.40:311-324.
11. Kanich, R. E., and J. E. Craighead. 1972. Human cytomegalovirus infection of cultured fibroblasts. II. Viral replicative sequence of awild and an adapted
strain.Lab.Invest. 27:273-282.
12. Kaplan, A. S. 1969. Herpes simplex and pseudorabies
929
VOL.12,1973
on November 10, 2019 by guest
http://jvi.asm.org/
SMITH AND DE HARVEN
viruses. Virol. Monogr. no. 5, Springer-Verlag, New
York.
13. McGavran, M. H., and M. G. Smith. 1965. Ultrastruc-tural, cytochemical andmicrochemicalobservationson
cytomegalovirus (salivary gland virus) infection of
human cells in tissue culture. Exp. Mol. Pathol. 4:1-10.
14. Morgan, C., H. M. Rose, M. Holden, and E.P. Jones. 1959.Electron microscopic observationsonthe
devel-opment of herpes simplex virus. J. Exp. Med.
110:643-656.
15. Nii, S., C. Morgan, and H. M. Rose. 1968. Electron microscopy ofherpes simplex virus. II. Sequence of
development. J. Virol. 2:517-536.
16. Roizman, B. 1971.Herpesviruses, membranes, andthe
socialbehavior of infected cells. p.37-77.In M.
San-ders and M. Schaeffer (ed.), Viruses affecting man
andanimals.W. H.Green, St. Louis.
17. Roizman, B. 1969. The herpesviruses-a biochemical definitionofthegroup.InCurrenttopicsin
microbiol-ogyandimmunology, vol. 49,pp. 1-79. Springer-Ver-lag, New York.
18. Ruebner, B. H., T.Hirano, R. Slusser, J. Osborn, and D. N.Medearis, Jr.1965. Human cytomegalovirus infec-tion.Electron microscopic andhistochemical changes in cultures of human fibroblasts. Amer. J. Pathol. 46:477-496.
19. Schwartz, J., and B. Roizman. 1969. Concerning the
egress of herpes simplex virus from infected cells:
electron and light microscope observations. Virology
38:42-49.
20.Scott, T. F. M. 1964. Herpes simplex. Diagnostic
proce-dures for viruses and rickettsial diseases, 3rded., p.
381-412. InE.H. Lennette andN. J.Schmidt (ed.),
pp. 381-412. American Public Health Ass., Inc., N.Y.
21. Scott,T.F. M. 1965.Theherpes virusgroup.Viral and
rickettsial infections ofman,4thed.,p.892-914.In I.
Tamm and F. L. Horsfall (ed.). J. B. LippincottCo., Philadelphia.
22. Scott, T. F. M., and D. L. McLeod. 1959. Cellular
responses to infection with strainsofherpes simplex virus. Ann. N. Y.Acad. Sci. 81:118-128.
23. Shipkey, F. H., R. A. Erlandson, R. B. Bailey, V. I. Babcock, and C. M. Southam.1967.Virusbiographies. II. Growth of herpes simplexvirus in tissue culture. Exp. Mol. Pathol. 6:39-67.
24. Siminoff, P., and M. G. Menefee. 1966. Normal and 5-bromo-deoxyuridine inhibited development ofherpes
simplex virus. An electronmicroscope study. Exp. Cell Res. 44:241-255.
25. Smith, J. D., and E. de Harven. 1973.Concentrationof herpesviruses. J. Virol. 11:325-328.
26. Smith, K. O., and L. Rasmussen. 1963. Morphology of cytomegalovirus (salivary gland virus). J. Bacteriol. 85:1319-1325.
27. Stackpole, C. W. 1969. Herpes-type virusof thefrog renal adenocarcinoma. I. Virus development intumor
trans-plants maintained at low temperature. J. Virol. 4:75-93.
28. Weller, T. H., J. B.Hanshaw, and D'M.E.Scott. 1960. Serologic differentiation of viruses responsible for
cy-tomegalicinclusion disease.Virology 12:130-132.
29. Wentworth,B. B., and L. French. 1969.Plaqueassayof
Herpesuirus hominisonhumanembryonic fibroblasts.
Proc. Soc. Exp.Biol. Med.131:588-592.
30. Wentworth, B. B., and L. French. 1970.Plaqueassayof cytomegalovirus strains of human origin. Proc. Soc. Exp. Biol. Med. 135:253-258.
31.Wildy, P., W. C.Russell,and R. W. Horne. 1960. The
morphologyofherpesvirus.Virology12:204-222.
32. Wilner, B. I.1969. Aclassificationofthemajorgroupsof
human and other animal viruses, 4th ed. Burgess Publishing Co., Minneapolis.