Copyright©1969 AmericanSociety forMicrobiology PrintedinU.S.A.
Differential
Inhibition
of
Attachment
and
Eclipse
Activities of HeLa
Cells for Enteroviruses1
IHOR ZAJAC AND RICHARD L. CROWELL'
DepartmentofMicrobiology, JeffersonMedicalCollege,Philadelphia, Pennsylvania19107,and
LangbordVirusLaboratory,Departmentof Microbiology,HahnemannMedical
College,
Philadelphia, Pennsylvania19102
Receivedforpublication4February 1969
Receptor activities ofHeLa cells were evaluated for ability to both attach and eclipseenteroviruses after exposureofcells toacid or heat. A modified procedure ofacid(pH1.5) elution of cell-associatedvirus, ascompared with otherprocedures, provided a general method for the optimal recovery of receptor-bound entero-viruses. With this procedure,eclipse ofvirusoperationallywas considered to be that amountofvirusinfectivity whichwasdeterminedinitially to becell-associated and whichwas notdissociable from thecells.HeLacellskilledbyheatingat56 C for 30 min could not attach or eclipse poliovirus TI, but they attached and eclipsed coxsackieviruses B1 and B3, and they attached echovirus 6 but did not eclipse it. HeLacells treatedatpH2.5for 10 minat 2 C could not attach or eclipsepoliovirus TI, but they attachedcoxsackievirusesB1 and B3 and echovirus 6, although these viruseswere noteclipsed. These resultsshowed that, within the operationaldefinition ofviruseclipse, the eclipse activity ofHeLa cells for some viruses can beirreversibly inactivated withoutimpairing the activityof thereceptorsforattachingthese viruses. The dataprovided additional evidence thatHeLa cells possess specific receptors for the different enteroviruses.
Initiation of infection of
susceptible
cellsby
enteroviruses has been shown to be
dependent
uponthe presenceof
specific
receptors to attach virus (14, 21, 22) and anefficient mechanism for viral eclipse (5). Extensivestudies
have beenperformed
in an attempt to characterizeentero-virusreceptorsby theuseof cellularhomogenates (8, 9,
21),
cellextracts(16, 19,
21),
redblood cells(18),
andliving
HeLa cells(1-4, 10, 23, 24)
as asource
of
receptoractivity.
These studies haveemphasized
the diverse characteristics of thecellu-larreceptors
responsible
for attachment ofpolio-viruses,
coxsackieviruses,
and echoviruses. This paper describes thestability
of the different enterovirus receptors of HeLa cells to acid and heat.The receptor activities of treatedcells wereevaluated for
capacity
both to attach and toeclipse virus.Inthese
studies,
thetermeclipse (7)
wasused
operationally
todenote thatamountofvirus
infectivity
whichwasdeterminedinitially
tobecell-associated and whichwas not dissociable fromthecellsbytreatmentwith
glycine
buffer(6)
atpH 1.5.
'Presented inpartatthe 66thAnnualMeetingof theAmerican Society forMicrobiology,LosAngeles,Calif., May,1966.
2ResearchCareerDevelopment Awardgranteeof the Public HealthService,National InstituteofAllergyandInfectious Dis-eases.
MATERIALS AND METHODS
HeLa cells. Cells designated HeLa (JJH) were culturedasmonolayers in growthmediumcontaining
calf serum (GMCaSlO), asdescribedelsewhere (23). Virus strains. Coxsackieviruses Bl and B3,
polio-virusTl,andthe method of plaque assay used in this
investigation were described previously (4). Echo-virus 6, kindly supplied by M. Klein of Temple
University Medical School, Philadelphia, Pa., was
propagated and assayed in a fashion similar to the other enteroviruses.
Determination of receptor activity to attach and eclipse enteroviruses. Determination of the receptor
activityof HeLa cells forattachingviruseswascarried out at 37 Cor at 2C, as described elsewhere (23).
Theamountof virus whichwasattachedtocellsat a
given time was determined by the difference between theamounts of free virus found and theinputvirus. In addition, in experiments conducted at 2 C, the
amountof virus whichwasattachedtocellswas deter-mined directly byelution of virus from the cells by use ofglycine buffer at pH 1.5. A close correlation
wasfound between thetwo methods used for deter-mining the amount of attached virus provided the system waskept cold. Determinations ofthe ability
of HeLa cellstoeclipseviruswerecarriedout at37C. Eclipsed virus (7) wasconsidered to be thatamount
of virusinfectivitywhichwasdeterminedinitiallytobe cell-associated (by difference between free virus and
inputvirus) and whichwas not dissociable from the
422
on November 11, 2019 by guest
http://jvi.asm.org/
cell by treatment with 0.05 M glycine buffer (6)
ad-justedtopH 1.5.
Preparation of cell-associated virus (CAV). A
sample of5 X 107 washed HeLa cellswascentrifuged
at225 X gfor 5 min, andthe sedimentedcellswere resuspended in 5 ml of virus at 2 C. [Input virus
multiplicity usually was approximately 10 plaque-forming units (PFU) forcoxsackievirus B3, 50 PFU
for coxsackievirusBi,and 100 PFU forpoliovirusTi and echovirus 6.] The cell-virus mixturewasincubated
at2 C for1 hr withfrequent agitation. Virus
attach-ment was stopped by diluting the cell-virus mixture
10-fold with cold balanced salt solution (BSS)-CaS3
solution and centrifuging thecells at225 X gfor 5
minat2 C. Thecellswerereadied foruseby washing
them four more times with cold BSS-CaS3 solution
(50ml eachwash)toreduce freevirus. The CAVwas
determined by the acid reversal method described below.
Reactivationof CAVbyacidpH.Theprocedureof
Fenwick and Cooper wasmodified and used (6). A
known number of cells with associated virus was diluted 100-fold in cold (2 C) 0.05 M glycine buffer
(pH 1.5). Aftertreatmentfor 1 min, thesample was
diluted 10-fold further intophosphate-buffered saline
(PBS), devoid of calcium andmagnesium,containing 3% calf-serum (PBSA-CaS3), which was adjusted topH10.9sothat the finalpHof theviruspreparation
was neutral. Theamountoftotal virusrecovered, as
determined by plaque assay and corrected for the
amount of virus remaining free prior to acid treat-ment, was consideredto be theamount ofinfectious viruswhichwascell-associated and boundtoreceptors atthe time of acid treatment.Fromacomparison of
several methods used for dissociation of
coxsackie-viruses from cell receptors, this method was found
to be themosteffective (seeTable2).
Chemicals and enzymes.Allchemicalswere
analyt-ical reagent grade.Chymotrypsin (3 Xcrystalline)was purchasedfromWorthingtonBiochemicalCorp.,
Free-hold,N.J. Freshenzymesolutionwaspreparedforeach
experimentand usedatafinal concentration of1%. RESULTS
To study effectively the receptor activity of HeLa cells for attachment and eclipse of
entero-viruses, we considered it important that the
following preliminary conditions be met: (i) the
viruses should attach andremain at the cell
sur-face for extended periods of time; and (ii) a
method should be used that will measure most accurately theamount of CAVpriortoinitiation of viraleclipse.The use oflow temperature has proved to be the only efficient method for main-taining the attached viruses on the cell surface,
and, aspresented below, the method of acid
re-versalof CAVappeared tofulfillthesecond
con-dition. Accordingly,experimentswere performed
in an attempt to follow the distribution of the total amount ofinput virus addedto HeLacells afterincubation for1hrat2C.
Six separate experiments with coxsackievirus
B3andthreeseparateexperiments with poliovirus
TI were performed in which the total PFU of virus was determined in each of five consecutive wash fluids and in the CAV fraction (Table 1). The resultsshowed that the major portion of the input virus remained in the fluid phase. The find-ing of almost complete recovery of the input virus inoculum attested to the efficacy of acid pH for
recovery of total CAV under conditions which prevented viral eclipse.
Capacity of assorted reagentstodissociate CAV. Interest in the fate of cell-associated virus has produced many procedures fordissociating virus-cellcomplexes (6, 7, 12, 15, 18). These procedures were compared for their capacity to dissociate CAV under conditions which precluded viral eclipse. Earlier experiments had shown that the acid pH method used todissociatevirus-antibody complexes (11, 20) and CAV (6, 15) effectively released coxsackievirus B3 from the surface of HeLacells (3). Todetermine the pHwhichwould optimally reverse CAV at 2 C, a wide range of acidpH was tested.
Preparation of CAV by the use ofpoliovirus TI, echovirus 6, and coxsackieviruses Bi and B3
TABLE 1. Distributioni ofiniput virus in the cellular andfluidphases of the HeLa cell suspensions
after incubationforIhrat2C
Amt ofinput virus recoveredb Phase'
Coxsackie B3 Polio Ti
Wash 1 82 i 6 86 i 3
Wash2 3±-0.5 6± 3
Wash 3 <1 <1
Wash 4 <1 <1
Wash 5 <1 <1
CAV(pH1.5) 13 ± 4 9-± 5
CAV lost 1 <1
Total recovery 99 ± 10 101 i 3
a The amount of free virus remaining in the fluid phase of the cell suspension used to deter-mine CAV was <1 for bothcoxsackievirusB3 and
poliovirusTi.
IPresented as the mean from six experiments with coxsackievirus B3 and three experiments with poliovirus T1 with the standard error of the mean, respectively. Washed HeLa cells (5 X 107)
were suspended in 5 ml of virus at 2 C. Input
multiplicity ofviruswas 10 PFUand 100PFUfor coxsackievirus B3 and poliovirus TI, respectively. After 1 hr of incubation at 2 C, the cell-virus
suspension was diluted to 50 ml with BSS-CaS3, and centrifuged at225 X g; the supernatant fluid (wash no. 1) was assayed for virus infectivity. The cells were washed accordingly four more
times, and thefluidswere assayedforvirus. After
the fifth wash, the CAV was determined, as indicated in Materials and Methods.
on November 11, 2019 by guest
http://jvi.asm.org/
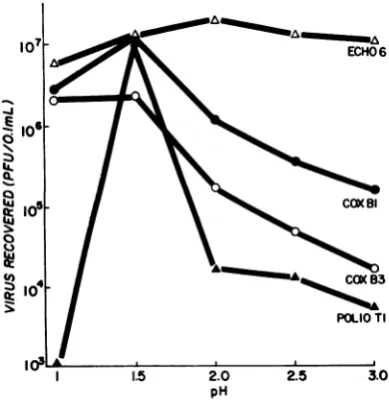
[image:2.494.247.440.347.486.2]was described in Materials and Methods. Cells with associated virus, kept at 2 C, were diluted 1:100 intocold0.05 M glycine bufferatdifferent pHvalues and held for1 minat2C,after which the pH wasbroughttoneutrality byafurther1:10 dilution into PBSA-CaS3. The cells were sedi-mented by centrifugation and the supernatant fluidswerecollectedandassayedtodetermine the amountof dissociated virus. The results of these experiments (Fig. 1) showed that optimal
re-covery of coxsackievirusesBi and B3 and
polio-virusTl occurredatpH1.5, whereas therecovery ofechovirus 6wasaccomplished
equally
wellatall pHvalues tested. Determination of virus stability inthemediumused showed that all of the viruses remainedcompletelystableatpH1.5at2Cforatleast 3 min. At pH 1.0,
poliovirus
Ti was in-activated within1min,andcoxsackievirusBi and echovirus6lost close to 50% of theirinfectivity,
whereas coxsackievirus B3 remained stable. The recovery ofCAVremained constant
irrespective
of whether the durationof the celltreatment at
pH
1.5at 2C wasfor30 sec orfor 5min. Treatment ofcellsfor 1minat2CatpH2.0or2.5 recovered only small amounts of the totalCAV.
However,
byincreasing the duration oftreatment,at
pH
2.0or2.5,to 5min,
progressive
elution of CAVwasfound to approach the
quantity
of virus disso-ciated atpH 1.5. It can beconcluded from these results and from those presented inTable 1 that, within the limits ofthe accuracy of theplaque
assay,pH1.5
yields
totalrecoveryofthe receptor-bound virus.In the next series ofexperiments,theefficacyto
7106
ILI \
a.0/ ^
[image:3.494.56.251.429.631.2]2.0 pH
FIG. 1. Effect of pHon the dissociation of entero-virusesfromHeLa cells.
elute CAV at pH 1.5, pH 2.5 or with 8 M urea, 6 M lithium chloride (LiCl), 4% sodiumdodecyl sul-fate (SDS), 1 % chymotrypsin, and alternate freezing and thawing was compared for each of the four viruses used previously. The cell-asso-ciated virus was prepared as described in Materials and Methods. The washed cells were taken up in a small volume of cold BSS-CaS3, and counted with a Coulter electronic cell counter. The cell samples were diluted in cold BSS-CaS3 to givea finalconcentration of 3 x 106cells/mlfor coxsackievirus B3,and 107cells/ml for the other three virus preparations. The CAV was deter-mined byfurther diluting each sample into acid solution, used for elution ofCAV, as described previously. The reversal of CAV with 8 M urea, 6 MLiCl, and 4% SDS wasperformedbymakinga 10-folddilution ofeachcell sample into the rea-gent followed bya 100-folddilution into PBSA-CaS3 . The amountof freevirus obtainedby each of the methods tested was determined by the plaque assay. In these experiments, no attempt wasmadetoaccount forthetotal amount of the input virus aswas donein the experiments pre-sented in Table 1.
The results of these experiments, compiled in Table 2, show that maximal recovery of cox-sackievirusesBi and B3 wasobtainedonly bythe method involving pH 1.5 and, therefore, the amounts obtained by this method were assigned values of 100%. The
high
saltconcentration
re-versed onlyasmallproportion,and SDS approxi-mately50%,
of thecoxsackieviruses
available at the cell surface. In contrast, the poliovirusTi
receptorcomplexwasdissociated equally wellby the
high
saltconcentration and pH 1.5, withSDS againbeing less efficient. Theamountofrecovery of echovirus6 wasapproximately
thesamefor all of the methodstested.Ingeneral,theprocedureof alternatefreezing
andthawingwasfoundtobean ineffective method for recoveringCAV.Effectof acidpHonthecapacity ofHeLa cells to attach and eclipse enteroviruses. The effective use ofacidpHfordissociation ofreceptor-virus complexespromptedastudyof thestabilityatlow pH of theviralreceptors of HeLacells.HeLacells were suspended in 5 ml of cold 0.05 M glycine buffer atpH2.5 toa finalconcentration of 107/ ml. At intervals during incubationat 2 C,
1-ml
samples were withdrawn and diluted 10-fold in PBSA-CaS3 to bring the pH to neutrality. The cellsweresedimented by centrifugation for5min at225 X g,washed, counted,andresuspendedin
BSS-CaS3
toa concentration of 107cells/ml.
Thetreated cells were tested for their capacity to attachpoliovirus Ti andcoxsackievirusB3during a60-min
period
at37 C. Asacontrol,
cellstreated atneutralpHweretestedconcurrently.
on November 11, 2019 by guest
http://jvi.asm.org/
TABLE 2. Capacity of assorted reagents to dissociate enterovirusesfrom HeLa cells
Amt ofvirusrecoveredafter 1 minat 2Ca
Reagents CoxsackieBi Coxsackie B3 PolioTi Echo 6
PFU/ml Percent PFU/ml Perc PFU/ml PerPcentPFUml Percent (108) Pe et (107) Pe ent (i01) Pe et (10'1) Pern
0.05 M Glycine (pH 1.5) 1.07 100 1.34 100 3.9 100 1.26 100
0.05 M Glycine (pH2.5) 0.058 5.4 0.03 2.2 0.05 1.3 1.34 106
8 M Urea 0.028 2.6 0.05 3.7 4.6 118 1.48 117
6M LiCl 0.022 2.0 0.008 0.6 3.5 90 1.56 124
4% SDSin BSSb 0.57 53 0.4 30 2.7 69 1.38 109
1% Chymotrypsinc
NDd ND 0.02 1.5 ND ND ND NDFreezing andthawing (5times) 0.16 15 0.11 8.2 0.18 4.6 1.30 103
None' 0.0078 0.7 0.002 0.2 0.01 0.2 0.24 19
aWashed HeLacells (5 X
107)
were suspended in5ml of virusat 2 C. Input multiplicities of viruswere 10, 50, and 100 PFU for coxsackievirus B3, coxsackievirus Bi, andpoliovirusTi and echovirus 6, respectively. After incubation for 1 hr at 2 C, the cell-virus suspensionwas dilutedto 50ml with
BSS-CaS3andcentrifugedat225 X g. Thecellswerewashedaccordinglyfourmoretimes. After the fifth wash, the cellswereadjustedto3X 106cells/mlforcoxsackievirus B3 andto10'cells/mlfor the other
viruses.CAVwasdetermined,asindicated in Materials and Methods. In theseexperiments,noattempt wasmadeto account for the totalamountofinputvirus.
b Duetotheformation ofaprecipitateat2C,the temperature of the SDSsolutionwas15C. cCells were treatedfor 30 minat37Cat aconcentrationof 3 X
106/ml.
dNot done.
eTheamountof virusin the fluidphaseofthe cellsuspensionpriorto treatment.
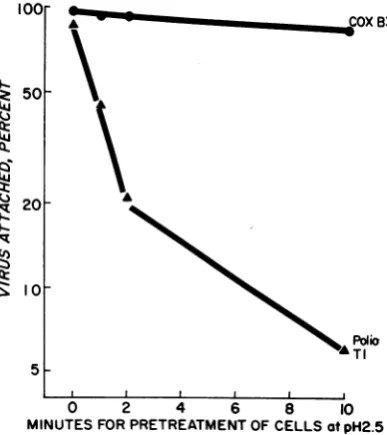
Theresultspresentedin
Fig.
2demonstrate that the receptor activity for coxsackievirus B3 re-mainedstableto treatmentatpH2.5for 10min, whereas the receptor activity for poliovirus Ti wasrapidlyinactivated.To determine the pH at which the cellular receptorsforeachoffourenteroviruses would be inactivated,replicatesamplesof HeLacells (2 x
107 cells) were resuspended in 10-ml volumes of cold 0.05 M glycine buffer
adjusted
to different pH levels. After a 10-min treatment at 2 C, the cells werewashed,resuspended in asmallvolumeof
BSS-CaS3,
andassayed
forcapacity
to attachand to eclipse each ofthe viruses at neutralpH
during
a 1-hrperiod
at 37 C. The amount ofattached virus which could not be recovered by 0.05Mglycine bufferatpH 1.5 wasconsidered to be eclipsed (7). The resultspresented inTable 3 show that the receptors for
attaching
coxsackie-viruses B1 and B3 and echovirus 6 were stable overthepHrange 1.0 to7.0,whereas thereceptors forpoliovirus TI were inactivated at a pH below 3.0. In contrast, theresults presented in Table 4 demonstrate thattheeclipse activityof HeLacells for each ofthe viruses tested was inactivatedby treatmentbelowpH 3.0. It should be noted that such treatmenthadnoeffectonthe attachment to HeLa cells of coxsackieviruses Bi and B3 and echovirus 6. Since HeLa-cellreceptors for polio-virusTi
were inactivated below pH3.0, thein-loor
2 50
Id10
Iq 2
QC ZIN 10
5
A - COX B3
F
PTI
0 2 4 6 8 10
MINUTES FOR PRETREATMENTOFCELLSatpH2.5 FIG. 2. Comparative rates of inactivation, at pH
2.5, ofthe ability of HeLa cells to attach coxsackie-virus B3 andpoliovirus Ti.
activationofeclipse below that level could not be demonstrated.
Effect oftemperature on the capacity of HeLa
cells to attach and eclipse enteroviruses. The
on November 11, 2019 by guest
http://jvi.asm.org/
TABLE3. Comparative stability ofenterovirus receptors ofHeLa cellsto acidpH
Virusattachedb Pretreatment of
cellsa Coxsackie Coxsackie Echo 6 PolioTi
Bi B3 Eco6PloT
pH % %
N
%1.0 99 97 74 2
2.0 96 97 86 7
3.0 99 99 80 54
4.0 98 99 82 65
5.0 98 97 76 55
6.0 99 97 77 57
7.0 98 97 80 64
BSS-CaS3 98 99 75 67
aReplicate samples of2 X 107 cells were
sus-pended in 10 ml ofcold 0.05 M glycine buffer at
thepH indicated and treated for 10 min at 2 C. BSS-CaS3atpH 7.0 served as acontrol.
bTreated cellswere sedimented, washed, resus-pended in 1 ml of BSS-CaS3, and counted. A
sample ofcellsuspension containing 107 cells/ml wasadjustedtovolumeof0.9ml,and 0.1ml of re-spective virus was added. Theinput PFU/0.1 ml was6.7 X 105 for coxsackieBi,2.9 X 105 for cox-sackie B3,4.2 X 105 for echo6, and5.5 X 105for
polioTi.Thecapacityof thecellsto attach each of theviruseswas determined at neutral pH for 1 hr at37 C.
porteddifferences in theheatstabilityofreceptors for enteroviruses (18, 21) suggested temperature as a means for the selective inactivation ofthe capacity of HeLa cells to attach and to
eclipse
enteroviruses. Replicate samples of 2 X 107 HeLa cellsweresuspendedin1.9 mlofBSS-CaS3 and heated at selectedtemperatures for 30 min. The cellsuspensions werecooledpriorto receiv-ingvirusinoculum, andthecapacityof the heat-treated cells to attach and to eclipse coxsackie-virusesBi andB3, poliovirus
TI,
andechovirus6 was determined at 37 C for 60 min.Replicate
samplesofcells which were notheatedservedas
controls.
The results presented in Table 5 indicate that the receptors for
attaching
poliovirus
Ti wereinactivated at 56C, revealingthat theywerethe mostlabileof thereceptors studied. The receptors for echo 6 were
partially
inactivated at 60C,
whereas a
slightly
greater temperature wasre-quired for inactivation of the receptors for
cox-sackieviruses Bi and B3. The results in Table 6 present the data
pertaining
tothethermostability
ofthe capacityofHeLa cells toeclipsethefour enteroviruses tested. These results demonstrated that, when HeLacellswereheatedat56 Cfor 30 min,theylost their
capacity
bothtoattach andtoeclipse
poliovirus
TI. On the otherhand,
thesecells attached but did not eclipse echovirus 6, whereas the attachment and eclipse mechanisms for coxsackieviruses Bi and B3 were affected
mninimally.
To confirm the observation that HeLa cells have a mechanism for eclipse of coxsackievirus
[image:5.494.59.251.92.237.2]Bi
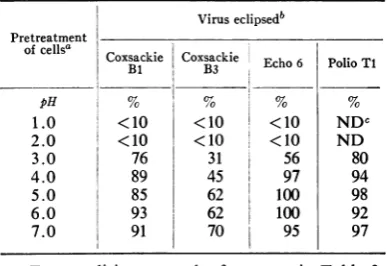
whichcouldbe separated from that for echo-virus 6, anadditional experimentwasperformed. HeLacellsweresuspendedin BSS-CaS3 at a con-centration of107cellsperml and heated at 56 C. Atintervals, samples ofcells were removed and testedfortheircapacity to attach and eclipse each virus as described previously. The results of this experiment (Table 7) confirmedthe previous ob-servationthatattachment ofbothviruses occurred normallytotheheatedcells. However, the eclipse mechanism for echovirus6 wasrapidly inactivated whereasthecellswerestillcapable ofeclipsing at least 60% of the attached coxsackievirus Bi (Table 7). These results reinforce the previous observations and suggest that the mechanism which is involved in the eclipse of coxsackievirus Bi is distinct fromthatfor echovirus 6.TABLE 4. Effect of acid pHon the capacity of
HeLacells toeclipse
eniteroviruses
Viruseclipsedb
Pretreatment
ofcellsa coxsackie Coxsackie Echo 6
Polio Ti
Bi B3
pH N S % %
1.0 <10 <10 <10 NDC
2.0 <10 <10 <10 ND
3.0 76 31 56 80
4.0 89 45 97 94
5.0 85 62 100 98
6.0 93 62 100 92
7.0 91 70 95 97
a For conditions, see the footnotes in Table 3. AftertreatingthecellsatthepHindicated,the
capacityof the cellstoattach (seeTable3)andto eclipse each of the viruses was determined at
neutral pH for 1 hr at 37 C. Eclipsed virus was
consideredtobe thatamountofinput virus
infec-tivity (seeTable3)whichwasdeterminedinitially tobecell-associatedandwhichwasnotdissociable from the cells by treatment with 0.05 M glycine
buffer (pH 1.5). The values of viruseclipsed were
calculated from thefollowing equations: (i) virus
attached (PFU) = input virus - unattached
virus; (ii) virus not eclipsed (PFU) = acid
re-covered virus - unattached virus; (iii) virus eclipsed (PFU) virus attached - virus not
eclipsed; (iv) viruseclipsed (%) = virus eclipsed
(PFU) X 100/virus attached (PFU).
Not done; the pretreatment inactivated the
capacity of the cellsto attach poliovirus Ti.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.494.264.458.353.486.2]TABLE5. Thermostability ofenterovirus receptors of HeLacells
Virusattachedb Pretreatment
of cellsa Coxsackie Coxsackie Echo6 PolioTi
Bi B3 Eco6PloT
C % No No %
37 98 96 77 81
40 NDc ND 80 74
45 ND ND 74 78
50 ND ND 67 62
56 99 98 69 15
60 98 97 42 <10
65 <10 <10 25 <10
70 <10 <10 <10 <10
aReplicate samples of 2 X 107 cells were sus-pended in 1.9 ml ofBSS-CaS3 and heated at the temperatureindicated for 30 min.
bCellsuspensionswerecooledpriortoreceiving the respective virus. The input PFU/0.1 ml was
1.98 X 106 for coxsackieBi,2.5X 107forcoxsackie
B3, 1.38 X 106 for echo 6, and 1.16 X 106 for
polioTI. Thecapacityof the cellsto attach each of the viruseswasdetermined at37Cfor 1hr.
c Not done.
TABLE 6. Thermostability of the capacityof HeLa cells toeclipse enteroviruses
Virus eclipsedb Pretreatment
ofcellsa Coxsackie Coxsackie Echo 6 Polio1
Bi B3 Eh oi
C % N % %
37 72 73 68 83
40 NDC ND 68 68
45 ND ND 61 82
50 ND ND 28 56
56 65 43 13 <10
60 63 39 <10 d
aFor conditions, see the footnotes in Table 5. bAfter treatment, the capacity of the cells to attach (see Table 5) and to eclipse each of the
viruseswas determined at neutralpH for 1 hrat
37 C. Eclipsed virus was considered to be that
amount of input virus infectivity (see Table 5)
which was determined initially to be cell-asso-ciated and which was not dissociable from the cells by treatment with 0.05 M glycine buffer (pH 1.5). The values of virus eclipse were
cal-culated from the following equations: (i) virus attached (PFU) = input virus-unattachedvirus; (ii) virus not eclipsed (PFU) = acid recovered virus - unattached virus; (iii) virus eclipsed (PFU) = virus attached - virus not eclipsed; (iv) virus eclipsed (%) = virus eclipsed (PFU) X 100/virus attached (PFU).
e Not done.
dPretreatment inactivated the capacity of the cellstoattach poliovirusTi.
TABLE7. Comparative ratesofinactivation of
the ability of HeLa cells toattach and eclipse coxsackievirusBland
echovirus 6
Coxsackie Bi Echo 6 Pretreatment of
cellsat 56CG
Attached Eclipsedb Attached Eclipsedb
min % % % %
None 96 78 88 68
5 97 61 81 5
15 96 61 72 <1
30 97 59 69 <1
aReplicate samples of 107 cells/0.9 ml of
BSS-CaS3 were heated. At time intervals indicated,
the cellswere removed andcooled prior to addi-tion ofvirus.Theinput PFU/0.1mlwas8.7 X 105 and 7.9 X 106for coxsackieBl andecho 6,
respec-tively. The capacity of the cells to attach and eclipse each of the viruseswasdetermined at 37C for 1 hr.
bValues were calculated from the equations given in Table 4.
DISCUSSION
Thecomparative experimentsdescribed inthis report represent butonephase of ourcontinuing
investigations
directed toward characterizing theproperties and functions of enterovirus receptors onintactHeLacells
(1-4, 10, 23, 24).
The results of theseexperiments
showedclearly
that the receptoractivity
ofHeLacellsforattaching polio-virus Ti wasmorelabiletoelevatedtemperatures andtolowpHthanwerethe receptoractivitiesforbinding
echovirus6and coxsackieviruses Bi andB3. These
findings,
in part, confirmed and ex-tended the results ofQuersin-Thiry
and Nihoul (21), who showed a difference between thethermostability
of receptoractivity
ofHeLa cellextractsfor
poliovirus
Ti and coxsackievirus B4. Thus, additional evidence is provided which serves to reemphasize the differences among the receptorsofHeLacells forattachingthedifferent enteroviruses.To determine the thermostability and acid
stability
ofHeLacellstoeclipse enteroviruses,
weconsidered it important first to obtain a method tomeasure accuratelytheamount ofviruswhich
becomes cell-associated under conditions pre-cluding viral eclipse. Thus, of all the methods tested, a brief exposure ofCAV to buffer at pH
1.5 at 2Cprovidedageneralmethod for recovery of maximal amounts of each of the viruses studied. Bydetermining the amount of receptor-bound virus at a given
time,
it was possible toassessthecapacityof HeLacellstoeclipse entero-virusesat37 Caftertreatmentof cells with acid or heat. Theresults showed that theeclipse activities
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.494.39.233.74.239.2] [image:6.494.244.439.79.214.2] [image:6.494.40.231.336.483.2]of cells (as defined in this paper) for coxsackie-viruses Bi and B3 and echovirus 6 were inacti-vated irreversibly at pH2.0 without impairing the activity of the receptors for attaching these viruses. In contrast, thefinding that both of these functions were inseparable for poliovirus Ti suggests that different events may be operative during theearlystagesof infectionby the different enteroviruses.
It was observed that even though heating of HeLacellsat56Cresulted in lossof cellviability, as determined by trypan blue staining (un-publisheddata),such "dead" cells, which did not attachpoliovirus Ti, attachedbutdidnoteclipse echovirus 6; yet these cells both attached and eclipsedcoxsackieviruses Bi and B3. Thefinding thatheat-killedcells remained capable of attach-ing and eclipsing coxsackieviruses Bi and B3
raises again the question whether the uncoating of coxsackieviruses occurs within phagocytic vesicles (13) oratthe receptors on thecell surface (3, 17). The improbability that heat-killed cells retainedtheability of engulfing virussupportsthe latter possibility. Resolution of this question, however, requires a better understanding of the relationship between the various operational de-finitions of viral eclipse and ofthe uncoating of the RNA genome ofeachofthe different
entero-viruses.
ACKNOWLEDGMENT
Thisinvestigation wassupported by Public Health Research grantAI-03771 toR. L. Crowellfromthe National Institute of Allergy andInfectiousDiseases.
LITERATURE CITED
1. Axler, D., and R. L. Crowell. 1968. Effect of anticellular serumonthe attachmentof enterovirusestoHeLacells.J. Virol.2:813-821.
2. Crowell,R.L. 1963.Specificviral interference in HeLa cell cultures chronically infected with coxsackie B5 virus. J. Bacteriol. 86:517-526.
3. Crowell, R. L. 1966.Specificcell-surfacealterationby
entero-virusesasreflectedby viral-attachment interference. J. Bac-teriol.91:198-204.
4. Crowell, R.L.,andJ.T.Syverton. 1961. The mammalian cell-virusrelationship. VI. Sustained infectionof HeLa cellsby coxsackieB3virusandeffectonsuperinfection. J. Exptl. Med. 113:419-435.
5. Darnell, J. E., andT.K.Sawyer. 1960.Thebasis for variation
insusceptibility to poliovirus in HeLacells. Virology 11: 665-675.
6. Fenwick,M.L.,andP. D.Cooper. 1962. Earlyinteractions between poliovirus and ERK cells: someobservations on the nature and significance of the rejected particles. Virology 18:212-223.
7. Holland, J. J.1962.Irreversible eclipse ofpoliovirus by HeLa cells. Virology 16:163-176.
8. Holland, J. J. 1964.Enterovirus entrance into specific host cells, and subsequentalterationsof cellprotein and nucleic acidsynthesis. Bacteriol.Rev. 28:3-13.
9. Kunin, C. M. 1962. Virus-tissue union andthe pathogenesis of enterovirusinfections.J.Immunol.88:556-569.
10. Levitt,N. H.,andR.L. Crowell. 1967.Comparativestudies ofthe regenerationof HeLa cellreceptors forpoliovirusTi andcoxtackievirusB3. J. Virol. 1:693-700.
11. Mandel,B.1961.Reversibilityofthereaction between polio-virusandneutralizingantibody of rabbit origin. Virology
14:316-328.
12. Mandel, B. 1962. The use of sodium dodecyl sulfate in studies on the interaction of poliovirus and HeLa cells. Virology 17:288-294
13. Mandel, B. 1967.The relationshipbetween penetration and uncoating ofpoliovirusin HeLa cells.Virology 31:702-712. 14. MeLaren, L.C.,J. J.Holland, and J. T. Syverton. 1959. The mammalian cell-virus relationship. I. Attachment of polio-virustocultivated cellsofprimateand non-primate origin. J. Exptl. Med. 109:475-485.
15. McLaren,L.C., J. J. Holland,andJ. T.Syverton. 1960. The mammalian cell-virus relationship. V. Susceptibility and resistance of cellsin vitrotoinfectionbycoxsackieA9 virus. J.Exptl. Med. 112:581-594.
16. McLaren, L. C.,J.V. Scaletti, and C. G. James.1968. Isolation andpropertiesofenterovirus receptors. Monograph no. 8, p.123-136. WistarInstitutePress,Philadelphia. 17.Philipson, L. 1967. Attachmentandeclipseof adenovirus. J.
Virol.1:868-875.
18. Philipson, L.,andS.Bengtsson. 1962. Interaction of entero-viruses with receptors from erythrocytes and host cells. Virology18:457-469.
19.Philipson, L.,and M.Lind.1964. Enterovirus eclipseina cell-free system. Virology23:322-332.
20. Pinheiro, F.,andG.D.Hsiung.1963.Astudyondissociation ofcoxsackieB4virus-antibody complex. Virology 20:457-463.
21. Quersin-Thiry, L.,and E.Nihoul. 1961. Interaction between cellularextracts andanimal viruses. II. Evidence forthe presenceofdifferentinactivatorscorrespondingtodifferent viruses.ActaVirol. 5:283-293.
22. Soloviev, V. D.,T. I. Krispin, V. G. Zaslavsky, andV. I. Agol. 1968. Mechanism of resistanceto enterovirusesof someprimate cells in tissueculture.J.Virol. 2:553-557. 23. Zajac, I.,and R.L.Crowell. 1965.Effectof enzymesonthe
interactionofenteroviruseswithlivingHeLacells.J. Bac-teriol.89:574-582.
24. Zajac,I.,and R.L. Crowell.1965.Location andregeneration ofenterovirusreceptorsofHeLacells.J. Bacteriol. 89:1097-1100.