Copyright© 1972 American Society for Microbiology Printed inU.S.A.
Saint
Louis
Encephalitis
Viral Ribonucleic Acid
Replication
Complex
ATEEF A. QURESHI AND DENNIS W. TRENT
Department of Microbiology, University ofTexas Medical School at San Antonio, SanAntonio, Texas 78229
Received for publication 26 October 1971
Pulse-labeled Saint Louis encephalitis viral ribonucleic acid (RNA) is found in
thecytoplasm of infected cells associated with a membranous structure which
sedi-ments with an average value of 250S. The integrity of the complex isdestroyed by
de-tergents and ribonuclease; however, it is stable in ethylenediaminetetraacetic acid
(EDTA) which differentiates this structure fromcellularpolyribosomes. With
cul-turesinwhich cellular RNA was highly labeled prior to infection, ribosomal RNA
couldnot be demonstrated inthecomplex isolated from EDTA-sucrosegradients.
Single-stranded 43S and the 26S and20S forms of viral RNA were found in the
com-plex.ViralRNApolymeraseactivity in sucrose-gradient fractions sedimented in the
same region as the fractions which contained the pulse-labeled viral RNA. The
polymerase incorporated8H-guanosinetriphosphate into acid-precipitable material
in theabsence ofadded template. It was also found that the replication complex
contains viral-specific proteins.
SynthesisofgroupAarboviral ribonucleic acid
(RNA) andenvelopment of thenucleocapsidare
associated with membranous structures (7, 8, 10,
17,18). Intracellular, complete,group B arbovirus
particles are observed inside membrane-bound
cytoplasmic vacuoles although the role of these
structures in viral macromolecular synthesis and
virion envelopmentis not clear (6, 15, 16).
Cali-guriand Tamm(3, 4)havereportedthat
transla-tionandtranscriptionofpolioviralRNA are
asso-ciated with distinct types of membranes which
canbeseparated by isopycnicsucrose
centrifuga-tion. Inpicornavirus-infected cells, RNA
synthe-sis is initiated in 130S structures found in the
smoothmembrane fraction whichmature toform
the late replication complex that has a
sedimen-tationcoefficientofapproximately250(3, 4).This
mature structurecontainsviral RNApolymerase,
single-stranded RNA, the replicative 20S form,
andthereplicativeintermediate 26S form of viral
RNA(4, 9). Thisstructureis definedasthe
viral-RNAreplication complex
(9).
Cytoplasmic
mem-brane-associated
(MRC)
structuressimilartothepicornavirusreplication complex arealso thesite
of viral RNAsynthesis in cells infected with the
group A arboviruses (7, 10).
Thiscommunicationdescribes theisolationand
preliminary
description
of areplication
complexfromBHK-21/13 cellsinfected with Saint Louis
encephalitis
(SLE)
virus. This structure has anaveragesedimentation coefficient of250,contains
viral RNA polymerase, single-stranded viral
RNA, and the replicative forms of SLE viral
RNA. Viral-specific proteinsappear to be an
in-tegral part of the SLE viralcomplex.
MATERIALS AND METHODS
Virus andcell culture.Pools of SLEvirus and pig
kidneycellmonolayer cultures were prepared
accord-ing to procedures described previously (11, 20). Baby hamsterkidney cells (13) obtained from the American Type Culture Collection were grown in a
lactalbumin medium(20).
Infection procedure. Confluent monolayercultures
of BHK cellswereinfectedwith SLEvirusat a
multi-plicity of 10plaque-forming unitsper cell. After an
absorption periodof 1 hr, warmEagle's medium (5)
with actinomycin D (0.5 ,ug/ml) and 5% calfserum
wasadded, and cultureswereincubated at37C. Cell fractionation. Cultures were pulse-labeled for
15minwith3H-uridine(50,uCi/ml)insingle-label
ex-periments.Indual-labelexperiments,50,ACiof
3H-uri-dine/mland10,ACiof'4C-leucine/mlwereused.After
theradioactive pulse,cells werescraped off theglass
and collected bycentrifugation, andcytoplasmic
ex-tracts were prepared (7). Insomeexperiments,
cyto-plasmicextracts were solubilized withBrij-58 or
so-diumdeoxycholate (DOC; 0.5%for 10 min at0C)
priortosucrosegradient analysis. Extracts were
lay-ered over a
28-mi,
5 to 40% (w/v) sucrose gradientprepared in eitherreticulocyte standardbuffer (RSB;
reference 22) or ethylenediaminetetraacetic acid (EDTA) buffer (9) andcentrifugedat25,000rev/min
for 120 minat4 CinaBeckman SW-27rotor. Frac-565
on November 10, 2019 by guest
http://jvi.asm.org/
tions collected from the gradients were analyzedfor opticaldensity,trichloroaceticacid-precipitable radio-activity (20), orboth.
Enzyme assay. Viral RNA polymerase activity in cell extracts andgradient fractionswasdeterminedby
themicrotechnique of LevinandFriedman(12).Fifty lambda of the enzyme preparation was incubated at
37 C for 60 min in a total volume of 0.1 ml which contained 0.05 M tris(hydroxymethyl)aminomethane,
pH8.0;0.01 MMgCl2; 0.001 Mdithiothreitol, 0.025jAg
ofactinomycin D, 0.01 M phosphoenol pyruvate;4jg
ofpyruvatekinase, and 10 nmeach ofadenosine tri-phosphate, uridine triphosphate, cytidine
triphos-phate, and 3H-guanosine triphosphate (GTP; 2.9 X
106counts permin).
Analysis ofRNAassociated withcytoplasmic struc-tures. RNA wasextracted frompooled, sucrose gra-dient fractions with sodium dodecyl sulphate-phenol
at 37 C andanalyzedbysucrose ratezonal
centrifuga-tion (20). One-half of each fraction was hydrolyzed
with ribonuclease (20,g/inl), and each sample was
precipitated with 10%ho trichloroacetic acid, collected
on glass fiber filters, and washed several times with
5%70 trichloroacetic acid, and dried, and the radio-activity wasdetermined (20).
Immunoprecipitation of cytoplasmic fractions.
Su-crosegradientfractions(200Sto300S) ofcytoplasmic
extracts of 3H-leucine-labeled SLE viral-infected or
noninfected cellswere pooledanddialyzed overnight
at4 C against 0.02 M phosphate buffer, pH 7.4. An
amount(25,uliters) ofthedialyzedsamplewaslayered
over 25 Muliters of SLE virus antibody-containing
mouseascitic fluid (2) and incubatedat room
temper-ature overnight. A positive test was indicated by a
zoneofprecipitatewhich formedattheinterphaseof
the solutions containing homologous antigen and antibody. Immunological precipitation of 3H-leucine-labeledproteininpooled gradientfractionswas quan-titated by the radioimmune precipitation technique
(19).
Chemicals. Ribonuclease was purchased from
Worthington Biochemical Corporation, Freehold,
N.J., and ribonuclease-free sucrose wasfrom Mann
Research Laboratories, N.Y. Actinomycin D was a
gift of Merck-Sharp and Dohme Research
Labora-tories, Rahway, N.J. 3H-uridine and 3H-guanosine
triphosphate were obtained from Schwarz/Mann,
Orangeburg, N.Y.; 14C- and 3H-leucine were pur-chased from New England Nuclear Corporation,
Boston, Mass.
RESULTS
Cytoplasmic fractions associatedwithviral RNA
synthesis. Cytoplasmic extracts prepared from
noninfected cellsweredivided, and a portion was
incubated with either DOC or Brij-58 before
centrifugation on RSB-sucrose density gradients
(Fig.
1).The A26ogradient profile of cytoplasmicextracts prepared without detergent treatment
shows materials which sediment with a size
dis-tribution of 300S to 236S, 74S ribosomes, and
lighter material. Brij-58 treatment prior to
gra-dient analysis increased the size distribution of
material in the major A260 peak from 340S to
200S. Sucrose gradients of extracts after DOC
treatmentcontainedrelativelyless material in the
200S to 300Sregion of the gradient.
Cells infectedfor 18 hr were labeledwith
3H-uri-dine for 15 min; cytoplasmic extracts were
pre-pared and treated with either DOC or Brij-58
and centrifuged through preformed RSB-sucrose
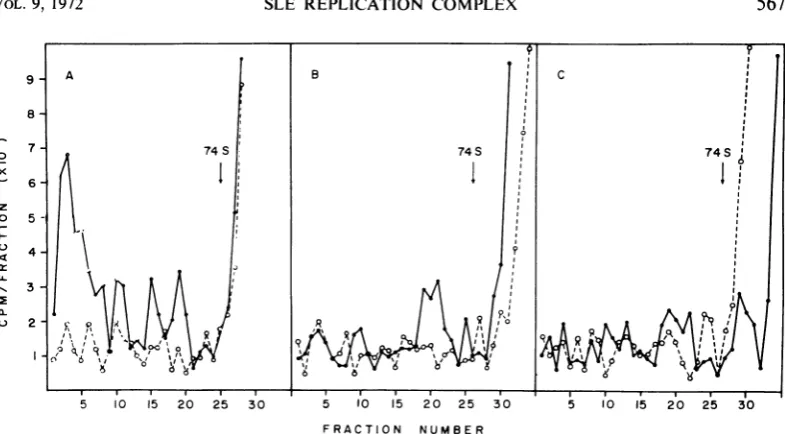
gradients (Fig. 2). The gradient profile of
non-detergent-treated cytoplasmic extracts had a
3H-uridine peak that contained approximately
40%o of the total radioactivity and sedimented
with an average value of 300S (Fig. 2A). Three
smallerpeaksofradioactivitywerealso observed
at 250S, 180S, and 140S.Afterincubationof the
extract with ribonuclease, less than
10%,
of theradioactivity observed in the fast-sedimenting
material of the untreated extract sedimented at
200S to 300S. Incubation of the extracts with
either Brij (Fig. 2B) or DOC (Fig. 2C) prior to
I10
E 236 S
c 0.5
o 300S 74S
o
_ _
(\J
>-01
01
0 5 10 15 20 25 30
FRACTION IUMBER
FIG.1.Effectof sodiuincleoxycholate
anid
Brij-58olncytoplasmic extracts fromnolninfcetedBHKcells.
Cy-toplasmic extract was prepared, divided, treated with
either DOC or Brij-58, anid centrifuged through pre-formed reticulocyte stanidard buffer-sucrose
gradienits.
Centtrifugationt was perfbrmed with a SW-40 rotor at
25,000 rev/minifor120 miii at 4 C. Fractiouis (0.5ml)
were collectedanidatnalyzedforoptical
denisity
at 260 mim.Symbols: (0) witreated cytoplasmicextract; (A) Brij-58-treated cytoplasmic extract;(0) deoxycholate-treated extract.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.500.269.458.300.553.2]7-74 S 74S 74S'
6
i
z o
5-
04-LL
-- 3
a.
2-~~~~~~~~~~~~~~~~~~~~
' , , . q
66 6
5 10 520 25 30 5 10 15 2025 30 5 10 I1 20 2530
FRACTION NUMBER
FIG. 2. Effectofsodiumdeoxycholate, Brij-58,and ribonuclease ont thecytoplasmicextractsfrom SaittLouis
enicephalilis virus-infectedcells. At 18hrpostinfection cultures werepulse-labeledwith 3H-uridinte (50 ,uCi/ml)
for 15miii. Cytoplasmicextracts wereprepared, divided, itncubatedwith
deoxycholate
orBrij-58,
anidcenztrifuged
throughpreformedreticulocytestandardbuffer-sucrosegradients.Fractiouisof1.0mlwerecollected,anidone-halfof eachfractionwasincubated with10 lAgofribontucleaseat37 Cfor10 miii.Riboniuclease-ireatedand
niontreated
fractions were analyzed for acid-precipitable radioactivity. Total 3H-uridine radioactivity(@);
ribontuclease-resistant3H-uridineradioactivity (0).A,
nondetergenit
treated;B,Brij-58
treated; C, deoxycholatetreated.centrifugation dissociated mostof the 3H-uridine
radioactivity from structures which sediment at
200S to 300S. When the250S 3H-uridine-labeled
peak from infectedcells was isolated from
RSB-sucrose
gradients,
treated withBrij
orDOC,
andrecentrifuged,the
gradient
profiles
weresimilartothose
presented
inFig.
2A and2B,
respectively.
Unlike
polyribosomes
from noninfectedcells,
whichwerestableto
recentrifugation
afterBrij-58
treatment, the faster
sedimenting
3H-uridine-la-beled structures fromvirus-infected cells are
dis-sociatedby the nonionic
detergent.
These resultsindicate that SLE viral RNA is associated with
detergent and ribonuclease-sensitive structures
which are distinct from
polyribosomes
observedinuninfected cells.
Characterization of the
replication complex.
Since the
rapidly
sedimenting
3H-uridine-labeled
structurefromSLEvirus-infectedcellssedimented
in sucrosegradientsina manner similar to
poly-ribosomes,
it wasthought
itmight
contain viralmessenger RNA
(mRNA)
and ribosomes or itcould be a
replication
complex.
Since EDTA isknownto causedissociationof mRNA and
ribo-somes (9), its effect on the sedimentation
profile
of the
complex
wasinvestigated.
Cytoplasmic
ex-tracts were
prepared
from cultures labeled with3H-uridine and '4C-leucine for 30 min at 16 hr
postinfection.
Halfof theextractwasadjusted
to0.02 M EDTA and
analyzed
by
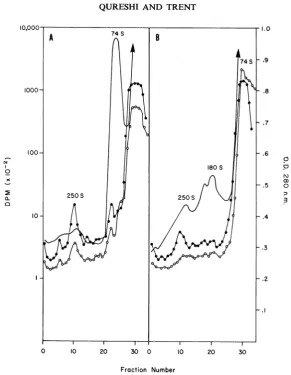
sedimentationonanEDTA-sucrosegradient (Fig. 3A). The control
extract was analyzed by sedimentation on a
RSB-sucrose gradient (Fig. 3B). TheA280 profile
ofthe control
(Fig.
3B) shows two peaks atap-proximately250S and 180S,the74S single
ribo-somes,andlighter materials. IntheRSB-sucrose
gradient,most of the3H-uridineradioactivity
re-mained nearthetop ofthe gradientwith asmall
amountoflabeled materialsedimenting inapeak
at2505.TheA28oprofile ofEDTA-treated extracts
(Fig. 3A) did not have a major peak at 180S
(polyribosomes)
but does show a broad peaksedimenting
in the 200S to 3005 region of thegradient. Radioactive RNA and protein
cosedi-mented in the EDTA-sucrose gradient in the
peak at 250S.These results differentiate between
structureswhichcontainpulse-labeledviral RNA
andare stablein EDTA(Fig. 3A) andthe
poly-ribosomes which sedimentatapproximately 180S
andaredissociated inthepresenceofEDTA.
To
investigate
whethercellularribosomalRNAis an integral part of theEDTA-resistant
struc-ture, uninfected BHK cells were incubated in
3H-uridine-containing
mediumfor 24 hrtolabelcellular ribosomal RNA. After an additional 12
hr in mediumwithoutlabel,the cellswereinfected
for 16hrin the presence of
actinomycin
(0.5
Mg/
ml) andpulse-labeledwith
'4C-leucine
for15 min.Cytoplasmicextractsprepared fromthe cellswere
divided and sedimented through either
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.500.51.444.57.274.2]0
0
a p
W
0 3
0 10 20 30 0 10 20 30
[image:4.500.113.405.54.430.2]Fraction Number
FiG. 3.Effectofethylenediaminetetraaceticacid(EDTA)onthereplication complex. 3H-Uridine (50 Ci/ml)
and14C-leucine (10,uCi/ml) wereaddedtocultures 16 hrpostinfection. Incorporation wasstopped after30 min.
(A) partoftheextract wasadjustedto0.02 MEDTAand sedimentedinanEDTA-sucrosegradient. (B) control
wassedimentedin an reticulocyte standardbuffer-sucrose gradient. Symbols: (0) acid-precipitable 3H-uridine
radioactivity; (0) acid-precipitable 14C-leucinieradioactivity; (-) A280.
sucrose orRSB-sucrose
gradients.
Fig.
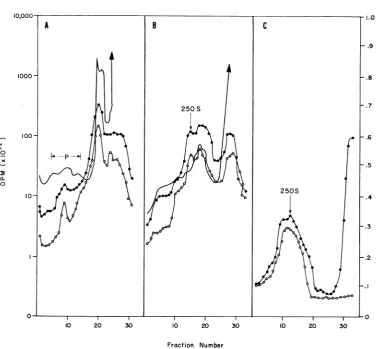
4AshowsthatuponEDTAtreatmentmostofthe3H-uridine
in RNA from prelabeled cells sediments at the
top of the
gradient
atapproximately
745 with acoincident
peak
of3H-uridine
and '4C-leucineradioactivity
sedimenting
atapproximately
250S.In contrasttothis, RSB-sucrose
gradients
ofex-tractsfromprelabeledcells
(Fig.
4B)
contained alarge prominent peak of 3H-uridine and
'4C-leu-cine radioactivity which sedimented at 180S to
250S.
Since a small percentage of 3H-uridine in the
EDTA-sucrose gradient sedimented inthe
heavy
regionof the gradient (Fig. 4A), fractions inthe
200Sto250S region of the
gradient
werepooledand resedimentedin EDTA
(Fig.
4C).Resedimen-tation ofthe 250S peak in EDTA shifted more
than 90% of the 3H-uridine counts in these
pooled fractionstothe topof thegradient.These
data indicate that prelabeled cellular ribosomal
RNAispartially dissociatedto structures smaller
than250SbyEDTAtreatment andthat some of
theremainingRNAwhichsedimentsat250Swill
partially dissociatetosmallerstructuresif treated
againwith EDTA. Theproteinin the250S peak
whichwaslabeledwith 14C-leucineduring the
in-fectionwas notdissociated from the complex by
this repeated EDTA treatment. These data
sug-gestthatmostofthecellularRNAfrom the
pre-labeledcellswhich sedimentswith the 250S
struc-tureisloosely bound.
Todetermine the type of RNA left in the 250S
region of thegradient from prelabeled cells after
the second centrifugation in EDTA, fractions 6
on November 10, 2019 by guest
http://jvi.asm.org/
0 c0
x
m-cz
,0
.9
0
OD
0 3
10 20 30 10 20 30 10 20 30
Fraction Number
FIG.4.Absenceofribosomes inthe replication complex. BHK cells were prelabeled with 50 ,uCi of3H-uridine
per mlfor 24 hr and then 12 hr in the absence of the isotope. The cells were infected for 16 hrinthepresence
ofactinomycinandpulse-labeledwith14C-leucine(10,uCi/ml)for 15 min,andcytoplasmic extractswereprepared. (A) partofthe extractwasadjusted to 0.02 Methylenediaminetetraacetic acid(EDTA)and sedimentedthrough anED TA-sucrose gradient. (B) control was sedimented through a reticulocyte standard buffer-sucrose gradient.
(C)fractions5to 15of the EDTA gradient indicated by the arrow in Fig. 4A were pooled, dilutedwithone-half
volume of ED TA buffer,anidanalyzed in an EDTA-sucrosegradient. Symbols: (-) trichloroacetic acid-precipitable 3H-uridineradioactivity; (0) trichloroacetic acid-precipitable14C-leucine radioactivity; (-) A280.
to 18 of the gradient shown in Fig. 4C were
pooled,and the RNAwasextracted andanalyzed
in sucrose gradients. Aheterogenous mixture of
ribonuclease-sensitive RNA species were found
with about 22% ribosomal, 3% single-stranded
viral43Sand theremaindersmall RNA species.
The RNA from the prelabeled 2505 peak
sedi-mented only in RSB(Fig.4B)was90%ribosomal
RNA.
Table 1 shows the percentage of 3H-uridine
counts remaining in the 250Sregion ofthe
gra-dient following centrifugationin RSB and
recen-trifugationin EDTA.Followingthe first
sedimen-tation in RSB, 82% of the RNA from the
pre-labeled cells was observed to sediment in the
200S to 3005 region ascompared to 15% inthe
extractsedimentedthroughEDTA.
Recentrifuga-tion of this 250S peak from prelabeled cells in
EDTA dissociated about 90% ofthe remaining
RNAfromthe complex to sediment nearthe top
of the gradient (Fig.
4).
The 250Speak
frompulse-labeled cells containedapproximately 15%
of thetotal RNA in EDTAgradientswhich did
not dissociate from the complex after isolation
andrecentrifugation.
To characterize thespecies of RNAwhich are
contained in thestructureswhich sediment in the
250S regionofthe gradient frominfected
pulse-labeled cells, cultures were infected and were
pulsed with 3H-uridine for 30 minat 16hr
post-infection, and the 250S peak was isolated from
EDTA gradients. The RNA was extracted and
569
on November 10, 2019 by guest
http://jvi.asm.org/
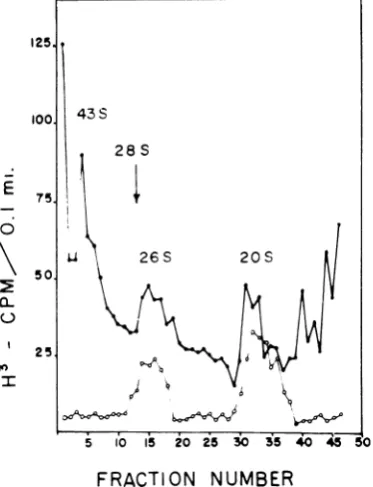
[image:5.500.47.437.67.416.2]analyzed on sucrose gradients (Fig. 5). Three
species of viral RNA areassociatedwiththe 250S
cytoplasmic structure from the infected cells. The
largest species has a sedimentation coefficient of
43, is ribonucleasesensitive,and isassumedto be
[image:6.500.61.256.151.273.2]newly synthesized single-stranded progeny type
TABLE 1. Absence of ribosomes in the replication complexa
Per centRNAremaining in the 250Sregion of thegradient Type ofgradient
Pulse-labeled Prelabeledb infected infected
RSB 15 82
1st EDTA 17 17
2nd EDTA 12 0.01
a RSB, reticulocyte standard
ethylenediaminetetraacetic acid.
bData taken from
Fig.
4.25
loo 43s
28 S
E 75;1
50 26s 2
50.
C)
25.
PO
I I5 Ib\
/1t\
+buffer; EDTA,
FRACTION
NUMBER
FIG. 5.Sucrosegradientanalysisof RNAextracted from viral replication complex. Cultures infected for 16
hr in the presentce of
actinomycint
were pulsed with3H-uridine for 30 miii; extracts were prepared and
centrifuged through ethylenediaminetetraacetic
acid-sucrosegradients.TheRNAinthe250S peak was then
extracted with sodium dodecylsulfate-phenol and an-alyzedon sucrose gradients. Trichloroacetic acid-pre-cipitable counts before (0)andafter(0) ribonuclease treatment. The designation 28S indicates the optical density peak of ribosomal RNA.
RNA. The 26S
partially
ribonuclease-resistantand 205 ribonuclease-resistant forms of viral
RNAwerealso present
(20).
These data lead to the conclusion that viral
RNA is contained in the 250S structure as a
tightly
boundintegral
part of the complex towhich small amounts ofribosomalRNA species
maybelooselyassociated.
RNA polymerase. Since the fast-sedimenting
material in the cytoplasmic extract contained the
replicativeforms of viralRNA,experimentswere
donetodetermineifviral RNApolymerase
activ-ity was present in the 250S particulate fraction.
Cytoplasmicextracts werepreparedfrom cultures
infected for 18 hr and analyzed on sucrose
gra-dients. Polymerase activity in gradient fractions
was determined by measuring the incorporation
of 3H-GTP into acid-precipitable material. As
shown inFig. 6, fractions8to 13 containedmost
of the enzyme activity which sedimented in the
sameregionofthegradientaspulse-labeled RNA
(Fig. 3B). Treatment of extracts with DOC or
Brij-58 either prior to or after gradient analysis
destroyedall detectable enzymeactivity.
SLE viralpolymerase hasproperties similar to
those reported for the group A arboviruses (14,
18) (Table 2). The enzyme required magnesium
ions and all four nucleotide triphosphates and was
inhibited by manganese ions and ribonuclease.
Incorporation of 3H-GTP into acid-precipitable
material was linearduring the first hour of
incu--- 8
74s A
100
-O .~i
t6 1,
C,
-4 C. 3
F,o--tio, N,rmbe,
FIG. 6. Sedimentation pattern of RNA polymerase
in cytoplasmic extract from cells infected with SLE virus. BHK cells wereinfected for 18 hr; cytoplasmic
extractwasprepared and centrifuged through an ethyl-enediaminetetraacetic acid-sucrose gradient. Fractions (1.0 ml) were collected and analyzed for RNA poly-meraseactivity.
1. 2
X.
a. 0
I
8
1 a.
I
ID
"I
:
-6
--, - --- ---I
:1 3.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.500.67.254.305.549.2] [image:6.500.269.452.394.588.2]TABLE 2. Effect ofMII2+, Mg2+,ntcleotides,
actinto-mycini D, ribolnuclease, and adeniosi,ie
triphos-phate-genteratinig system olt Sainlt Loulis
enl-cephalitis viralpolymerase
Total Reactionsystema radio-activityb
Complete assay... 1,572
Complete assay without
Mg2... 315
ATP . 173
CTP 250
UTP . 187
ATP, CTP, UTP 73
Phosphoenol pyruvate and
phospho-enol pyruvate kinase 817
Actinomycin D 1,436
Complete assay with
Ribonuclease (0.5 jAg) 506
Mn'+... ... 346
Uninfected cells (complete assay) 184
aCTP, cytidine triphosphate; UTP, uridine triphosphate.
bCounts per minute of
3H-guanosine
ttiphos-phate incorporated into acid-precipitable ma-terial.
bation and then declined rapidly. Cytoplasmic
extracts of uninfected BHK cells did not
incor-porate a significant amount of 3H-GTP into
acid-precipitable material in the presence of
actinomycinD.
Viral proteins in the replication complex.
Im-munoprecipitationtests wereperformedto
deter-mineiftheprotein associated with thereplication
complex wasviralspecific. Fractions inthe 250S
region of the gradient shown in Fig. 3A were
pooledand reacted withSLEviral-hyperimmune
ascitic fluid. A visible zone of precipitate
devel-oped at the
interphase
between the SLE viralantibodyandfractionswhichcontained the
repli-cationcomplex.Noprecipitatewasobservedafter
incubation of2505 gradient fractions from
non-infected cellswithviral antiserum or when normal
ascitic fluid was used. Most (92%) of the
radio-activity in those fractions which contained the
replication complex was precipitated with SLE
viral antibody, in comparison to less that 10%
fromcytoplasmic fractions of noninfectedcellsor
when normal, mouse ascitic fluid was used. The
replication complex therefore does contain
viral-specific proteins. Whether these proteins are
structural, nonstructural, or
both,
is not known(21).
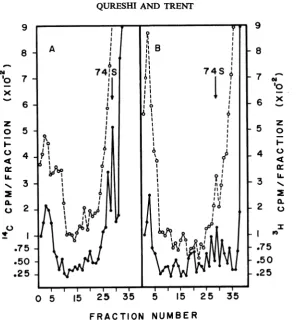
Kinetics oflabelingthereplication complex. The
experimentsdescribed above have established the
existence of a cytoplasmic replication complex
(CRC) in SLE virus-infected cells. To determine
the time in the growth cycle when this complex
was most active in RNA synthesis, cultures
in-fected for 12 hr and 18 hr were doubly labeled
with3H-uridine and '4C-leucine for 15 min.
Cyto-plasmic extracts from these cells were analyzed on
sucrose-EDTA gradients (Fig. 7). It is observed
thatfractions which containmaximal protein and
RNA (trichloroacetic acid-precipitable
radioac-tivity) arecoincident in the gradient. The amount
of "4C-leucine counts in thefractions containing
the CRC from 12-hr(Fig. 7A) and 18-hrinfected
cells (Fig. 7B) is similar, whereas the amount of
3H-uridine radioactivity in the 2505 region from
the 18-hr cultures had increased approximately
twofold. The CRC in SLEviral-infected cells
ap-pears to contain either a constant amount of
structuralprotein, enzymatic protein, or both, or
at 12 hrpostinfectionthe synthesis and release of
protein in the complex has reached a constant
rate. These experiments do not distinguish
be-tween thesepossibilities.However, since the CRC
does not appear to contain polyribosomes and
therefore isprobably notinvolvedinprotein
syn-thesis, we feel that these data probably reflect the
CRCcomposition. The increasedincorporation of
3H-uridine into viral RNA in the replication complex during the viral growth cycle is
concomi-tantwith the increase in totalcell-associated viral
RNA synthesis (20).
DISCUSSION
Pulse-labeled SLE viral RNA is found in a
2505 cytoplasmic structure which contains viral
RNA polymerase activity and the nascent and
replicative forms of viral RNA. These results
sug-gest that this structure is a replicationcomplex,
i.e., the site of SLE viral RNA synthesis. The
replicationcomplexis first detected 6 hr after
in-fection (unpublisheddata) and is most active in
RNA synthesis during the rapid phase of viral
growth when the rate of viral RNA synthesis is
maximal(20).
Input Semliki Forest viral RNA bindsto
mem-braneousstructureswhich have many similarities
tothe SLE viral MRCstructures (8, 17). The
ob-servationthatcycloheximide inhibits the binding
ofSindbis viral RNAtothe MRCstructuresmay
explain our previous observation that
cyclohexi-mide treatment prior to the end of the latent
period inhibits SLE virusgrowth (21).Incontrast
to the polioviral replication complex (9), the
structure and functional
integrity
of the MRCstructures from the Sindbis (17) and SLE viral
CRCare destroyed by
detergent
treatment. Thiswouldsuggestthat the arboviralreplication
com-plex may be composed of subunits which are
membrane associated. Dissociation of these
on November 10, 2019 by guest
http://jvi.asm.org/
9
8
w 0
x
z 0
I-4
IL N.I
0
0L
C1)
7
6
5
4
3
2
.75 .50 .25
9
8
7
^F
'O
6 _
z 0 Z
I-4 0
4
cl:
3
20
2 a.
I
I n
.75 .50 .25
0 5 15 25 35 5 15 2 5 35
FRACTION NUMBER
FIG. 7. Sucrose gradient analysis ofthe replicalion complex pulse-labeled early andlate during viral RNA
synthesis.BHKcellsinfectedfor12hr and 18 hrweresimultaneously pulse-labeledwith3H-uridineand"4C-leucine
for15min; cytoplasmicextracts werepreparedandcentrifuged through ethylenediaminetetraaceticacid-sucrose
gradients. Symbols:(0) 3H-uridine trichloroacetic acid-precipitableradioactivity; (0) 14C-leucinetrichloroacetic
acid-precipitable radioactivity.A, 12 hrpostinfection;B, 18 hrpostinfection.
subunits
during theisolation
procedure mayex-plain
the failure of arboviral RNApolymerase
preparations to
synthesize
progeny-type,single-stranded RNA (14).
Grimley
etal.(10)
havede-scribed adistinct type ofcytoplasmic vacuole in
Sindbis virus infections
which
they
believetobethe site of viral RNA
synthesis.
Many smallervacuoles with similar features are found
in
theperinuclear region of SLE virus-infected cells,
however, their rolein viral RNAsynthesis isnot
now clear(15;unpublisheddata).
In contrast to picornaviral polyribosomes
which are stabletodetergent treatment and can
readily be isolated(9),repeatedattempts toisolate
viral polyribosomesfromSLEviral-infectedcells
using many techniques have
failed
(unpublisheddata).
TheSLE viral CRCdoesnot appear tobeassociated with viral protein synthesis or to
con-tain ribosomes. The cellular site of SLE viral
proteinsynthesis isnot nowknown. Thepresence
of viral
antigen(s)
intheCRCmay beexplainedby theincorporation of viral protein(s) into the
membranes from whichthe complex develops or
associates. TheSindbis viral envelope isacquired
bybudding of the
nucleocapsid
throughcytoplas-mic or internal membranes (10). The envelope
contains a single polypeptide which gives the
virion
hemagglutinating, complement-fixing
andneutralizing antibody-blocking
activities (1). Viralantigen in the SLE viral CRC may reflect the
presence of a viral-modified membrane as a
structuralpartofthisstructure. Antigens inthe
CRCmaybe identicalto theenvelope
polypep-tides of the complete SLE virion (21). Further
experimentsareneededtocharacterizethe
mem-branes associated with the replication of SLE
viral RNA,stiuctural and nonstructuralproteins,
and theenvelopmentprocess.
ACKNOWLEDGMENTS
This investigation was supported by Public Health Service research grantAI-09397 from the National Institute of Allergy andInfectious Diseases.
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.500.113.411.55.376.2]LITERATURE CITED
I. Appleyard, G., J. D.Oram,and J.L.Stanley. 1970.
Dissocia-tionofSemliki Forest virus into biologically active
com-ponents.J. Gen.Virol. 9:179-189.
2.Brandt, W. E., E. L. Buescher, and F. M. Hetrick. 1967. Productionandcharacterization of arbovirus antibody in
mouseasciticfluid. Amer. J. Trop. Med. Hyg. 16:339-347. 3. Caliguri,L.A., andI.Tamm. 1970. The role of cytoplasmnic
membranes inpoliovirusbiosynthesis. Virology 42:100-111. 4. Caliguri, L. A., and I. Tamm. 1970. Characterization of
poliovirus-specific structures associated with cytoplasmic membranes. Virology42:112-122.
5.Eagle, H. 1959. Amino acidmetabolism in mammalian cell cultures. Science 130:432-437.
6. Filishie, B. K., and J. Rehacek. 1968. Studiesof the
morphol-ogy ofMurray Valley encephalitis and Japanese B
en-cephalitis virus growingin cultured mosquito cells. Virology 34:435-443.
7. Friedman, R. M., andI. K.Berezensky. 1967. Cytoplasmic
fractions associated with Semliki Forest virus ribonucleic acidreplication. J. Virol. 1:374-383.
8. Friedman, R. M., and T. Sreevalsan. 1970. Membrane binding cfinputarbovirus ribonucleic acid: effect of interferonor
cycloheximide. J. Virol. 6:169-175.
9. Girard, M.,and D.Baltimore. 1967.Thepoliovirus replica-tion complex: site for synthesis of poliovirus RNA. J. Mol. Biol.24:59-74.
10. Grimley,P.M.,1.K.Berezesky, andR.M. Friedman. 1968.
Cytoplasmic structures associated with an arbovirus
in-fection: loci of viral ribonucleic acid synthesis. J. Virol. 2:1326-1338.
11. Inoue, Y. K.,and R. Ogura. 1962. Studies ofJapanese B encephalitis virus.III. PropagationandassayofJapanese
Bencephalitisinstablelineofporcine kidney cells.Virology 18:500-501.
12. Levin, J. G., and R. M. Friedman. 1971.Analysis of arbo-virusribonucleic acid forms bypolyacrylamidegel electro-phoresis.J. Virol. 7:504-514.
13. Macpharson, I.,and M.Stocker. 1962.Polyoma transforma-tion of hamster cell clones-an investigation ofgenetic factorsaffectingcellcompetence.Virology 16:147-151. 14. Martin, E. M., and J.A.Sonnabend. 1967.Ribonucleicacid
polymerase catalyzing synthesis ofdouble-stranded arbo-virusribonucleicacid. J.Virol.1:97-109.
15. Murphy,F.A., A. K.Harrison, G.W.Gary, S. G.Whitfield, and F. T.Forrester. 1968.St.Louisencephalitisvirus in-fection in mice. Electron microscopic studies of central
nervoussystem.Lab.Invest.19:652-662.
16. Ota, Z. 1965. Electronmicroscopestudy of thedevelopment ofJapanese B encephalitis virus in porcine stable (PS) cells.Virology 25:372-378.
17. Sreevalsan, T. 1970. Associationofviral ribonucleic acidwith cellular membranes in chick embryo cells infected with Sindbis virus. J. Virol. 6:438-444.
18.Sreevalsan, T., and F. H. Yin. 1969. Sindbis virus-induced viral ribonucleicacid polymerase. J. Virol.3:599-604. 19. Summers, D. F., andL. Levintow. 1965. Constitution and
function of polyribosomes of polio virus infected HeLa cells.Virology 27:44-53.
20. Trent, D. W., C. C. Swensen, and A. A. Qureshi. 1969. Synthesis of Saint Louis encephalitis virus ribonucleic acid inBHK-21/13cells.J.Virol. 3:385-394.
21. Trent D. W., and A. A. Qureshi. 1971. Structural and
non-structural proteins of Saint Louis encephalitis virus. J. Virol. 7:379-388.
22. Warner,J., P. Knopf,andA. Rich. 1963. Amultiple ribo-somal structure in protein synthesis. Proc. Nat. Acad. Sci.U.S.A.49:122-126.