Copyright© 1986,American Societyfor Microbiology
Mouse
Peritoneal Cells Confer
an
Antiviral State
on
Mouse
Cell
Monolayers: Role
of
Interferon
ENRICOPROIETTI,1* SANDRAGESSANI,1 FILIPPO
BELARDELLI,'
AND IONGRESSER2Departmentof Virology, Istituto Superiore di Sanita', Rome,
Italy,'
andLaboratory of Viral Oncology,Institut de Recherches Scientifiques surleCancer, Villejuif, France2Received4March1985/Accepted 3June 1985
Vesicular stomatitis virus and encephalomyocarditis virus do not multiply in the majority of peritoneal macrophages freshly explanted from 4- to 8-week-old male or female mice. However, when peritoneal macrophages werecultivatedin vitro for 3 to 5 days,these cells became permissive for both viruses. The loss of antiviral state in "aged" macrophages paralleled a significant decrease in the intracellular levels of
(2'-5')oligo-adenylate synthetase activity. Although biologically active interferon was not detected in the
nutrientmediumofmacrophage cultures, freshly harvested
peritonea!
cells could confer an antiviral state on monolayer cultures of mouse cells (aged macrophages, embryonic fibroblasts, and L cells) but not on heterologous chicken embryo, rabbit kidney, or human cells infected with vesicular stomatitis virus or encephalomyocarditis virus. The conferred antiviral state required at least 7 h to develop in target cells and wastotallyinhibited by the presence of antibody to mouse interferon
a/1
but not tointerferonyin thecocultures.Heterologous guinea pig and rabbit peritoneal cells could not transfer an antiviral state to targetmouse cells. Donorperitoneal cells from mice preinjected with antibody to interferon
a$P
couldnot transfer an antiviral statetotarget mouse cells. This ensemble of results indicating that freshly harvested peritoneal cells transfer interferon(which is responsible forinducing an antiviral state in susceptible mouse target cells)adds
further experimental evidence that interferon is spontaneously expressed in normal mice and plays an important role in maintaining some host cells in anantiviral state.Theresistance of restingmouseperitonealmacrophages to
vesicular stomatitis virus (VSV) and encephalomyocarditis
virus (EMCV) can be abolished by injection of mice with
antibody to interferon ox/, (1). Further evidence that
inter-feron was responsible for the antiviral state of peritoneal
macrophagesstemmedfromrecentexperiments in whichan
inverse correlation was observed between the intracellular
levelsof(2' -5')oligo-adenylate(2-5A)synthetaseactivityin
peritoneal macrophages and the permissivity ofthese cells
for VSV (14).Thusthe level of2-5Asynthetase
activity
washigh inresting peritoneal macrophages from adultmice and
low in peritoneal macrophages from mice injected with
antibody to interferon
od/I
(14). These data stronglysug-gested that injection of mice with antibody to interferon
resultedin theneutralization ofendogenousinterferon
spon-taneously expressed innormal mice. However, most
inves-tigators have not been able to demonstrate biologically
activeinterferonintheperitonealwashingsortissueextr-acts
ofthemajority of mice tested(11,18;J.Galabru,N.Robert,
C.
Buffet-Janvresse,
Y. Riviere, and A. G.Hovanessian,
J.Gen. Virol., in press).
The in vivo resistance ofperitoneal macrophages to
sev-eral animal virusespersists when these cellsarefirstplaced
in culture. Although interferon was not recovered in the nutrient medium ofsuspensions offreshly explanted
perito-neal cells, we could demonstrate that these cells conferred an antiviral state on target mouse cell monolayers in coculture experiments. The evidence indicates that
inter-feronwastransferred fromtheperitonealcells to the
under-lying target cells. The results of these experiments are
presented herein.
* Correspondingauthor.
MATERIALS AND METHODS
Mice. Male and female DBA/2, Swiss, C57B1/6, and BALB/c mice 5 to8weeks oldwereobtained from Charles River, Italia S.p.A. (Milan, Italy). Nude mice (BALB/c genetic background) were purchased from Charles River
Breeding Laboratories, Inc. (Wilmington, Mass.) and kept underspecific-pathogen-free conditions during experiments. Preparation of donormousecells. (i) Total peritoneal cells. The peritoneum was washed with 2.5 ml of RPMI 1640 nutrient medium containing 10% fetal calf serum (FCS;
GIBCO Laboratories, Grand -Island, N.Y.).
(ii) Mouse peritoneal macrophages. Mouse peritoneal cells (106 cellsperml)wereallowedtoattachtoapetriplastic dish (Nunc, Roskilde, Denmark) in 10 ml of RPMI nutrient medium containing 10% FCS for 20 min at 37°C. The monolayer was washed three times with RPMI medium without serum to remove nonadherent cells, the remaining cellmonolayerwastrypsinized, thecellswerecounted, and viabilitywas determined. By this technique, almost 50% of the cellsattachedtotheplastic substrate couldberecovered.
(iii) Thymocytes and lymph node cells. Thymocytes and mesenteric lymph node cells were obtained by teasing and filtering the cellsthrough ametallic grid.
Preparation of target cells. For preparation of virus-permissive macrophage monolayers, peritoneal washings were prepared as described above and seeded in 24-well plastic plates (Nunc), each well containing approximately 106 cells perml. Cellswere allowedtoattach totheplastic culture dish at 37°C for 3 h, and nonadherent cells were discarded. The criteria usedto define these firmlyattached cells as macrophages have been previously described (1). Macrophages were cultivated in vitro for 4 to 5 days, at 456
on November 10, 2019 by guest
http://jvi.asm.org/
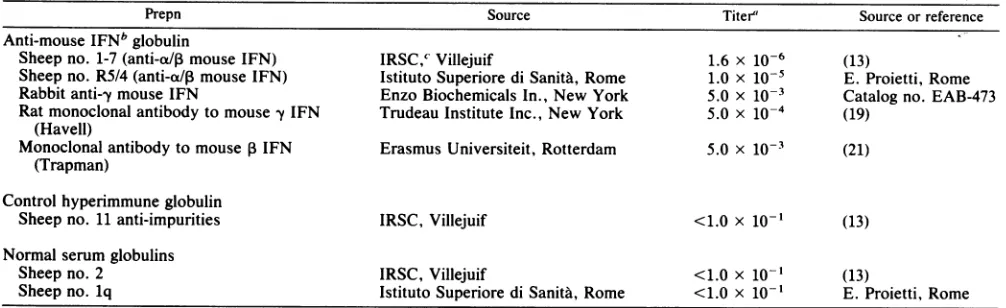
TABLE 1. Source and activity of the different immunoglobulin preparations
Prepn Source Titer' Source or reference
Anti-mouseIFNb globulin
Sheep no. 1-7(anti-a/I mouseIFN) IRSC,c Villejuif 1.6 x 10-6 (13)
Sheep no. R5/4(anti-ap mouse IFN) Istituto Superiore diSanita,Rome 1.0 X 10-5 E. Proietti, Rome Rabbit anti--y mouse IFN EnzoBiochemicals In., New York 5.0 x 10-3 Catalogno. EAB-473 Ratmonoclonal antibody to mouse fy IFN Trudeau Institute Inc., New York 5.0 x 10-4 (19)
(Havell)
Monoclonal antibody to mousePIFN Erasmus Universiteit, Rotterdam 5.0 x 10-3 (21) (Trapman)
Control hyperimmune globulin
Sheep no. 11 anti-impurities IRSC, Villejuif <1.0 x
1o-1
(13)Normal serum globulins
Sheep no. 2 IRSC, Villejuif <1.0 x 10-1 (13)
Sheep no. lq Istituto Superiore di SanitA, Rome <1.0 x 10- E. Proietti, Rome
aNeutralizing titer against 4 to 8 U of mouse interferon. bIFN,Interferon.
cIRSC, Institut de Recherches Scientifiques sur le Cancer.
which time they becamepermissive for VSV and EMCV and
were usedastargetcell monolayers. Monolayer cultures of "aged" macrophages showed the same sensitivity as
monolayer cultures of L cellstothe antiviral action ofmouse
interferona113.
MouseL929cells, human Helaand Wishcells, andrabbit RK13 cells were cultivated in RPMI 1640 medium
supple-mentedwith 10% FCS. Chicken embryo fibroblasts (kindly provided by F. Tat6, Rome) were cultivated in Dulbecco
modified Eagle minimum essential medium (Eurobio, Paris) supplemented with 1% chicken serum, 4% FCS, and 10%
tryptose phosphate broth (Difco Laboratories, Detroit, Mich.). Mouse (Swiss) embryo fibroblasts, obtained from 16-day-old Swiss mouse fetuses, were cultivated in RPMI medium supplemented with 10% FCS, 1% nonessential amino acids (GIBCO), and 1% sodiumpyruvate (GIBCO).
Viruses. Theorigin, methodsofpreparation, andassayof VSV (Indiana strain) and EMCV have beendescribed
pre-viously (1).
Hyperimmune and normal serumglobulins. All serawere
decomplemented and extensively absorbedon murinecells. Theimmunoglobulin fractionswereseparated by ammonium
sulfate precipitation (protein concentration varied between 20 and33 mg/ml) and showntobe devoid ofanytoxicity for mouse peritoneal macrophage. The anti-mouse interferon
ax/, globulins didnotneutralize interferon-y.
Thesourcesandactivities of thedifferentimmunoglobulin preparations arepresentedin Table 1.
Preparation of interferon. Mouseinterferon al3 was
pro-duced, partiallypurified (specific activity, 107reference units
permgofprotein), andassayedaspreviouslydescribed(20). One ofthe unitsasexpressedin thetextistheequivalentof four interferon reference units.
Assay for induction of antiviralstatein targetcell
monolay-ers.(i) VSV yieldassay.A0.1-mlsampleof medium contain-ing freshly harvested total peritoneal cells, macrophages, or othercelltypes(usually 2.5 x 106cells)wasfirst mixed with 0.2 ml of soft agar (Bacto-agar [Difco] 0.33% in 5% FCS-RPMI 1640)topreventthedonorperitoneal cells from adhering to the underlying target cell monolayers. After solidification for5minat4°C,asecondlayer of softagarwas applied. After incubation for 24 h at 37°C, agarand donor peritoneal cells were discarded, and the remaining monolayer was washed three times with medium (2%
FCS-RPMI 1640). A 0.2-ml sample of a viral dilution
(mul-tiplicity ofinfection ofapproximately 2) was added toeach
well. After 1 h of incubation at 37°C the cell sheet was
washedthoroughly, and 1ml of nutrientmediumwasadded.
After incubation for18 h at37°Cin a 5%
C02-air
incubator,thecell-free nutrientmedium wasassayedfor virus in mouse
L929cells.
(ii) Plaque reduction assay. Donor
peritoneal
cells(usually
5 x 106cells) were mixed in 0.2 ml in soft agar as described
above beforeadding toconfluent cell monolayersin35-mm
tissue cultureplates (Nunc). Asdescribedabove, a second
layerof softagar was
applied,
theculturewasincubatedfor24 h, the agar containing donor peritoneal cells was
re-moved, thecellmonolayerwaswashed,and agivennumber
ofplaqueforming units was added(2).
RESULTS
Peritoneal macrophages developpermissivity for VSV and EMCV during in vitro culture. VSV and EMCV do not
multiply in the vast
majority
of peritoneal macrophagesfreshlyexplanted from4- to7-week-old maleorfemalemice
(1). However,when
peritoneal
macrophageswerecultivatedin vitro for 3 to 5 days, these cells became
permissive
forVSV(Fig. 1A) or EMCV (datanot shown).
Inanotherexperimentwedetermined the development of
permissivitytoVSV and the levelof2-5Asynthetase
activity
in peritoneal macrophages (DBA/2 mice) duringthe first 5
days of culture. As
previously
noted (14), there was anexcellent inverse correlation between the level of 2-5A
synthetase activity in
peritoneal macrophages
and thepermissivity
of these cells for VSV. Thus, cultivation ofmacrophages in vitro for5 days rendered these cells
com-pletelypermissive forVSVandwasassociatedwitha20-fold
decreaseinthelevelof2-5Asynthetase
activity
with respect tofreshly
explanted
peritoneal macrophages (data
not shown).Freshly harvested peritoneal cellsconfer an antiviral state onunderlying monolayercultures ofagedpermissive macro-phages. Cultivation of macrophages for 5 days rendered these cells
permissive
for VSV (Fig. 1B). The addition of freshly harvested mouseperitonealcells inducedanantiviral state in the underlying macrophagemonolayer
which was clear cut after 7 h ofcocultivation and became more pro-nounced thereafter(Fig. 1B).on November 10, 2019 by guest
http://jvi.asm.org/
5
4
No
0 3 0
L 2
A
0 1 2 3 4 5 0Z 6b 1U 14 18 22 26 Days ofcultivationinvitro Time (hrs)
FIG. 1. Decayof antiviralstateinperitoneal macrophages during in vitro cultivation (A) and recovery of antiviralstateby cocultivation withfreshly harvested peritoneal cells (B). (A) Peritoneal cells were harvested from theperitoneal cavity of 6-week-old Swiss male mice and seeded in 24-well plastic plates (106 cells per ml ofmedium). Cells wereallowed to fix tothe plastic culture dish at 37°C for 3 h, and nonadherentcells were discarded.Atdifferent days after cellseeding,peritoneal macrophageswereinfected withVSV, and virusyieldswere determined. There werethreemacrophage cultures for each experimental point. (B) Peritoneal cellswereharvested from6-week-old Swiss male mice, andperitoneal macrophageswerecultivated in vitro for 5daystorender thempermissivefor VSV. Peritoneal cells(2.5 x 106,
freshly harvested from theperitoneal cavity of 5-week-old Swissmalemice)wereadded in agarcontaining mediumasdescribed in Materials and Methods for different timesof cocultivation. Agar and donor cells wereremoved, and theunderlying peritoneal macrophageswere infected with VSV. VSVyieldsweredeterminedasdescribed in Materials and Methods. Therewere three macrophage cultures for each point. Symbols: 0,in vitro-aged macrophage monolayer alone; *,peritoneal cells addedtoinvitro-agedmacrophage monolayer.
Table 2 shows theresults of similar
experiments
inwhichperitoneal cells fromdifferent strains of mice were added to
syngeneic in vitro-aged macrophage cultures permissive for
VSV. In each instance, theaddition offreshperitoneal cells
conferred an antiviral state (to VSV or EMCV) in the
underlying monolayercellculture. Induction ofthisantiviral
state did not occur when antibody to interferon aol was included duringthe periodofcocultivation (Table 2).
TABLE 2. Effect of addition of freshly harvested peritoneal cells fromdifferent strains of mice on the multiplication of VSV or
EMCV in monolayer cultures of agedmacrophages" Virus yield/0.2 ml(logl0meant SE)
Strain Challenge Mouse Mouseperitonealcells,
ofmouse virus Controlb peritoneal anti-Interferona/W
cellsadded added
DBA/2 VSV 3.6
±
0.2.0.5
3.8±
0.3SWISS
VSV
3.3±
0.3 0.6±
0.1 3.6±
0.5C57B1 VSV 2.5
±
0.2.0.5
2.7±
0.2BALB/c VSV 3.2
±
0.2 1.6±
0.1 4.1±
0.2 BALB/c VSV 3.4±
0.2 1.7±
0.3 4.2±
0.4nude
DBA/2 EMCV 5.3 + 0.3 2.7
±
0.1 5.5±
0.3aPeritoneal macrophages explanted from different strains of6-week-old male orfemale mice were cultivated in vitro for 4 days to render them
permissive for VSVorEMCV. Syngeneic fresh peritoneal cells(2.5 x 106) from 5 to6-week-old male mice were added asdescribedinMaterials and
Methods. After 18 h, donor peritoneal cells were removed, and target
macrophage monolayerswereinfected with VSV.Therewere three
macro-phage cultures foreachexperimentalsample.
bTarget macrophage cultures alone.
'Sheep1-7antibodytointerferon (x/ (1:1,000, final dilution).
In the experiments described above, amaximal antiviral
state was conferred whenfreshly harvested peritoneal cells
were cocultivated with permissive macrophage monolayer
cell culturesfor12 to18 h before removal oftheperitoneal
cells in the agar overlay (Fig. 1B). It was of interest to
determinewhetherthisperiodof 12 h was necessary for the
underlyingpermissive macrophage monolayertodevelopan
antiviral state or whether it was necessary for the donor
peritoneal cells tobe present for 12h. Accordingly, mouse
peritoneal cellsin agar wereaddedtoprepared macrophage
cultures,and the donor cellswereremovedatdifferenttimes
thereafter. The underlying macrophage cultures were then
furtherincubatedat37°Cfor 12 hbefore infectionwith VSV.
Theresults ofthese
experiments
showedclearlythata5-minperiod of cocultivation offreshly
peritoneal
cells withpre-pared macrophage cellmonolayerswassufficienttoinducea
2-log1o
reduction of VSV yield (compared with controlmacrophages alone).Thisantiviralstate didnot occurwhen
antibody to interferon
at/l
was added to the cell mixture(data notshown).
Freshly harvested mouseperitoneal cells conferanantiviral state on target mouse cells but not on heterologous cells. Freshlyharvested mouse peritoneal cellsconferred an
anti-viral state on target monolayer cultures of in vitro-aged
mouse macrophages,mouseembryo fibroblasts, andmouse Lcells(Table3). This effect was abrogated bythepresence
ofantibody to mouse interferon
a/c
during the period ofcocultivation. Mouse peritoneal cells did not confer an
antiviral state on targetmonolayerculturesofheterologous
chicken embryo fibroblasts, rabbit kidney cells, or human Wish cells. A minimal effect may have occurred in HeLa cells.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.141.473.71.293.2]TABLE 3. Freshly harvested mouse peritoneal cells confer an antiviral state on target mouse cells but not on heterologous cells
VSV yield/0.2 ml MeanVSV plaques/
Addition Cellmonolayertarget (log1omeant SE)' dish(%of
control)b
None Aged mouse macrophage monolayers (5 3.3 ± 0.2
days)
Peritoneal cells 1.0 ± 0.1
Peritoneal cells, antibodyc 3.4 ± 0.3
None Mouse embryo fibroblasts 5.5 ± 0.2 21.5
Peritoneal cells 2.5 + 0.1 2.5 (11)
Peritoneal cells,antibody 6.0 ± 0.4 17.0 (79)
None MouseLcells 52.0
Peritoneal cells 11 (21)
Peritoneal cells, antibody 53 (102)
None Chicken embryo fibroblasts 5.9 ± 0.3 15.5
Peritoneal cells 5.8 ± 0.2 13.5 (87)
Peritoneal cells, antibody 6.0± 0.2 13.5(87)
None Rabbit kidney (RK13) 5.5 ± 0.1 32.5
Peritoneal cells 5.7 ± 0.3 32.5(100)
Peritoneal cells, antibody 5.7 ± 0.1 36.5(112)
None HeLa cells 5.7 ± 0.3 9.0
Peritoneal cells 4.5 + 0.4 5.0(56)
Peritoneal cells, antibody 5.7 ± 0.1 8.5(94)
None HumanWish cells 52.0
Peritoneal cells 38.5(74)
Peritoneal cells, antibody 45.0(87)
a Different types of cellmonolayers (permissive for VSV) were cocultivated with 2 x 106peritoneal cells(freshly harvested from 5-week-oldSwiss female mice). After 18 h at 37°C the donor cells in agar were removed, and the target cell monolayers were infected wih VSV. There were three infectedculturesfor each experimental sample.
bDifferenttypes ofcell monolayers were cocultivated with 6x106peritoneal cells(freshly explanted from 5-week-old Swissmalemice). After18 h at37°C cell
monolayerswerewashed and further processed forVSV plaque assay as described in Materials and Methods. There were twocellcultures for eachexperimental sample.
Sheep 1-7 antibody to interferonal/P(1:500,final dilution).
Relationship between the number ofmouseperitoneal cells
embryo
fibroblasts(Fig. 2A).
An antiviral state did notand theircapacity to confer an antiviralstate.Amounts of 2 x developinmousefibroblastswhen
antibody
tointerferona/I106to 3 x 106peritonealcells wererequiredtoinducea50% wasincorporated into thenutrientmedium
regardless
of theinhibition ofVSV plaques in monolayer cultures ofmouse numberof
peritoneal
cells added.N.ofperitoneal cells per dish(log,O) N.4.5ofperitoneal5 macrophages5.5 perwel6 (log,,)6.5
FIG. 2. Relationship between the number of mouse peritoneal cells and their capacity to confer an antiviral state. (A) Various numbers oftotal peritoneal cells (harvested from 5-week-old Swiss female mice) were cocultivated with confluent monolayers ofmousefibroblasts. After 18 h at 37°C, peritoneal cells were removed, and the fibroblast monolayers were processed for VSV plaque assay. The values are
expressed as the percentage of plaques (mean of four individual cultures) with respect to the mean numberof plaquesincontrol culturesof untreated fibroblasts (58 + 4.1). Symbols: 0, in presence of antibody to mouse interferona/l during the cocultivation period(sheep 1-7; 1:1,000, final dilution). 0, in absence ofantibody during the cocultivation period. (B) Peritoneal macrophages, freshly harvested fromn 5-week-old Swiss female mice, were separated from other peritoneal cells by adherence to the plastic substrate. Aftertrypsinization, different numbers of these macrophages were suspended in agar containing medium and cocultivated with 4-day-old monolayer cultures of macrophages. After 18 h at 37°C the donor cells in agar were removed, and the macrophage monolayers were washed and infected withVSV.
There were three infected cultures for each point.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.136.471.424.631.2]TABLE 4. Capacity ofdifferent celltypes to conferanantiviral state ontarget mousecell monolayer cultures
MeanVSVplaques! VSV
yieldl0.2
ml Expt Donorcells Targetcells dish(%ofcontrol) (Logl0 mean± SE)
1" None Mousefibroblasts 76(100)
Mouse peritonealcells Mousefibroblasts 0
Mouse peritoneal cells, antibody to IFNb Mousefibroblasts 78(103)
Mouse thymocytes Mouse fibroblasts 33(43)
Mouse thymocytes, antibody to IFN Mouse fibroblasts 90(118)
2C None Aged in vitro mouse macrophage cultures 2.8± 0.3
Mouse peritoneal cells Agedin vitromousemacrophage cultures c0.5
Mouseperitoneal cells,antibodyto IFN Aged in vitro mouse macrophage cultures 2.6± 0.1 Guineapig peritoneal cells Agedinvitromousemacrophage cultures 2.5± 0.1 Guineapig peritoneal cells, antibody toIFN Aged in vitro mouse macrophage cultures 2.5± 0.3 Rabbit peritoneal cells Aged in vitromouse macrophage cultures 3.0 ± 0.2
3d
None Agedin vitromouse macrophage cultures 4.5 ± 0.1Mouseperitoneal cells from normal Aged invitromouse macrophage cultures 1.8 ± 0.4 untreated mice
Mouse peritonealcellsfrommice Aged in vitro mouse macrophagecultures 4.9 ± 0.4 preinjectedwith antibodyto IFN
Peritonealcells andthymocyteswereexplanted from 6-week-old Swissmale mice, and 5 x 106 cellswerecocultivatedwith mouseembryo fibroblastsas
describedinMaterials and Methods. Insome cell cultures antibodyto mouseinterferon ox/p(sheep 1-7;final dilution, 1:500)was added.After 18 h at37°C fibroblast monolayerswerewashedandfurtherprocessed forVSVplaque assay.There werefour dishes foreachexperimental condition.
bIFN, Interferon.
' Peritoneal macrophages (from 6-week-old Swissmalemice)cultivated in vitro for4daysweretreated with2 x 106 peritoneal cells as indicated. After 18 h at
37'Cperitoneal cellswereremoved,and cellmonolayerswereinfectedwithVSV. Therewerethreeinfectedcell cultures for eachgroup.
"Peritoneal macrophages (from 7-week-old Swiss male mice) cultivated in vitro for4daysweretreatedwith 2.5 x 106 peritoneal cellsfreshlyharvestedform 8-week-oldSwissmale mice(either untreatedorpreinjectedintraperitoneallywithantibodytointerferon4daysbefore [sheep1-7;diluted1:10,0.2 ml permouse]). After18 h at37°C, peritoneal cellswereremoved, and cell monolayers wereinfectedwithVSV.There were three infected cellculturesfor eachgroup.
Figure 2Billustrates the results of an experiment in which
weused different numbers of freshly harvested mouse peri-toneal macrophages (separated from those periperi-toneal cells which did not adhere firmly to the plastic substrate). Over
98% of the attached cells were stained for nonspecific
esterase(1). When these cells were detached by trypsin and cocultivated with aged monolayer cultures of macrophages
permissivefor virus replication, less than105donor cells was
sufficient to induce 50% inhibition of VSV yield. These
results indicated that peritoneal macrophages constitute a veryefficient cell population for the transfer of the antiviral state.
Type of cells capable of conferring an antiviral state on mousecellmonolayers permissive forVSV. It wasof interest todetermine whether other types offreshlyexplanted mouse
cells and cells from other species could also confer an
antiviralstate on mousecell monolayers. Mouse thymocytes
conferredonly aslightinhibition on mouse fibroblasts
com-paredwith mouseperitoneal cells (experiment 1 in Table 4).
Likewise, mouse mesenteric lymph node cells conferred
only a slight antiviral state on mouse fibroblasts (data not
shown). In contrast, heterologous guinea pig and rabbit
peritoneal cells (experiment 2 in Table 4) did not induce an
antiviral state in underlying mouse macrophage monolayer cultures. Likewise, cocultivation of RK13 cells with mouse embryonicfibroblasts was notassociated with the induction
ofan antiviral state (data not shown).
Peritoneal cells from mice preinjected with antibody to
interferon did not confer an antiviral state on underlying target mousemacrophage monolayer cultures (experiment 3 in Table 4).
Induction of the antiviral state by mouse peritoneal cells is specifically abolished by antibodies to mouse interferon
a/P.
The above experiments showed that the capacity of perito-neal cells to confer an antiviral state on target monolayercultures of mouse embryonic fibroblasts or aged mouse macrophages was abrogated when sheep 1-7 antibody to interferon
o/,I
wasincorporated in the coculture medium. It was ofinterest to test the specificity of this effect of antibody and to determine the minimal amount of neutralizing anti-body required. Two different sheep antibodies to mouse interferon a/1 (including a purified fraction) were highly effective in abrogating the capacity of mouse peritoneal cells to induce the antiviral state in target mouse cells, whereas polyclonal rabbit and monoclonal rat antibody to mouse interferon y, sheep antibody to the impurities in the inter-feronoJ/I
preparations, and normal sheep immunoglobulin were ineffective (Table 5). A rat monoclonal antibody to mouse interferon fi exerted some neutralizing effect, al-though this effect was much less marked than that afforded by the same number of neutralizing units of antibody to interferon ox/,.Figure 3 shows the relationship between the amount of neutralizing antibody of sheep 1-7 in the coculture and the extentof the antiviral state induced by donor peritoneal cells
onmonolayercultures ofmouseembryonic fibroblasts(Fig.
3A) and macrophages (Fig. 3B). DISCUSSION
Resting peritoneal macrophages from normal mice were resistant to VSV and EMCV in vivo (1) and when first placed in culture, as shown herein. The evidence indicating that interferon was responsible for this antiviral state may be summarizedasfollows. First, peritoneal macrophages taken from mice injected with antibody to mouse interferon ot/l
(but not injected with control immunoglobulins) were fully permissive for these viruses (1). Second, 2-5A synthetase activity was considered a sensitive marker of interferon activity (14), and we found an excellentinverse correlation between the level of2-5A synthetase activity of peritoneal
on November 10, 2019 by guest
http://jvi.asm.org/
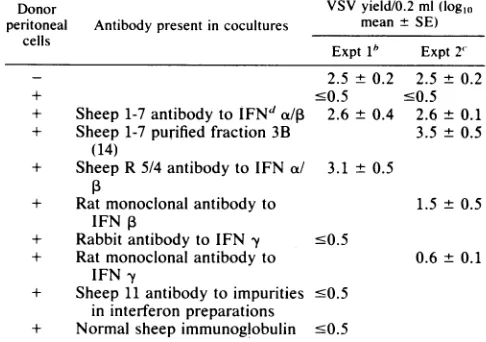
TABLE 5. Effect of different antibodies on the capacity of peritoneal cells to induce an antiviral state in peritoneal
macrophages permissive forVSVa
Donor VSVyield/0.2 ml(logl,
peritoneal Antibody presentin cocultures meant SE) cells
Expt1" Expt 2' 2.5 ± 0.2 2.5 ± 0.2
+ '0.5 '0.5
+ Sheep1-7antibody to
IFNd
do/, 2.6 ± 0.4 2.6 ± 0.1 + Sheep1-7purified fraction 3B 3.5 ± 0.5(14)
+ SheepR 5/4 antibodytoIFNa/ 3.1 + 0.5
+ Ratmonoclonalantibody to 1.5 ± 0.5 IFN P
+ Rabbitantibody to IFN y -0.5
+ Ratmonoclonal antibody to 0.6 ± 0.1 IFN y
+ Sheep11antibodytoimpurities .0.5 ininterferon preparations
+ Normal sheepimmunoglobulin 0.5
'Peritoneal macrophages explanted from5 to 6-week-old Swissmalemice
werecultivated invitro torender them permissiveforvirus replicationand usedastargetmonolayer cultures. After4days.macrophageswere
coculti-vatedat37°Cwith2 x106peritoneal cells freshly explantedfrom5-week-old Swiss male mice. The neutralizing titers of the anti-interferon globulin preparations(finaldilutionintheexperimental test)wereasfollows.
Experi-ment1:anti-a/13 mouseinterferon globulin (sheep1-7),2 x 103against 8 Uof
a/1 mouseinterferon; anti-ax/1 mouseinterferonglobulin (sheepR5/4). 1 x
103 against 8 U ofcx/1 mouse interferon; rabbit anti-mouse y interferon globulin, 1x 103against 8U of mouseyinterferon. Experiment2:anti-a/,8
mouse interferon globulin (sheep 1-7), 4 x 10' against 8 Uofc/P13 mouse
interferon;anti-a/13mouseinterferon globulin (sheep1-7;fraction3B), 3x103
against 8 U of cs/1 mouse interferon; antibody to y mouse interferon (monoclonalantibody),2.5x 103against10Uof mouseyinterferon;antibody
to1 mouse interferon (monoclonal antibody), 5x 102against 5U of1Bmouse
interferon. After18 hofcocultivation at37°C,peritonealcellswereremoved,
andmacrophageswereinfectedwithVSV. Virusyieldsweredeterminedas
described in MaterialsandMethods.
hThere were three infected macrophage cultures for each experimental
condition.
'There were five infected macrophage cultures for each experimental condition.
dIFN,Interferon.
macrophages and their permissivity for VSV (see above).
Nevertheless, we have been unable to recover biologically
active interferon in theperitoneum ofnormal miceorinthe
nutrient medium of cultures of peritoneal macrophages.
Galabru and co-workers (in press) reported low levels of
circulatinginterferon in14of 160 normalmice.The
question
remains, therefore, whether interferon is present in most
normalmice and whether it isresponsible fortheseemingly
inherent antiviral state ofresting peritoneal macrophages
andprobablyother cells ofthenormalmouse.
Thefollowingresults presented hereinarguestronglythat
interferonisresponsible for this antiviralstate.
(i)Mouseperitoneal cellsareresistanttoVSV andEMCV
when first placed in culture and show high levels of2-5A
synthetase activity. After 4 to 5 days in culture these cells become permissive to these viruses, and the level of 2-SA
synthetase activity
declines.
(ii) Freshly harvested mouseperitoneal cells (and proba-bly macrophages constitute the most effective population;
Fig. 2B)canefficientlytransfer this antiviralstate to
permis-sive target cells, provided they are also of mouse origin
(monolayer cultures of aged macrophages, embryonic
fibro-blasts, or L cells). The antiviral state cannot be conferred
efficiently or at all on heterologous cells known to be
insensitive to mouse interferon, such as chicken embryo,
and rabbit kidney, and human HeLa and Wish cells (Table
3).
(iii)Thekinetics of transfer of the antiviral state(Fig. 1B)
are in accord with previous observations by Dianzani and Baron(8) on thetime necessary for exogenous interferonto induce a full antiviral state in susceptible cells.Furthermore,
the experiments show that cocultivation ofperitoneal cells
with target cellsforonly a few minutes suffices to transferan antiviral state, provided that sufficient timeis subsequently
permitted forthe development of the antiviral state in the
target cells.
(iv) Transfer of the antiviral state was in all instances
completely abolished by the addition ofantibody to
inter-feron al/1 in the mixed cell cultures.
(v) Heterologous guinea pig or rabbit peritoneal macro-phages cannot transfer an antiviral state to mouse cells (Table 4).
(vi) Peritoneal cells from mice injected with antibody to interferon ao/1, which are permissive for VSV (1), cannot transfer the antiviral state to target mouse cells (Table 4).
The failure of two different preparations of antibody againist mouse interferon y to inhibit the phenomenon, the incomplete effect of antibody to interferon 1, and the com-pleteinhibition afforded by two preparations of antibody to interferon ac/ indicate that the transfer of the antiviral state is mediated by interferon a or
P
or both, but not by interferony.
This ensemble of results suggests that mouse peritoneal cells contain biologically active interferon (either intra-cellularly oratthe surface) and that these cells transfer this interferon to susceptible mouse cells. Furthermore, the experimental resultsindicate clearly that the peritoneal cells transfer interferon rather than inducing the production of interferon in the target cells. Nevertheless, it is somewhat surprising to us that, in view of the extent of the antiviral effect induced in the target cells by peritoneal cells resus-pended in agar, we have not been able to recover any interferon in the nutrient medium of cocultures. It seems possible thatthe mechanism of transferof the antiviral state by macrophages is more effective than might be suspected, as was suggested many years ago by Glasgow (9).
There have been a few reports of the transfer of the antiviral state in coculture. In the experiments of Blalock and co-workers (3-5), the transfer of the antiviral state occurred between interferon-treated cells and heterologous targetcells and was notinhibited by the presence of antibody to interferon. Their phenomenon appears to be totally dif-ferent, therefore, from the one we have described. Ito and co-workers (15) showed that mouse peritoneal cells from normal orgermfree mice alsotransferred an antiviralstateto homologous mouse cells (but not to heterologous human cells), and the establishment of this antiviral state was partially blocked by the presence of antibody to type I interferon. They concluded, therefore, that interferon was the responsible factor. Soldateschi and co-workers (D. Soldateschi, D. Boraschi, and A. Tagliabue, Antiviral Res., in press) found that co-culture of mouse peritoneal cells and mouse embryonic fibroblasts resulted in an inhibition of EMCV multiplication, but they concluded that this effect was not mediated by interferon because there was not a direct correlation between anti-EMCV activity and inter-feron production. The recent experimental results of Vogel and Fertsch (22) are inaccordwith the observation ofIto et al. (15) and ourselves. They showed that the treatment of C3H/HeN mouse peritoneal macrophages in vitro with
on November 10, 2019 by guest
http://jvi.asm.org/
A
.n
>n 1
B
Neutralizingtiterofantibody(logO) titerofantibody (loglo)
FIG. 3. Capacity of different amounts of antibody to interferon ot/, to abrogate the antiviral state in mouse target cells induced by peritoneal cells. (A) Confluent monolayers of mouse embryo fibroblasts were cocultivatedwith 5x 106 peritoneal cells (freshly explanted from 5-week-old Swiss male mice) in the presence of different amounts of antibody to interferon a/,B. After 18 h at 37°C, peritoneal cells were removed, and mouse fibroblasts were processed for VSV plaque assay. There were four fibroblast cultures for each experimental condition. Thevalues are expressed as percentage of plaque (means of four individual cultures) with respecttothe meannumber of plaques in control untreatedfibroblasts (42±2.5). No plaqueswere detected in fibroblast cultures cocultivated withperitonealcells in theabsence of antibody. Theratio of peritoneal cells to embryonic fibroblasts was2.5:1.(B)Peritoneal macrophages explanted from 8-week-old Swiss male mice were cultivated in vitrotorender thempermissive for VSV. After4days, macrophages were cocultivated with 3 x 106 peritoneal cells, freshly explanted from 6-week-old Swiss male mice,inthe presence ofdifferent amounts of antibodytointerferon
a/P.
After 18hat37°C,peritoneal cells were removed, and macrophage cultures were infected with VSV. There were three macrophage cultures for each experimental condition. The ratio of peritoneal cellstomacrophage cultureswas 10:1.bodytointerferonot/agreatly reduced the capacity of these
macrophages to phagocytose opsonized erythrocytes.
Fur-thermore, they demonstrated that this phagocytic activity
could be transferred in coculture from C3H/HeN
macro-phages to macrophages from endotoxin-hyporesponsive
C3H/HeJ miceandconcluded thatthiseffect wasmediated
by interferon (22).
Although thereis accumulating evidencethatinterferon is spontaneously expressed under some conditions in normal
hunians(16), rabbits(7),and mice(1,11, 14, 15, 22; Galabru
et al., in press), several important questions remain to be
answered. Asregards mice, dothe peritoneal cellsproduce
interferon or do they bind interferon which is produced
elsewhere?Isinterferonproducedconstitutively byavariety
of host cells in normal mice, or is it continually induced
because ofthebacterial florainmicein thealimentarycanal
assuggestedby theexperiments ofGalabruetal. (inpress)?
Whatever the mechanism, the presence of low levels of
interferon in normal animals appears to be an integral componentin the hostdefenseagainstviralinfection. It may well be that these low levels of interferon before viral infection arefar moreimportant for the host than the much greater amounts of interferon which are produced in the course of viral infection. Likewise, it is possible that low levelsofendogenous interferon alsoexertothereffects in the
animal, such as restricting the growth of neoplastic cells.
Thus, injection of mice with antibody to interferon
ct/n
markedly enhanced the tumorigenicity of xenogeneic (17)
andsyngeneic(11) tumor cells. It remains to be determined
whether these low levels of interferon also play a role in regulating the function of certain host cells, as well as the
expression of certain surface antigens,suchas
histocornpat-ibility antigens, which are known to be affected by
exogenousinterferon(10). Ourexperimental resultssupport
a hypothesis proposed by Bocci (6) of a "physiological
interferon response" whichismanifested bytheproduction
of interferon "at specific sites" and which "can maintain active defense systems essential for survival."
ACKNOWLEDGMENTS
This work was supported in part by grants from Consiglio Nazionale delle Ricerche (Progetto Finalizzato Controllo delle Malattie da Infezione No. 83.02916.52 and Progetto Finalizzato OncologiaNo. 84.00498), theRichardLounsbery Foundation, the SimoneetCinodel DucaFoundation,theDirection de Recherches et Techniques (contract 82-420), the Association pour le Develop-pement de la Recherche sur le Cancer at Villejuif, the Institut National delaSanteetdela RechercheMedicale (contract 127 012), and the Foundationpourla RechercheMedicale
Frangaise.
We are indebted to the skillful technical assistance of L. Santurbano.
LITERATtJRECITED
1. Belardelli, F., F. Vignaux, E. Proietti, and I. Gresser. 1984. Injectionofmicewithantibodytointerferonrendersperitoneal macrophages permissivefor vesicularstomatitis virus and en-cephalomyocarditis virus. Proc. Natl. Acad. Sci. USA 81:602-606.
2. Benedetto, A.,G. B.Rossi,C. Amici,F. Belardelli,L.Cioe,G. Carruba, and L. Carrasco. 1980. Inhibition of animal virus products by means of translation inhibitors impermeable to 2c
0 u
z
NZ
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.163.456.70.295.2]normal cells. Virology 106:123-132.
3. Blalock, J. E., and S. Baron. 1977. Interferon-inducedtransfer of viral resistance between animal cells. Nature (London) 269:422-425.
4. Blalock, J. E., and S. Baron. 1979. Mechanisms of interferon-induced transfer of viral resistance between animal cells. J. Gen. Virol. 42:362-372.
5. Blalock, J. E., and G. J. Stanton. 1978. Efficient transfer of interferon-induced virus resistance between human cells. J. Gen. Virol. 41:325-331.
6. Bocci, V. 1981. Production and roleof interferon in physiologi-calconditions. Biol. Rev.56:49-85.
7. Bocci, V., M. Muscettola, L. Paulesu, and G. Grasso. 1984.The physiological interferon response. II. Interferon is present in lymph but not in plasma of healthy rabbits. J. Gen. Virol. 65:101-108.
8. Dianzani, F., and S. Baron. 1975. Unexpectedlyrapid action of humaninterferon inphysiological conditions.Nature(London) 257:682-683.
9. Glasgow, L. A. 1970. Transfer ofinterferon-producing macro-phages: new approach to viral chemotherapy. Science 170:854-856.
10. Gresser, I. 1984. The effect of interferonontheexpression of surface antigens, p. 113-132. In J. Vilcek and E. De Maeyer (ed.), Interferons and the immune system. Elsevier/North-HollandPublishing Co., Amsterdam.
11. Gresser, I., F. Belardelli, C. Maury, M. T. Maunuory, and M.G. Tovey. 1983.Injection of mice with antibodytointerferon enhancesthegrowth oftransplantable murine tumors.J. Exp. Med. 158:2095-2107.
12. Gresser, I., C. Bourali, M. T. Thomas, and E. Falcoff. 1968. Effect of repeated inoculation of interferon preparations on infection of mice with encephalomyocarditis virus (32723). Proc. Soc. Exp. Biol. Med. 127:491-496.
13. Gresser, I., M. G. Tovey, M. T. Bandu, C. Maury, and D.
Brouty-Boye. 1976. Role of interferon in the pathogenesis of virus diseases in mice as demonstrated by the use of anti-interferon serum. I. Rapid evolution ofencephalomyocarditis virusinfection. J. Exp. Med. 144:1305-1315.
14. Gresser, I., F. Vignaux, F. Belardelli, M. G. Tovey, and M. T. Maunoury. 1985. Injection of mice with antibody to mouse interferon a/,Bdecreases the level of 2'-5'oligo-adenylate syn-thetase inperitonealmacrophages.J. Virol.53:221-227. 15. Ito, Y., H.Aoki, Y. Kimura, M. Takano, K. Shimokata, and K.
Maeno.1981. Naturalinterferon-producingcells in mice. Infect. Immun.31:519-523.
16. Lebon, P., S. Girard, F. Thepot, and C. Chany. 1981.Presence constante de interferon a dans les liquides amniotiques humains.C.R. Acad. Sci.293:69-71.
17. Reid, M. L,N. Minato,I. Gresser, J. Holland, A. Kadish, and B. R.Bloom. 1981. Influence of anti-mouse interferon serum on thegrowthandmetastasis oftumorcells.Proc.Natl.Acad. Sci. USA78:1171-1175.
18. Sen, G. S. 1982. Mechanism of interferon action: progress toward itsunderstanding. Prog. Nucleic AcidRes. Mol. Biol. 27:105-156.
19. Spitalny,G. L., and E. A. Havell.1984. Monoclonal antibodyto murinegammainterferon inhibitslymphokine-induced antiviral and macrophage tumoricidal activities. J. Exp. Med. 159:1560-1565.
20. Tovey, M.G., J. Begon-Lours, and I. Gresser. 1974. Amethod forthelarge scaleproduction ofpotent interferonpreparation. Proc. Soc. Exp. Biol. Med. 146:809-815.
21. Trapman, J. 1983. Distinctantigenic characteroftwo compo-nentsofpoly(I)-poly(C)-inducedmouse Lcellinterferon. FEBS Lett. 109:137-140.
22. Vogel, S. N., and D. Fertsch. 1984. Endogenous interferon production by endototoxin-responsive macrophages provides an autostimulatory differentiation signal. Infect. Immun. 45:417-423.