0022-538X/91/116320-05$02.00/0
Cloning, Sequencing,
and Functional
Analysis of
a
Marek's Disease
Virus
Origin of DNA
Replication
HEIDI S. CAMP,1 PAUL M. COUSSENS,1 ANDROBERT F. SILVA2*
Molecular VirologyLaboratory, Department of Animal Science, MichiganState University,EastLansing, Michigan
48824,1
andAvianDisease andOncology Laboratory,
AgriculturalResearchService, U.S.Department ofAgriculture,3606 East MountHope Road, EastLansing, Michigan488232
Received6May 1991/Accepted30July 1991
Previously, we isolated a replicon from a defective Marek's disease virus (MDV), analogous to defective herpes simplex viruses (amplicons). Defective viruses containcis-actingelementsrequiredfor DNAsynthesis
andviruspropagationsuch as anoriginof DNAreplicationand apackaging-cleavage signalsite. In thisreport, the MDVrepliconwas utilized to locate anorigin ofMDVDNAreplication. A comparisonof MDVreplicon sequences with other herpesvirus replication origin sequences revealed a 90-bp sequence containing 72%
identity to thelytic origin
(oris)
ofherpessimplexvirustype1. This90-bp sequence displayednosimilaritytobetaherpesvirus or gammaherpesvirus replication origins. The 90-bp sequenceis arranged as an imperfect palindrome centered around an A+T-rich region. This sequence also contains a 9-bp motif (5'CGTTCG CAC3') highlyconserved in alphaherpesvirus replication origins. To test functionalityof the 90-bpputative MDVreplicationorigin,weconductedDpnI replication assays with subclonesgeneratedfrom the4-kbpMDV
replicon.A700-bpMDVreplicon subfragment containingthe90-bpputativeMDVreplicationorigin sequence
iscapableofreplicatinginchickenembryofibroblast cells cotransfected withhelpervirus DNA. Inconclusion, weidentified afunctionalorigin ofDNAreplicationin MDV. Similarityof MDVorigin sequencestothoseof
alphaherpesviruses supportsthe currentcontentionthat MDV is moreclosely relatedto alphaherpesviruses
than togammaherpesviruses.
Marek's disease virus (MDV) is a highly cell-associated avian herpesvirus. Inchickens, MDV is the etiologic agent ofMarek'sdisease, amalignant T-celllymphoma(7). There are three MDV serotypes. Serotype 1 includes oncogenic MDVsand their attenuated derivatives, serotype 2 includes the closely related, naturally occurring nononcogenic chicken herpesviruses, and serotype 3 is the antigenically related nononcogenic turkey herpesviruses. MDV pathology has beenextensively characterized (7, 23). Molecular anal-ysis of MDV, however, has lagged behind that of other herpesviruses, primarily because of technical difficulties presented by the tightly cell-associated nature of MDV infection.
The MDV genome, a double-stranded linear DNA mole-cule of approximately 160 to 180 kbp, consists of a unique long (UL) and a unique short
(Us)
segment flanked by inverted repeats (TRL, IRL,IRS,
TRS). MDV has been classified as a gammaherpesvirus based on its lymphotro-pism (25). However, the overall genomic structure and colinearity of many MDV genes to those of alphaherpesvi-ruses such as herpes simplex virus (HSV) andvaricella-zostervirus suggest that MDV should be reclassified (5). Previously, we reported the isolation and characterization of a defective serotype 2 MDV (8). The defective MDV (replicon)genomeexists as a high-molecular-weight head-to-tail concatemer consisting of 4-kbp viral monomeric repeats and appears to be analogousto HSV amplicons (12). A 4-kbp repeat was cloned into the EcoRI site in pUC19 and was designated pA5 (8).
HSV amplicons contain multiple head-to-tail reiterations of monomeric repeat units derived from either end of the
Us
fragment orfrom two noncontiguous regions within the
Us
* Corresponding author.
andULfragmentsof the HSV genome(11). HSV amplicons replicateand propagate in the presenceofahelpervirus(11, 12, 19, 30, 34, 35). Molecular analysis of HSV amplicons revealed three lytic replication origins (oriL andtwocopies
of
oris)
andapackaging-cleavage signalsite within the HSVgenome (11, 12, 30, 31, 33-35).
DNA sequencing. DNA sequencing of the 4-kbp MDV replicon was initiatedtoidentify a potential origin of repli-cation in MDV. pA5Iwas constructed by cloningthe 4-kbp replicon into pUC19in theinverse orientationwith respect to pA5. Unidirectional deletion mutants were created from pA5 andpA5I with exonuclease III and Si nuclease (Boehr-ingerMannheimBiochemicals,Indianapolis, Ind.), and then deleted cloneswere recircularized with T4 DNA ligase (16). DNAsequencingwasperformedwithdouble-stranded plas-mid templates and the dideoxy chain termination method (26). In all sequencing reactions, the Sequenase enzyme (United States Biochemical Corp., Cleveland, Ohio) was usedas recommended bythe manufacturer. MDV replicon sequences were compared with sequences in the GenBank data base with the MacVector (International Biotechnolo-gies, Inc., NewHaven, Conn.) computer program.
A90-bp A+T-rich sequence, closely related to HSV and varicella-zoster virus replication origin sequences, was lo-cated betweenaPstI-to-BdnHI site of pA5 (9, 20, 33) (Fig. 1A). Overall, the 90-bp sequence contains 72% sequence identityto core regions of the HSV-1
oris
andoriL and 66% sequence identity to origins of varicella-zoster virus and equineherpesvirus 1 replication (2, 9, 20, 24, 32, 33) (Fig. 1B). Neither the replication origins of Epstein-Barr virus (oriP and oriLyt) nor those of cytomegalovirus contained sequences similarto theputative origin of serotype 2 MDV replication (1, 13, 14, 15, 37).The 90-bp sequence is arranged in an imperfect palin-dromecontaining30bp of alternatingATresidues, whereas 6320
on November 10, 2019 by guest
http://jvi.asm.org/
A. NJV ori
10 20 30 40 50
A0XCICAC G(iiGCACCG CGAACCAATA TAPGATTATA TATATAATAT
60 70 80 90
ATIATGCG
CAGG'IGA_ACCCCrC
03CAATICGGG
10 20 30 40 50 60 70 80
B. MDV ori ACGCGTCAGC GTTCGCACCG CGAACCAATA TAAGAITATA TATATAATATAITATTGGCG CAAGGTGCGA ACGCCCGTCC GCGCAATCGG
HSV-1 oris ACGCG-.AGC GITCGCAC.. CG..CCAATA T---A-TATA TATAT----T ATrA- .GGCG -AA-GGTGCGA AC...CG.C. GC.
HSV-1 oriL ACGCG-.AGC GTTCGCAC. G-.CC.A-A TAA--- TATA TATAT ----T ATTA--GG-. CAA.IGTGCGA ACG- -C.TC. G
VZV ori ---.CA.C GTTCGCAC...ATA T-A.A-TATA TATAT-ATAT A-TAT.G.-G -AA.G.G.GA ---- GT
EHV-1 oris -.G.G. -..C GTTCGCACC. ...ACCA- -A TAA-.T-.-. T-.ATAAT-. ATTATT... CAA.GGTGO3GA AC.
FIG. 1. Identification of a putative origin of MDV serotype 2 replication by DNA sequence analysis of the MDV replicon. DNA sequencingwasperformed by the dideoxy chain termination method. (A) Nucleotide sequence of the 90-bp putative origin of MDV serotype 2 replication. Black bars indicate a 9-bp sequence shown to be recognized by the HSV-1 origin-binding protein. (B) Alignment of MDV serotype 2replication origin withalphaherpesviruslytic origins ofreplication. Black bars indicate highly conserved sequences among other origins of herpesvirus replication. Dots (.) represent mismatched sequences, and dashes (-) indicate gaps.
the
oris
andoriL
of HSV contain a nearly perfect palindrome with an 18-bp A+T-rich region (9, 33). Lockshon and Galloway (20) reported that the palindrome center, contain-ing the 18-bp A+T-rich sequence, is essential for HSV type 2 (HSV-2) replication. However, expansion of the 18-bp A+T-rich region to 52 bp by introducing alternating AT sequences did not abolish the HSV-2 replication function (20). Interestingly, theputative MDV replication origin also contained a 9-bp sequence (CGTTCGCAC) that is highly conservedinalphaherpesvirus replication origins (2, 24, 32, 33). This 9-bp sequence is a subset of an 11-bp motif (CGTTCGCACTT)shown to be recognized by the HSV type 1 (HSV-1) origin-binding protein, the product of UL9 gene expression (10, 18). Recently, Bruckner et al. (4) reported that the HSV-1 origin-binding protein contains helicase activity, possibly used for unwinding DNA duplex at the origin. Othershaverecentlyreportedthat properinteraction of origin-binding protein withoris,
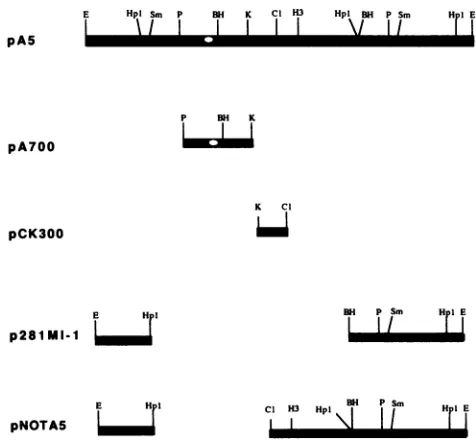
containing the 11-bp motif, isessential forDNA replication(17).Plasmid replication. To determine whether the putative MDV origin, identified by DNA sequence analysis, could function as areplication origin, we constructed a series of subclones from pA5 (Fig. 2). pNOTA5 was derived from pA5 byreplacing a2.5-kbp HpaI and ClaIfragment of pA5 withNotIlinkers(agiftfrom A. Finkelstein). p281MI-1 was constructed by cloning a 2-kbp BamHI fragment from the 281MI/1 genome into pUC18 at the BamHI site.
pCK300
containsa300-bp ClaI-to-KpnI
subfragment
ofpA5
inserted between the ClaI and KpnI sites inpUC18.
A700-bp
PstI-to-KpnI subfragment of pA5 was isolated and cloned intopUC18 byusingthe PstI andKpnIsitestocreate
pA700.
All pA5 subclones as well as pA5 were
propagated
in Escherichia coli DH5a competentcells,
aDam'
E. coli strain.Also,the4-kbpEcoRIreplicon
fragment
wasisolated frompA5andreligated.
We conducted DpnI resistance assays
(33)
onpA5
sub-clones as well as the circular form of the4-kbp
replicon.
DpnI cleaves only
methylated
GATC sequences;therefore,
DNApropagated from
Dam'
strainsofE. coli ismethylated
at the adenine residues within GATC and
susceptible
to DpnI digestion. DNAreplicated
ineucaryotic
cells is not methylatedatGATC sequences and is therefore resistantto DpnI cleavage.High-molecular-weight
DNA was isolated from chicken embryo fibroblast(CEF) cells infectedwith281MI/1
passage15, aserotype 2 MDV(36). Preparation andmaintenanceof primary or secondary CEF cultures have been previously described (28). Virus stocks were determined to be free of reticuloendotheliosis and avian leukosis viruses by either complement fixation or enzyme-linked immunosorbent
as-say(29). Upon evidenceof extensivecytopathiceffect,cells
were lysed in 150 mM NaCl-100 mM EDTA-1% sodium dodecyl sulfate-100 ,ugofproteinase Kpermlfor 4 to24h
at 37°C. DNAwasextracted twice withanequalvolume of phenol-chloroform-isoamylalcohol(25:24:1,vol/vol/vol)and
once with chloroform-isoamyl alcohol (24:1,
vol/vol)
and precipitated with 2.5 volumes of100% ethanol. The DNAwasresuspendedin TE (1mMEDTA, 10 mMTris-HCl, pH 8.0)and stored at4°C.
Secondary CEF cells were cotransfected with 500 ng of
pA5
pA700
E Hpl Sm P RH K Cl H3 Hpl BH P Sm Hl E
/
\I
Ilm
I
P BwiK
I~
pCK300
E HDI p281MI-1
E H I
pNOTAS
BH Psm HIE
Cl H3 Hpl\ S H[E
FIG. 2. Generation of pA5 subclones. All subclones were
in-serted into pUC18. The location of the
putative
MDV origin ofreplication is indicated as an open circle (o). E, EcoRI;
Hpl,
HpaI;Sm,SmaI;P,PstI;BH,BamHI;K, KpnI;Cl,ClaI;andH3, HindIll.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.65.557.69.203.2] [image:2.612.317.555.454.674.2]0
ee
55.5
5.2-2
4.0_op
2.8.a
I..
1 2 3 4 5
FIG. 3. Replication of the 4-kbpreplicon and pA5. The 4-kbp
replicon andpA5weresubjectedtotheDpnI resistanceassaytotest their abilitytoreplicate in CEF cells. CEF cellswerecotransfected
with eitherthe circularform of the 4-kbpreplicon DNA(lane 3)or
pA5 (lane 4) and 281MI/1 DNA. Seven days aftercotransfection,
total cellular DNA was digested with EcoRI and DpnI, resolved
througha0.8%agarosegel, and transferredtoanylon membrane.
The membrane was probed with 32P-labeled 4-kbp replicon DNA
andvisualized by autoradiography. Lanes: 1, mock-infected CEF
DNA; 2,DNA from CEFtransfected with281MI/1DNA; 3, DNA
from CEFcotransfectedwith281MI/1DNAand thecircular form of
the 4-kbp replicon DNA; 4, DNA from CEF cotransfected with
281MI/1 DNA and pA5; 5, 281MI/1 DNAmixed with methylated
inputpA5.Numbersonleft show size in kilobasepairs.
plasmid DNA and 20 ,ug of 281MI/1 DNA by calcium phosphate coprecipitation (22), exceptthat Dulbecco
mini-mal essentialmediumwasused throughout theexperiments.
When 80% of CEFmonolayersexhibited cytopathic effect(5
to 8 days after cotransfection), total high-molecular-weight
cellularDNA was isolatedas described above.
Approximately 6,ug oftotal cellular DNA was digested with EcoRI andDpnI (International Biotechnologies, Inc.)
under conditions recommended by the manufacturer.
Di-gestedDNA fragments wereresolvedon 0.8%agarose gels
andtransferredtoZetaProbe membranes (Bio-Rad
Labora-tories, Inc., Richmond, Calif.) as recommended by the
supplier. Probes were prepared with [a-32P]dCTP with a
randomprimed labeling kit (Bethesda Research
Laborato-ries,Inc., Gaithersburg, Md.) as specified by the
manufac-turer.Nylonmembranesweresubsequently hybridized with
appropriate32P-labeled probes(6).
As expected, EcoRI and DpnI cleavage of
281MI/1-in-fected CEFDNAproduced5.5-, 5.2-, and 2.8-kbp fragments
which hybridizedtothe4-kbp repliconprobe (Fig. 3,lane2).
The 4-kbp replicon probe, however, did not
hybridize
to uninfected CEF DNA(Fig. 3, lane1).Cotransfection
ofCEF cells with circularEcoRI
4-kbp DNA and281MI/1
DNA resulted in aDpnI-resistant
EcoRI
fragmentof4kbp(Fig.
3, lane 3) in addition to the threeEcoRI
fragments from the281MI/1
helpervirus genome. DNAisolatedfrom CEF cells cotransfected with intactpA5and281MI/1
DNA alsoyielded
a 4-kbp
EcoRI
fragment resistant toDpnI
cleavage(Fig. 3,
lane 4) in addition to the three
EcoRI
fragments.Methylated
inputpA5 DNA mixed with
281MI/1-infected
CEF DNAwas cleaved byDpnI,
indicating thatDpnI
resistance was not a result of inhibition ofDpnI
activity (Fig. 3, lane 5).Results of the
DpnI
resistance assay confirmed that the 4-kbp circular form of the MDV replicon was able toreplicate in CEFcells in the presenceof
281MI/1
helper
virus DNA. pA5, containing the 4-kbp replicon inserted into theEcoRI
site of pUC19, was also replicated as anepisome in CEF cells cotransfected with281MI/1
helper virus DNA. Therefore, we concluded that sequences spanningtheEcoRI
site within MDV replicon DNA are not required for ampli-fication of replicon DNA. However,replication ofthe 4-kbp replicon was at least fivefold more efficient than thatof pA5. Insertion of pUC19 sequences within theEcoRI
site of replicon DNA may have reduced the ability of pA5 to replicate. The decrease in replication activity of pA5 may also be due to some unknown inhibitory sequences within pUC19.Replication activity of pA5 subclones was indicated by the presence of unit-length
DpnI-resistant
EcoRI
subclone DNA following digestion of cotransfected total cell DNA with EcoRI andDpnI
(EcoRI
introduces a single cut within the pA5 subclones). During cotransfection experiments, we ob-served MDV plaque reduction by certain pA5 subclones. Thus, 32P-labeled pA5 was used as a probe to detect both plasmid DNA as well as helper virus DNA. As expected, intact pA5 was able to replicate in the presence of helper virus DNA (Fig. 4, lane 5), generatingDpnI-resistant
EcoRI
fragments of 4 and 2.7 kbp corresponding to virus- and plasmid-specific bands, respectively. However, pA5 alone did not show replication activity, suggesting that functions provided in trans by a helper virus are essential for replica-tion (Fig. 4, lane 3). Methylated input pA5, used as a control, was susceptible to
DpnI
cleavage (Fig. 4, lane 1). Among tested subclones, only pA700, which contains the putative MDV origin of replication identified by DNA sequence analysis, replicated as indicated by aDpnI-resistant
EcoRI
fragment of 3.4 kbp (2,686-bp pUC18 plus 700-bp
PstI-to-BamHIMDV DNA) (Fig. 4, lane 9). CEF cells cotransfected with 281MI/1 DNA and pNOTA5, p281MI-1, or pCK300 (Fig. 2) representing the remainder of MDV replicon DNA were not
DpnI
resistant, indicating that they do not contain an origin of replication (Fig. 4, lanes 6 to 8). In all cases, a fraction of input plasmid DNA was susceptible toDpnI
cleavage and migrated as low-molecular-weightmaterial,
evident after prolonged exposures of the autoradiogram (data not shown).Results of
DpnI
resistance assays indicated that a func-tional origin of MDV replication is located in a 700-bp PstI-to-KpnI subfragment of the MDV replicon. The com-plete nucleotide sequence of the 700-bp fragment is shown in Fig. 5. Comparison of pA700 nucleotide sequences with the GenBank data base revealed no significant sequence homol-ogy to any known viral sequences. The exact location of the MDV replication origin in the 281MI/1 viral genome is unknown because of the lack of complete restriction endo-nuclease maps or genomic clones of serotype 2 MDV.on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.118.247.72.369.2]4.0-..
2.- s
I
_,~~~~~~~~~~~.
1 2 3 4 5 6 7 8 9
FIG. 4. Replicationanalysis of pA5 subclones. Each of the pA5 subclones was tested for replication activity based on the DpnI resistance assay as described in Fig. 3. Each lane contains total transfectedcellular DNA digested withEcoRIandDpnI.32P-labeled pA5 was used to detect plasmid DNA as well as 281MI/1 DNA. Lanes: 1, pA5 Dam'; 2, mock-infected CEF DNA; 3, DNA from CEF cells transfectedwith pA5; 4, DNA from CEF cells transfected with 281MI/1 DNA; 5, DNA from CEF cells cotransfected with 281MI/1DNAandpA5; 6, DNAfromCEF cells cotransfected with 281MI/1DNAand pNOTA5; 7, DNA from CEF cells cotransfected with 281MI/1DNA andp281MI-1; 8, DNA from CEF cells cotrans-fected with 281MI/1 DNA and pCK300; 9, DNA from CEF cells cotransfected with281MI/1 DNAandpA700.
Studies from our laboratory indicate that there are two copies of the replicon sequences within the 281MI/1 viral genome (27). This suggests that there are at least two replication origins in MDV.
HSVcontains two copiesof
oris
withinTRs
andIRS
and was localized by utilizing HSV amplicons (11, 12). By analogy with HSV, the MDV replicon sequence is most likelytobe present in the repeatsflankingtheUs
segmentof the MDV genome,assumingthat the structure of serotype 2 MDV DNAis similartothatof serotype 1 and 3MDVDNA. However, on the basis ofDNA sequence identity to lytic origins of alphaherpesvirus replication, Bradleyetal.(3)and Morgan et al. (21) have located aputative serotype 1MDV originofreplicationin repeatsflankingtheUL region.MDVserotype2functionalreplication originsequences, identified in this study, share 82% identity to theputative serotype 1
MDVreplication
origin
sequences.In conclusion, we identified a functional origin of MDV replication within a 700-bp subfragment of the MDV repli-con.Systematic deletion
analysis
within the700-bpfragment
willbe useful todeterminetheminimumsequences
required
for MDV DNAreplication.
Nucleotide sequenceaccession number. The GenBank ac-cession number for the nucleotide sequences reported here
is M74523.
PSTI 20 40 60
CIGCAGVICG GGAAGAGCGG GGGAGGCGCCGICCGAGCG CCGIGAG GTIIGGGG
80 100 120
Cg@3lArG ¶GGIGGGGAAGGGAAGGGG AGACGGCCGA AACCTACGCG T7 CGCGC
140 160 180
TCCGATCCGG GA.TCr-CC GACGGGCt'r C!I' TCCGIATCCG 7CIr'TCCGCCA
200 220 240
CCI'I~CGTC G'IrGCGGGAACCGGACCG GCGCICrAAG CGGAGATCCGGCGC7CCGCr
260 280 300
I I'rA5GACC GGGCCGGI= GAGGGCGTAACGATCAGIG A7GCAATGCAAACGAGCGGG
320 340 360
GCAGACGCGr CAGC_GIIGC
ACCGC:A:ATAAGAT
TATATATATAATATAITAIT380 400 420
GGCGCAAGGT GCGAACGCCG C CGGGAAGCGG GATCCTAIGC CACLGII1
440 460 480
UGICCGGCCG CGCCCGCGC CGGGCTAGA AAC':3 CCCWC G GGGGCGATC
500 520 540
GGGGACA GGCCrACAAA TACGCGAGCG GPG7CCGGC GGGGACCGT CGIGCIG
560 580 600
GCCGGCCCGC C)G2CCGAAAG CGCGGGACCG CGGTAATAA GCGCCCGCCG TCGCGGATCG
620 640 660
GAi?wlwlIwl3(; lCGTrrACCGCGGGC GAACCGCG GCGAACGAAC CCGITCCCGr
680 700 720
GGGATGCAG GCGGCCGGGA PGCATCGCG CGCCGxTCCC AACGAtG CrACGGCrCG
740 760
C21ItGCGGG GGTCGCCGAC GGGI3GAAOGGGGAP.G1XAC
KNI
FIG. 5. Nucleotide sequences of pA700. pA700, containing a
700-bpsubfragment ofthe MDV replicon DNA,was sequencedin bothorientationsby using dideoxy chain terminations (26). Under-lines represent an MDV replication originsequence that shares a
significantsequenceidentitytoalphaherpesvirus replicationorigins.
Blackbarsindicatethe9-bp motif(CGTTCGCAC) highly conserved among alphaherpesvirus replication origins. Rectangles represent possible Spl-binding sites (GGGCGG), and circles (<>) depict
possible CAAT bindingregions.
We thank A. Finkelstein for providing the pNOTA5 construct.
WealsothankL. F.Velicer andJ. D.Reilly for criticalreviewofthe manuscript.
This research was supported in part by competitive research grants85-CRCR-1-1709 and88-37266-3983 awarded to R. F. Silva andP. M. Coussens,respectively, underthe competitive research grantsadministeredby the U.S. Department ofAgricultureand the MichiganAgriculturalExperimentStation.H.S.Campissupported in part by a Michigan State University biotechnology research fellowshipaward.
REFERENCES
1. Anders, D. G., and S. M. Punturieri. 1991. Multicomponent
originofcytomegaloviruslytic-phaseDNAreplication.J.Virol. 65:931-937.
2. Baumann, R. P., R. R. Yalamanchili,and D. J. O'Callaghan.
1989. Functional mapping and DNA sequence of an
equine
herpesvirus 1originofreplication.J. Virol.63:1275-1283. 3. Bradley, G., M. Hayashi, G. Lancz, A. Tanaka, and M.
Nonoyama. 1989. Structure of the Marek's disease virus BamHI-H genefamily:genesofputativeimportancefortumor
induction.J. Virol.63:2534-2542.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.89.270.72.325.2] [image:4.612.316.558.72.425.2]4. Bruckner, R.C., J. J.Crute, M.S.Dodson,andI. R.Lehman.
1991. The herpessimplexvirus 1originbinding protein: aDNA
helicase. J. Biol. Chem. 266:2669-2674.
5. Buckmaster, A. E., S. D. Scott, M. J. Sanderson, M. E. G. Boursnell, N. L. J.Ross, and M. M. Binns. 1988. Genesequence
andmapping data fromMarek's disease virus andherpesvirusof
turkeys: implications for herpesvirus classification. J. Gen.
Virol. 69:2033-2042.
6. Budowle, B., and F. S. Baechetel. 1990. Modifications to im-provethe effectiveness of restrictionfragment length polymor-phism typing. Appl. Theor. Electro. 1:181-187.
7. Calnek, B. W., and R. L. Witter. 1991. Marek's disease, p. 342-385. In B. W. Calnek, H. J. Barnes, C. W. Beard, W. M.
Reid, and H. W. Yoder, Jr. (ed.), Diseases of poultry. Iowa State University Press, Ames, Iowa.
8. Carter, J. K., and R.F.Silva. 1990. Cell cultureamplificationof adefective Marek's disease virus.VirusGenes4:225-237. 9. Deb, S., and M. Doelberg. 1988. A 67-base-pairsegment from
theori-s region of herpes simplex virus type 1 encodesorigin function. J. Virol. 62:2516-2519.
10. Elias, P., and I. R. Lehman. 1988. Interaction oforigin binding protein with anorigin of replication of herpes simplexvirus 1.
Proc. Natl. Acad. Sci. USA 85:2959-2963.
11. Frenkel, N.1980.Defective interferingherpesviruses,p.91-120.
In A.J.Nahmias, W. R. Dowdle, and R. F. Schinazi (ed.), The human herpesviruses. Elsevier Science Publishing, Inc., New York.
12. Frenkel, N., H. Locker, W. Batterson, G. S. Hayward, and B.
Roizman. 1976. Anatomy of herpes simplex virus DNA. VI. Defective DNA originates from the S component. J. Virol. 20:527-531.
13. Gahn, T. A., and C. L. Schildkraut. 1989. The Epstein-Barr virus origin of plasmid replication, orip, contains both the initiation andtermination sites of DNA replication. Cell 58:527-535.
14. Hammerschmidt, W., and B. Sugden. 1988. Identification and characterization oforiLyt, alytic origin of DNA replication of
Epstein-Barr virus. Cell 55:427-433.
15. Hamzeh,F. M.,P.S.Lietman, W. Gibson, and G. S. Hayward.
1990. Identification of the lytic origin of DNA replication in humancytomegalovirus byanovel approach utilizing
ganciclo-vir-induced chain termination. J.Virol.64:6184-6195.
16. Henikoff,S. 1987. Unidirectional digestion withexonuclease III inDNAsequenceanalysis. Methods Enzymol. 155:156-159.
17. Hernandez, T. R., R. E. Dutch, I. R. Lehman, C. Gustafsson,
andP. Elias. 1991. Mutations ina herpes simplex virustype1
origin that inhibit interaction with origin-binding protein also
inhibit DNAreplication. J. Virol.65:1649-1652.
18. Koff, A., and P. Tegtmeyer. 1988. Characterization of major
recognition sequencesforaherpes simplex virustype 1
origin-binding protein. J. Virol. 62:4096-4103.
19. Kwong, A. D., and N. Frenkel. 1984. Herpes simplex virus
amplicons: effect of sizeonreplication ofconstructed defective
genomescontaining eucaryotic DNA sequences. J. Virol. 51:
595-603.
20. Lockshon, D., and D. A. Galloway. 1988. Sequence and
struc-turalrequirements ofa herpes simplex viralDNA replication
origin. Mol. Cell. Biol. 8:4018-4027.
21. Morgan, R. W., J. L. Cantello, J. A. J. Claessens, and P. Sondermeyer. 1991. Inhibition of Marek's disease virus DNA
transfection bya sequencecontaininganalphaherpesvirus
ori-gin ofreplicationandflankingtranscriptionregulatoryelements. Avian Dis. 35:70-81.
22. Morgan, R. W., J. L. Cantello, and C. H. McDermott. 1990.
Transfection of chicken embryo fibroblasts with Marek's
dis-easevirus DNA. Avian Dis. 34:345-351.
23. Payne, L. N. 1989. Pathogenesis of Marek's disease virus, p. 307-316. In S.Kato,T.Horiuchi,T.Mikami,andK. Hirai (ed.), Advances in Marek's disease virus research. Japanese Associ-ation on Marek'sDisease, Osaka, Japan.
24. Polvino-Bodnar, M., P. K. Orberg, and P. A. Shaffer. 1987.
Herpes simplex virus type 1 oriL is not required for virus replication or for the establishment and reactivation of latent infection in mice. J. Virol. 61:3528-3535.
25. Roizman, B. 1990. Herpesviridae: a briefintroduction, p. 1787-1793. In B. N. Fields and D. M. Knipe (ed.), Fieldsvirology. RavenPress, New York.
26. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
27. Silva, R. F., J. Carter, and R. L. Witter. 1988. Construction of a shuttle vectorand its utilization as a vehicle to insertforeign DNA into the Marek's disease virusgenome, p. 50-55. In S. Kato,T. Horiuchi, T. Mikami, and K. Hirai (ed.), Advances in Marek's disease research. Japanese Association on Marek's Disease, Osaka,Japan.
28. Silva, R. F., and L. F. Lee. 1984. Monoclonalantibody-mediated immunoprecipitation of proteins from cells infected with Marek's disease virus orturkey herpesvirus.Virology 136:307-320.
29. Smith, E. J., A. Fadly, and W. Okazaki. 1979. Anenzyme-linked immunosorbentassay fordetecting avian leukosis-sarcoma vi-ruses.Avian Dis. 33:698-707.
30. Spaete, R. R., and N. Frenkel. 1985. The herpes simplex virus amplicon: analysis of cis-acting replication functions. Proc. Natl.Acad. Sci. USA 82:694-698.
31. Stow, N. D. 1982. Localization of an origin of DNA replication within theTRs/IRs repeated region ofthe herpessimplexvirus type 1 genome. EMBO J. 1:863-867.
32. Stow, N. D., and J. Davison. 1986.Identification of a varicella-zoster virus origin of DNA replication and its activation by herpes simplex type 1 gene products. J. Gen. Virol. 67:1613-1623.
33. Stow, N. D., and E. C. McMonagle. 1983. Characterization of theTRs/IRs originof DNA replication of herpessimplex type 1. Virology 130:427-438.
34. Vlazny, D. A., and N. Frenkel. 1981. Replication of herpes simplex virus DNA: localizationof replication recognition sig-nals within defective virus genomes. Proc. Natl. Acad. Sci. USA78:742-746.
35. Weller, S. K., A.Spadaro, J. E. Schaffer, A. W. Murray, A. M. Maxam, and P. A. Shaffer. 1985. Cloning, sequencing, and functional analysis ofori-L, a herpes simplex virus type 1 origin of DNAsynthesis. Mol. Cell. Biol. 5:930-942.
36. Witter, R. L. 1983. Characteristicsof Marek'sdisease viruses isolatedfrom vaccinatedcommercial chickenflocks: association ofviral pathotype with lymphomafrequency. Avian Dis. 27: 113-132.
37. Yates, J., N. Warren, D. Reisman, and B. Sugden. 1984. A cis-acting element from the Epstein-Barr viral genome that permits stable replication of recombinant plasmids in latently infectedcells. Proc. Natl. Acad. Sci. USA 81:3806-3810.